
Structural and Functional Changes of Reconstituted High-Density Lipoprotein (HDL) by Incorporation of α-synuclein: A Potent Antioxidant and Anti-Glycation Activity of α-synuclein and apoA-I in HDL at High Molar Ratio of α-synuclein


Abstract
:1. Introduction
2. Results
2.1. Purification and Characterization of α-syn
2.2. Phospholipid Binding Ability
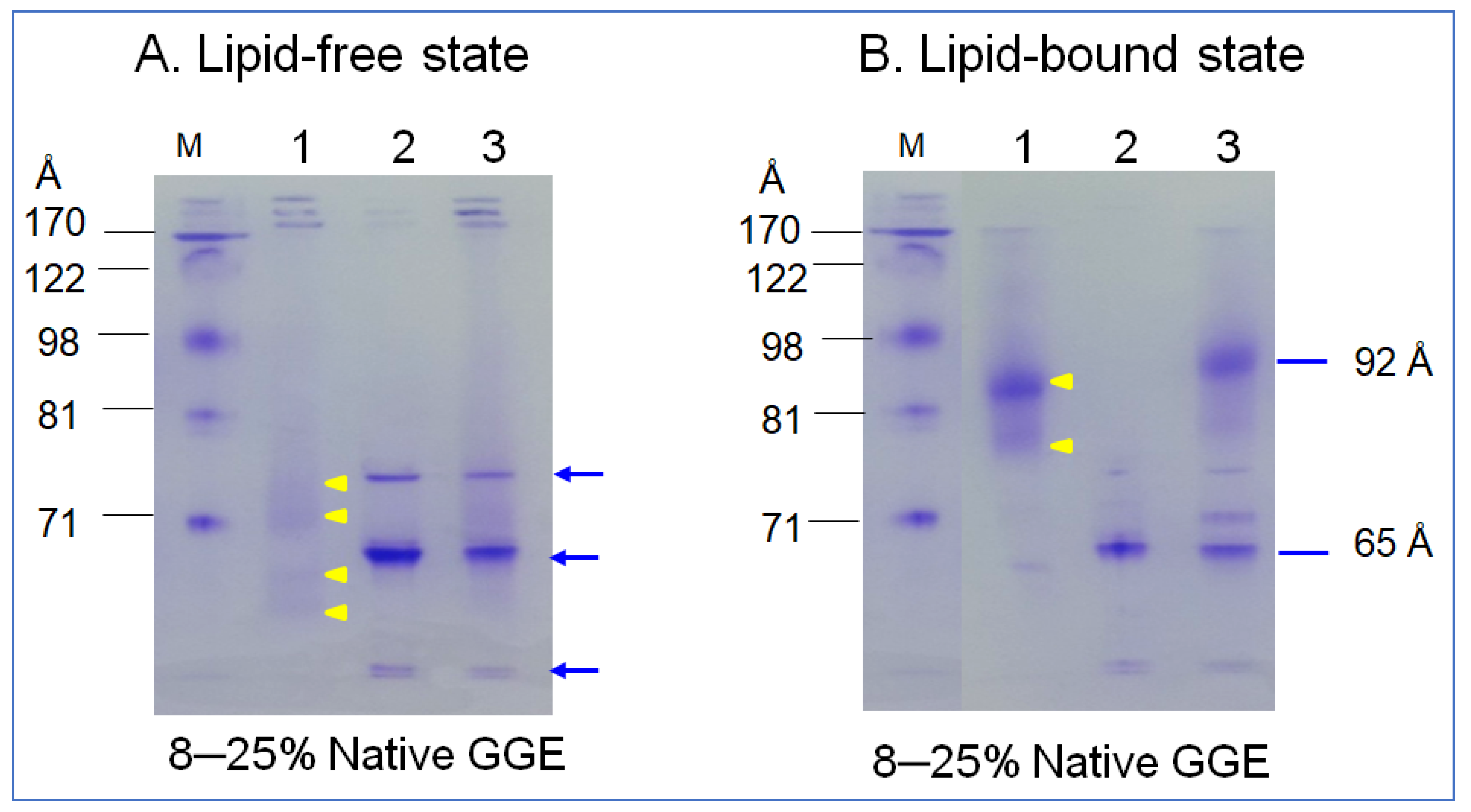
2.3. Synthesis of rHDL
2.4. Secondary Structure Analysis
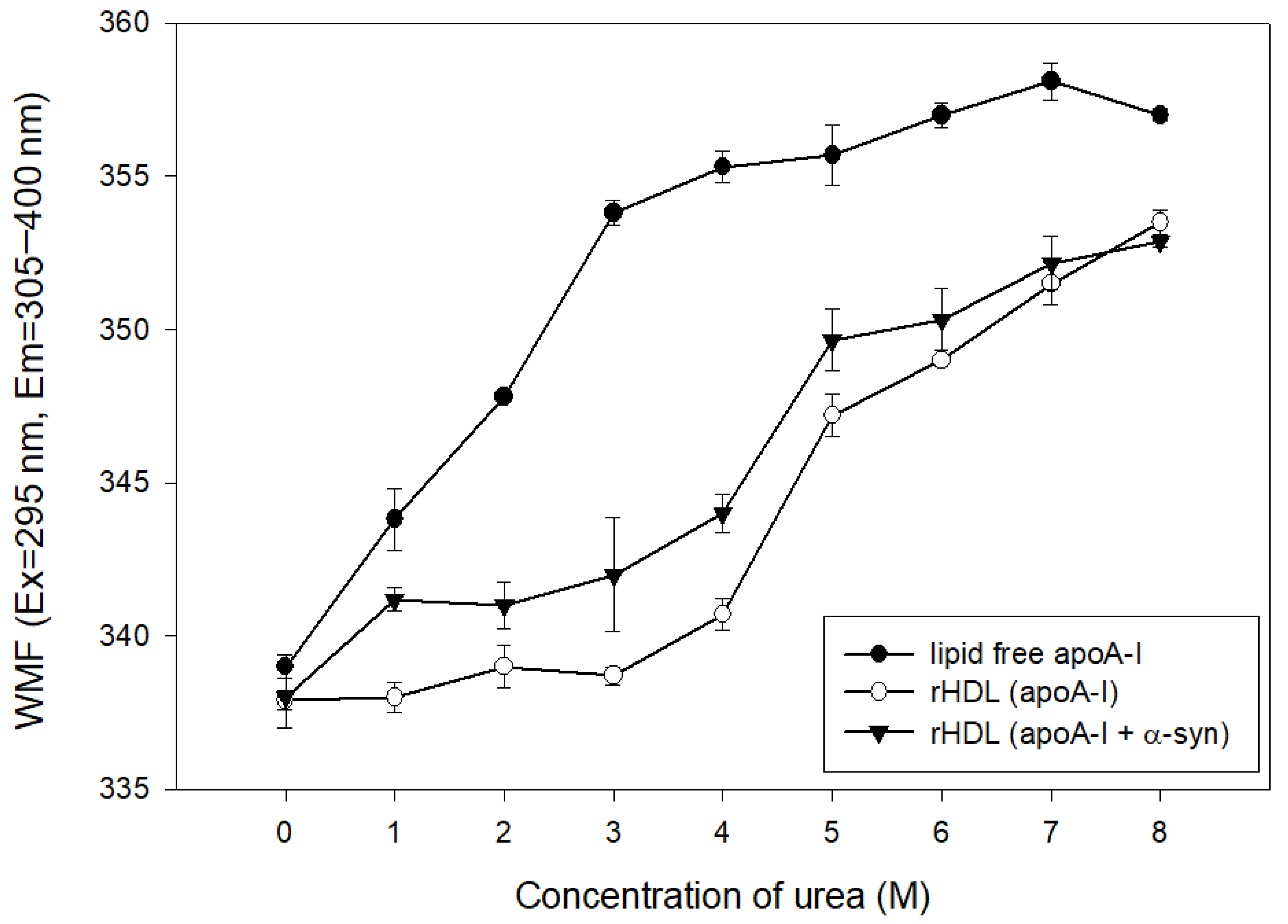
2.5. α-syn Caused More Rapid Isothermal Denaturation
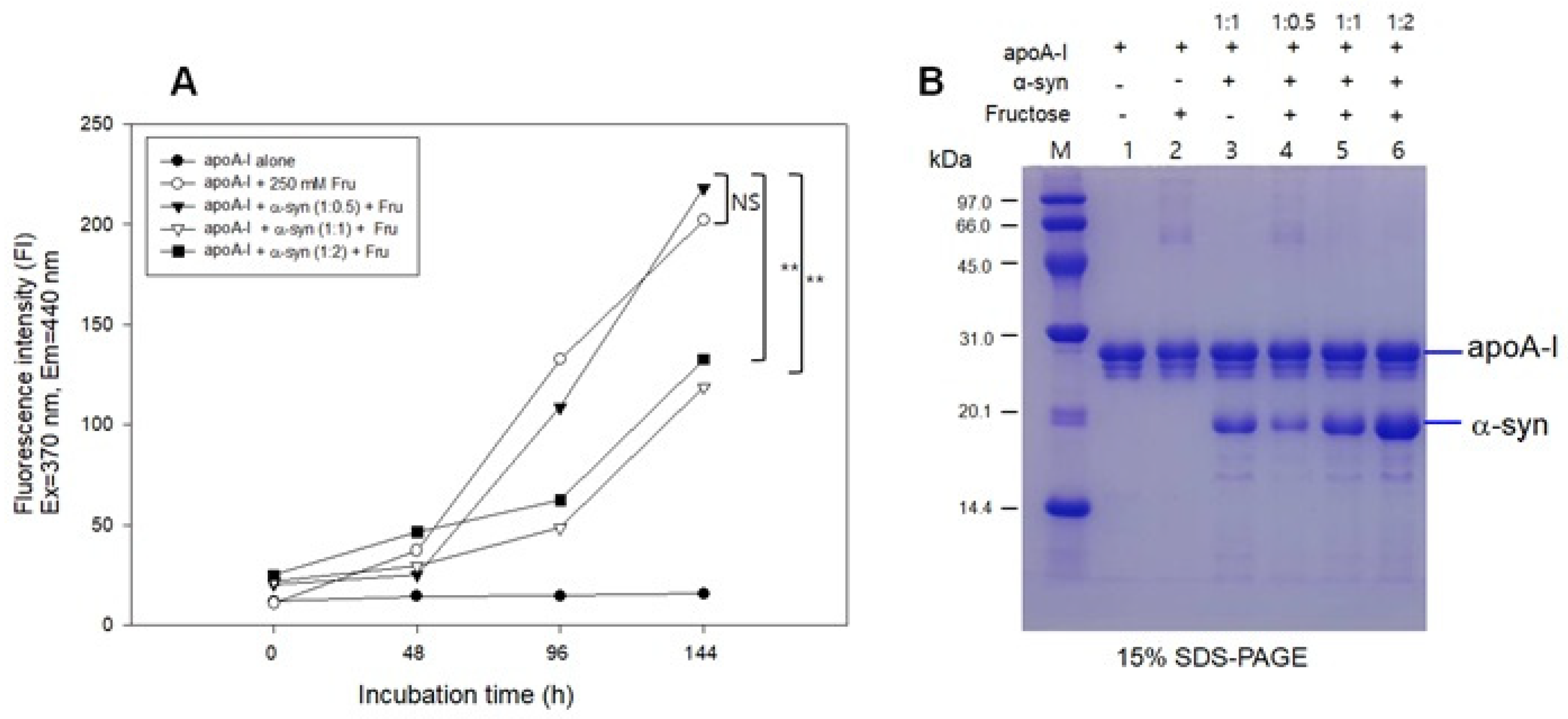
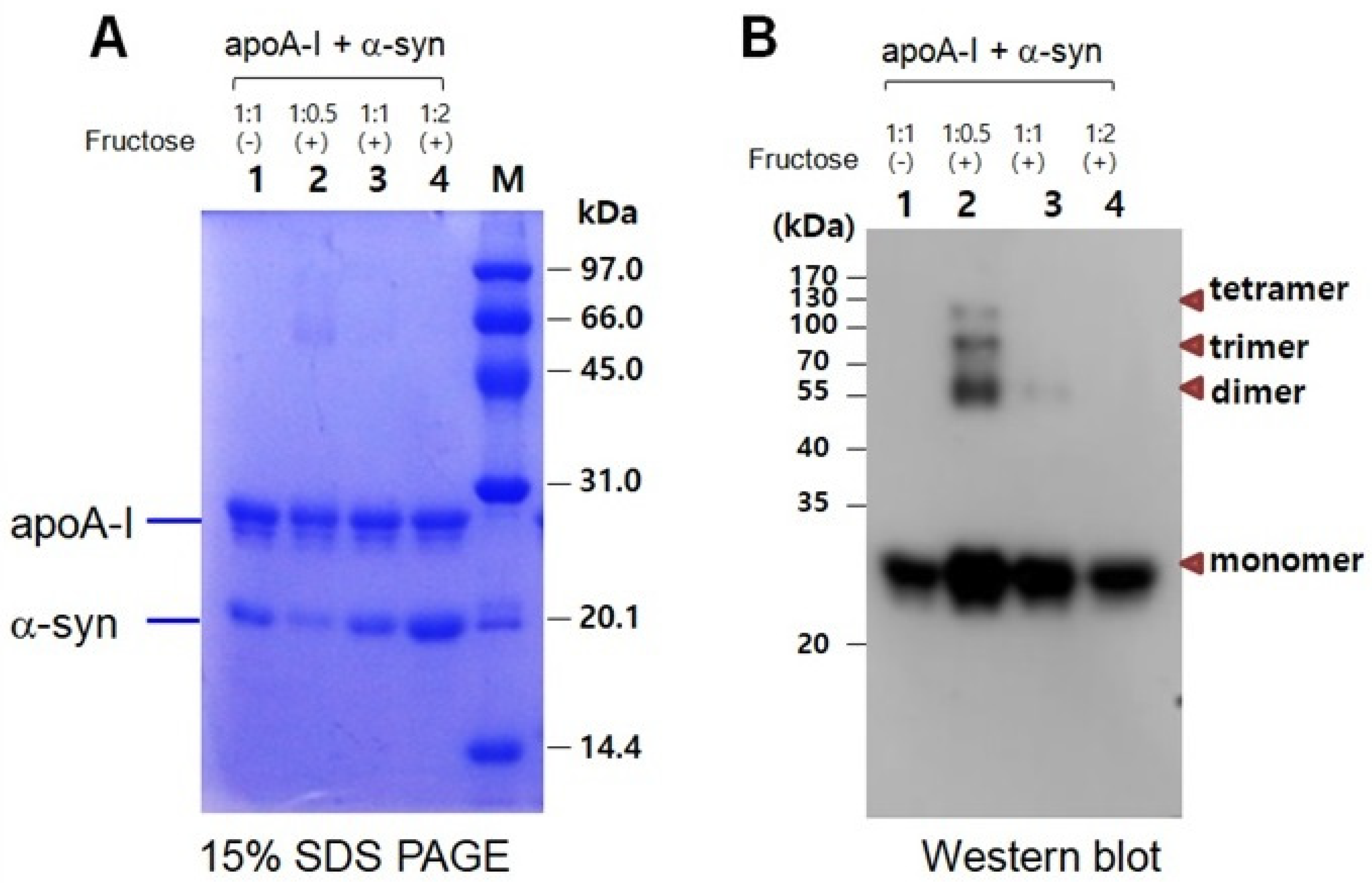
2.6. Inhibition of Glycation by α-syn
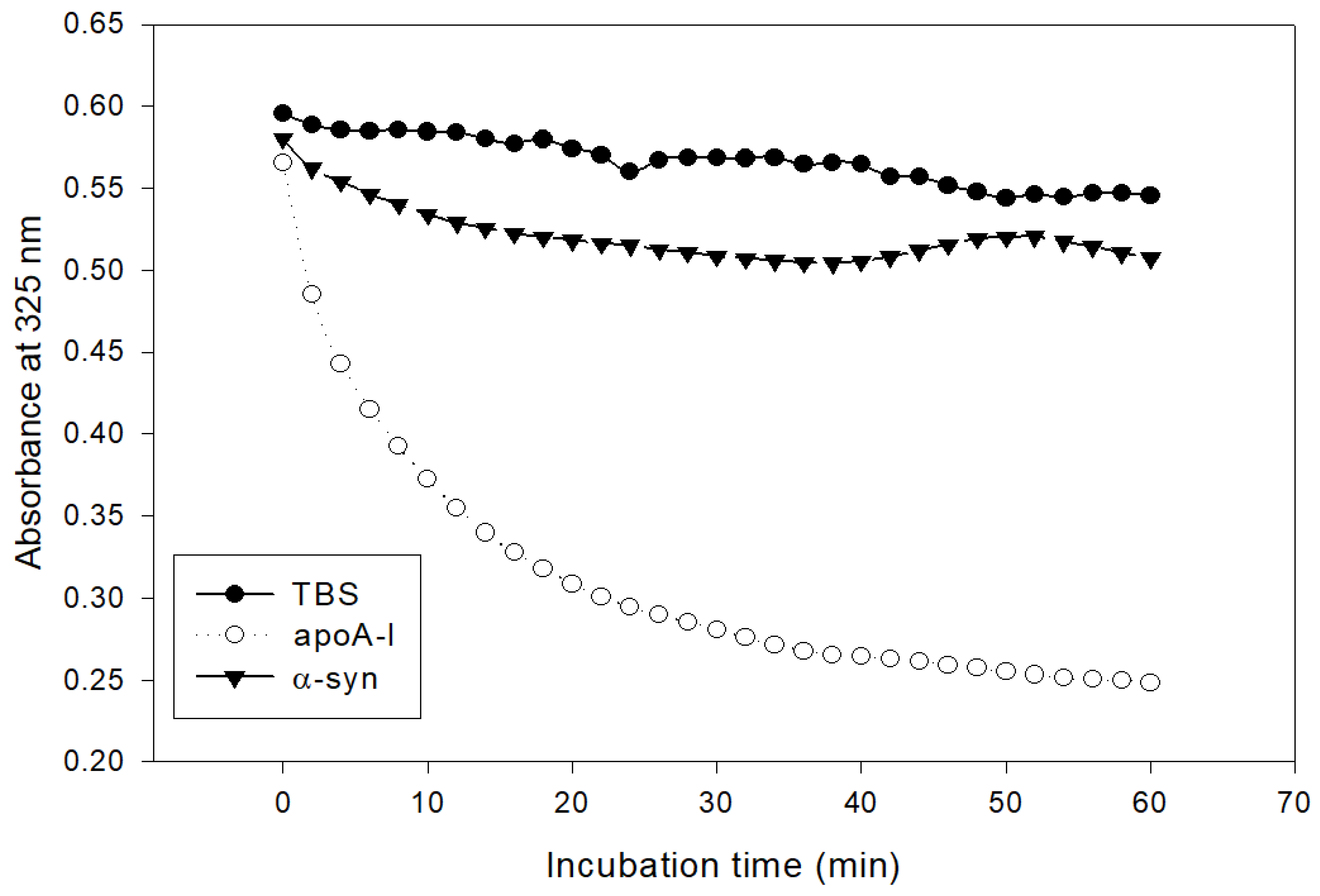
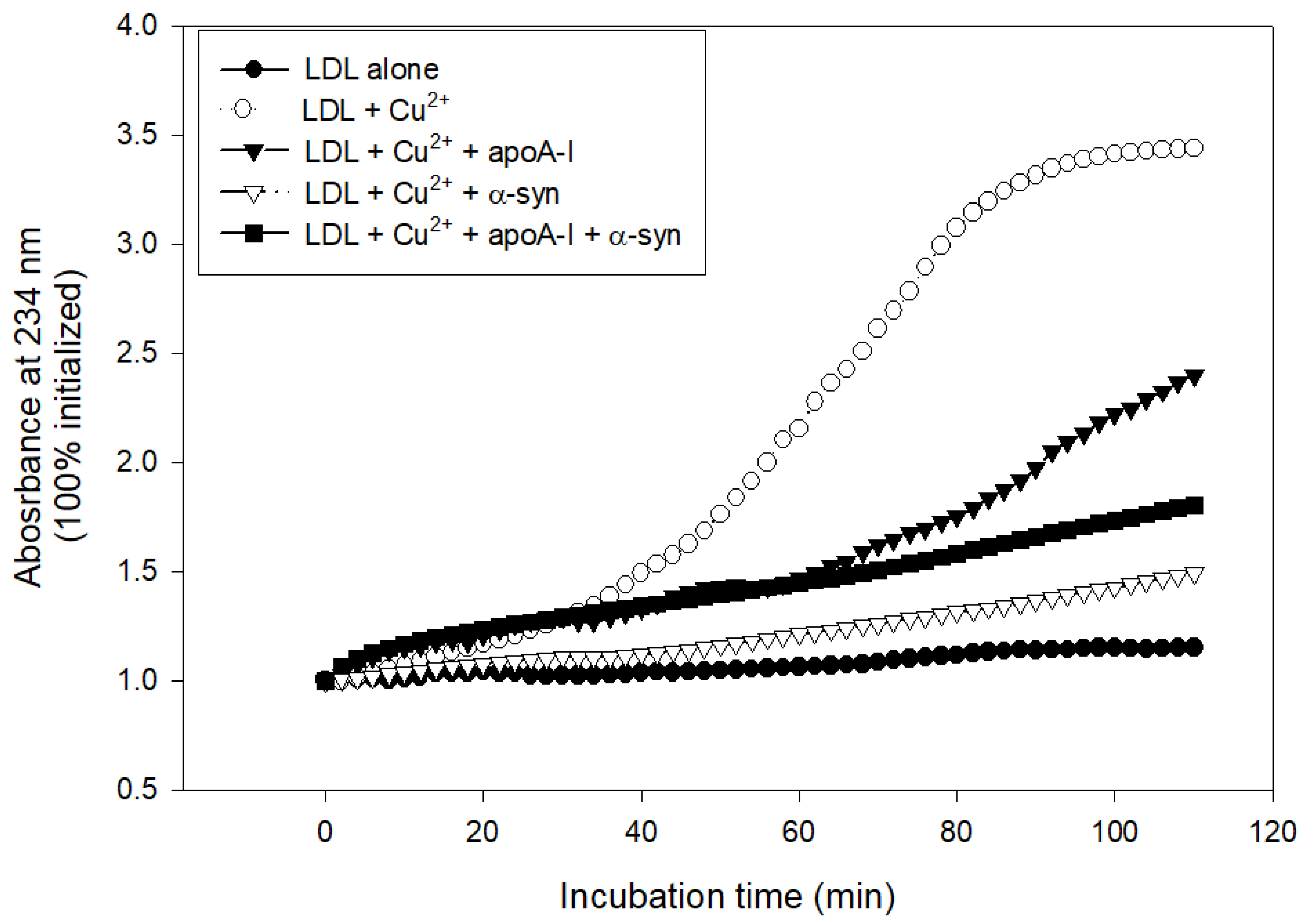
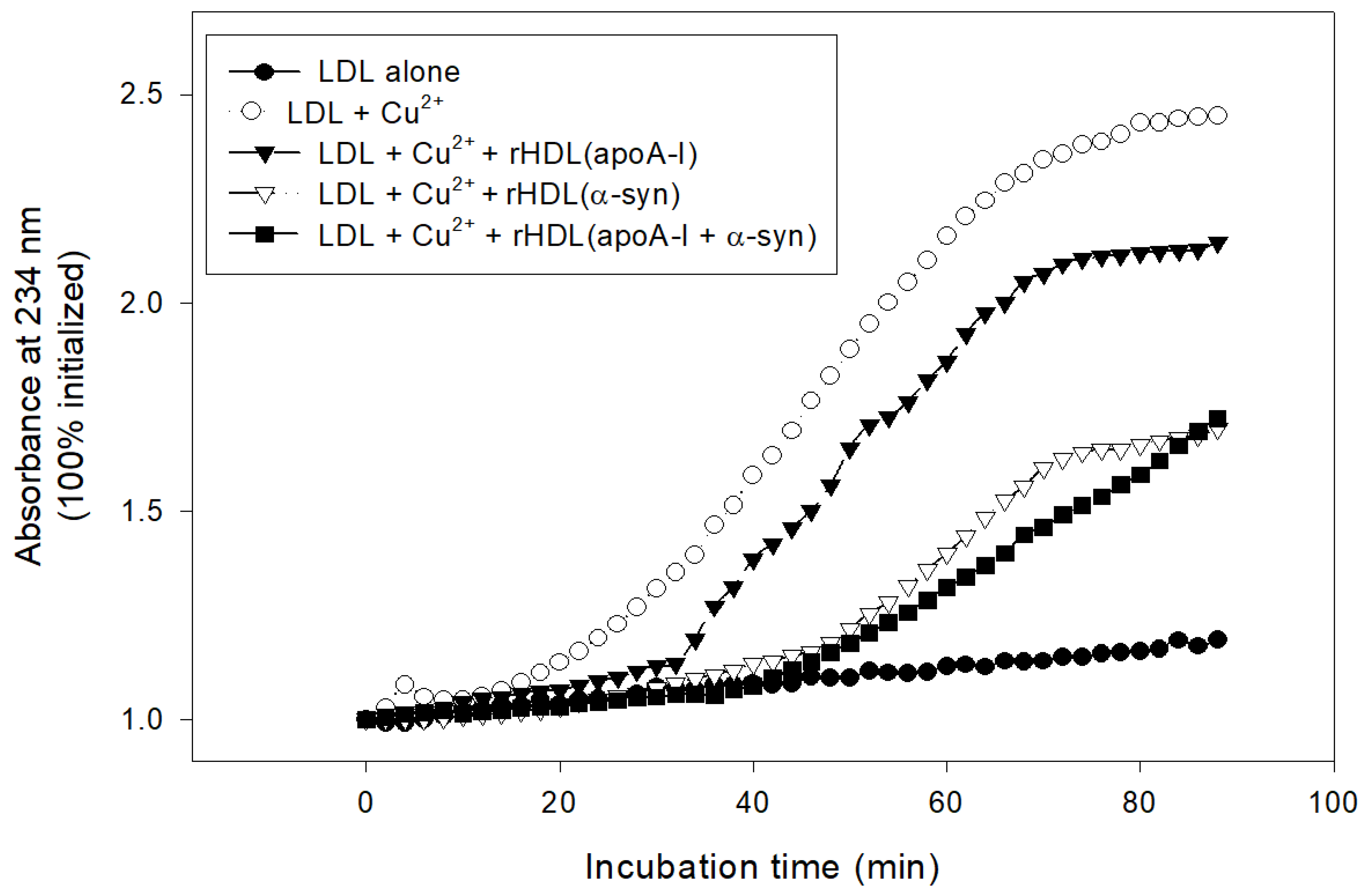
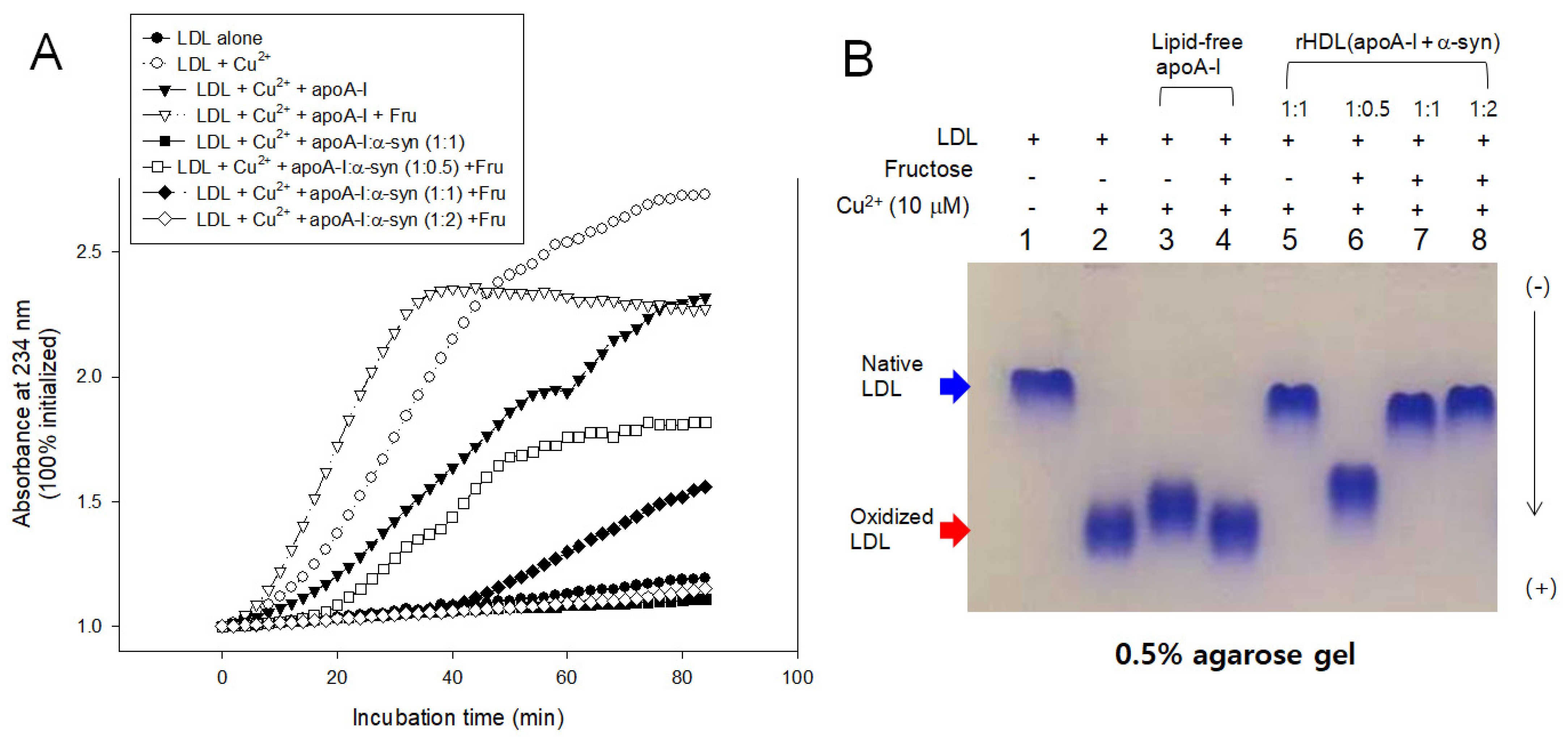
2.7. Inhibition of LDL Oxidation
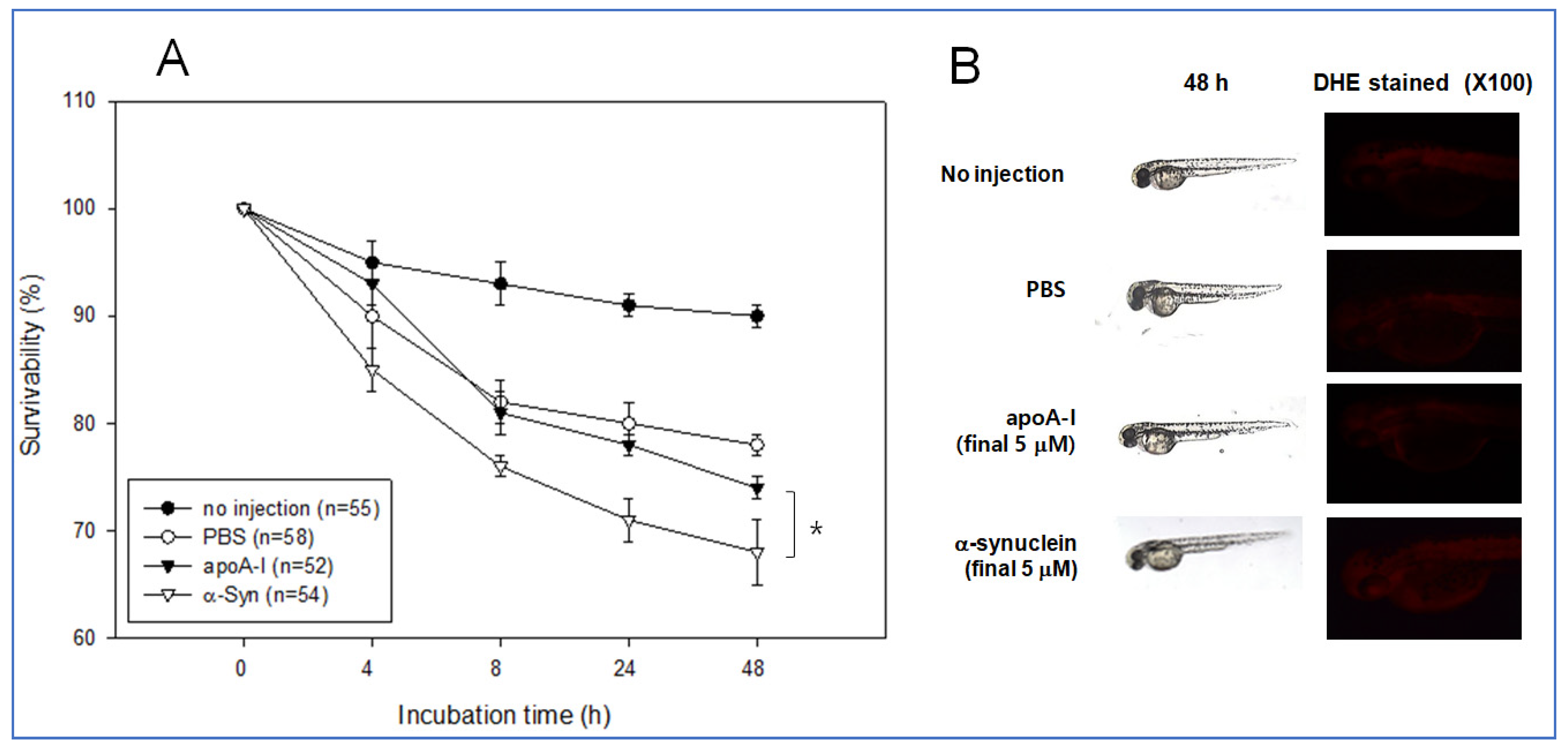
2.8. Embryo Survivability by Microinjection
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Expression and Purification of α-synuclein
4.3. Protein Sequencing and Isoelectric Focusing
4.4. Characterization of Secondary Structure by Circular Dichroism
4.5. Trp Fluorescence during Isothermal Denaturation
4.6. Purification of Human Lipoproteins
4.7. Purification of Human apoA-I
4.8. Oxidation of LDL
4.9. Synthesis of Reconstituted HDL
4.10. DMPC Clearance Assay
4.11. Glycation of apoA-I with α-syn
4.12. Western Blotting
4.13. Inhibition of LDL Oxidation
4.14. Zebrafish
4.15. Microinjection into Zebrafish Embryos
4.16. Statistical Analysis
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kalia, L.V.; Lang, A.E. Parkinson’s disease. Lancet 2015, 386, 896–912. [Google Scholar] [CrossRef]
- Karimi-Moghadam, A.; Charsouei, S.; Bell, B.; Jabalameli, M.R. Parkinson disease from mendelian forms to genetic susceptibility: New molecular insights into the neurodegeneration process. Cell. Mol. Neurobiol. 2018, 38, 1153–1178. [Google Scholar] [CrossRef] [Green Version]
- Gibb, W.R.; Lees, A.J. The relevance of the Lewy body to the pathogenesis of idiopathic Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 1988, 51, 745–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, U.; Park, S.J.; Park, S.M. Cholesterol metabolism in the brain and its association with Parkinson’s disease. Exp. Neurobiol. 2019, 28, 554–567. [Google Scholar] [CrossRef] [Green Version]
- Polymeropoulos, M.H.; Lavedan, C.; Leroy, E.; Ide, S.E.; Dehejia, A.; Dutra, A.; Pike, B.; Root, H.; Rubenstein, J.; Boyer, R.; et al. Mutation in the α-synuclein gene identified in families with Parkinson’s disease. Science 1997, 276, 2045–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasanen, P.; Myllykangas, L.; Siitonen, M.; Raunio, A.; Kaakkola, S.; Lyytinen, J.; Tienari, P.J.; Pöyhönen, M.; Paetau, A. A novel α-synuclein mutation A53E associated with atypical multiple system atrophy and Parkinson’s disease-type pathology. Neurobiol. Aging 2014, 35, 2180.e1–2180.e5. [Google Scholar] [CrossRef] [PubMed]
- Swanson, C.R.; Berlyand, Y.; Xie, S.X.; Alcalay, R.N.; Chahine, L.M.; Chen-Plotkin, A.S. Plasma apolipoprotein A1 associates with age at onset and motor severity in early Parkinson’s disease patients. Mov. Disord. 2015, 30, 1648–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson, C.R.; Li, K.; Unger, T.L.; Gallagher, M.D.; Van Deerlin, V.M.; Agarwal, P.; Leverenz, J.; Roberts, J.; Samii, A.; Gross, R.G.; et al. Lower plasma apolipoprotein A1 levels are found in Parkinson’s disease and associate with apolipoprotein A1 genotype. Mov. Disord. 2015, 30, 805–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, W.; Wan, X.; Liu, B.; Rong, X.; Zhu, L.; Li, P.; Li, J.; Wang, L.; Cui, L.; Wang, X. Specific changes of serum proteins in Parkinson’s disease patients. PLoS ONE 2014, 9, e95684. [Google Scholar] [CrossRef] [Green Version]
- Qiang, J.K.; Wong, Y.C.; Siderowf, A.; Hurtig, H.I.; Xie, S.X.; Lee, V.M.; Trojanowski, J.Q.; Yearout, D.; Leverenz, J.; Montine, T.J.; et al. Plasma apolipoprotein A1 as a biomarker for Parkinson disease. Ann. Neurol. 2013, 74, 119–127. [Google Scholar] [CrossRef]
- Vitali, C.; Wellington, C.L.; Calabresi, L. HDL and cholesterol handling in the brain. Cardiovasc. Res. 2014, 103, 405–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Eckel, R.H. What are lipoproteins doing in the brain? Trends Endocrinol. Metab. 2014, 25, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Fantini, J.; Carlus, D.; Yahi, N. The fusogenic tilted peptide (67–78) of alpha-synuclein is a cholesterol binding domain. Biochim. Biophys. Acta 2011, 1808, 2343–2351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, J.T.; Halliday, G.M.; Kim, W.S. α-synuclein regulates neuronal cholesterol efflux. Molecules 2017, 22, 1769. [Google Scholar] [CrossRef] [Green Version]
- Emamzadeh, F.N.; Allsop, D. α-synuclein Interacts with Lipoproteins in Plasma. J. Mol. Neurosci. 2017, 63, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.H. High-Density Lipoproteins as Biomarkers and Therapeutic Tools: Volume 2. Improvement and Enhancement of HDL and Clinical Applications, 1st ed.; Springer: New York, NY, USA, 2019. [Google Scholar]
- He, Y.; Kothari, V.; Bornfeldt, K.E. High-density lipoprotein function in cardiovascular disease and diabetes. Arterioscler. Thromb. Vasc. Biol. 2018, 38, e10–e16. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.H. Biomedicinal implications of high-density lipoprotein: Its composition, structure, functions, and clinical applications. BMB Rep. 2009, 42, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.H. High-Density Lipoproteins as Biomarkers and Therapeutic Tools: Volume 1. Impacts of Lifestyle, Diseases, and Environmental Stressors on HDL, 1st ed.; Springer: New York, NY, USA, 2019. [Google Scholar]
- Nam, G.E.; Kim, S.M.; Han, K.; Kim, N.H.; Chung, H.S.; Kim, J.W.; Han, B.; Cho, S.J.; Yu, J.H.; Park, Y.G.; et al. Metabolic syndrome and risk of Parkinson disease: A nationwide cohort study. PLoS Med. 2018, 15, e1002640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Ji, L.; Zhang, D.; Tong, X.; Pan, B.; Liu, P.; Zhang, Y.; Huang, Y.; Su, J.; Willard, B.; et al. Nonenzymatic glycation of high-density lipoprotein impairs its anti-inflammatory effects in innate immunity. Diabetes Metab. Res. Rev. 2012, 28, 186–195. [Google Scholar] [CrossRef]
- Guerrero, E.; Vasudevaraju, P.; Hegde, M.L.; Britton, G.B.; Rao, K.S. Recent advances in α-synuclein functions, advanced glycation, and toxicity: Implications for Parkinson’s disease. Mol. Neurobiol. 2013, 47, 525–536. [Google Scholar] [CrossRef]
- Vicente Miranda, H.; Outeiro, T.F. The sour side of neurodegenerative disorders: The effects of protein glycation. J. Pathol. 2010, 221, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Olsen, L.K.; Dowd, E.; McKernan, D.P. A role for viral infections in Parkinson’s etiology? Neuronal Signal. 2018, 2, NS20170166. [Google Scholar] [CrossRef]
- Belin, A.C.; Ran, C.; Anvret, A.; Paddock, S.; Westerlund, M.; Håkansson, A.; Nissbrandt, H.; Söderkvist, P.; Dizdar, N.; Ahmadi, A.; et al. Association of a protective paraoxonase 1 (PON1) polymorphism in Parkinson’s disease. Neurosci. Lett. 2012, 522, 30–35. [Google Scholar] [CrossRef]
- Kondo, I.; Yamamoto, M. Genetic polymorphism of paraoxonase 1 (PON1) and susceptibility to Parkinson’s disease. Brain Res. 1998, 806, 271–273. [Google Scholar] [CrossRef]
- Cho, K.H.; Kim, J.R.; Lee, I.C.; Kwon, H.J. Native high-density lipoproteins (HDL) with higher Paraoxonase exerts a potent antiviral effect against SARS-CoV-2 (COVID-19), while glycated HDL lost the antiviral activity. Antioxidants 2021, 10, 209. [Google Scholar] [CrossRef] [PubMed]
- Sulzer, D.; Antonini, A.; Leta, V.; Nordvig, A.; Smeyne, R.J.; Goldman, J.E.; Al-Dalahmah, O.; Zecca, L.; Sette, A.; Bubacco, L.; et al. COVID-19 and possible links with Parkinson’s disease and parkinsonism: From bench to bedside. NPJ Parkinsons Dis. 2020, 6, 18. [Google Scholar] [CrossRef]
- Searles Nielsen, S.; Checkoway, H.; Zhang, J.; Hofmann, J.N.; Keifer, M.C.; Paulsen, M.; Farin, F.M.; Cook, T.J.; Simpson, C.D. Blood alpha-synuclein in agricultural pesticide handlers in central Washington State. Environ. Res. 2015, 136, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Ganguly, U.; Singh, S.; Pal, S.; Prasad, S.; Agrawal, B.K.; Saini, R.V.; Chakrabarti, S. Alpha-synuclein as a biomarker of Parkinson’s disease: Good, but not good enough. Front. Aging Neurosci. 2021, 13, 702639. [Google Scholar] [CrossRef]
- Zhou, A.L.; Swaminathan, S.K.; Curran, G.L.; Poduslo, J.F.; Lowe, V.J.; Li, L.; Kandimalla, K.K. Apolipoprotein A-I crosses the blood-brain barrier through clathrin-independent and cholesterol-mediated endocytosis. J. Pharmacol. Exp. Ther. 2019, 369, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Uversky, V.N.; Fink, A.L. Protein Misfolding, Aggregation, and Conformational Diseases, Part B: Molecular Mechanisms of Conformational Diseases, 1st ed.; Springer: New York, NY, USA, 2007; pp. 61–110. [Google Scholar]
- Emamzadeh, F.N. Alpha-synuclein structure, functions, and interactions. J. Res. Med. Sci. 2016, 21, 29. [Google Scholar] [CrossRef]
- Corraliza-Gomez, M.; Sanchez, D.; Ganfornina, M.D. Lipid-binding proteins in brain health and disease. Front. Neurol. 2019, 10, 1152. [Google Scholar] [CrossRef]
- Wilhelmus, M.M.; Bol, J.G.; Van Haastert, E.S.; Rozemuller, A.J.; Bu, G.; Drukarch, B.; Hoozemans, J.J. Apolipoprotein E and LRP1 increase early in Parkinson’s disease pathogenesis. Am. J. Pathol. 2011, 179, 2152–2156. [Google Scholar] [CrossRef]
- Davis, A.A.; Inman, C.E.; Wargel, Z.M.; Dube, U.; Freeberg, B.M.; Galluppi, A.; Haines, J.N.; Dhavale, D.D.; Miller, R.; Choudhury, F.A.; et al. APOE genotype regulates pathology and disease progression in synucleinopathy. Sci. Transl. Med. 2020, 12, eaay3069. [Google Scholar] [CrossRef]
- Emamzadeh, F.N.; Aojula, H.; McHugh, P.C.; Allsop, D. Effects of different isoforms of apoE on aggregation of the α-synuclein protein implicated in Parkinson’s disease. Neurosci. Lett. 2016, 618, 146–151. [Google Scholar] [CrossRef] [Green Version]
- Eichmann, C.; Campioni, S.; Kowal, J.; Maslennikov, I.; Gerez, J.; Liu, X.; Verasdonck, J.; Nespovitaya, N.; Choe, S.; Meier, B.H.; et al. Preparation and characterization of stable α-synuclein lipoprotein particles. J. Biol. Chem. 2016, 291, 8516–8527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Videira, P.A.Q.; Castro-Caldas, M. Linking glycation and glycosylation with inflammation and mitochondrial dysfunction in Parkinson’s disease. Front. Neurosci. 2018, 12, 381. [Google Scholar] [CrossRef]
- Munch, G.; Lüth, H.J.; Wong, A.; Arendt, T.; Hirsch, E.; Ravid, R.; Riederer, P. Crosslinking of alpha-synuclein by advanced glycation end products—an early pathophysiological step in Lewy body formation? J. Chem. Neuroanat. 2000, 20, 253–257. [Google Scholar] [CrossRef]
- Padmaraju, V.; Bhaskar, J.J.; Prasada Rao, U.J.; Salimath, P.V.; Rao, K.S. Role of advanced glycation on aggregation and DNA binding properties of alpha-synuclein. J. Alzheimers Dis. 2011, 24, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Qin, Z.J.; Hu, D.; Munishkina, L.A.; Fink, A.L. α-synuclein can function as an antioxidant preventing oxidation of unsaturated lipid in vesicles. Biochemistry 2006, 45, 8135–8142. [Google Scholar] [CrossRef] [PubMed]
- Angelova, P.R.; Horrocks, M.H.; Klenerman, D.; Gandhi, S.; Abramov, A.Y.; Shchepinov, M.S. Lipid peroxidation is essential for α-synuclein-induced cell death. J. Neurochem. 2015, 133, 582–589. [Google Scholar] [CrossRef] [Green Version]
- Santner, A.; Uversky, V.N. Metalloproteomics and metal toxicology of alpha-synuclein. Metallomics 2010, 2, 378–392. [Google Scholar] [CrossRef]
- Kim, S.H.; Yadav, D.; Kim, S.J.; Kim, J.R.; Cho, K.H. High consumption of iron exacerbates hyperlipidemia, atherosclerosis, and female sterility in zebrafish via acceleration of glycation and degradation of Serum Lipoproteins. Nutrients 2017, 9, 690. [Google Scholar] [CrossRef] [Green Version]
- Koldamova, R.P.; Lefterov, I.M.; Lazo, J.S. Apolipoprotein A-I directly interacts with amyloid precursor protein and inhibits amyloid beta aggregation and toxicity. Biochemistry 2001, 40, 3553–3560. [Google Scholar] [CrossRef] [PubMed]
- Emamzadeh, F.N. Role of apolipoproteins and α-synuclein in Parkinson’s Disease. J. Mol. Neurosci. 2017, 62, 344–355. [Google Scholar] [CrossRef]
- De Boer, A.G.; van der Sandt, I.C.; Gaillard, P.J. The role of drug transporters at the blood-brain barrier. Annu. Rev. Pharmacol. Toxicol. 2003, 43, 629–656. [Google Scholar] [CrossRef]
- Yoon, J.H.; Cho, K.H. A point mutant of apolipoprotein A-I (V156K) showed enhancement of cellular insulin secretion and potent activity of facultative regeneration in zebrafish. Rejuvenation Res. 2012, 15, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.H. Structural and functional impairments of reconstituted high-density lipoprotein by incorporation of recombinant β-Amyloid42. Molecules 2021, 26, 4317. [Google Scholar] [CrossRef] [PubMed]
- Matsudaira, P. Sequence from picomole quantities of proteins electroblotted onto polyvinylidene difluoride membranes. J. Biol. Chem. 1987, 262, 10035–10038. [Google Scholar] [CrossRef]
- Jang, W.; Shim, J.; Lee, D.Y.; Dutta, P.; Kim, J.R.; Cho, K.H. Rapid detection of dysfunctional highdensity lipoproteins using isoelectric focusing-based microfluidic device to diagnose senescence-related disease. Electrophoresis 2011, 32, 3415–3423. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Yang, J.T.; Martinez, H.M. Determination of the secondary structures of proteins by circular dichroism and optical rotatory dispersion. Biochemistry 1972, 11, 4120–4131. [Google Scholar] [CrossRef]
- Han, J.M.; Jeong, T.S.; Lee, W.S.; Choi, I.; Cho, K.H. Structural and functional properties of V156K and A158E mutants of apolipoprotein A-I in the lipid-free and lipid-bound states. J. Lipid Res. 2005, 46, 589–596. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.H.; Jonas, A. A key point mutation (V156E) affects the structure and functions of human Apolipoprotein A-I. J. Biol. Chem. 2000, 275, 26821–26827. [Google Scholar] [CrossRef]
- Park, K.H.; Shin, D.G.; Kim, J.R.; Cho, K.H. Senescence-related truncation and multimerization of apolipoprotein A-I in high-density lipoprotein with an elevated level of advanced glycated end products and cholesteryl ester transfer activity. J. Gerontol. A. Biol. Sci. Med. Sci. 2010, 65, 600–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brewer, H.B., Jr.; Ronan, R.; Meng, M.; Bishop, C. Isolation and characterization of apolipoproteins A-I, A-II, and A-IV. Methods Enzymol. 1986, 128, 223–246. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Cho, K.H. Synthesis of reconstituted high density lipoprotein (rHDL) containing apoA-I and apoC-III: The functional role of apoC-III in rHDL. Mol. Cells 2009, 27, 291–297. [Google Scholar] [CrossRef]
- Pownall, H.J.; Massey, J.B.; Kusserow, S.K.; Gotto, A.M., Jr. Kinetics of lipid–protein interactions: Effect of cholesterol on the association of human plasma high-density apolipoprotein A-I with L-alpha-dimyristoylphosphatidylcholine. Biochemistry 1979, 18, 574–579. [Google Scholar] [CrossRef]
- Park, K.H.; Jang, W.; Kim, K.Y.; Kim, J.R.; Cho, K.H. Fructated apolipoprotein A-I showed severe structural modification and loss of beneficial functions in lipid-free and lipid-bound state with acceleration of atherosclerosis and senescence. Biochem. Biophys. Res. Commun. 2010, 392, 295–300. [Google Scholar] [CrossRef]
- McPherson, J.D.; Shilton, B.H.; Walton, D.J. Role of fructose in glycation and cross-linking of proteins. Biochemistry 1988, 27, 1901–1907. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid-Free | Lipid-Bound (POPC:FC:apoA-I: α-syn = 95:5:1:1) | |||||
|---|---|---|---|---|---|---|
| α-helix Content | WMF | Isoelectric Point (pI) | α-helix Content | WMF | Particle Size (Å) | |
| apoA-I alone | 56 | 341 | 6.4 | 76 | 337 | 86, 78 |
| α-syn alone | - | - | 4.5 | - | 65 | |
| apoA-I + α-syn | 51 | 332 | - | 45 | 339 | 92, 74, 69, 65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, K.-H. Structural and Functional Changes of Reconstituted High-Density Lipoprotein (HDL) by Incorporation of α-synuclein: A Potent Antioxidant and Anti-Glycation Activity of α-synuclein and apoA-I in HDL at High Molar Ratio of α-synuclein. Molecules 2021, 26, 7485. https://doi.org/10.3390/molecules26247485
Cho K-H. Structural and Functional Changes of Reconstituted High-Density Lipoprotein (HDL) by Incorporation of α-synuclein: A Potent Antioxidant and Anti-Glycation Activity of α-synuclein and apoA-I in HDL at High Molar Ratio of α-synuclein. Molecules. 2021; 26(24):7485. https://doi.org/10.3390/molecules26247485
Chicago/Turabian StyleCho, Kyung-Hyun. 2021. "Structural and Functional Changes of Reconstituted High-Density Lipoprotein (HDL) by Incorporation of α-synuclein: A Potent Antioxidant and Anti-Glycation Activity of α-synuclein and apoA-I in HDL at High Molar Ratio of α-synuclein" Molecules 26, no. 24: 7485. https://doi.org/10.3390/molecules26247485
APA StyleCho, K.-H. (2021). Structural and Functional Changes of Reconstituted High-Density Lipoprotein (HDL) by Incorporation of α-synuclein: A Potent Antioxidant and Anti-Glycation Activity of α-synuclein and apoA-I in HDL at High Molar Ratio of α-synuclein. Molecules, 26(24), 7485. https://doi.org/10.3390/molecules26247485