
Water Regime Monitoring of the Royal Walnut (Juglans regia L.) Using Sap Flow and Dendrometric Measurements


Abstract
:1. Introduction
2. Materials and Methods
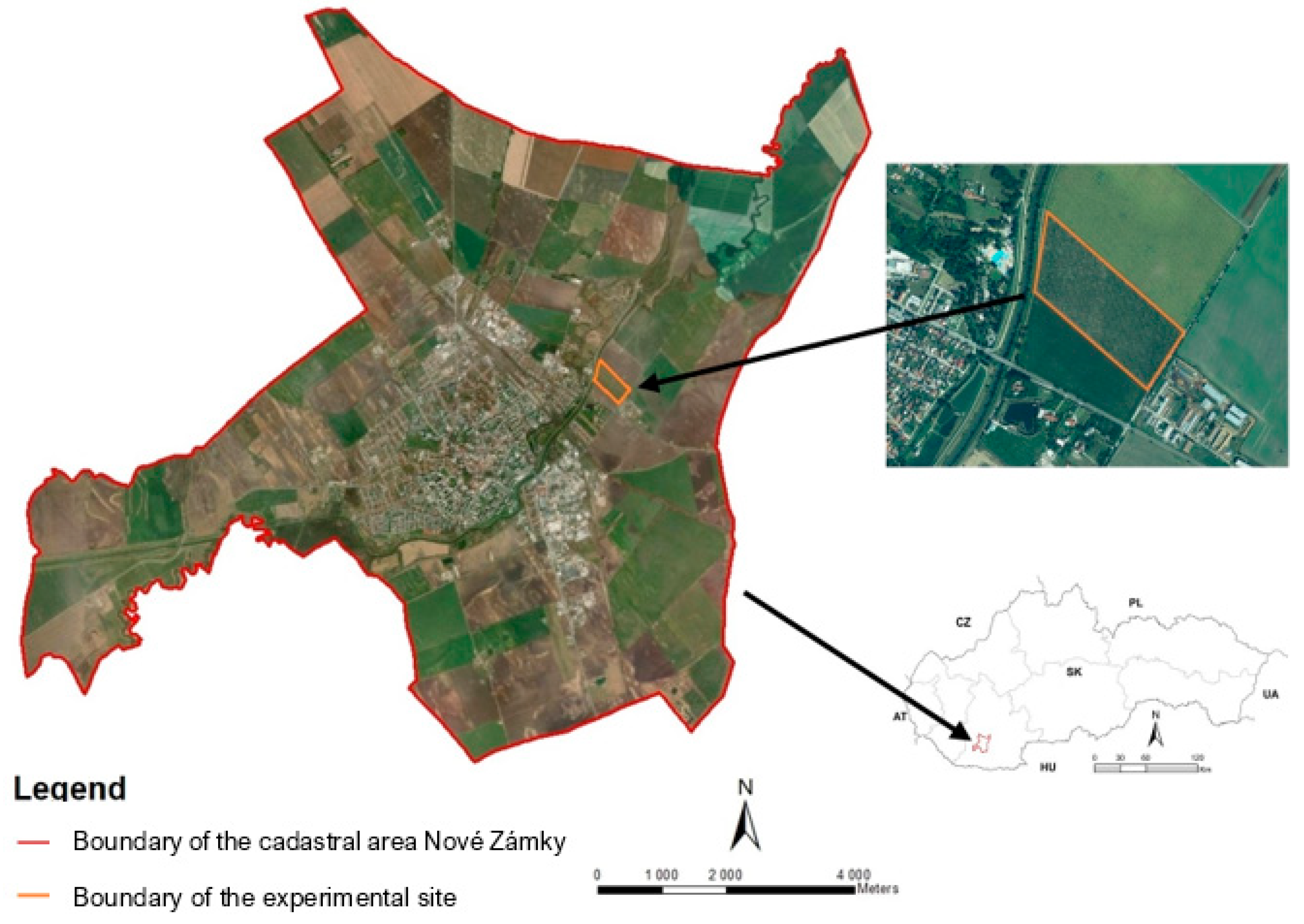
2.1. Experimental Site
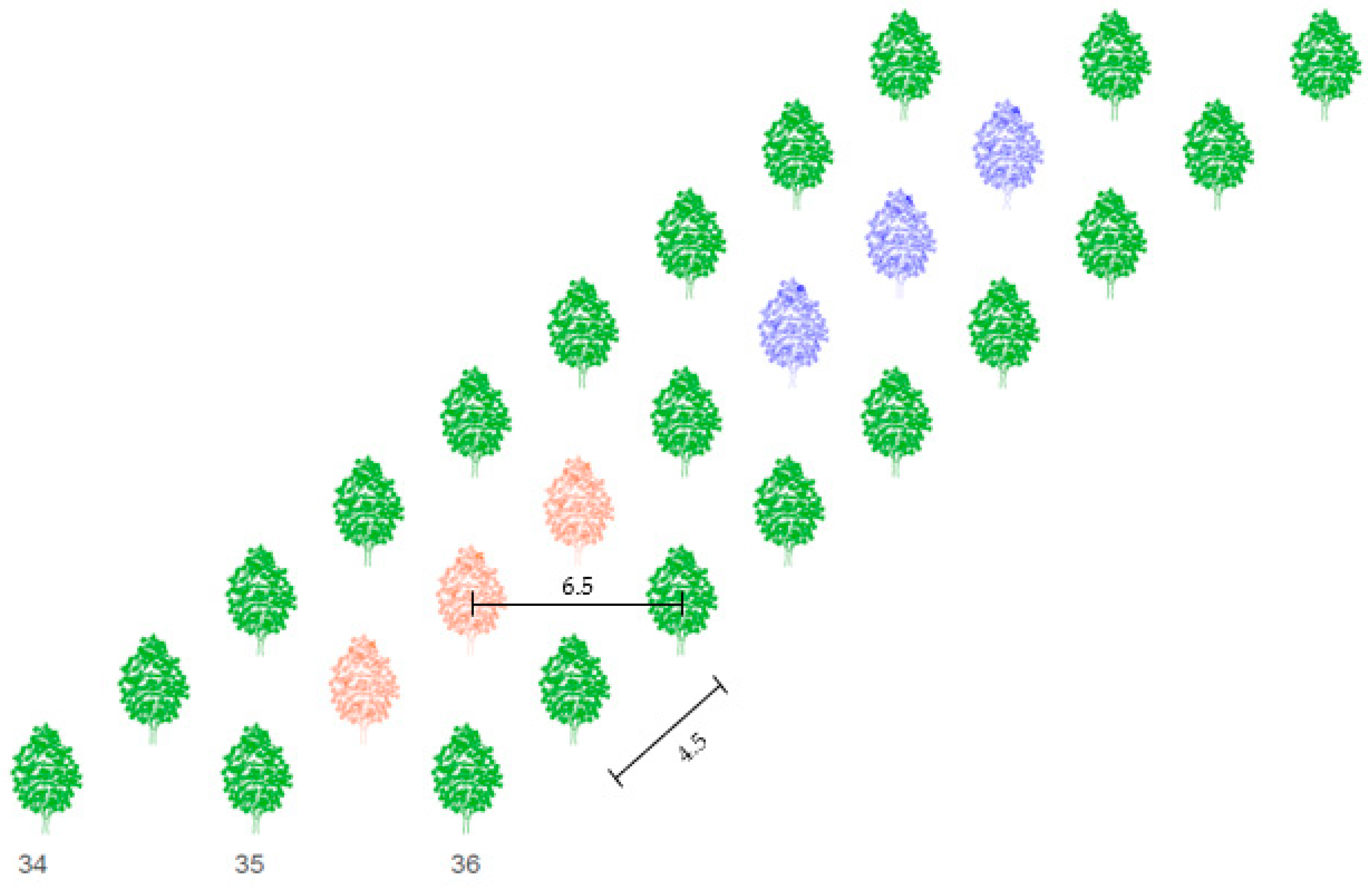
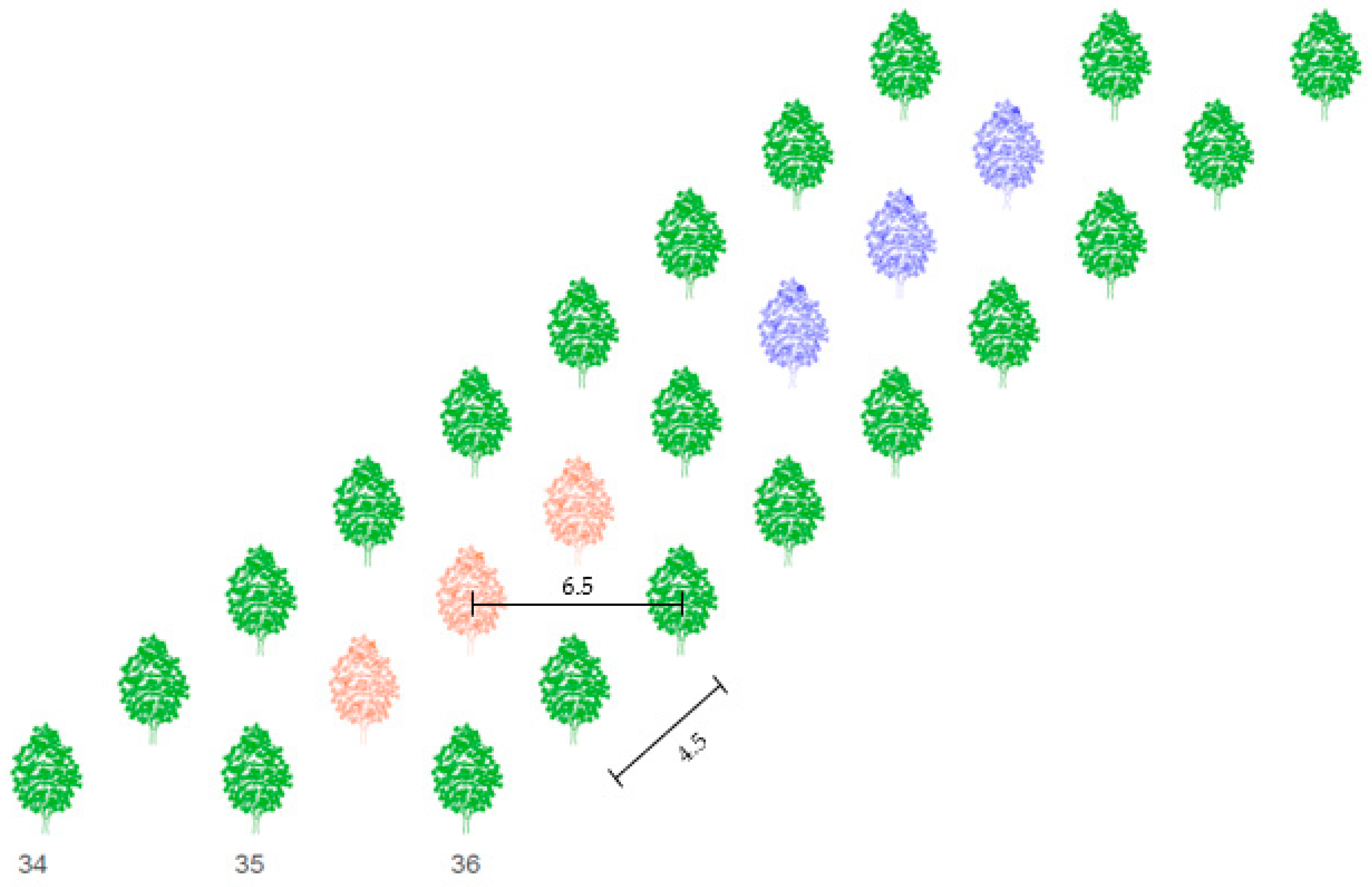
2.2. Experimental Design
Crop
2.3. Field Measurements
2.3.1. Soil Moisture
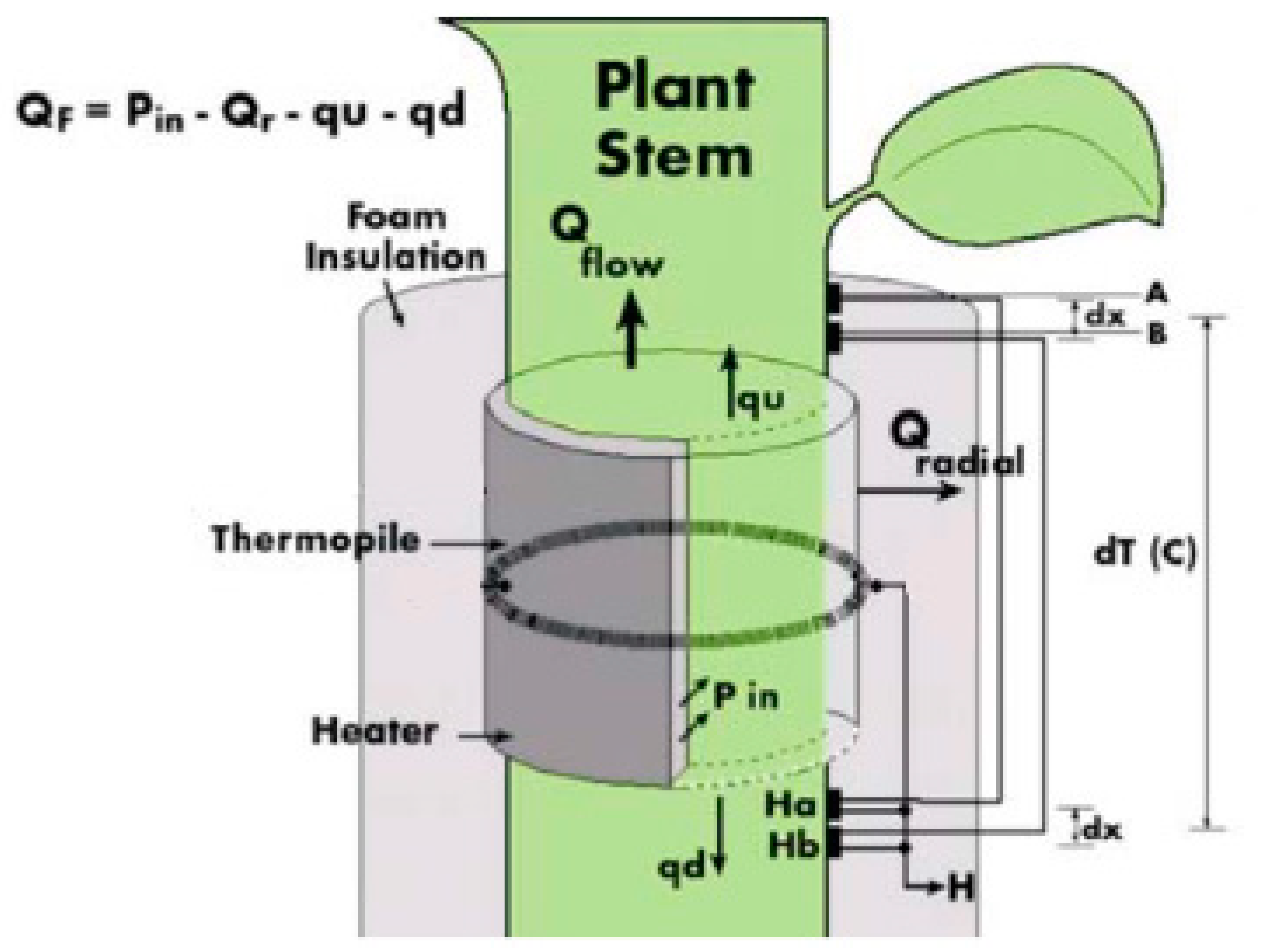
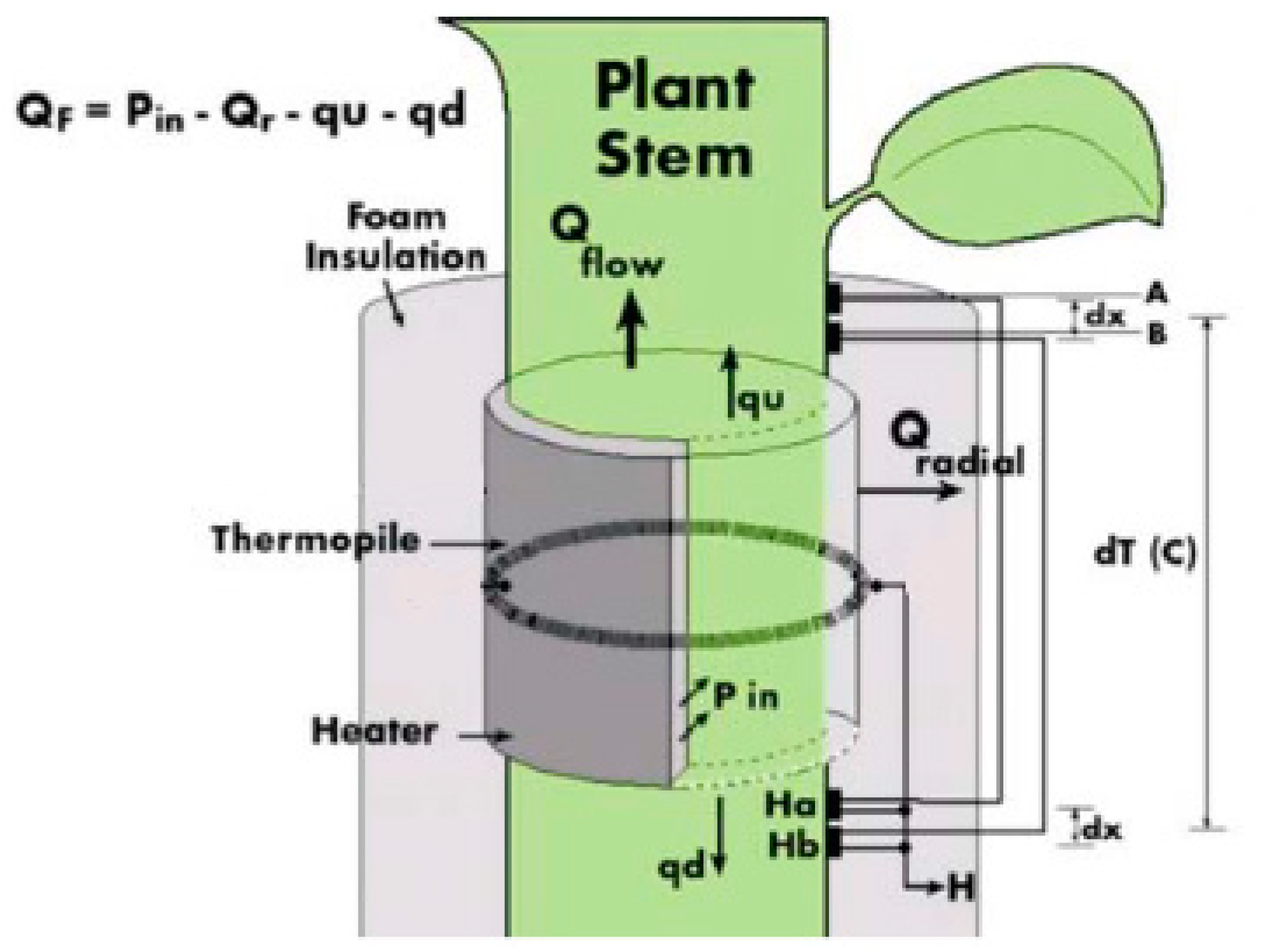
2.3.2. Sap Flow
2.3.3. Dendrometric Measurements
2.4. Statistical Analyses
- Sap flow and air temperature;
- Sap flow and soil moisture;
- Dendrometric changes and air temperature;
- Dendrometric changes and soil moisture;
- Sap flow and dendrometric changes.
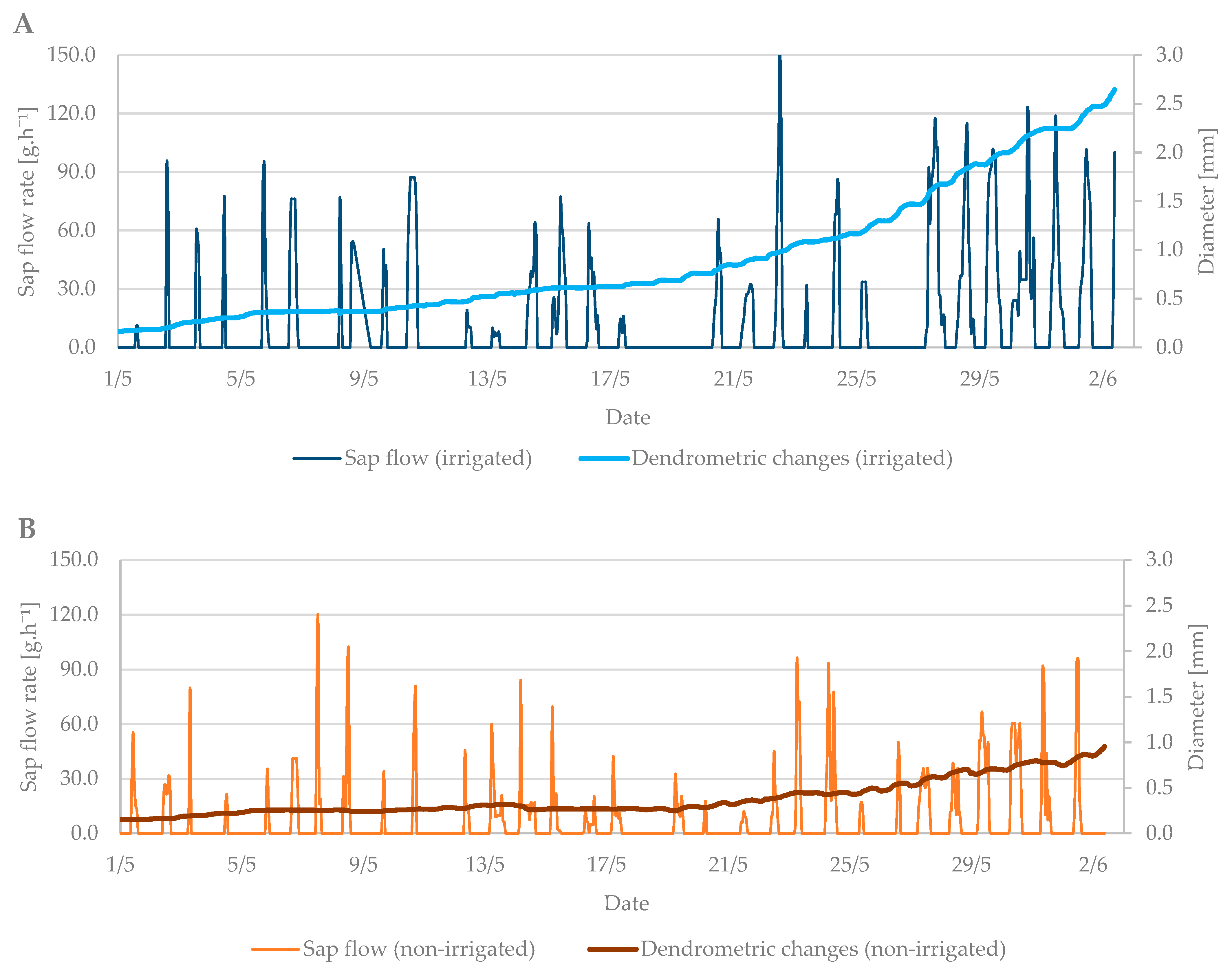
3. Results
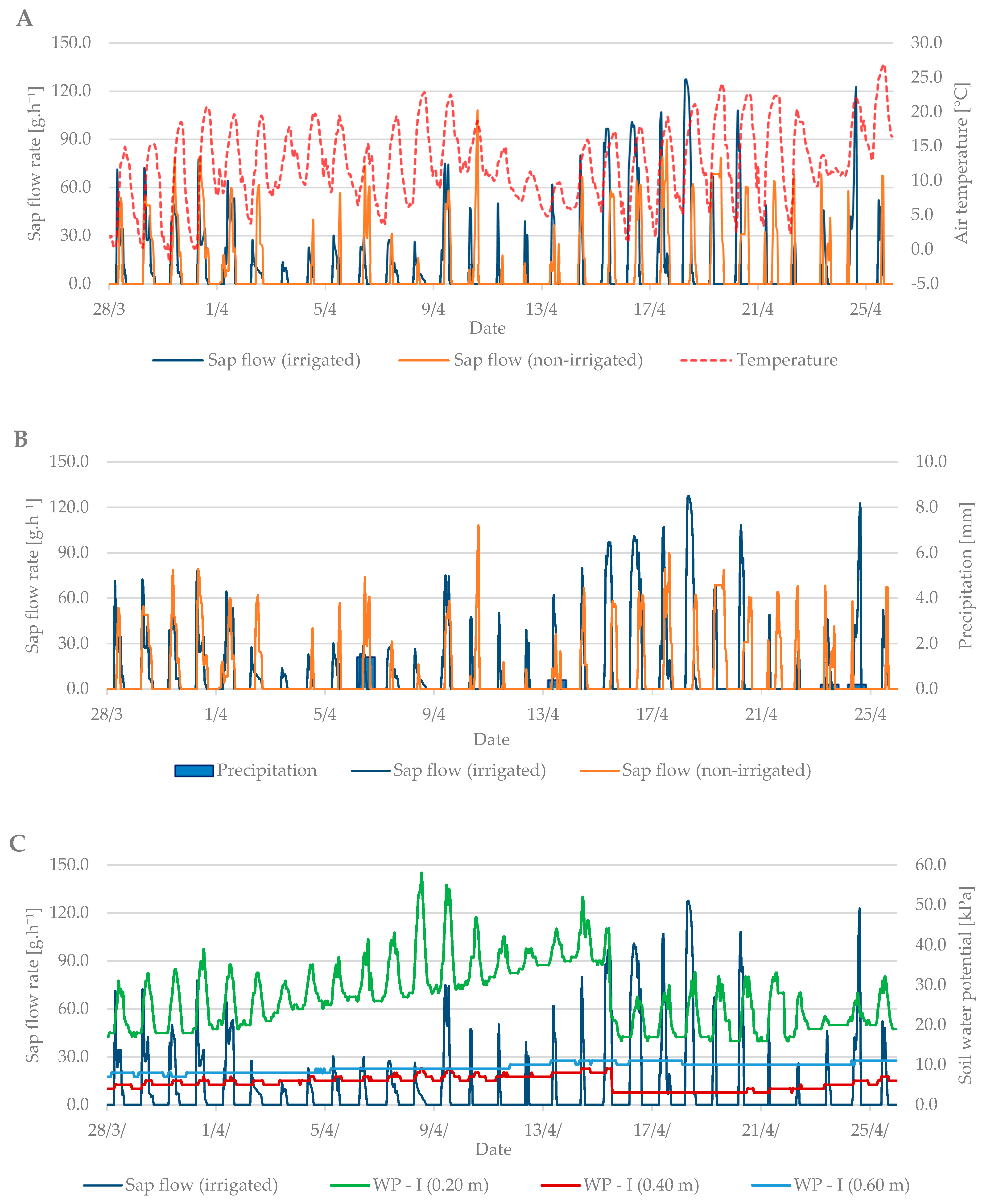
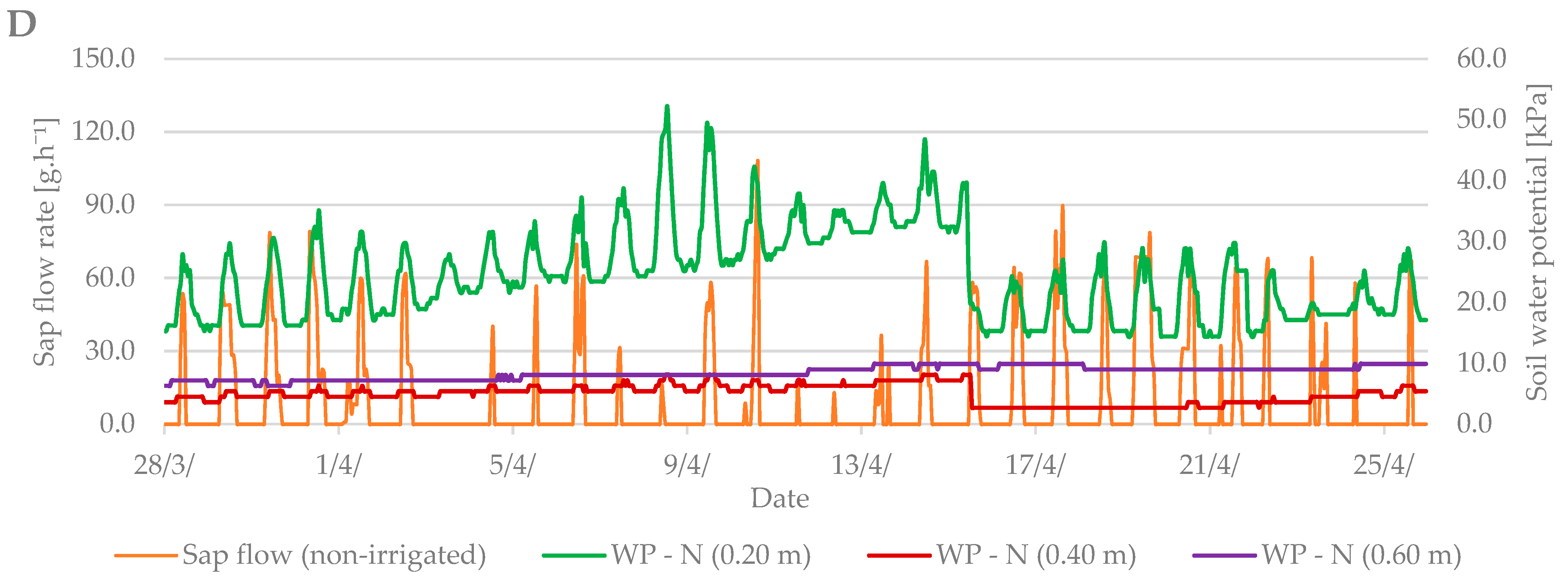
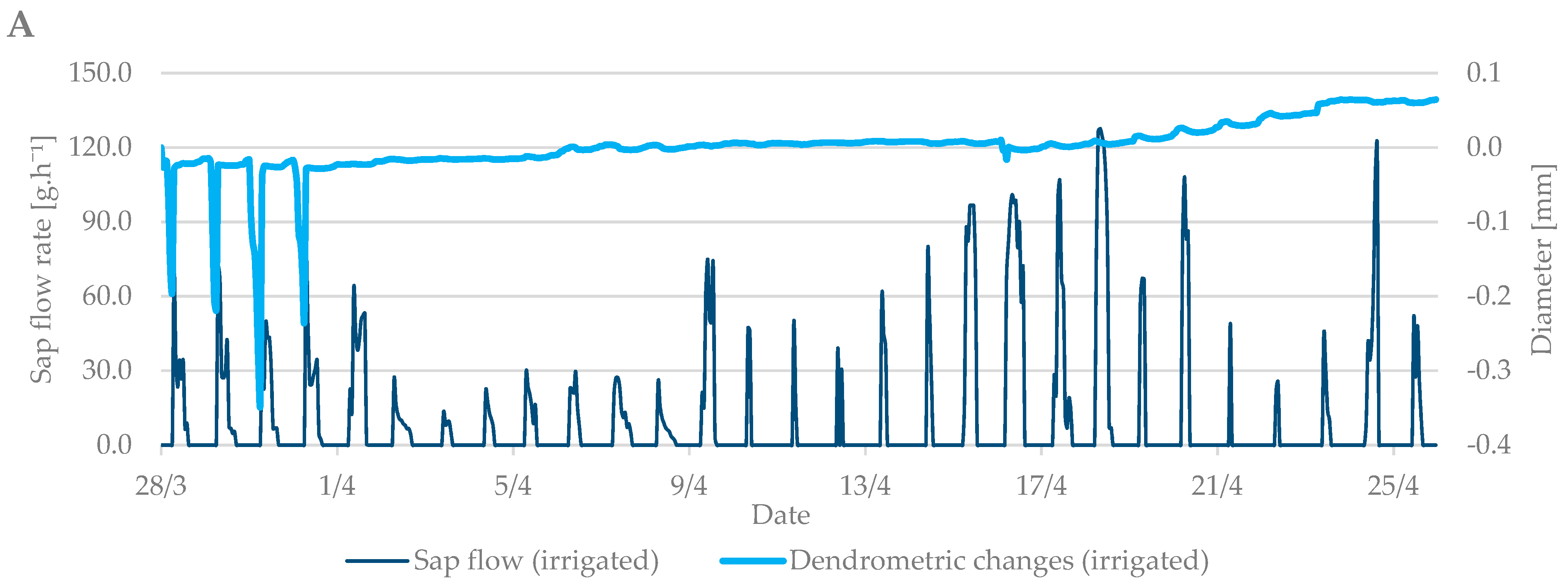
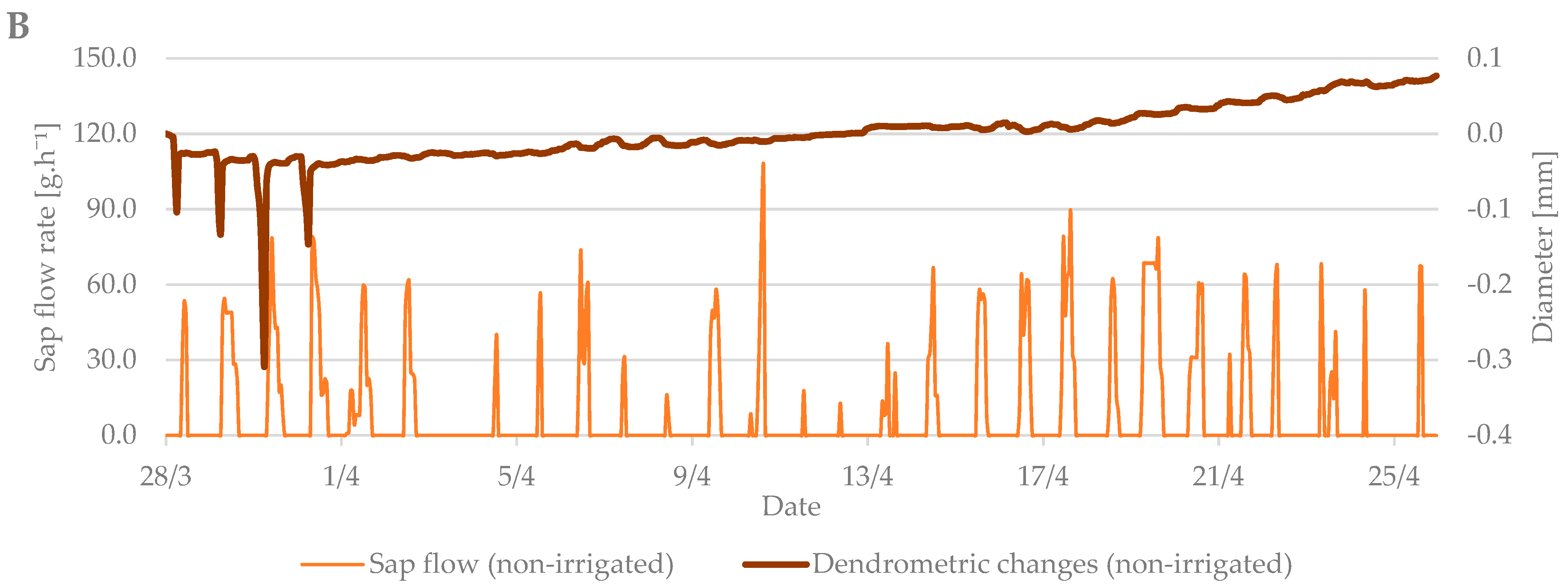
3.1. Budbreak Period
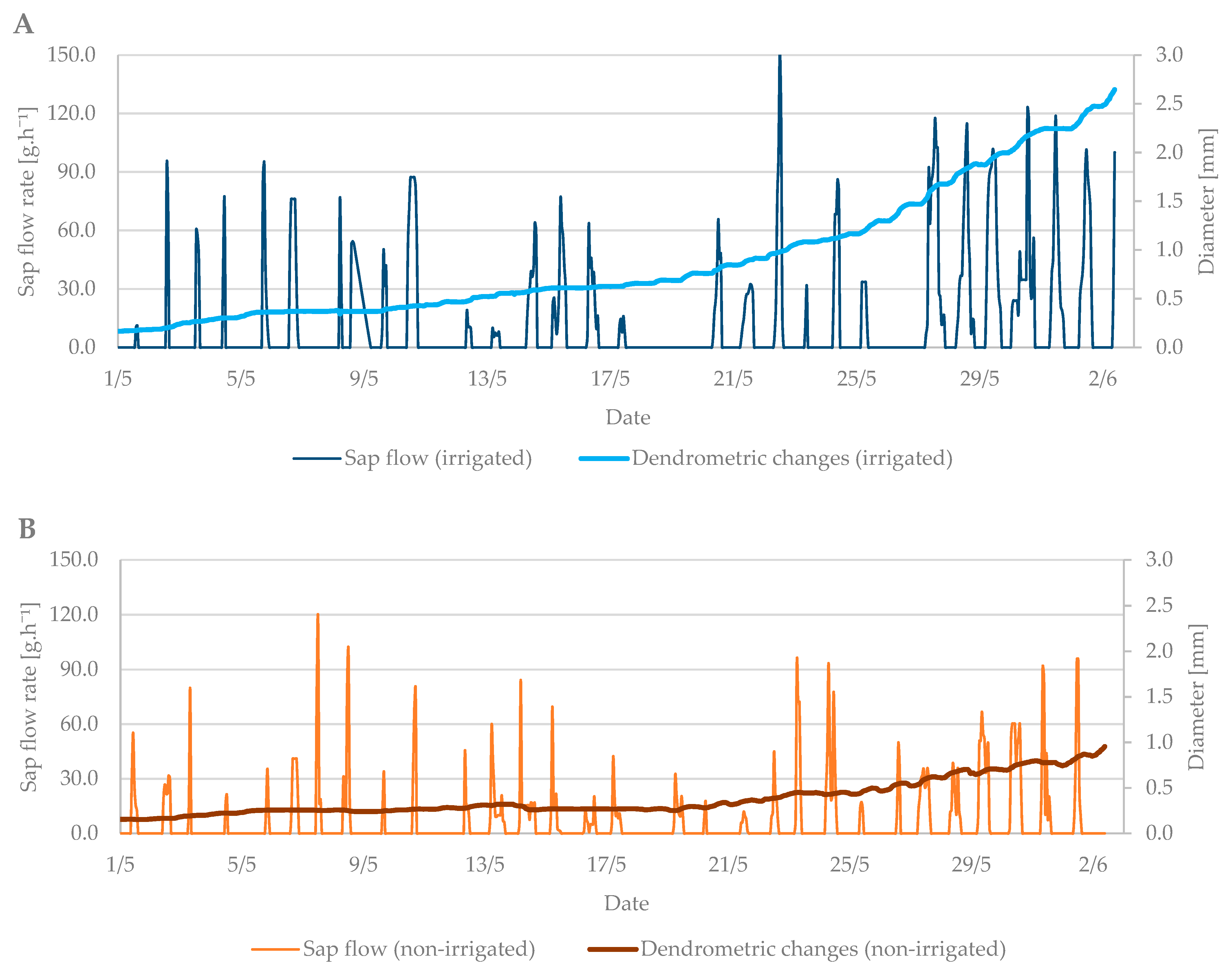
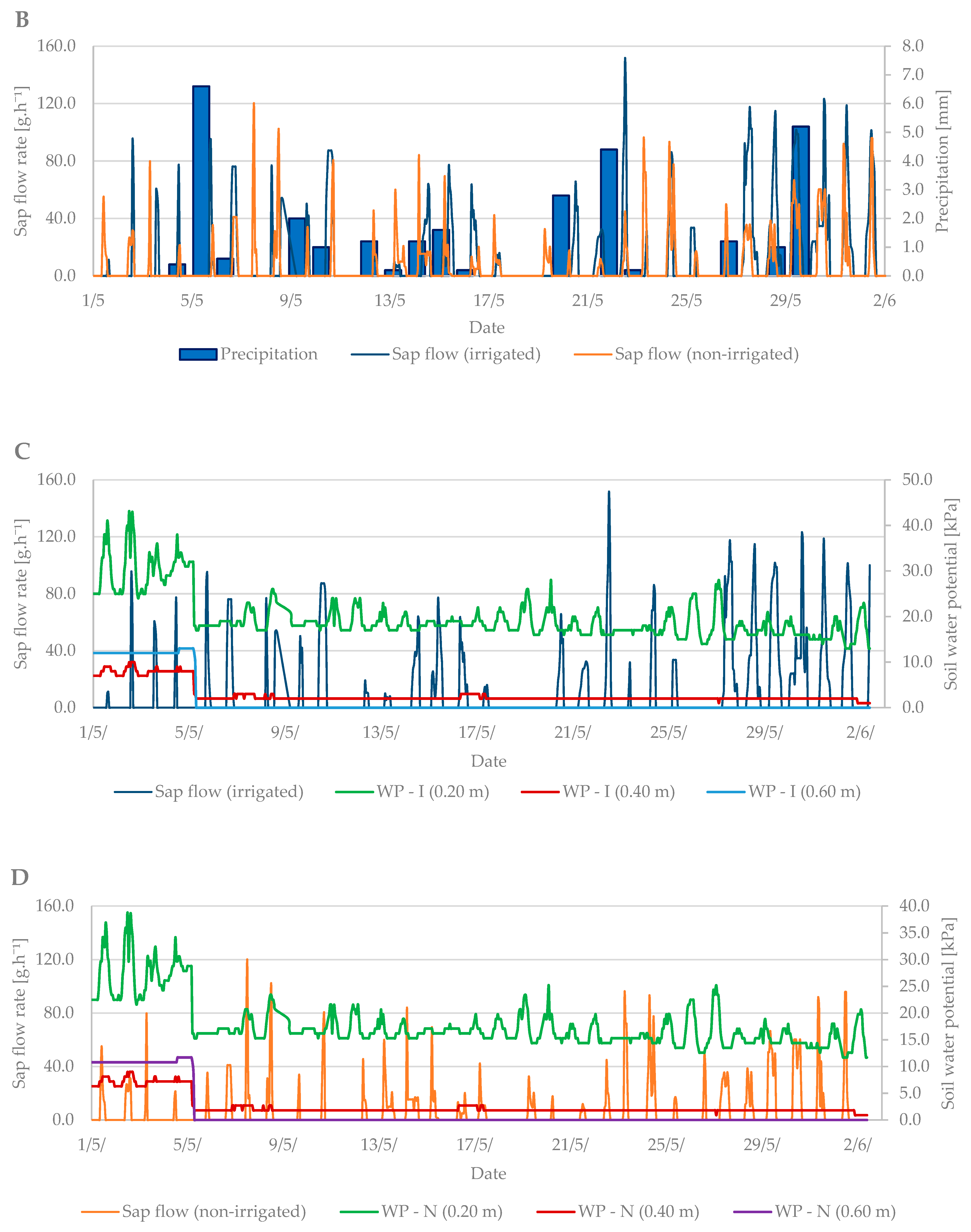
3.2. Flowering Period
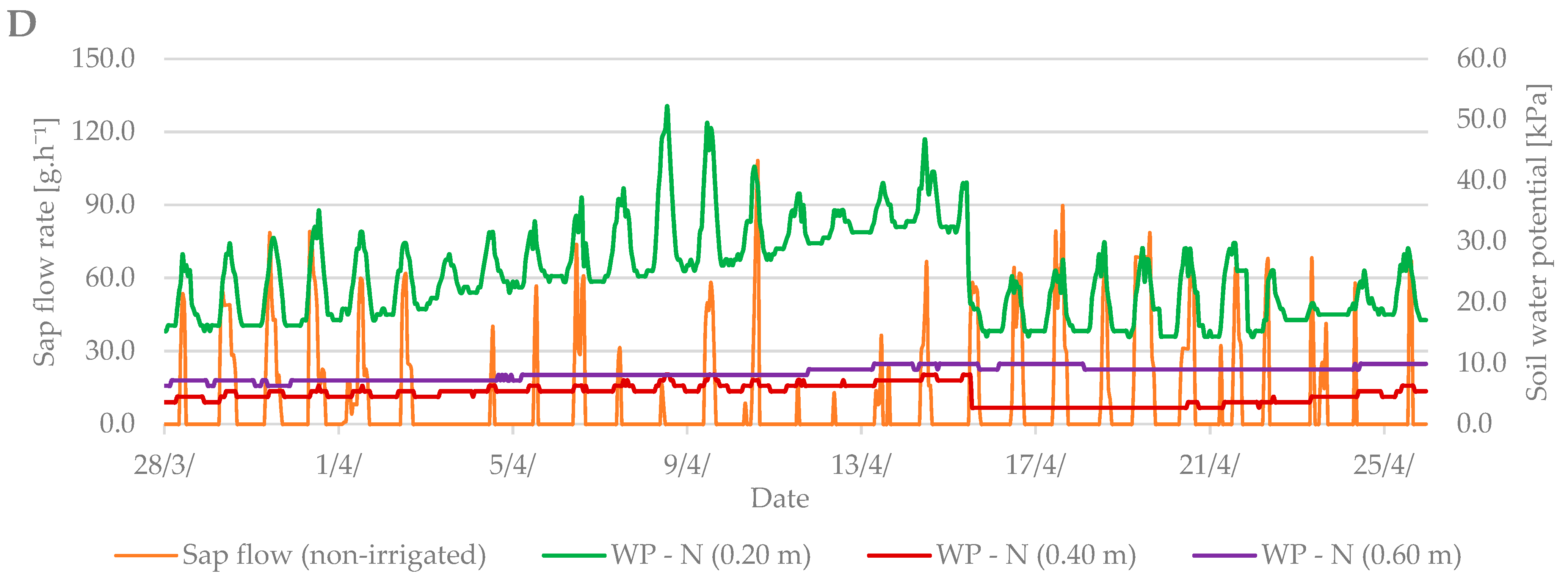
3.3. Relation between Sap Flow, Dendrometric Changes and Selected Properties of Environment (Air Temperature and Soil Moisture)
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Čimo, J.; Aydın, E.; Šinka, K.; Tárník, A.; Kišš, V.; Halaj, P.; Toková, L.; Kotuš, T. Change in the Length of the Vegetation Period of Tomato (Solanum lycopersicum L.), White Cabbage (Brassica oleracea L. var. capitata) and Carrot (Daucus carota L.) Due to Climate Change in Slovakia. Agronomy 2020, 10, 1110. [Google Scholar] [CrossRef]
- Brázdil, R.; Trnka, M. Sucho v Českých Zemích—Minulost, Současnost, Budoucnost (Drought in the Czech Lands—Past, Present, Future); Centrum Výzkumu Globální Změny Akademie věd České Republiky: Brno, Czech Republic, 2015; p. 402. [Google Scholar]
- Chhapekar, S.S.; Jaiswal, V.; Ahmad, I.; Gaur, R.; Ramchiary, N. Progress and Prospets in Capsicum Breeding for Biotic and Abiotic Stresses. In Biotic and Abiotic Stress Tolerance in Plants; Vats, S., Ed.; Department of Bioscience & Biotechnology, Banasthali Vidyapith: Rajastna, India, 2018; pp. 279–322. [Google Scholar]
- Bárek, V.; Halaj, P.; Igaz, D. The Influence of Climate Change on Water Demands for Irrigation of Special Plants and Vegetables in Slovakia. In Bioclimatology and Natural Hazards; Springer: Dordrecht, The Netherlands, 2009; pp. 271–282. [Google Scholar]
- Wetherald, R.T.; Manabe, S. Simulation of hydrologic changes associated with global warming. J. Geophys. Res. Atmos. 2003, 107, 15. [Google Scholar] [CrossRef] [Green Version]
- Trenberth, K.E.; Van der Schriee, G.; Jones, P.D.; Barichivich, J.; Briffa, K.R.; Sheffield, J. Global warming and changes in drought. Nat. Clim. Chang. 2014, 4, 17–22. [Google Scholar] [CrossRef]
- Sangiorgio, D.; Cellini, A.; Pastore, C.; Onofrietti, C.; Spinelli, F. Facing Climate Change: Application of Microbial Biostimulants to Mitigate stress in Horticultural Crops. Agronomy 2020, 10, 794. [Google Scholar] [CrossRef]
- Walnut Production Worldwide from 2012/13 to 2019/20. Available online: https://www.statista.com/statistics/675967/walnut-production-worldwide/ (accessed on 30 September 2021).
- Baran, M.F.; Gökdoğan, O.; Oğuz, H.İ. Determining the Energy Usage Efficiency of Walnut (Juglans regia L.) Cultivation in Turkey. Erwerbs-Obstbau 2017, 59, 77–82. [Google Scholar] [CrossRef]
- Vahdati, K.; Lotfi, N. Abiotic Stress Tolerance in plants with Emphasizing on Drought and Salinity Stresses in Walnut. In Abiotic Stress—Plant Responses and Applications in Agriculture; InTech: Rijeka, Croatia, 2013; pp. 307–365. [Google Scholar]
- Sadat-Hosseini, M.; Vahdati, K.; Boroomand, N.; Mehdi-Arab, M.; Asadi-Yekta, S. How Walnut Roots Respond to Drought Stress: A Morphological and Anatomical Study. South-West. J. Hortic. Biol. Environ. 2019, 10, 51–64. [Google Scholar]
- Kozlowski, T.; Pallardy, S. Acclimation and Adaptive Responses of Woody Plants to Environmental Stresses. Bot. Rev. 2002, 68, 270–334. [Google Scholar] [CrossRef]
- Lampinen, B.; Buchner, B.; Fulton, A.; Grant, J.; Mills, N.; Prichard, T.; Schwankl, L.; Shackel, K.; Gilles, C.; Little, C.; et al. Irrigation management in walnut using evapotranspiration, soil and plant based data. Calif. Walnut Board 2004, 24, 113–136. [Google Scholar]
- Buchner, R.P.; Lindow, S.E.; Adaskaveg, J.E.; Gilles, C.K.; Koutsoukis, R. Walnut blight control investigation Tehama 2008. Calif. Walnut Board 2008, 7, 195–200. [Google Scholar]
- Jerszurki, D.; Couvreur, V.; de Carvalho Ramos Silva, L.; Matsumoto, N.; Shackel, K.; Moretti de Souza, J.L.; Hopmans, J. Impact of growth and hydraulic conductance on canopy carbon-water relations of young walnut trees (Juglans regia L.) under drought. Sci. Hortic. 2017, 226, 342–352. [Google Scholar] [CrossRef]
- Navara, J. Orech kráľovský (Juglans regia L.) a Jeho Pestovanie (Californian Walnut (Juglans regia L.) and its cultivation); Veda: Bratislava, Slovakia, 2010; p. 76. [Google Scholar]
- Baker, J.M.; Van Bavel, C.H.M. Measurement of mass flow of water in the stems of herbaceous plants. Plants Cell Environ. 1987, 10, 777–782. [Google Scholar]
- Fernández, E.; Čermák, J.; Cohen, Y.; Ferreira, I.; Nadezhdina, N.; Testi, L.; Steppe, K. Methods to estimate sap flow. ISHS Work. Group Sap Flow 2017, 9, 1–9. [Google Scholar]
- Kišš, V.; Manina, M. Optimization of irrigation based on SAP flow measurement. In Proceedings of the 18th International Multidisciplinary Scientific GeoConference SGEM2018, Albena, Bulgaria, 2–8 July 2018; p. 8. [Google Scholar]
- ENVIROPORTAL Atlas Krajiny Slovenskej republiky. Available online: http://geo.enviroportal.sk/atlassr/ (accessed on 8 February 2021).
- Manažment sucha—Zhodnotenie rokov 2011 a 2012 podľa indexov sucha. Available online: http://www.shmu.sk/File/Hydrologia/Publikacna_cinnost/2014/2014_DenDunaja_Stastny_Turna_Manazment%20sucha%20-%20podla%20indexov%20sucha.pdf (accessed on 10 August 2020).
- Mills, N.J.; Lampinen, B.; Julier, J.; Grant, J. 2001 Managing codling moth in Walnuts: Parasitism and nut susceptibility. Walnut Res. Rep. Database 2001, 14, 363–376. [Google Scholar]
- Mora-Sala, B.; Berbegal, M.; Abad-Campos, P. The use of qPCR reveals a high frequency of Phytophthora quercina in two Spanish holm oak areas. Forests 2018, 9, 697. [Google Scholar] [CrossRef] [Green Version]
- Šiška, B.; Špánik, F.; Repa, Š.; Gálik, M. Praktická biometeorológia, 5th ed.; SUA: Nitra, Slovakia, 2008; pp. 104–144. [Google Scholar]
- Hájková, L.; Richterová, D.; Kohut, M. Časová variabilita nástupu fenofází ovocných dřevin sledovaných ve fenologické síti ČHMÚ za období 1991–2010. In Proceedings of the Rostliny v Podmínkách Měnícího se Klimatu Conference (Plants in Conditions of Changing Climate, Conference), Lednice, Czech Republic, 20–21 October 2011; p. 12. [Google Scholar]
- Śpitalniak, M.; Lejcuś, K.; Dąbrowska, J.; Garlikowski, D.; Bogacz, A. The influence of a water absorbing geocomposite on soil water retention and soil matric potential. Water 2019, 11, 1731. [Google Scholar] [CrossRef] [Green Version]
- Ishimwe, E.; Garner, C.D.; Coffman, R.A. Full-scale hydro-mechanical properties of vertical flow bed treatment soils. Geotech. Geol. Eng. 2018, 36, 2735–2748. [Google Scholar] [CrossRef]
- Irrometer Soil Water Management Since 1991. Available online: https://www.irrometer.com/basics.html (accessed on 12 February 2021).
- Lascano, R.J.; Goebel, T.S.; Booker, J.; Baker, J.T.; Gitz, D.C., III. The stem heat balance method to measure transpiration: Evaluation of a new sensor. Agric. Sci. 2016, 7, 604–620. [Google Scholar] [CrossRef] [Green Version]
- Dynagage Sap Flow Sensor User Manual. Available online: http://www.dynamax.com/images/uploads/papers/Dynagage_Manual.pdf (accessed on 16 February 2021).
- Han, M.; Zhang, H.; DeJonge, K.C.; Comas, L.H.; Gleason, S. Comparison of three crop water stress index model with sap flow measurements in Maize. Agric. Water Manag. 2018, 203, 366–375. [Google Scholar] [CrossRef]
- Smith, D.M.; Allen, S.J. Measurement of sap flow in plant stems. J. Exp. Bot. 1996, 47, 1833–1844. [Google Scholar] [CrossRef] [Green Version]
- Grime, V.L.; Morrison, J.I.L.; Simmonds, L.P. Including the heat storage term in sap flow measurements with the stem heat balance method. Agric. For. Meteorol. 1995, 74, 1–25. [Google Scholar] [CrossRef]
- Wilson, K.B.; Hanson, P.J.; Mulholland, P.J.; Baldochci, D.D.; Wullschleger, S.D. A comparison of methods for determining forest evapotranspiration and its components: Sap-flow, soil water budget, eddy covariance and catchment water balance. Agric. For. Meteorol. 2001, 106, 153–168. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, F.; Yuan, L.; Wang, Q.; Zheng, K.; Zhao, C. Environmental factors effect on stem radial variations of Picea crassifolia in Qilian Mountains, Northwestern China. Forests 2016, 7, 210. [Google Scholar] [CrossRef] [Green Version]
- Takáč, J.; Bárek, V.; Halaj, P.; Igaz, D.; Jurík, Ľ. Possible impact of climate change on soil water content in Danubian Lowland. Cereal Res. Commun. 2008, 36, 1623–1626. [Google Scholar]
- He, Q.-Y.; Yan, M.-J.; Miyazawa, Y.; Chen, Q.-W.; Cheng, R.-R.; Otsuki, K.; Yamanaka, N.; Du, S. Sap flow changes and climatic responses over multiple-year treatment of rainfall exclusion in a sub-humid black locust plantation. For. Ecol. Manag. 2020, 457, 11. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, Z.; Li, Z.; Tang, J.; Caldwell, J.; Zhang, W. Biophysical control of whole tree transpiration under an urban environment in Nothern China. J. Hydrol. 2011, 402, 388–400. [Google Scholar] [CrossRef]
- Chang, X.; Zhao, W.; He, Z. Radial pattern of sap flow and response to microclimate and soil moisture in Qinghai spruce (Picea crassifolia) in the upper Heihe River Basin of arid northwestern China. Agric. For. Meteorol. 2014, 187, 14–21. [Google Scholar] [CrossRef]
- Jones, T.; Winbourne, J.B.; Hutyra, L.R. Ribbonized sap flow: An emerging technology for the integration of sap flow sensor components onto a single platform. Ecosphere 2020, 11, 6. [Google Scholar] [CrossRef]
- Santiago, L.S.; Goldstein, G.; Meinzer, F.C.; Fownes, J.; Mueller-Dombois, D. Transpiration and forest structure in relation to soil waterlogging in a Hawaiian montane cloud forest. Tree Physiol. 2000, 20, 673–681. [Google Scholar] [CrossRef] [Green Version]
- Lavrič, M.; Eler, K.; Ferlan, M.; Vodnik, D.; Gričar, J. Chronological Sequence of Leaf Phenology, Xylem and Phloem Formation and Sap Flow of Quercus pubescens from Abandoned Karst Grasslands. Front. Plant Sci. 2017, 8, 314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zha, T.; Qian, D.; Jia, X.; Bai, Y.; Bourque, C.P.-A.; Ma, J.; Feng, W.; Wu, B.; Peltola, H. Soil moisture control of sap-flow response to biophycal factors in a desert-shrub species, Artemisia ordosica. Biogeosciences 2017, 14, 4533–4544. [Google Scholar] [CrossRef] [Green Version]
- Nakano, Y.; Iwasaki, N. Determination of phloem sap flow rate using a combination of the heat balance method and girdling in Cintrus. Agric. For. Meteology 2019, 278, 7. [Google Scholar]
- Lenz, A.; Hoch, G.; Körner, C. Early season temperature controls cambial activity and total tree ring width at the alpine treeline. Plant Ecol. Divers. 2013, 6, 365–375. [Google Scholar] [CrossRef]
- Sánchez-Costa, E.; Poyatos, R.; Sabaté, S. Contrasting growth and water use strategies in four co-occurring Mediterranean tree species revealed by concurrent measurements of sap flow and stem diameter variations. Agric. For. Meteorol. 2015, 207, 7. [Google Scholar] [CrossRef] [Green Version]
- Dang, H.; Lu, P.; Yang, W.; Han, H.; Zhang, J. Drought-induced reductions and limited recovery in the radial growth, transpiration, and canopy stomatal conductance of Mongolian Scots Pine (Pinus sylvestris var. Mongolica Litv.): A five-year observation. Forests 2019, 10, 1143. [Google Scholar] [CrossRef] [Green Version]
- Güney, A.; Küpper, M.; Rathgeber, C.; Şahin, M.; Zimmermann, R. Intra-annual stem growth dynamics of Lebanon Cedar along climatic gradients. Trees 2017, 31, 587–606. [Google Scholar] [CrossRef] [Green Version]
- Tian, Q.; He, Z.; Xiao, S.; Du, J.; Peng, X.; Chen, L.; Lin, P.; Zhu, X.; Ding, A. Growing season stem water status assessment of Qinghai spruce throught the sap flow and stem radiam variations in the Qilian mountains of China. Forests 2018, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Herzog, K.M.; Häsler, R.; Thum, R. Diurnal changes in the radius of subalpine Norway spruce stem: Their relation to the sap flow and their use to estimate transpiration. Trees 1995, 10, 94–101. [Google Scholar] [CrossRef]
- Malheiro, A.C.; Pires, M.; Conceição, N.; Claro, A.M.; Dinis, L.-T.; Moutinho-Pereira, J. Linking sap flow and trunk diameter measurements to assess water dynamics of Touriga-nacional grapevines trained in Cordon and Guyot systems. Agriculture 2020, 10, 315. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, X.; Zhao, S.; Qi, G.; Guo, S. The depth of water taken up by Walnut trees during different phenological stages in an irrigated arid hilly area in the Taihang mountains. Forests 2019, 10, 121. [Google Scholar] [CrossRef] [Green Version]
- Bhusal, N.; Han, S.-G.; Yoon, T.-M. Impact of drouhght stress on photosynthetic response, leaf water potential and stem sap flow in two cultivars of bi-leader apple trees (Malus x domestica Borkh.). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Fernández, J.E.; Green, S.R.; Caspari, H.W.; Diaz-Espejo, A.; Cuevas, M.V. The use of sap flow measurements for scheduling irrigation in olive, apple and Asian Pear trees and in grapevines. Plant Soil 2008, 305, 91–104. [Google Scholar] [CrossRef]
- Cammalleri, C.; Rallo, G.; Agnese, C.; Minacapilli, M.; Provenzano, G. Combined use of eddy covariance and sap flow techniques for partition of ET fluxes and water stress assessment in an irrigated olive orchard. Agric. Water Manag. 2013, 120, 89–97. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Precipitation | Air Temperature | ||||
|---|---|---|---|---|---|---|
| Total (mm) | % of Normal | Description | Average (°C) | Deviation from Normal (°C) | Description | |
| March | 17.7 | 65.56 | dry | 8.3 | +5.3 | warm |
| April | 2.8 | 3.11 | extremely dry | 13.2 | +2.5 | very warm |
| May | 29.8 | 53.21 | dry | 14.0 | +1.6 | warm |
| June | 9.2 | 15.08 | extremely dry | 23.9 | +5.3 | extremely warm |
| July | 50.7 | 99.41 | normal | 23.4 | +3.2 | extremely warm |
| August | 7.4 | 12.76 | extremely dry | 22.9 | +3.4 | extremely warm |
| September | 5.8 | 14.87 | extremely dry | 15.8 | +0.3 | normal |
| October | 17.7 | 55.31 | dry | 12.5 | +2.3 | warm |
| Budbreak Period | Flowering Period | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Treatment | T (°C) | w (mm) | T (°C) | w (mm) | |||||
| 0.20 | 0.40 | 0.60 | 0.20 | 0.40 | 0.60 | ||||
| Sap flow | Irrigated | 0.27 *** | 0.25 *** | 0.34 *** | 0.50 *** | 0.06 | 0.05 | 0.03 | 0.05 |
| Non-irrigated | 0.21 ** | 0.02 | 0.18 * | 0.14 | 0.89 | 0.01 | 0.001 | 0.004 | |
| D.C. | Irrigated | 0.35 *** | 0.01 | 0.42 | 0.58 *** | 0.42 *** | 0.47 *** | 0.43 *** | 0.40 *** |
| Non-irrigated | 0.28 *** | 0.13 ** | 0.21 *** | 0.72 *** | 0.38 *** | 0.47 *** | 0.42 *** | 0.39 *** | |
| Treatment | Budbreak Period | Flowering Period | |
|---|---|---|---|
| Dendrometric Changes | |||
| Sap flow | Irrigated | 0.24 ** | 0.17 ** |
| Non-irrigated | 0.16 * | 0.23 *** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bárek, V.; Kováčová, M.; Kišš, V.; Paulen, O. Water Regime Monitoring of the Royal Walnut (Juglans regia L.) Using Sap Flow and Dendrometric Measurements. Plants 2021, 10, 2354. https://doi.org/10.3390/plants10112354
Bárek V, Kováčová M, Kišš V, Paulen O. Water Regime Monitoring of the Royal Walnut (Juglans regia L.) Using Sap Flow and Dendrometric Measurements. Plants. 2021; 10(11):2354. https://doi.org/10.3390/plants10112354
Chicago/Turabian StyleBárek, Viliam, Martina Kováčová, Vladimír Kišš, and Oleg Paulen. 2021. "Water Regime Monitoring of the Royal Walnut (Juglans regia L.) Using Sap Flow and Dendrometric Measurements" Plants 10, no. 11: 2354. https://doi.org/10.3390/plants10112354
APA StyleBárek, V., Kováčová, M., Kišš, V., & Paulen, O. (2021). Water Regime Monitoring of the Royal Walnut (Juglans regia L.) Using Sap Flow and Dendrometric Measurements. Plants, 10(11), 2354. https://doi.org/10.3390/plants10112354