Abstract
An electronic nose (E-nose) system equipped with a gas sensor array and real-time control panel was developed for a fast diagnosis of whitefly infestation in tomato plants. Profile changes of volatile organic compounds (VOCs) released from tomato plants under different treatments (i.e., whitefly infestation, mechanical damage, and no treatment) were successfully determined by the developed E-nose system. A rapid sensor response with high sensitivity towards whitefly-infested tomato plants was observed in the E-nose system. Results of principal component analysis (PCA) and hierarchical clustering analysis (HCA) indicated that the E-nose system was able to provide accurate distinguishment between whitefly-infested plants and healthy plants, with the first three principal components (PCs) accounting for 87.4% of the classification. To reveal the mechanism of whitefly infestation in tomato plants, VOC profiles of whitefly-infested plants and mechanically damaged plants were investigated by using the E-nose system and GC-MS. VOCs of 2-nonanol, oxime-, methoxy-phenyl, and n-hexadecanoic acid were only detected in whitefly-infested plants, while compounds of dodecane and 4,6-dimethyl were only found in mechanically damaged plant samples. Those unique VOC profiles of different tomato plant groups could be considered as bio-markers for diagnosing different damages. Moreover, the E-nose system was demonstrated to have the capability to differentiate whitefly-infested plants and mechanically damaged plants. The relationship between sensor performance and VOC profiles confirmed that the developed E-nose system could be used as a fast and smart device to detect whitefly infestation in greenhouse cultivation.
1. Introduction
The U.S. is one of the world leaders in fresh tomato production, with around 2.7 billion pounds of fresh tomatoes produced in 2015 with a market value of about USD 1.22 billion [1]. However, tomatoes are subject to attack by a number of insect pests from the first emergence as seedlings until harvest. Among the biotic stresses from insects, damage caused by whitefly is a major constraint in the growth and productivity of tomatoes and other vegetable crops. Whiteflies, a family of Aleyrodidae, are small insects with a wingspan less than 3 mm. In warm or tropical climates, for example, in greenhouses, whitefly damage presents major problems in crop protection [2]. Nearly 25% of yield reduction of fresh tomatoes is caused by insects and virus damage [3,4]. Within the list of 27 arthropod pests causing yield reduction, whitefly is ranked as the most serious one, affecting 90% percent of the acreage grown [5] Whitefly, especially Trialeurodes vaporariorum and Bemisia tabaci, has been found worldwide in greenhouse crops and open-field vegetables over the past 20 years [6]. Direct crop damages occur when whiteflies feed on the foliage, which weakens the plants, reduces the yield, and even kills the seedlings. From a psychopathological perspective, the main concern is the plant viruses transmitted by whiteflies, causing serious reduction of crop production and total crop failure.
Many approaches for insect control have been developed in past decades [7,8,9]. Most methods use an integrated strategy for insect control employing a combination of chemical control, biological control, and genetic engineering. However, the extensive use of pesticide raises food safety concerns because of the potential of residues remaining on food as well as potentially serious environmental problems and can result in insecticide-resistant pests [10]. Genetic engineering may provide a solution to address the difficulty of resistant gene transfer, but, with respect to commercial utilization, it is still under development [11]. Therefore, a technology with low-cost, fast, and accurate diagnosis of insect infestation at early stages is advantageous to minimize the use of chemicals. It may further help to establish a smart platform for insect control and pest management.
Plants release a large amount of volatile organic compounds (VOCs) into their surroundings, which delivers functional information responding to their growth, defense, and health [12,13]. For example, the composition of VOCs keeps changing according to variable environments, such as pathogen infection, herbivore feeding, and mechanical damage [14]. In other words, VOCs emitted from plants indicate their real-time physiological health statuses, and, thus, they can be used as bio-information for disease diagnosis. Recent methods utilizing plant VOC profiles for insect control have received extensive interest [15,16]. Utilization of VOC fingerprints as unique information of plants has become an emerging field for food quality evaluation, insect management, and food safety, benefiting from promising advantages of non-invasiveness. Using GC-MS to analyze VOC profiles has become a widely used technique to identify individual VOCs from target objects [17,18]. However, such technology is expensive and time-consuming and requires expertise. Besides, GC-MS detection is a laboratory-based test, which will limit its applications in greenhouses and in open fields. Therefore, the development of a cost-efficient, fast, and accurate technology detecting VOC profiles is advantageous and a promising tool for insect control in greenhouses.
The electronic nose system (E-nose system) is designed to mimic the mammalian olfactory system. It has different types of sensor arrays and is coupled with pattern recognition algorithms that emulate the roles of the olfactory bulb and brain. In detail, the sensor array, which is the core of the system, is used to transform VOC information into electronic or digital signals using conditioning circuits to modulate signals and pattern recognition algorithms to classify aromas. It has been employed in diverse applications ranging from medical diagnosis to the food industry, environmental protection, and agriculture [19,20,21,22]. Given the fact that VOC profiles of plants play significant roles in responding to pest attacks, fungal infection, and other damages, the E-nose system is considered to have potential in detecting insect attacks and fungal infections. Specifically, the E-nose system has been employed in detecting insect damage and insect population in the host plant. In the case of the spider mite (Tetranychus urticae Koch), for instance, the E-nose system was used to investigate VOC profile changes of tomato plants with/without spider mite infestation [23,24]. The reliability of the E-nose system was also confirmed by detecting rice plants infested by striped stemborer (Chilo suppressalis) and brown planthopper (Nilaparvata lugens) [25]. Moreover, an early-stage diagnosis of aphid-infested tomato plants was performed by using a lab-developed E-nose system in our group [26].
However, the research on using smart E-nose systems for insect control in greenhouses and open fields is still in the early stage of its development. Moreover, the commercial E-nose system is expensive and not universal for all plants.
To address the above challenges, in this study a low-cost, non-invasive, portable E-nose system was to be designed and developed to accurately diagnose whitefly infestation in tomato plants. Specific objectives of this research were: (1) development of a portable E-nose system based on optimized sensor array; (2) determination of VOC biomarkers of tomato plants with and without infestation of whiteflies; (3) development of practical interface software for real-time recording and fast diagnosis; and (4) evaluation of the performance of the developed E-nose system under real-time applications. The long-term goal is to provide a reliable and practicable E-nose platform for fast and smart diagnosis for pest control in greenhouses and open fields.
2. Methods and Materials
2.1. Fabrication of E-Nose System
After optimization based on our previous work [26], a portable E-nose system (36 cm × 30 cm × 15 cm), equipped with a sensor array, a gas chamber, a data acquisition unit, signal conditioning module, temperature control system, and micro pumps, as illustrated in Figure 1, was developed in the Plant Sensing Lab (Department of Food, Agricultural, and Biological Engineering, Ohio State University, Wooster, OH) and in the Intelligent Sprayer Lab (USDA Agricultural Research Service (ARS), Wooster, OH). Specifically, the system included: (1) four gas sensors (MQ-138, MQ-135, MQ-3, and TGS 2602, the details of each sensor as listed in Table 1. As listed in Table 1, each sensor has a specific gas sensing spectrum, e.g., MQ-3(S1) is sensitive to alcohol family VOCs, while MQ-138 (S2) is sensitive to ketones and aromatic compounds; therefore, those sensor arrays result in a broad VOC spectrum, leading to a reliable and accurate detection); (2) a gas chamber (15 cm × 10 cm × 8 cm); (3) two micro pumps (Model NMS020L, KNF Neuberger Inc., Trenton, NJ, USA) for pumping air through the gas chamber; (4) and (5) two signal conditional modules; (6) a data acquisition unit (USB6200, National Instrumental Inc (NI), Austin, TX) designed to control the entire system; (7) a circulation fan (OD8025-12HSS, DC Brushless Fans, Hangzhou, China) used to keep the temperature inside the system consistent; and (8) a multiple power supply (RT-50 A, Mean Well USA, Inc. Fremont, CA, USA), employed to provide various voltages, such as 12 V DC, for the developed E-nose system. An interface software (as shown in Figure 2) based on the Labview platform (National Instrumental Inc., Austin, TX, USA) was designed to control the entire system, display sensor real-time performance, and automatically record the data for further analysis. The photos of the developed E-nose system are illustrated in Supplementary Materials (see Figure S1 in Supplementary Materials).
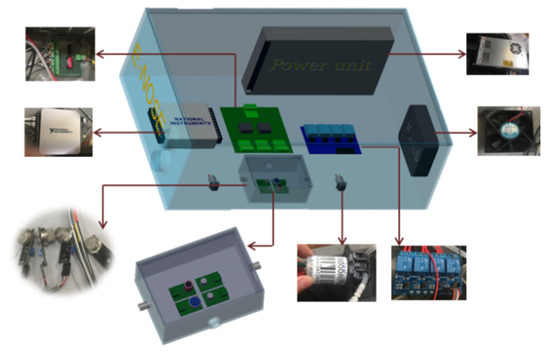
Figure 1.
Components of the developed E-nose system.

Table 1.
Gases detected by each sensor employed in E-nose system.
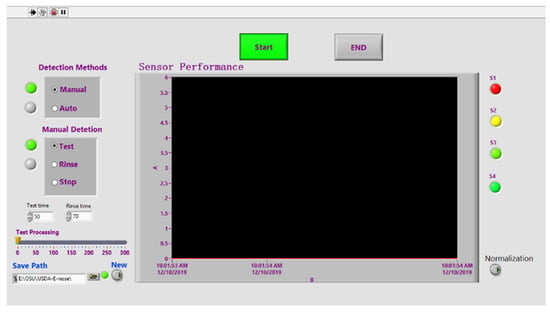
Figure 2.
Interface panel of the developed E-nose system.
2.2. Plant and Insects
Tomato seeds (Tomato, Burpee’s Big Boy Hybrid, Burpee Seed Company, Warminster, PA, USA) were sown in 10 cm pots for 2–3 weeks under indoor conditions (1/4 inch deep in pot, 75 °F). As soon as seedlings grew to 2–3 inches, they were transplanted to 1 L pots with rich soil mixture. Plants were watered daily, fertilized weekly, and grown in a greenhouse under controlled conditions (light 15 h per day, temperature at 80.6 °F). Plants at 5 weeks of age were used for experiments and divided into three groups (15 plants for each group). Virus-free greenhouse whiteflies (Trialeuodes vaporariorum) were obtained from a laboratory colony raised in the Controlled Environments Laboratory (Department of Entomology, Ohio Agricultural and Development Center, Wooster, OH, USA). The infestation by whitefly of the tomato plants (100 adult whiteflies for one plant) was performed in a cage within 4 h in the morning. Infested tomato plants inside a cage, healthy plants (controls), and the plants designed for mechanical damage were cultured in the same greenhouse under the same conditions. As for mechanical damage, acupuncture treatment (10 times for one leaf, total 5 leaves on one plant) was performed on tomato plants.
2.3. Experimental Design
The developed E-nose differed from previous studies in that the optimized system could be used for open-field tests [26]. As illustrated in Figure 3, pump 2 discharged the headspace air of a plant sample, which was covered in a Teflon bag for around an hour, to the developed E-nose system, and pump 1 helped to pull out the tested air flow and draw back to the sample plant. This system design provided a stable closed loop, thereby partially reducing interferences of temperature and humidity from the external environment. The parameters of temperature and humidity were not collected during E-nose detection.
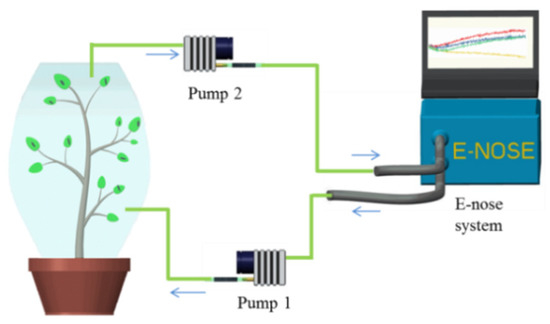
Figure 3.
Illustration of tomato plant detection using the E-nose system.
Tomato plants were cultivated under the three different treatments of whitefly infestation, mechanical damage, and no stress (control group). Before testing, selected plant samples were covered with Teflon bags for about an hour to generate headspace air. Then, the air was pumped into the E-nose system for 10 min. Nitrogen was used to rinse the E-nose system for 5 min before the next sample testing. For each treatment (15 plants for each treatment group), ten tomato plants of similar size were selected and tested as replicates.
2.4. GC-MS Tests
For the GC-MS test, the headspace air of tomato plants was collected using solid-phase microextraction (SPME) from the same plants used for E-nose tests. Three replicates were randomly selected from the tree plant groups. The collected VOCs were desorbed, separated through a column (ZB-5HT, 30 mm × 0.25 mm × 0.25 mm, Zebron, Phenomenex, CA, USA), and then identified by MS in the GC-MS system (QP 2010 SE, Shmadzu, Kyoto, Japan). The GC-MS operational parameters were as follows: splitless injection temperature for 3 min at 200 °C; column temperature programmed for 50 °C for 1 min to 120 °C at 5 °C per min and then to 240 °C at 8 °C per min for 5 min; ion source set at 230 °C, 70 eV, with a mass ranging from 40 to 350 amu. Compounds were identified by comparing the recorded mass spectra with the National Institute of Standards and Technology mass spectral library, retention index (RI), and previous literature and published index data (http://www.flavornet.org, accessed on 17 October 2021). The retention indices were calculated for all of the VOCs using a homologous series of n-alkanes (C8 to C20) (Sigma-Aldrich, Inc. St. Louis, MO, USA).
2.5. Data Analysis
Sensor signal voltages were normalized following the equation below before further qualitative and quantitative analysis.
where v is the normalized sensor signal voltage, is the sensor voltage at the ith second, and is the initial voltage at 0 s.
Principle component analysis (PCA) and hierarchical clustering analysis (HCA) were employed for qualitative classification. All of the data analysis was performed by MATLAB (Mathworks Inc., Natick, MA, USA).
3. Results and Discussion
3.1. Sensor Response
The developed E-nose system provided a fast, non-invasive, and real-time diagnosis of whitefly infestation of tomato plants at early stages. The sensor performances of the E-nose system towards different tomato plants were represented in Figure 4. As shown, the sensor array exhibited different performance patterns between healthy tomato plants (Figure 4a) and plants infested by whiteflies (Figure 4b). Specifically, the sensor performance of whitefly-infested plants showed a pattern of increasing and higher voltages of each sensor compared with that of healthy plants. For healthy plants, the sensor voltage increased within the first 450 s and tended stable afterwards, while for whitefly-infested plants the sensor response increased more quickly and kept increasing after 450 s. That might be because the infested plants released VOCs at higher concentrations in order to defend against attacks from insects. Moreover, it is worth noticing that sensor 3 (S3), which was sensitive to the alcohol class of VOCs, showed a significant and continuous increase when the E-nose system analyzed the infested plants, while it showed little response in the healthy plants. This suggested that some alcohol VOCs were newly generated after whitefly damage to tomato plants.
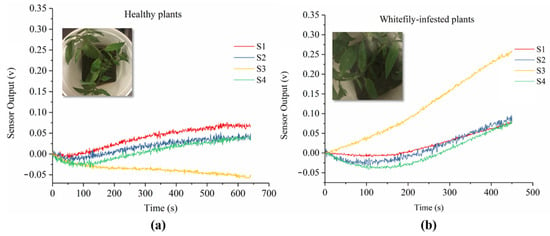
Figure 4.
Sensor performance of E-nose system towards healthy (a) and whitefly-infested (b) tomato plants.
3.2. E-Nose Performance
Based on the reliable sensor response towards different tomato plants, two pattern recognition algorithms, PCA and HCA, were employed for further diagnosis. Results in Figure 5 illustrate that the infested plants and healthy plants were clearly different in the PCA plot (Figure 5a), with the first three PCs accounting for 87.4% of classification. Also, the two clusters representing healthy and infested plants were clearly observed in the HCA plot (Figure 5b). Specifically, the loading plot of Figure 5c indicates that sensor S3 made the most significant contribution to differentiation between infested plants and healthy plants, followed by sensor S1, while sensors S2 and S4 made relatively less of a contribution to classification. Those results suggested that VOC profiles of healthy and infested plants were different and the E-nose system had a promising potential to diagnose the difference. In order to investigate the compounds that corresponded to the VOC profile changes and led to different sensor responses, a deeper understanding and identification of the structure and concentration of each VOC was necessary.
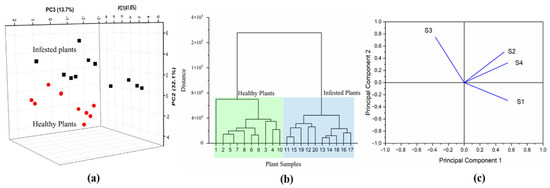
Figure 5.
PCA result (a) and HCA result (b) of the developed E-nose system detecting healthy and infested tomato plants; PCA loading plot (c).
3.3. GC-MS Results
Two typical GC-MS spectra of VOC profiles of a healthy and a whitefly infested tomato plant are shown in Figure 6 and Figure 7, respectively. Chemical structures of some crucial volatile compounds, of which the relative concentration was more than 1%, were marked in the spectrum figures. Comparing the spectrum of whitefly-infested plant (Figure 7) with that of a healthy one (Figure 6), it is clear that the infested plant had a richer VOC profile than the healthy one, and two new compounds were observed in the infested tomato plants. Those new compounds were 2-nonanol and oxime-, methoxy-phenyl. Compound 2-nonanol, with an odor of cucumber, is a simple alcohol, and it has been reported to have bio-activity [27]. Cheng et al. reported that 2-nonanol has the function of defending against plant-parasitic nematodes as a bio-insecticide, possessing nematicidal activity and fumigant activity [27]. In terms of whitefly infestation, 2-nonanol might have a similar activity to antagonize the continuous infestation from whiteflies. Oxime-, methoxy-phenyl has been identified as one of the eight different components for insect resistance, such as aphid resistance [28]. Moreover, the concentration of oxime-, methoxy-phenyl was the second highest of the compounds identified from the whitefly infested tomato plants.
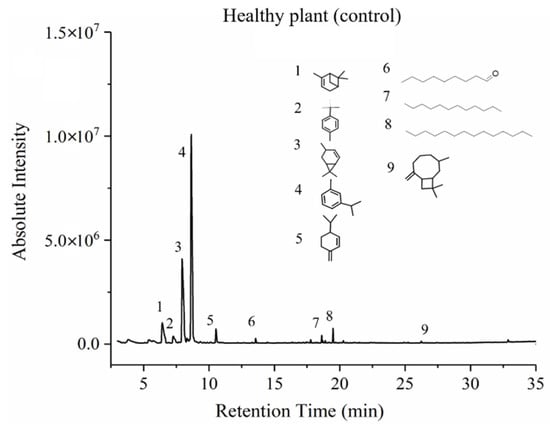
Figure 6.
A typical GC-MS spectrum of VOCs from healthy tomato plant.
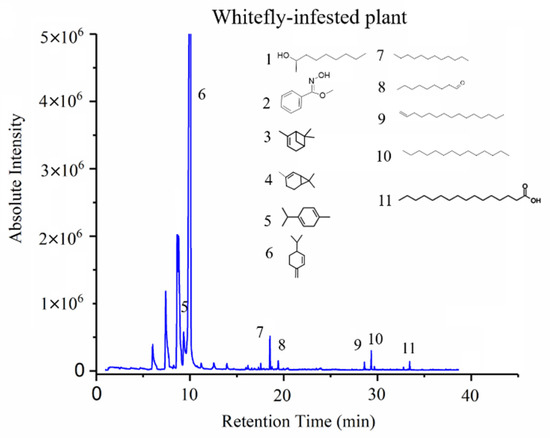
Figure 7.
A typical GC-MS spectrum of VOCs from a whitefly-infested tomato plant.
Table 2 lists the VOCs with averaged concentration in healthy plants, infested plants, and mechanically damaged tomato plants. Besides the aforementioned two new compounds, n-hexadecanoic acid was also found in the infested plants. In comparison with healthy plants, the concentrations of 4-carene (from 28.05% to 14.66%) and beta-phellandrene (from 53.7% to 30.65%) significantly decreased after infestation by whiteflies, while the new compound oxime-, methoxy-phenyl had a significantly high concentration (17.67%). One of the probable reasons was that some of the VOCs of healthy plants might be generated from a different synthesis path and be configured as new compounds with the ability to combat the infestation and to deliver messages for communication between plants. All those findings indicated that VOC profiles of tomato plants changed after infestation. Such variations provided fundamentals for E-nose system development.
Table 2.
Typical VOCs released from whitefly-infested tomato plant, healthy plant, and mechanically damaged plant.
3.4. Sensor Performance between Mechanical Damage and Whitefly Infestation
Whiteflies are a highly polyphagous insect pest that cause considerable damage to crops by direct phloem feeding and by indirect biotrophic infection resulting from whitefly transmission of plant viruses. Moreover, phloem-feeders cause mechanical damage to plants, along with saliva components and endosymbiotic-borne stimulation, and they are believed to contribute to the induction of plant defense responses against whiteflies. As studies have demonstrated, plant VOCs play a role in the direct defense against herbivores, pathogens, and other damages, and the VOC profiles in the response by different species of plants subjected to natural or imposed herbivory, viral infection, and mechanical damages might share some common constituents [29], for instance, the well-known compounds such as terpenoids, phenylpropanoids, fatty acids, and amino acid derivatives [30]. Various studies have qualitatively identified the difference between herbivory-induced VOCs and mechanical injury-induced VOCs [31]. However, only a few studies have investigated quantitative differences between plant VOCs triggered by insect infestation and mechanical damages. In the present study, the developed E-nose system was targeted to distinguish this difference, with the purpose of exploring a new technique for insect control in greenhouses.
As indicated in Figure 8, sensor performance towards whitefly-infested plants and mechanically damaged plants were apparently different (Figure 8a,b), demonstrating that the developed E-nose system was able to differentiate the variation of VOCs emitted from whitefly-infested plants and mechanically damaged plants. In comparison, the sensor S1 response to mechanically damaged plants had a higher amplitude than that in whitefly-infested plants, while for three other sensors (S2, S3, and S4), it was noticeable that the response to whitefly-infested plants continued to increase (Figure 8a) but those of mechanically damaged plants increased first and then decreased to a stable level. Several reasons might explain this phenomenon. Whitefly is a phloem-feeder, causing less damage to plants compared with other feeding guilds and the mechanical damages by physical acupuncture in this study. Thus, S1 displayed a higher sensor amplitude when exposed to mechanically damaged plants. On the other hand, previous research found that whitefly infestation in tomato plants induced a direct and rapid resistance to pathogen or bacterial infection at the beginning stage of plant defense by releasing corresponding VOCs [32], while mechanical damages to plants may not trigger that biological defense [33]. Third, VOC emission in whitefly-infested plants is a dynamic process that involves a time-dependent variation in the VOC composition [34]. Thereby we can explain the continuous increase in the four sensors’ responses.
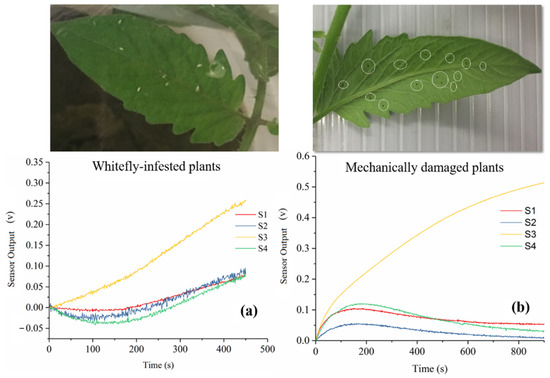
Figure 8.
Sensor performance of E-nose system towards whitefly-infested plant (a) and mechanically damaged plant (b).
The PCA plot in Figure 9 showed the results of the E-nose system detecting the difference between the three plant groups (i.e., mechanically damaged group, whitefly-infested group, and control group). As shown, the mechanically damaged group and the whitefly-infested group were separated but with slight overlap, while those two groups were clearly separated from the control group. This indicated that the VOCs released from the mechanically damaged group and the whitefly-infested group were different but had some compounds in common, but the VOC profiles of the damaged groups were significantly different from those of the healthy group. Given the above findings, it could be demonstrated that the developed E-nose system is able to differentiate those two damages in tomato plants. From the perspective of plant metabolic pathways, one of the key VOCs emitted after insect damage is indole, which belongs to the alcohol class and is bio-synthesized via the tryptophan pathway [35], while damage-induced VOCs are synthesized upon cell disruption and possess the ability to trigger a systemic response and herbivore resistance [36]. Most likely, after mechanical damage or wounding, two metabolic synthesis phases may be performed [37]. In the first stage, VOC synthesis is initiated within seconds of tissue damage by delivering enzymes, such as cut green leaves (GLVs). In the second stage, synthesis is induced, within hours of damage perception, of compounds such as phenolic compounds and terpenes [37]. This helps to explain why the sensors’ performance pattern for damaged leaves (Figure 8b) increased within the first 200 s and then decreased over 800 s.
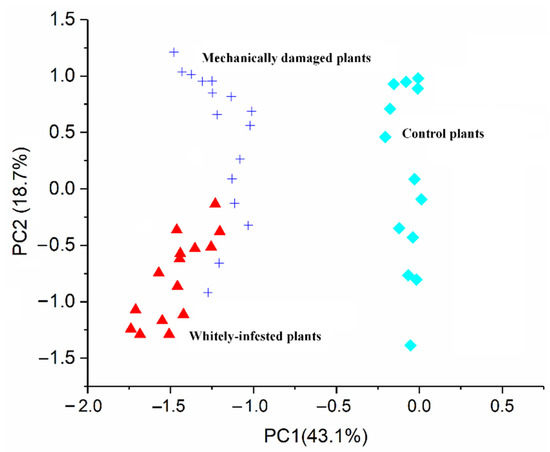
Figure 9.
PCA result of the three plant groups detected by E-nose system.
3.5. Relationship between VOCs Composition and Sensor Performance
As listed in Table 2, the VOC profiles of tomato plants significantly changed in response to whitefly infestation and mechanical damage. Overall, in comparison with control plants, VOCs detected in whitefly-infested plants and mechanically damaged plants showed different emission patterns. For VOCs released from insect attacked plants: (1) around 38.9% were not detected in controls but were detected after whitefly infestation; and (2) 27.8% showed obvious variation during infestation, while another 27.8% showed no significant variation. For VOCs emitted from mechanically damaged plants: (1) 22.2% were only detected in acupuncture treated plants; and (2) 44.4% of VOCs showed no significant variation compared with VOCs from controls, while 22% displayed a large increase or decrease.
Specifically for VOCs emitted from whitefly-infested plants and mechanically damaged plants, as illustrated in Figure 10, it was found that: they shared 55.6% of VOCs; 33.3% of VOCs were only detected in whitefly-damaged plants; and 12% were only detected in mechanically damaged ones. Among the common VOC elements, 23% of VOCs exhibited a large variation between these two types of attacked tomato plants. Clearly, according to those results, it is reasonable that sensors showed similar tendencies but different performances when directed towards whitefly-infested and mechanically damaged plants. More importantly, the VOCs of 2-nonanol, oxime-, methoxy-phenyl, and n-hexadecanoic acid, which were only found in the VOC profiles of infested plants, and the VOCs dodecane and 4,6-dimethyl, which were only observed in mechanically damaged plants, might function as VOC bio-markers to distinguish their differences by using the developed E-nose system. Moreover, it is notable that 2-nonanol and n-hexadecanoic acid, which were only detected in whitefly-infested plants, have been reported to function as an anti-parasitic against nematodes and in resistance to fungi Thus, it is suggested that tomato plants’ response to whitefly attack might trigger resistance to pathogens or fungi, which has been demonstrated in other reports [38,39]. In conclusion, the VOCs emission in whitefly-infested plants and mechanically damaged plants is a concomitant process, and the corresponding VOC profiles are different, providing fundamental support for the feasibility of differentiation by sensors. In addition, it demonstrated the ability of the developed E-nose system to distinguish whitefly-infested plants from controls and mechanically damaged ones.
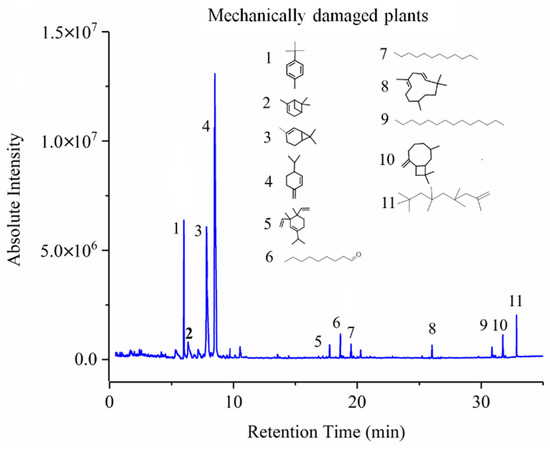
Figure 10.
GC-MS spectrum of mechanically damaged plant.
Given the obtained results, it could be concluded that the developed E-nose system is able to distinguish infested plants from healthy plants, providing a fast and acceptable classification. However, there are some limitations for potential applications in open fields. Different from greenhouses, the collection of crop VOCs in an open field is challenging, due to unstable environmental surroundings, low VOC concentrations, and multiple variables. For example, there might be more than one insect infestation or damage in one plant, and infestation from insects probably cause different viral infections, leading to unreliable VOC fingerprints. Therefore, subsequent research will focus on the development and optimization of the E-nose system for open-field tests.
4. Conclusions
A portable E-nose system was developed for a fast and smart diagnosis of whitefly infestation in tomato plants in greenhouses. On the basis of the PCA results (the first three PCs accounting for 87.4% of the classification) and the HCA results, the developed E-nose system demonstrated its great potential as a smart device for successful diagnosis of whitefly infestation. The GC-MS results indicated that whitefly-stressed tomato plants released some new VOCs and confirmed that terpene compounds might be used to combat whitefly infestation. Moreover, the difference in VOC profiles between whitefly-stressed plants and mechanically damaged plants indicated that the damage caused by whitefly could trigger resistance to fungal and bacterial infection during the response to the attack. The variation in VOC fingerprints between plants under different treatments helped to reveal the principles of sensor responses when directed towards different samples, thus providing a baseline for further optimization. Results from GC-MS analysis and E-nose performance demonstrated its promising ability as a non-invasive and portable tool for insect control in tomato plants in greenhouses. Further investigations will be needed to validate the developed E-nose system’s reliability and repeatability in different plants and in open fields.
Supplementary Materials
The following are will be available online at https://www.mdpi.com/article/10.3390/chemosensors9110297/s1, Figure S1: Photos of developed E-nose system: sampling chamber (a), sensory chamber (b), a photo of E-nose system (c), and control panel (d).
Author Contributions
S.C.: methodology, validation, investigation, writing—draft, writing—review and editing. L.C.: software and programming. N.A.: methodology. H.Z. and P.P.L.: funding acquisition, supervision, project administration, writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by USDA NIFA SCRI (Grant No. 2015-51181-24253).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No data reported.
Acknowledgments
The authors would like to thank Mary Wicks for reviewing the manuscript.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Guan, Z.F.; Biswas, T.; Wu, F. The US Tomato Industry: An Overview of Production and Trade. 2018. Available online: https://edis.ifas.ufl.edu/fe1027 (accessed on 17 October 2021).
- Xu, H.X.; Qian, L.X.; Wang, X.W.; Shao, R.X.; Hong, Y.; Liu, S.S.; Wang, X.W. A salivary effector enables whitefly to feed on host plants by eliciting salicylic acid-signaling pathway. Proc. Natl. Acad. Sci. USA 2019, 116, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Moodley, V.; Gubba, A.; Mafongoya, P.L. A survey of whitefly-transmitted viruses on tomato crops in South Africa. J. Crop Prot. 2019, 123, 21–29. [Google Scholar] [CrossRef]
- Nault, B.A.; Speese, J. Major insect pests and economics of fresh market tomato in eastern Virginia. J. Crop Prot. 2002, 21, 359–366. [Google Scholar] [CrossRef]
- Pico, B.; Diez, M.J.; Nuez, F. Viral diseases causing the greatest economic losses to the tomato crop. II. The tomato yellow leaf curl virus—A review. Sci. Hortic. 1996, 67, 151–196. [Google Scholar] [CrossRef]
- Jesus, N.C.; Elvira, F.O.; Sonia, S.C. Emerging Virus Disease Transmitted by whiteflies. Annu. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar]
- Gantz, V.M.; Akbari, O.S. Gene editing technologies and applications for insects. Curr. Opin. Insect Sci. 2018, 28, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Kaur, A. Control of insect pests in crop plants and stored food grains using plant saponins: A review. LWT 2018, 87, 93–101. [Google Scholar] [CrossRef]
- Lombardo, L.; Coppola, G.; Zelasco, S. New technologies for insect-resistant and herbicide tolerant plants. Trends Biotechnol. 2016, 34, 49–57. [Google Scholar] [CrossRef]
- Bhatia, V.; Uniyal, P.L.; Bhattacharya, R. Aphid resistance in Brassica crops: Challenges, biotechnological progress and emerging possibilities. Biotechnol. Adv. 2011, 29, 879–888. [Google Scholar] [CrossRef]
- Kumar, S.; Atri, C.; Sangha, M.K.; Banga, S.S. Screening of wild crucifers for resistance to mustard aphid, Lipaphis erysimi (Kaltenbach) and attempt at introgression of resistance genes(s) from Brassica fruticalosa to Brassica juncea. Euphytica 2011, 3, 461–470. [Google Scholar] [CrossRef]
- Matsui, K. A portion of plant airborne communication is endorsed by uptake and metabolism of volatile organic compounds. Curr. Opin. Plant Biol. 2016, 32, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Oilkawa, P.Y.; Lerdau, M.T. Catabolism of volatile organic compounds influences plant survival. Trends Plant Sci. 2013, 18, 695–703. [Google Scholar] [CrossRef]
- Maffei, M.E. Sites of synthesis, biochemistry and functional role of plant volatiles. S. Afr. J. Bot. 2010, 76, 612–631. [Google Scholar] [CrossRef]
- Noman, A.; Aqeel, M.; Qasim, M.; Haider, I.; Lou, Y. Plant-insect-microbe interaction: A love triangel between enemies in ecosystem. Sci. Total Environ. 2020, 699, 134181. [Google Scholar] [CrossRef]
- Maffei, M.E.; Mithofer, A.; Boland, W. Insects feeds on plants: Rapid signals and response preceding the induction of phytochemucal release. Phytochemistry 2007, 68, 2946–2959. [Google Scholar] [CrossRef]
- Chang, K.P.P.; Zakaria, A.; Nasir, A.S.A.; Yusuf, N.; Thriumani, R.; Shakaff, A.Y.M.; Adom, A.H. Analysis and Feasibility Study of Plant Disease using E-Nose. In Proceedings of the 2014 IEEE International Conference on Control System, Computing and Engineering (ICCSCE 2014), Penang, Malaysia, 28–30 November 2014. [Google Scholar]
- Lan, Y.B.; Zheng, X.Z.; Westbrook, J.K.; Lopez, J.; Lacey, R.; Hoffmann, W.C. Identification of stink bugs using an electronic nose. J. Bionic Eng. 2008, 5, 72–180. [Google Scholar] [CrossRef]
- Khatoon, Z.; Fouad, H.; Alothman, O.Y.; Hashem, M.; Ansari, Z.A.; Ansari, S.A. Doped SnO2 Nanomaterials for E-nose based electrochemical sensing of biomarkers of lung cancer. ACS Omega 2020, 5, 27645–27654. [Google Scholar] [CrossRef]
- Cui, S.Q.; Yang, L.C.; Wang, J.; Wang, X.L. Fabrication of a sensitive gas sensor based on PPy/TiO2 nanocomposites films by layer-by-layer self-assembly and its application in food storage. Sens. Actuators B. Chem. 2016, 233, 337–346. [Google Scholar] [CrossRef]
- Tastan, M.; Gokozan, H. Real-time monitoring of indoor air qualifty with internet of things-based E-nose. Appl. Sci. 2019, 9, 3435. [Google Scholar] [CrossRef]
- Cui, S.Q.; Ling, P.; Zhu, H.P.; Keener, H.M. Plant pest detection using an artificial nose system: A review. Sensors 2018, 18, 378. [Google Scholar] [CrossRef]
- Cellini, A.; Blasioli, S.; Biondi, E.; Bertaccini, A.; Braschi, I.; Spinelli, F. Potential applications and limitations of electronic nose devices for plant disease diagnosis. Sensors 2017, 17, 2596. [Google Scholar] [CrossRef]
- Okur, S.; Sarheed, M.; Hubber, R.; Zhang, Z.; Heinke, L.; Kanbar, A.; Woll, C.; Nick, P.; Lemmer, U. Identification of mint scents using a QCM based E-nose. Chemosensors 2021, 9, 31. [Google Scholar] [CrossRef]
- Cheng, S.-M.; Wang, J.; Wang, Y.-W.; Wei, Z.-B. Discrimination of Different Types Damage of Tomato Seedling by Electronic Nose. In Proceedings of the ITM Web of Conferences, Wuhan, China, 24–26 March 2017. [Google Scholar]
- Cui, S.Q.; Inocnete, E.A.A.; Acosta, N.; Keener, H.M.; Zhu, H.P.; Ling, P.P. Development of E-nose system for fast diagnosis of aphids-stressed tomato plants at early stage. Sensors 2019, 19, 3480. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Yang, J.; Nie, Q.; Huang, D.; Yu, C.; Zheng, L.; Cai, M.; Thomashow, L.S.; Weller, D.M.; Yu, Z.; et al. Volatile organic compounds from paehibacillus polymyxa KM2501-1 control meloidogyne incognita by multiple strategies. Sci. Rep. 2017, 7, 16213–16225. [Google Scholar] [CrossRef]
- Wang, R.; Shen, X.; Wang, C.; Ge, R.; Zhang, Z.; Guo, X. Analysis of leaf volatiles of crabapple (Malus sp.) individuals in different aphids’ resistance. Am. J. Plant Sci. 2014, 5, 3295–3301. [Google Scholar] [CrossRef]
- McCormick, A.C.; Unsicker, S.B.; Gershenzon, J. The specificity of herbivore-induced plant volatiles in attracting herbivore enemies. Trends Plant Sci. 2012, 17, 303–310. [Google Scholar] [CrossRef]
- Dudareva, N.; Negre, F.; Nagegowda, D.A.; Orlova, I. Plant volatiles: Recent advances and future perspectives. Crit. Rev. Plant Sci. 2006, 25, 417–440. [Google Scholar] [CrossRef]
- Piesik, D.; Łyszczarz, A.; Tabaka, P.; Lamparski, R.; Bocianowski, J.; Delaney, K. Volatile induction of three cereals: Influence of mechanical injury and insect herbivory on injured plants and neighbouring uninjured plants. Ann. Appl. Biol. 2010, 157, 425–434. [Google Scholar] [CrossRef]
- Kant, M.R.; Ament, K.; Sabelis, M.W.; Haring, M.A.; Schuurink, R.C. Differential timing of spider mite induced direct and indirect defenses in tomato plants. Plant Physiol. 2004, 135, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Rodiguez-saona, C.; Crafts-brandner, S.J.; Canas, L.A. Volatile emissions triggered by multiple herbivore damage: Beet armyworm and whitefly feeding on cotton plants. J. Chem. Ecol. 2003, 29, 2539–2550. [Google Scholar] [CrossRef] [PubMed]
- Ángeles López, Y.I.; Martínez-Gallardo, N.A.; Ramírez-Romero, R.; López, M.G.; Sánchez-Hernández, C.; Délano-Frier, J.P. Cross-kingdom effects of plant-plant signaling via volatile organic compounds emitted by tomato (Solanum lycopersicum) plants infested by the greenhouse whitefly (Trialeurodes vaporariorum). J. Chem. Ecol. 2012, 38, 1376–1386. [Google Scholar] [CrossRef] [PubMed]
- Frey, M.; Stettner, C.; Paré, P.W.; Schmelz, E.A.; Tumlinson, J.H.; Gierl, A. An herbivore elicitor activates the gene for indole emission in maize. Proc. Natl. Acad. Sci. USA 2000, 97, 14801–14806. [Google Scholar] [CrossRef]
- Quintana-Rodriguez, E.; Duran-Flores, D.; Heil, M.; Camacho-Coronel, X. Damage-associated molecular patterns (DAMPs) as future plant vaccines that protect crops from pests. Sci. Hortic. 2018, 237, 207–220. [Google Scholar] [CrossRef]
- Meents, A.K.; Mithofer, A. Plant-plant communication: Is there a role for volatile damage-associated molecular patterns? Front. Plant Sci. 2020, 11, 1538–1550. [Google Scholar] [CrossRef]
- Liu, P.P.; Von Dahl, C.C.; Park, S.W.; Klessig, D.F. Interconnection between methyl salicylate and lipid-based longdistance signaling during the development of systemic acquired resistance in Arabidopsis and tobacco. Plant Physiol. 2011, 155, 1762–1768. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yi, H.S.; Heil, M.; Adame-alvarez, R.M.; Ballhorn, D.; Ryu, C.M. Airborne induction and priming of plant resistance to a bacterial pathogen. Plant Physiol. 2009, 151, 2152–2161. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).