
Identification of Sorghum (Sorghum bicolor (L.) Moench) Genotypes with Potential for Hydric and Heat Stress Tolerance in Northeastern Mexico

 , and
, and 
Abstract
:1. Introduction
2. Results
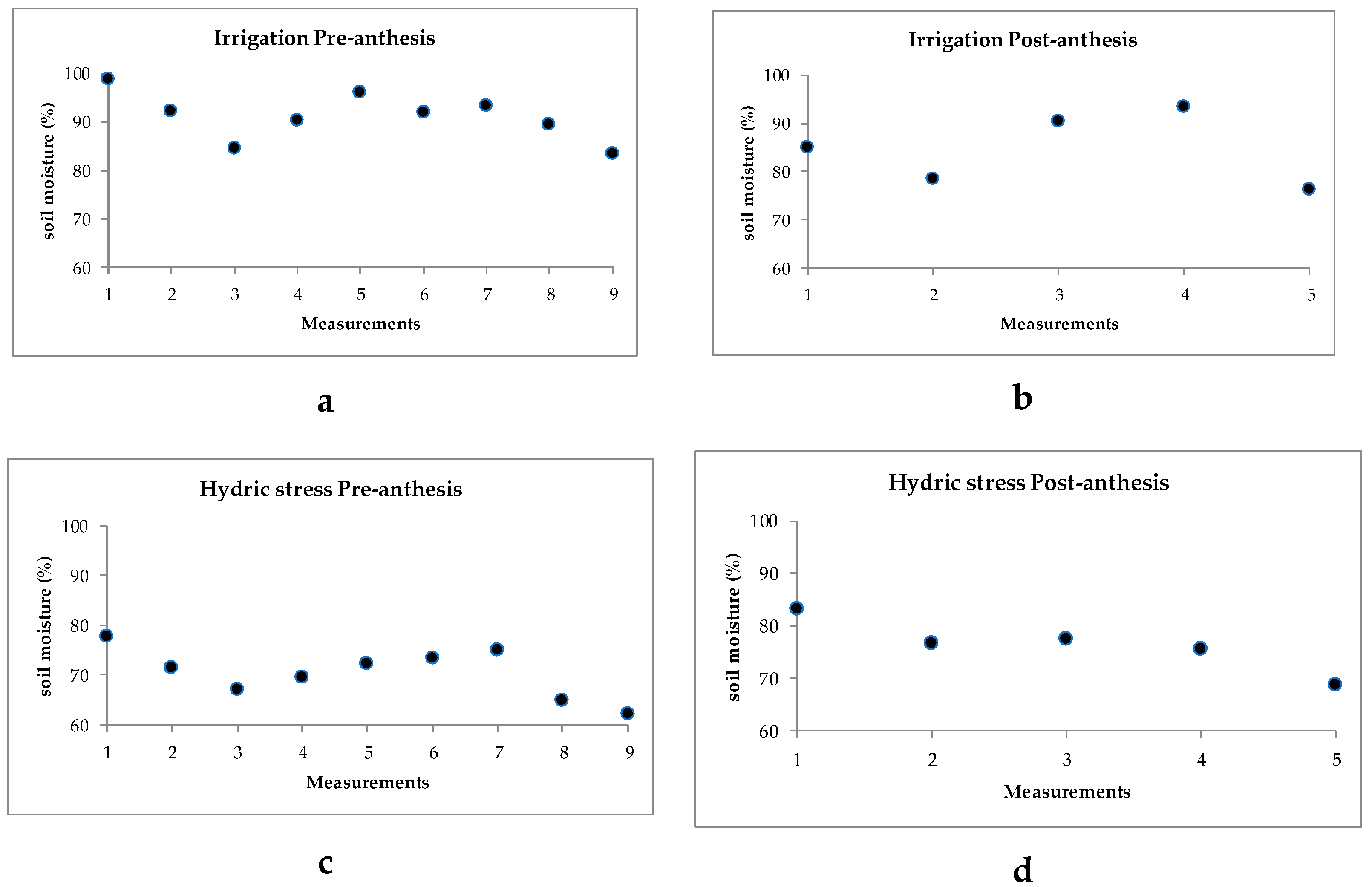
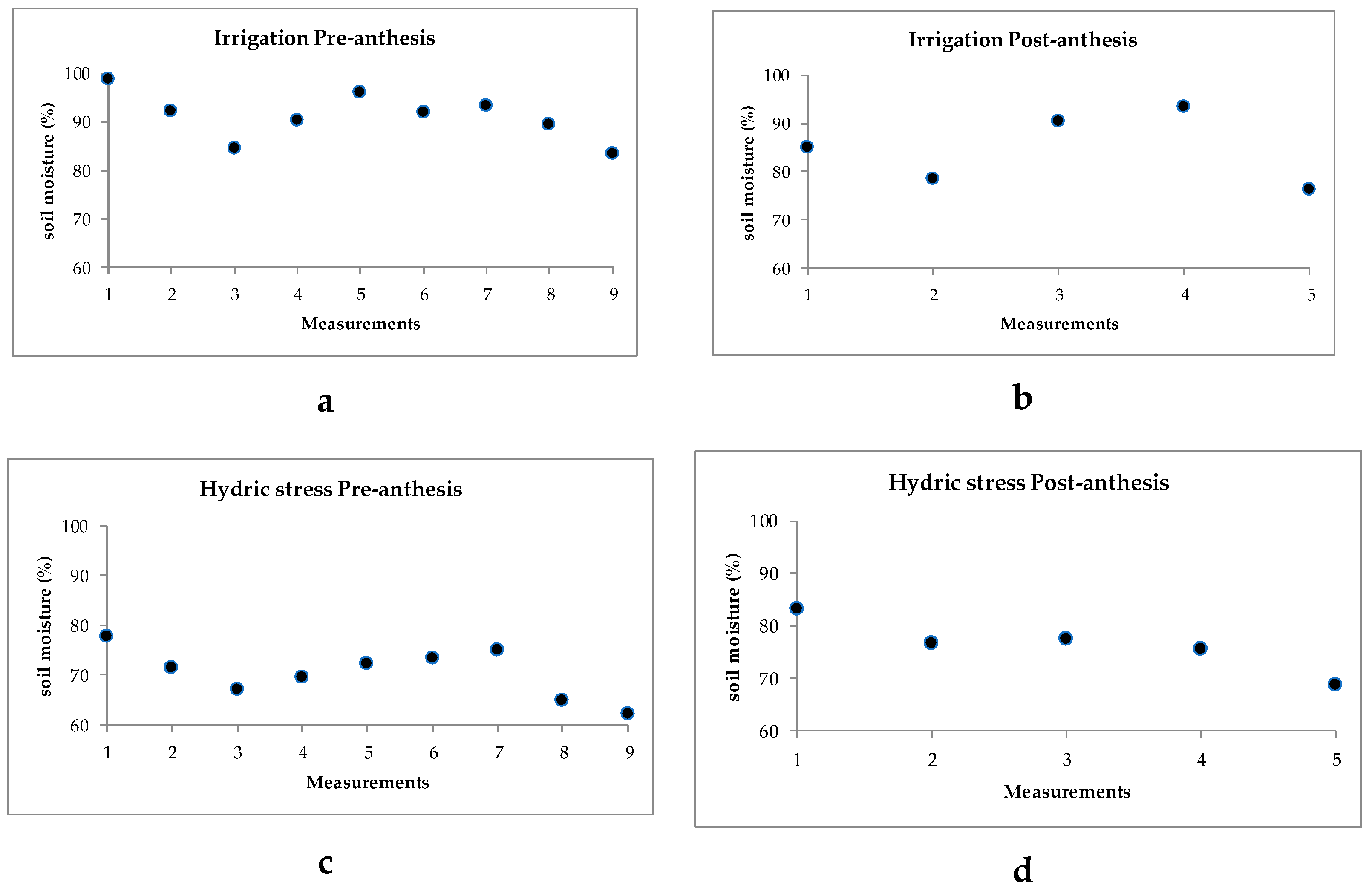
2.1. Ambient Temperatures and Soil Humidity
2.2. Physiological Variables
2.2.1. Maximum Quantum Yield of PSII (Fv/Fm) under Hydric Stress
2.2.2. Photosynthetic Rate (PN), Stomatal Conductance (gs), and Transpiration (E)
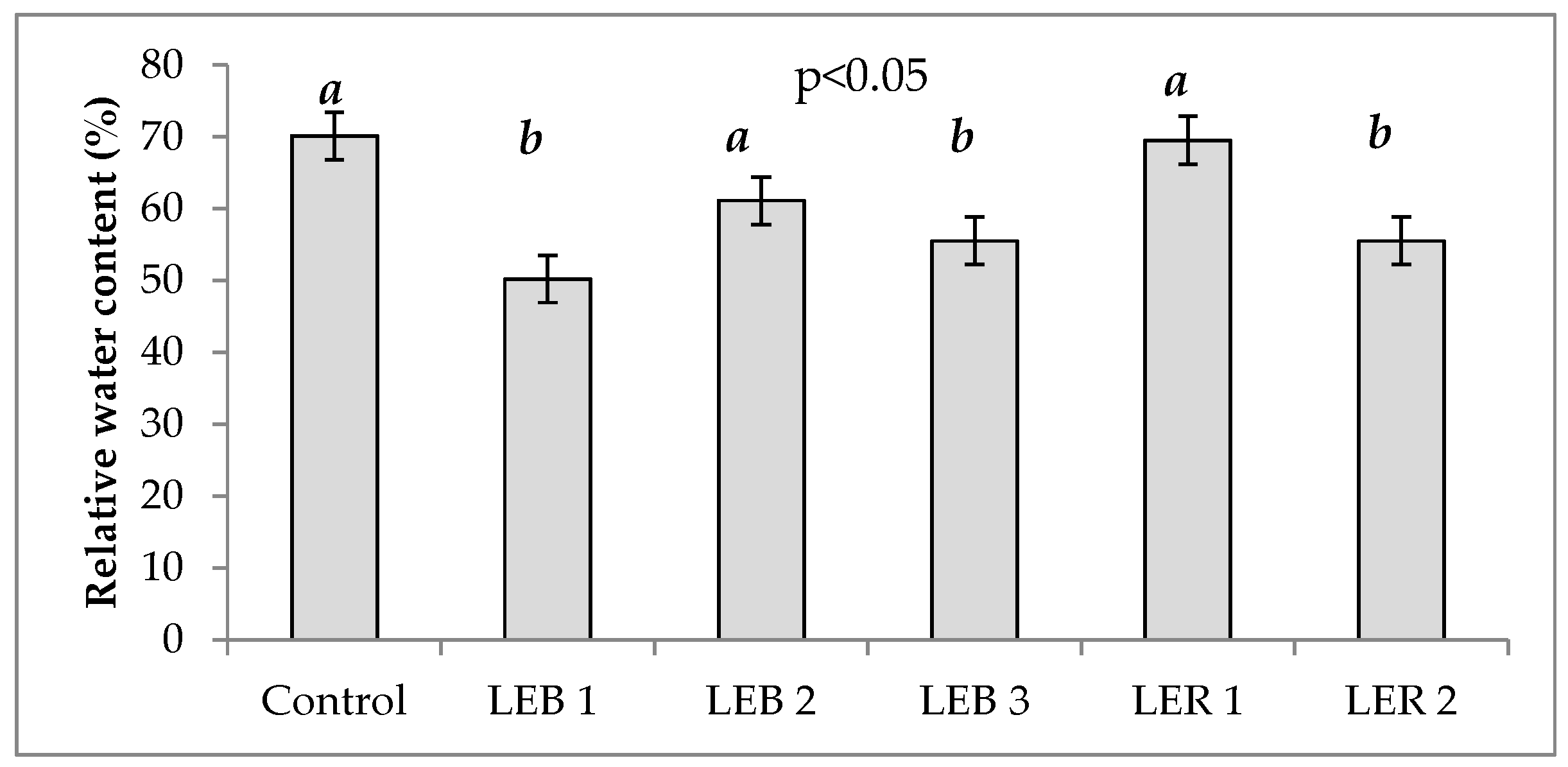
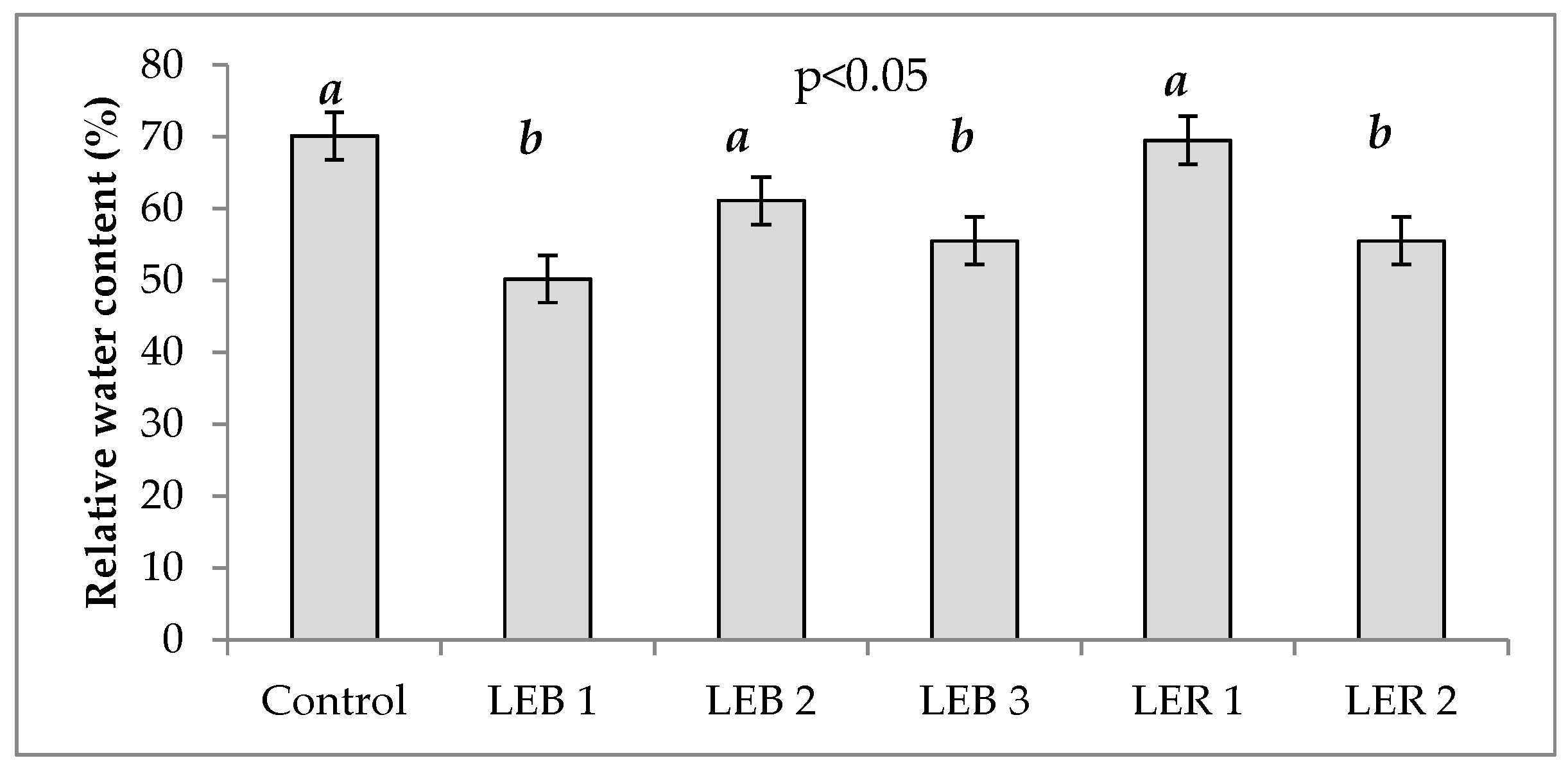
2.2.3. Relative Water Content (RWC)
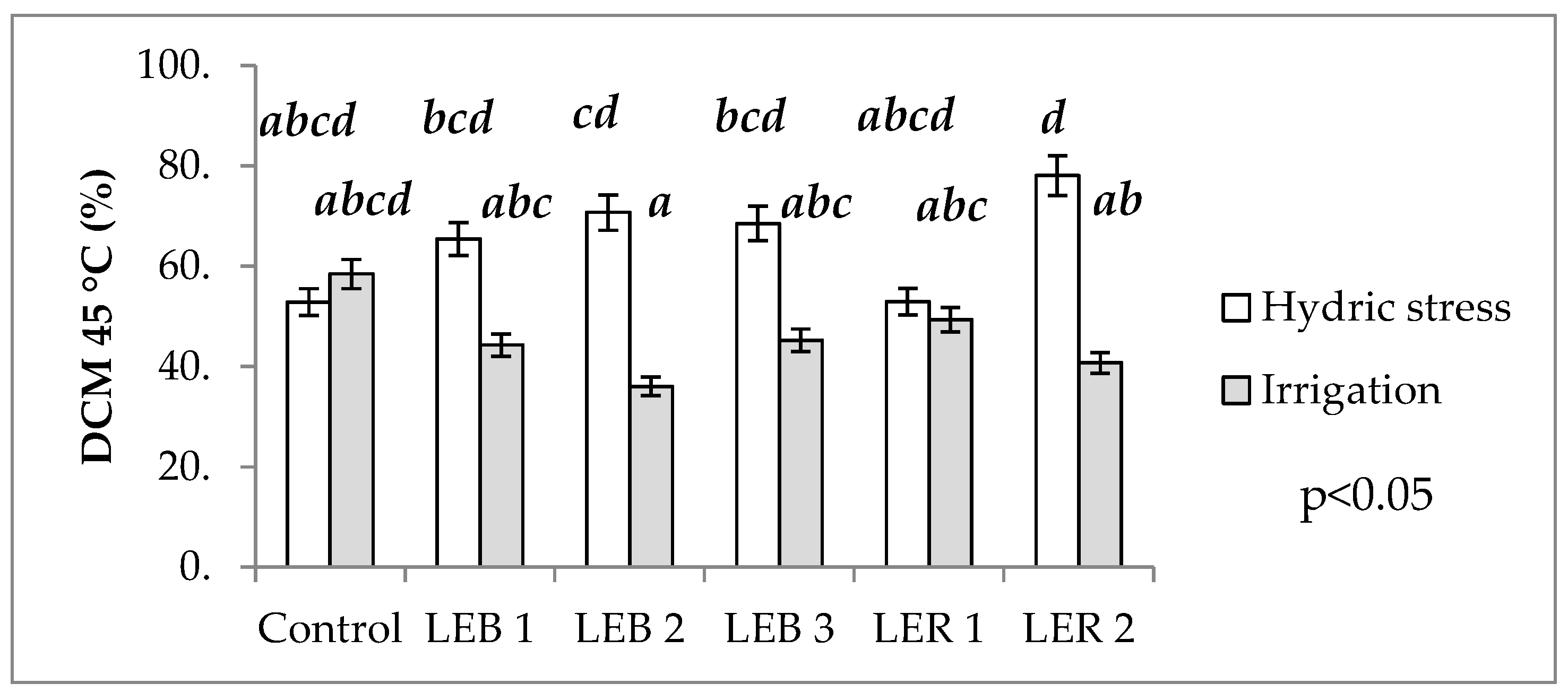
2.2.4. Damage to Cell Membranes at 40 and 45 °C
2.3. Agronomic Variables
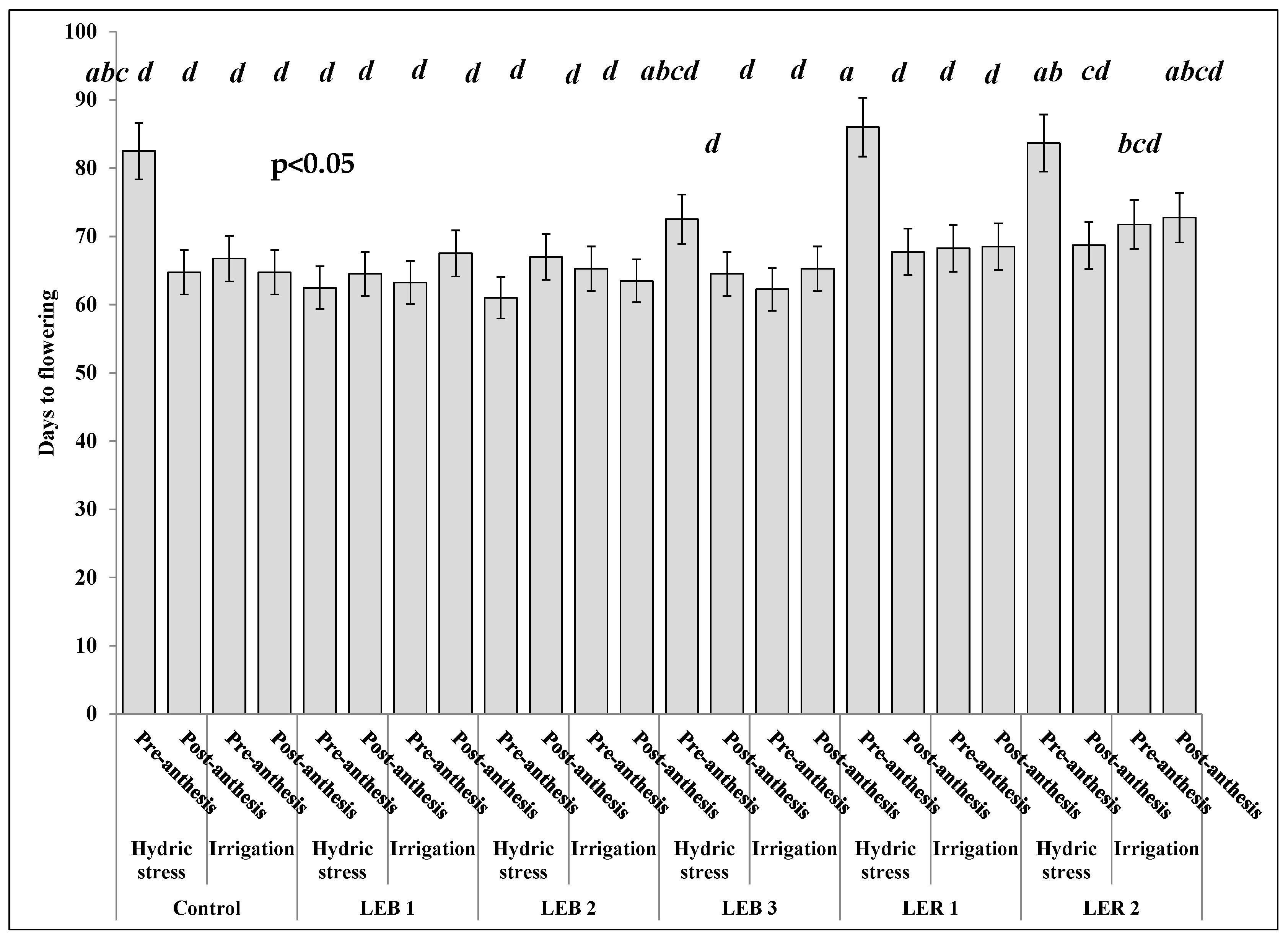
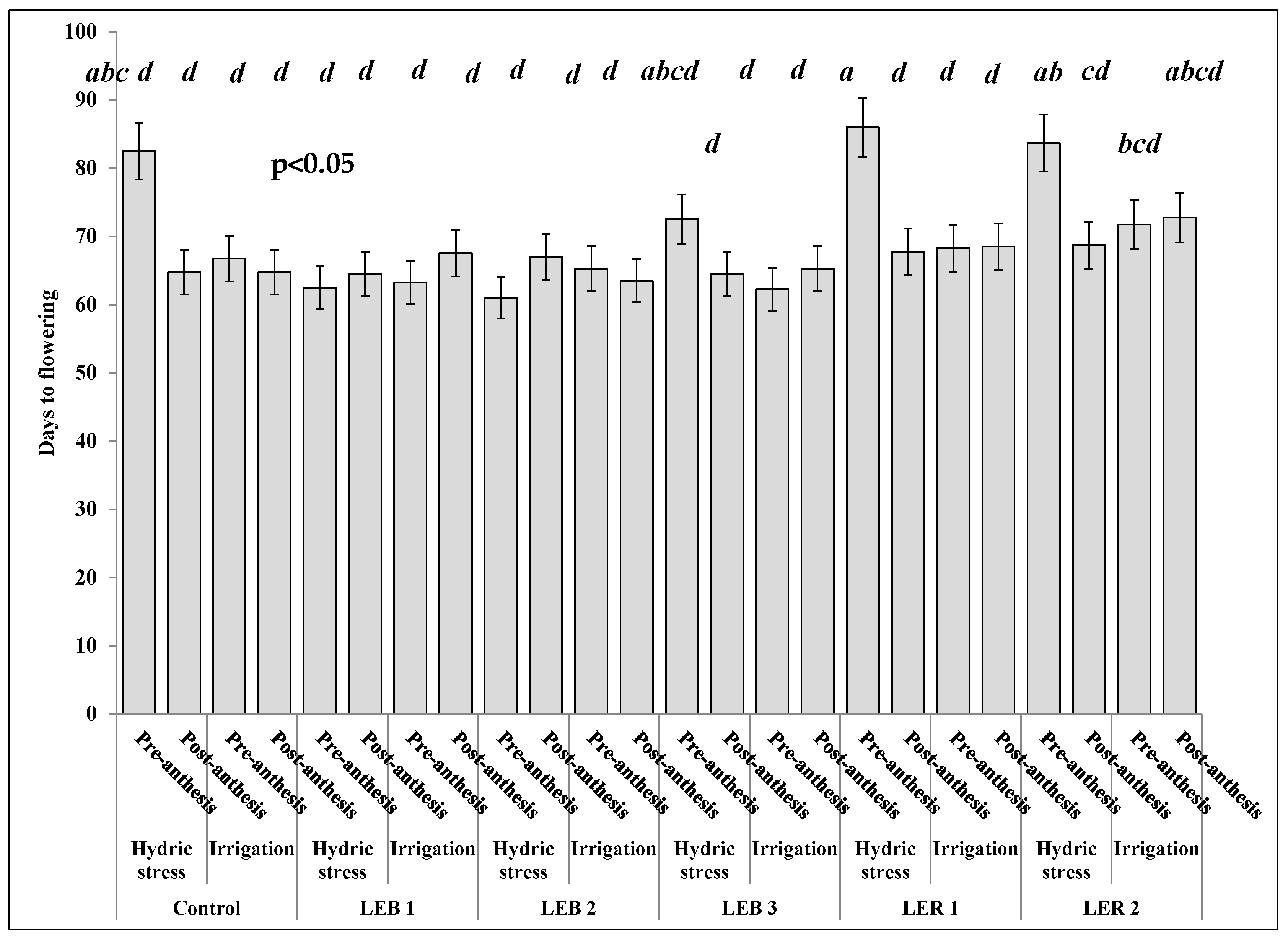
2.3.1. Days to Flowering
2.3.2. Plant Height
2.3.3. Panicle Length
2.3.4. Grain Weight per Panicle (GWP), Weight of 100 Grains (W100G), and Number of Grains per Panicle (NGP)
3. Discussion
3.1. Physiological Variables
3.1.1. Maximum Quantum Yield of PSII (Fv/Fm) under Hydric Stress
3.1.2. Photosynthetic Rate (PN), Stomatal Conductance (gs), and Transpiration (E)
3.1.3. Relative Water Content (RWC)
3.1.4. Damage to Cell Membranes at 40 and 45 °C
3.2. Agronomic Variables
3.2.1. Days to Flowering
3.2.2. Plant Height
3.2.3. Panicle Length
3.2.4. Grain Weight per Panicle, Weight of 100 Grains, and Number of Grains per Panicle
4. Materials and Methods
4.1. Experimental Location
4.2. Experimental Conditions
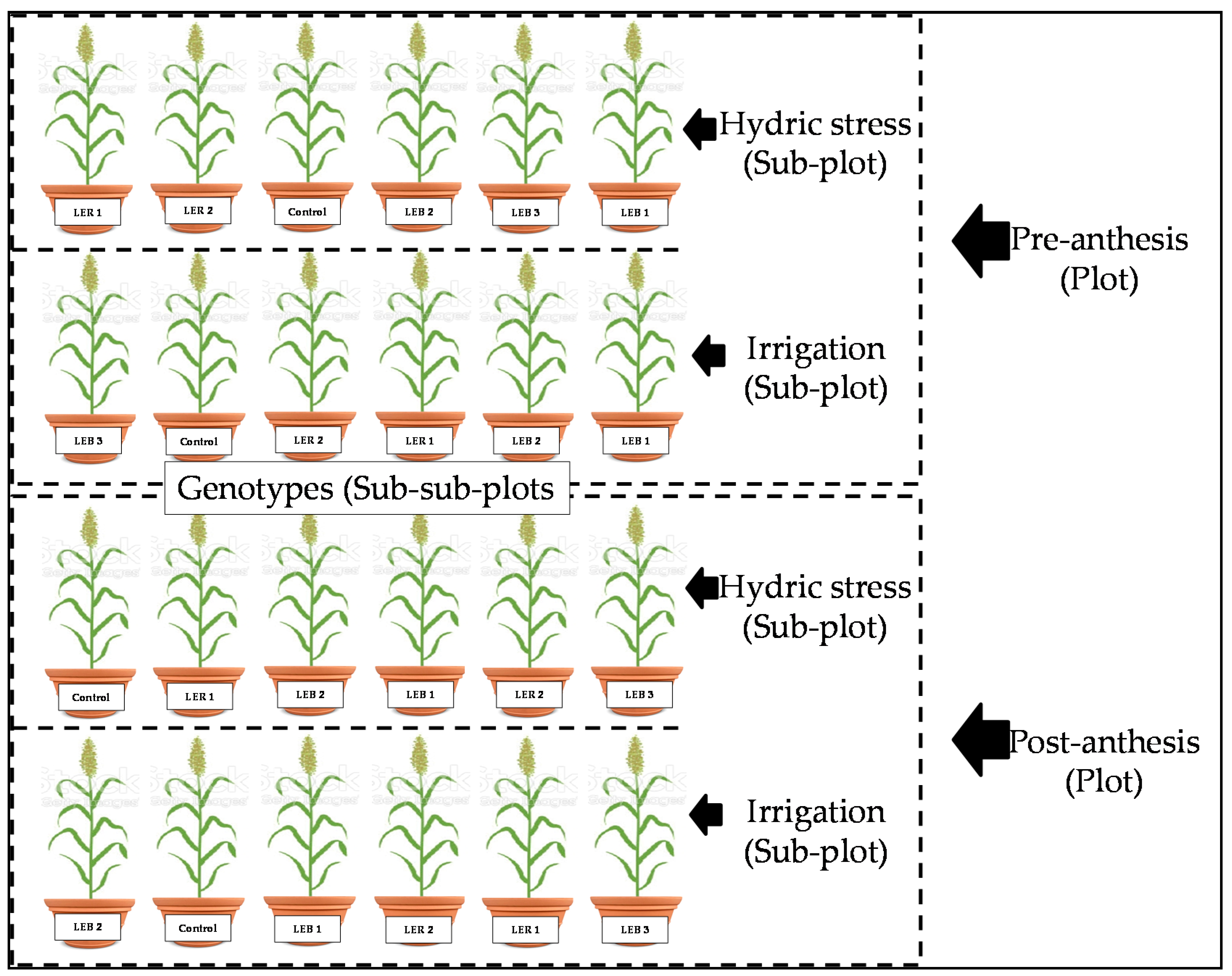
4.3. Experimental Design
4.4. Physiological Variables
- F0 = Minimum leaf chlorophyll fluorescence after dark acclimation
- Fm = Maximum leaf chlorophyll fluorescence after dark acclimation
- Fv = Variable leaf chlorophyll fluorescence after dark acclimation
4.5. Agronomic Variables
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Henckel, P. Physiology of Plants under Drought. Annu. Rev. Plant Physiol. 1964, 15, 363–386. [Google Scholar] [CrossRef]
- Loffroy, O.; Hubac, C.; Vieira da Silva, J. Effect of Temperature on Drought Resistance and Growth of Cotton Plants. Physiologia Plantarum 1983, 59, 297–301. [Google Scholar] [CrossRef]
- Siddique, M.; Hamid, A.; Islam, M. Drought Stress Effects on Water Relations of Wheat. Bot. Bull. Acad. Sin. 2000, 41, 35–39. [Google Scholar]
- Lipiec, J.; Doussan, C.; Nosalewicz, A.; Kondracka, K. Effect of Drought and Heat Stresses on Plant Growth and Yield: A Review. Int. Agrophys. 2013, 27, 463–477. [Google Scholar] [CrossRef]
- Secretaría de Agricultura y Desrrollo Rural. Servicio de Información Agroalimentaria y Pesquera—SIAP; Secretaría de Agricultura y Desrrollo Rural: Mexico City, Mexico, 2010. [Google Scholar]
- Davies, J. The importance of sorghum in the semi-arid tropics. In Proceedings of the International Workshop on Sorghum Diseases, Hyderabad, India, 11–15 December 1978; pp. 6–7. [Google Scholar]
- Smith, R.; Bhaskaran, S. Sorghum [Sorghum bicolor (L.) moench]. In Crops I; Springer: Berlin/Heidelberg, Germany, 1986; pp. 220–233. [Google Scholar]
- Taylor, J. Overview: Importance of sorghum in Africa. In Proceedings of the Afripro, Workshop on the Proteins of Sorghum and Millets: Enhancing Nutritional and Functional Properties for Africa, Pretoria, South Africa, 2–4 April 2003; Volume 2. [Google Scholar]
- Eck, H.; Musick, J. Plant Water Stress Effects on Irrigated Grain Sorghum. I. Effects on Yield 1. Crop Sci. 1979, 19, 589–592. [Google Scholar] [CrossRef]
- Wright, G.; Smith, R.; McWilliam, J. Differences between Two Grain Sorghum Genotypes in Adaptation to Drought Stress. I. Crop Growth and Yield Responses. Aust. J. Agric. Res. 1983, 34, 615–626. [Google Scholar] [CrossRef]
- Craufurd, P.; Peacock, J. Effect of Heat and Drought Stress on Sorghum (Sorghum Bicolor). II. Grain Yield. Exp. Agric. 1993, 29, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Queiroz, M.S.; Oliveira, C.E.; Steiner, F.; Zuffo, A.M.; Zoz, T.; Vendruscolo, E.P.; Silva, M.V.; Mello, B.; Cabra, R.; Menis, F.T. Drought Stresses on Seed Germination and Early Growth of Maize and Sorghum. J. Agric. Sci. 2019, 11, 310–318. [Google Scholar] [CrossRef]
- Bulmer, M. The Effect of Selection on Genetic Variability. Am. Nat. 1971, 105, 201–211. [Google Scholar] [CrossRef]
- Gilbert, J.; Lewis, R.; Wilkinson, M.; Caligari, P. Developing an Appropriate Strategy to Assess Genetic Variability in Plant Germplasm Collections. Theor. Appl. Genet. 1999, 98, 1125–1131. [Google Scholar] [CrossRef]
- Akman, H.; Zhang, C.; Ejeta, G. Physio-morphological, Biochemical, and Anatomical Traits of Drought-tolerant and Susceptible Sorghum Cultivars under Pre-and Post-anthesis Drought. Physiologia Plantarum 2021, 172, 912–921. [Google Scholar] [CrossRef]
- Rosenow, D.; Quisenberry, J.; Wendt, C.; Clark, L. Drought Tolerant Sorghum and Cotton Germplasm. Agric. Water Manag. 1983, 7, 207–222. [Google Scholar] [CrossRef]
- Rauf, S.; Al-Khayri, J.M.; Zaharieva, M.; Monneveux, P.; Khalil, F. Breeding strategies to enhance drought tolerance in crops. In Advances in Plant Breeding Strategies: Agronomic, Abiotic and Biotic Stress Traits; Springer: Berlin/Heidelberg, Germany, 2016; pp. 397–445. [Google Scholar]
- Singh, M.; Srivastava, J.; Kumar, A. Cell Membrane Stability in Relation to Drought Tolerance in Wheat Genotypes. J. Agron. Crop Sci. 1992, 168, 186–190. [Google Scholar] [CrossRef]
- Rad, M.R.N.; Kadir, M.A.; Yusop, M.R.; Jaafar, H.Z.; Danaee, M. Gene Action for Physiological Parameters and Use of Relative Water Content (RWC) for Selection of Tolerant and High Yield Genotypes in F2 Population of Wheat. Aust. J. Crop Sci. 2013, 7, 407–413. [Google Scholar]
- Tsago, Y.; Andargie, M.; Takele, A. In Vitro Selection of Sorghum (Sorghum bicolor (L) Moench) for Polyethylene Glycol (PEG) Induced Drought Stress. Plant Sci. Today 2014, 1, 62–68. [Google Scholar] [CrossRef]
- Hemantaranjan, A.; Bhanu, A.N.; Singh, M.; Yadav, D.; Patel, P.; Singh, R.; Katiyar, D. Heat Stress Responses and Thermotolerance. Adv. Plants Agric. Res. 2014, 1, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rehman, S.U.; Bilal, M.; Rana, R.M.; Tahir, M.N.; Shah, M.K.N.; Ayalew, H.; Yan, G. Cell Membrane Stability and Chlorophyll Content Variation in Wheat (Triticum aestivum) Genotypes under Conditions of Heat and Drought. Crop Pasture Sci. 2016, 67, 712–718. [Google Scholar] [CrossRef]
- Polash, M.A.S.; Sakil, M.A.; Tahjib-Ul-Arif, M.; Hossain, M.A. Effect of Salinity on Osmolytes and Relative Water Content of Selected Rice Genotypes. Trop. Plant Res. 2018, 5, 227–232. [Google Scholar] [CrossRef]
- Chen, W.; Yao, X.; Cai, K.; Chen, J. Silicon Alleviates Drought Stress of Rice Plants by Improving Plant Water Status, Photosynthesis and Mineral Nutrient Absorption. Biol. Trace Elem. Res. 2011, 142, 67–76. [Google Scholar] [CrossRef]
- Da Silva, E.C.; Nogueira, R.; da Silva, M.A.; de Albuquerque, M.B. Drought Stress and Plant Nutrition. Plant Stress 2011, 5, 32–41. [Google Scholar]
- Meng, S.; Zhang, C.; Su, L.; Li, Y.; Zhao, Z. Nitrogen Uptake and Metabolism of Populus simonii in Response to PEG-Induced Drought Stress. Environ. Exp. Bot. 2016, 123, 78–87. [Google Scholar] [CrossRef]
- Anjum, S.A.; Xie, X.; Wang, L.; Saleem, M.F.; Man, C.; Lei, W. Morphological, Physiological and Biochemical Responses of Plants to Drought Stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Fathi, A.; Tari, D.B. Effect of Drought Stress and Its Mechanism in Plants. Int. J. Life Sci. 2016, 10, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Maghsoudi, K.; Emam, Y.; Pessarakli, M. Effect of Silicon on Photosynthetic Gas Exchange, Photosynthetic Pigments, Cell Membrane Stability and Relative Water Content of Different Wheat Cultivars under Drought Stress Conditions. J. Plant Nutr. 2016, 39, 1001–1015. [Google Scholar] [CrossRef]
- Sinclair, T.; Bennett, J.; Muchow, R. Relative Sensitivity of Grain Yield and Biomass Accumulation to Drought in Field-Grown Maize. Crop Sci. 1990, 30, 690–693. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Gómez, M.; Azcón, R. Influence of Different Glomus Species on the Time-Course of Physiological Plant Responses of Lettuce to Progressive Drought Stress Periods. Plant Sci. 1995, 110, 37–44. [Google Scholar] [CrossRef]
- Yin, L.; Wang, S.; Liu, P.; Wang, W.; Cao, D.; Deng, X.; Zhang, S. Silicon-Mediated Changes in Polyamine and 1-Aminocyclopropane-1-Carboxylic Acid Are Involved in Silicon-Induced Drought Resistance in Sorghum bicolor L. Plant Physiol. Biochem. 2014, 80, 268–277. [Google Scholar] [CrossRef]
- Xu, W.; Cui, K.; Xu, A.; Nie, L.; Huang, J.; Peng, S. Drought Stress Condition Increases Root to Shoot Ratio via Alteration of Carbohydrate Partitioning and Enzymatic Activity in Rice Seedlings. Acta Physiol. Plant. 2015, 37, 9. [Google Scholar] [CrossRef]
- Ding, J.; Johnson, E.A.; Martin, Y.E. Optimization of Leaf Morphology in Relation to Leaf Water Status: A Theory. Ecol. Evol. 2020, 10, 1510–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornic, G.; Massacci, A. Leaf photosynthesis under drought stress. In Photosynthesis and the Environment; Springer: Berlin/Heidelberg, Germany, 1996; pp. 347–366. [Google Scholar]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-Induced Responses of Photosynthesis and Antioxidant Metabolism in Higher Plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef] [PubMed]
- Zivcak, M.; Brestic, M.; Balatova, Z.; Drevenakova, P.; Olsovska, K.; Kalaji, H.M.; Yang, X.; Allakhverdiev, S.I. Photosynthetic Electron Transport and Specific Photoprotective Responses in Wheat Leaves under Drought Stress. Photosynth. Res. 2013, 117, 529–546. [Google Scholar] [CrossRef]
- Wang, Z.; Li, G.; Sun, H.; Ma, L.; Guo, Y.; Zhao, Z.; Gao, H.; Mei, L. Effects of Drought Stress on Photosynthesis and Photosynthetic Electron Transport Chain in Young Apple Tree Leaves. Biol. Open 2018, 7, bio035279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dąbrowski, P.; Baczewska-Dąbrowska, A.H.; Kalaji, H.M.; Goltsev, V.; Paunov, M.; Rapacz, M.; Wójcik-Jagła, M.; Pawluśkiewicz, B.; Bąba, W.; Brestic, M. Exploration of Chlorophyll a Fluorescence and Plant Gas Exchange Parameters as Indicators of Drought Tolerance in Perennial Ryegrass. Sensors 2019, 19, 2736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stirbet, A.; Lazár, D.; Kromdijk, J.; Govindjee. Chlorophyll a Fluorescence Induction: Can Just a One-Second Measurement Be Used to Quantify Abiotic Stress Responses? Photosynthetica 2018, 56, 86–104. [Google Scholar] [CrossRef]
- Romanowska-Duda, Z.; Grzesik, M.; Janas, R. Maximal Efficiency of PSII as a Marker of Sorghum Development Fertilized with Waste From a Biomass Biodigestion to Methane. Front. Plant Sci. 2019, 9, 1920. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, U.; Hormann, H.; Neubauer, C.; Klughammer, C. Assessment of Photosystem II Photochemical Quantum Yield by Chlorophyll Fluorescence Quenching Analysis. Funct. Plant Biol. 1995, 22, 209–220. [Google Scholar] [CrossRef]
- Bilger, W.; Schreiber, U.; Bock, M. Determination of the Quantum Efficiency of Photosystem II and of Non-Photochemical Quenching of Chlorophyll Fluorescence in the Field. Oecologia 1995, 102, 425–432. [Google Scholar] [CrossRef]
- Bradbury, M.; Baker, N.R. Analysis of the Slow Phases of the in Vivo Chlorophyll Fluorescence Induction Curve. Changes in the Redox State of Photosystem II Electron Acceptors and Fluorescence Emission from Photosystems I and II. Biochimica et Biophysica Acta 1981, 635, 542–551. [Google Scholar] [CrossRef]
- Eichelmann, H.; Laisk, A. Cooperation of Photosystems II and I in Leaves as Analyzed by Simultaneous Measurements of Chlorophyll Fluorescence and Transmittance at 800 Nm. Plant Cell Physiol. 2000, 41, 138–147. [Google Scholar] [CrossRef] [Green Version]
- Hamdani, S.; Wang, H.; Zheng, G.; Perveen, S.; Qu, M.; Khan, N.; Khan, W.; Jiang, J.; Li, M.; Liu, X. Genome-wide Association Study Identifies Variation of Glucosidase Being Linked to Natural Variation of the Maximal Quantum Yield of Photosystem II. Physiologia Plantarum 2019, 166, 105–119. [Google Scholar] [CrossRef]
- Cerda Hurtado, I.M. Identificación de Marcadores Moleculares Ligados a Caracteres de Interés En Materiales Élite de Sorgo Dulce Sorghum Bicolor (L.) Moench de La FA-UANL; UANL: San Nicolás de los Garza, Mexico, 2014. [Google Scholar]
- Mendoza-Onofre, L.E.; Cisneros-López, M.E.; Galicia-Juárez, M.; Hernández-Martínez, M. Líneas Mantenedoras (Líneas B) y Restauradoras (Líneas R) de Sorgo Granífero Adaptadas a Los Valles Altos Centrales de Mexico. Revista Fitotecnia Mexicana 2017, 40, 107–110. [Google Scholar] [CrossRef]
- Kohzuma, K.; Cruz, J.A.; Akashi, K.; Hoshiyasu, S.; Munekage, Y.N.; Yokota, A.; Kramer, D.M. The Long-term Responses of the Photosynthetic Proton Circuit to Drought. Plant Cell Environ. 2009, 32, 209–219. [Google Scholar] [CrossRef]
- Kapanigowda, M.H.; Perumal, R.; Djanaguiraman, M.; Aiken, R.M.; Tesso, T.; Prasad, P.; Little, C.R. Genotypic Variation in Sorghum [Sorghum bicolor (L.) Moench] Exotic Germplasm Collections for Drought and Disease Tolerance. SpringerPlus 2013, 2, 650. [Google Scholar] [CrossRef] [Green Version]
- Netondo, G.W.; Onyango, J.C.; Beck, E. Sorghum and Salinity: II. Gas Exchange and Chlorophyll Fluorescence of Sorghum under Salt Stress. Crop Sci. 2004, 44, 806–811. [Google Scholar] [CrossRef]
- Qaseem, M.F.; Qureshi, R.; Shaheen, H. Effects of Pre-Anthesis Drought, Heat and Their Combination on the Growth, Yield and Physiology of Diverse Wheat (Triticum Aestivum L.) Genotypes Varying in Sensitivity to Heat and Drought Stress. Sci. Rep. 2019, 9, 6955. [Google Scholar] [CrossRef] [Green Version]
- Elsheery, N.I.; Cao, K.-F. Gas Exchange, Chlorophyll Fluorescence, and Osmotic Adjustment in Two Mango Cultivars under Drought Stress. Acta Physiologiae Plantarum 2008, 30, 769–777. [Google Scholar] [CrossRef]
- Bethke, P.C.; Drew, M.C. Stomatal and Nonstomatal Components to Inhibition of Photosynthesis in Leaves of Capsicum Annuum during Progressive Exposure to NaCl Salinity. Plant Physiol. 1992, 99, 219–226. [Google Scholar] [CrossRef]
- Kao, W.-Y.; Tsai, T.-T.; Shih, C.-N. Photosynthetic Gas Exchange and Chlorophyll a Fluorescence of Three Wild Soybean Species in Response to NaCl Treatments. Photosynthetica 2003, 41, 415–419. [Google Scholar] [CrossRef]
- Condon, A.G.; Richards, R.; Rebetzke, G.; Farquhar, G. Breeding for High Water-Use Efficiency. J. Exp. Bot. 2004, 55, 2447–2460. [Google Scholar] [CrossRef] [Green Version]
- Cornic, G. Drought Stress Inhibits Photosynthesis by Decreasing Stomatal Aperture—Not by Affecting ATP Synthesis. Trends Plant Sci. 2000, 5, 187–188. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Cornic, G. Photosynthetic Carbon Assimilation and Associated Metabolism in Relation to Water Deficits in Higher Plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, M.; Balota, M.; Delgado, M.; Amani, I.; Fischer, R. Physiological and Morphological Traits Associated with Spring Wheat Yield under Hot, Irrigated Conditions. Funct. Plant Biol. 1994, 21, 717–730. [Google Scholar] [CrossRef]
- Amani, I.; Fischer, R.; Reynolds, M. Canopy Temperature Depression Association with Yield of Irrigated Spring Wheat Cultivars in a Hot Climate. J. Agron. Crop Sci. 1996, 176, 119–129. [Google Scholar] [CrossRef]
- Fischer, R.; Rees, D.; Sayre, K.; Lu, Z.; Condon, A.; Saavedra, A.L. Wheat Yield Progress Associated with Higher Stomatal Conductance and Photosynthetic Rate, and Cooler Canopies. Crop Sci. 1998, 38, 1467–1475. [Google Scholar] [CrossRef]
- Condon, A.; Reynolds, M.; Brennan, J.; van Ginkel, M.; Trethowan, R.; Rebetzke, G.; Bonnett, D.; Richards, R.; Farquhar, G. Stomatal aperture related traits and yield potential in bread wheat. In Proceedings of the International Symposium on Wheat Yield Potential: Challenges to International Wheat Breeding—Extended Abstracts, Ciudad Obregón, Mexico, 20–24 March 2006; p. 126. [Google Scholar]
- Mahfouz, H.; Megawer, E.A.; Maher, A.; Shaaban, A. Integrated Effect of Planting Dates and Irrigation Regimes on Morpho-Physiological Response, Forage Yield and Quality, and Water Use Efficiency of Clitoria (Clitoria ternatea L.) in Arid Region. Arch. Agron. Soil Sci. 2020, 66, 152–167. [Google Scholar] [CrossRef]
- Agami, R.A.; Alamri, S.A.; Abd El-Mageed, T.; Abousekken, M.; Hashem, M. Role of Exogenous Nitrogen Supply in Alleviating the Deficit Irrigation Stress in Wheat Plants. Agric. Water Manag. 2018, 210, 261–270. [Google Scholar] [CrossRef]
- ElBasyoni, I.; Saadalla, M.; Baenziger, S.; Bockelman, H.; Morsy, S. Cell Membrane Stability and Association Mapping for Drought and Heat Tolerance in a Worldwide Wheat Collection. Sustainability 2017, 9, 1606. [Google Scholar] [CrossRef] [Green Version]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat Tolerance in Plants: An Overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Tahir, A.; Qadir, M.; Saif, R.; Sattar, S.; Tahir, S. Morphological and Yield Response of Pulses against Drought Stress: A Review. Turk. J. Agric. Food Sci. Technol. 2019, 7, 202–208. [Google Scholar] [CrossRef] [Green Version]
- Reddy, B.V.; Ramaiah, B.; Ashok Kumar, A.; Reddy, P.S. Evaluation of Sorghum Genotypes for the Stay-Green Trait and Grain Yield. J. SAT Agric. Res. 2007, 3, 1–4. [Google Scholar]
- Jabereldar, A.A.; El Naim, A.M.; Abdalla, A.A.; Dagash, Y.M. Effect of Water Stress on Yield and Water Use Efficiency of Sorghum (Sorghum bicolor L. Moench) in Semi-Arid Environment. Int. J. Agric. For. 2017, 7, 1–6. [Google Scholar]
- Assefa, Y.; Staggenborg, S.A.; Prasad, V.P. Grain Sorghum Water Requirement and Responses to Drought Stress: A Review. Crop Manag. 2010, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ashok Kumar, A.; Reddy, B.V.S.; Sharma, H.C.; Hash, C.T.; Srinivasa Rao, P.; Ramaiah, B.; Reddy, P.S. Recent Advances in Sorghum Genetic Enhancement Research at ICRISAT. Am. J. Plant Sci. 2011, 2, 589–600. [Google Scholar] [CrossRef] [Green Version]
- García, E. Modificaciones al Sistema de Clasificación Climática de Köppen; Universidad Nacional Autónoma de Mexico: Mexico City, Mexico, 2004. [Google Scholar]
- Blum, A.; Ebercon, A. Cell Membrane Stability as a Measure of Drought and Heat Tolerance in Wheat1. Crop Sci. 1981, 21, 43. [Google Scholar] [CrossRef]
- Sade, N.; Galkin, E.; Moshelion, M. Measuring Arabidopsis, Tomato and Barley Leaf Relative Water Content (RWC). Bio-protocol 2015, 5, e1451. [Google Scholar] [CrossRef] [Green Version]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; González, L.; Tablada, M.; y Robledo, C.W. Infostat—Software Estadístico; Grupo InfoStat, FCA, Universidad Nacional de Córdoba: Córdoba, Argentina, 2008. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. V. | D.F. | Fv/Fm | PN (μmol CO2 m−2 s−1) | gs (mol H2O m−2 s−1) | E (mmol H2O m−2 s−1) | RWC (%) | DCM 40 (%) | DCM 45 (%) |
|---|---|---|---|---|---|---|---|---|
| Stage (S) | 1 | 0.01 ns | 18.2 ns | 0.02 ns | 24.7 * | 45.7 ns | 2737.1 ns | 646.0 ns |
| Error (a) | 6 | 0.02 | 18.2 | 0.02 | 2.7 | 153.3 | 708.6 | 203.3 |
| Humidity (H) | 1 | 5.47 ** | 12,248.2 ** | 1.83 ** | 1045.5 ** | 25,696.7 ** | 752.1 ns | 7744.9 ** |
| H x S | 1 | 0.05 ns | 69.5 ns | 0.12 ** | 6.5 ns | 19.3 ns | 2377.5 ns | 831.3 ns |
| Error (b) | 6 | 0.02 | 15.9 | 0.01 | 1.4 | 115.9 | 513.4 | 182.4 |
| Genotype (G) | 5 | 0.09 * | 14.1 ns | 0.01 ns | 3.4 ns | 359.1 * | 267.5 ns | 121.8 ns |
| H x G | 5 | 0.11 * | 5.9 ns | 0.01 ns | 2.8 ns | 300.24 ns | 854.2 ns | 1073.6 * |
| S x G | 5 | 0.01 ns | 20.6 ns | 0.01 ns | 2.4 ns | 147.1 ns | 261.0 ns | 130.5 ns |
| H x S x G | 5 | 0.01 ns | 18.7 ns | 0.01 ns | 2.3 ns | 158.3 ns | 494.2 ns | 279.9 ns |
| Error (c) | 60 | 0.04 | 10.9 | 0.01 | 1.8 | 140.2 | 516.0 | 259.0 |
| C.V. | 38.8 | 35.5 | 60.6 | 27.8 | 23.2 | 71.3 | 29.3 |
| Variable | Hydric Stress | Irrigation |
|---|---|---|
| PN (µmol CO2 m−2 s−1) | −1.9 b | 20.6 a |
| gs (mol H2O m−2 s−1) | 0.06 b | 0.34 a |
| E (mmol H2O m−2 s−1) | 1.5 b | 8.1 a |
| RWC (%) | 34.8 b | 67.5 a |
| DCM 40 (%) | 32.6 | 24.9 |
| DCM 45 (%) | 81.2 a | 51.4 b |
| S. V. | d. f. † | Days to Flowering. | d. f. | Plant Ht. (cm) | d. f. | Panicle Lenght (cm) | d. f. | GWP (gr) | W100G (gr) | NGP |
|---|---|---|---|---|---|---|---|---|---|---|
| Stage (S) | 1 | 130.8 ns | 1 | 6750.3 ** | 1 | 968.7 ** | 1 | 2214.9 ** | 0.31 ns | 1,684,749 ns |
| Error (a) | 6 | 26.3 | 6 | 258.6 | 6 | 23.4 | 6 | 119.9 | 0.34 | 1,150,454.00 |
| Humidity (H) | 1 | 165.0 * | 1 | 9861.8 ** | 1 | 709.9 ** | 1 | 20,009.5 ** | 6.09 ** | 40,694,690 ** |
| H × S | 1 | 383.3 ** | 1 | 7332.5 ** | 1 | 995.1 ** | 1 | 1846.9 ** | 0.64 * | 1,296,639 ns |
| Error (b) | 6 | 16.1 | 6 | 185.6 | 6 | 10.4 | 5 | 101.3 | 0.05 | 465,604 |
| Genotype (G) | 5 | 199.0 ** | 5 | 296.8 ns | 5 | 39.9 * | 5 | 1520.7 ** | 1.30 ** | 1,003,595 ns |
| H × G | 5 | 36.3 ns | 5 | 63.6 ns | 5 | 8.2 ns | 5 | 51.5 ns | 0.49 ** | 288,298 ns |
| S × G | 5 | 80.6 ** | 5 | 147.7 ns | 5 | 19.9 ns | 5 | 170.8 ns | 0.22 * | 350,376 ns |
| H × S × G | 5 | 66.7 ** | 5 | 27.6 ns | 5 | 16.4 ns | 4 | 191.9 ns | 0.16 ns | 265,983 ns |
| Error (c) | 50 | 19.6 | 60 | 192.5 | 55 | 15.5 | 26 | 295.5 | 0.09 | 872,887 |
| C. V. | 6.5 | 16.8 | 16.6 | 41.2 | 17.1 | 40.8 |
| Stage | Soil Humidity | Days to Flowering | Plant ht. (cm) | Panicle Length (cm) | GWP (gr) | W100G (gr) | NGP | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pre-anthesis | Hydric stress † | 74 | a | 55.4 | b | 13.9 | b | 8.2 | b | 1.03 | b | 880 | b |
| Irrigation | 66 | b | 93.1 | a | 26.2 | a | 61.6 | a | 2.04 | a | 3000 | a | |
| Post-anthesis | Hydric stress | 66 | b | 89.6 | a | 27.4 | a | 16.2 | b | 1.32 | b | 1068 | b |
| Irrigation | 67 | b | 92.4 | a | 26.5 | a | 46.5 | a | 1.81 | a | 2693 | a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galicia-Juárez, M.; Zavala-García, F.; Sinagawa-García, S.R.; Gutiérrez-Diez, A.; Williams-Alanís, H.; Cisneros-López, M.E.; Valle-Gough, R.E.; Flores-Garivay, R.; Santillano-Cázares, J. Identification of Sorghum (Sorghum bicolor (L.) Moench) Genotypes with Potential for Hydric and Heat Stress Tolerance in Northeastern Mexico. Plants 2021, 10, 2265. https://doi.org/10.3390/plants10112265
Galicia-Juárez M, Zavala-García F, Sinagawa-García SR, Gutiérrez-Diez A, Williams-Alanís H, Cisneros-López ME, Valle-Gough RE, Flores-Garivay R, Santillano-Cázares J. Identification of Sorghum (Sorghum bicolor (L.) Moench) Genotypes with Potential for Hydric and Heat Stress Tolerance in Northeastern Mexico. Plants. 2021; 10(11):2265. https://doi.org/10.3390/plants10112265
Chicago/Turabian StyleGalicia-Juárez, Marisol, Francisco Zavala-García, Sugey Ramona Sinagawa-García, Adriana Gutiérrez-Diez, Héctor Williams-Alanís, María Eugenia Cisneros-López, Raúl Enrique Valle-Gough, Rodrigo Flores-Garivay, and Jesús Santillano-Cázares. 2021. "Identification of Sorghum (Sorghum bicolor (L.) Moench) Genotypes with Potential for Hydric and Heat Stress Tolerance in Northeastern Mexico" Plants 10, no. 11: 2265. https://doi.org/10.3390/plants10112265
APA StyleGalicia-Juárez, M., Zavala-García, F., Sinagawa-García, S. R., Gutiérrez-Diez, A., Williams-Alanís, H., Cisneros-López, M. E., Valle-Gough, R. E., Flores-Garivay, R., & Santillano-Cázares, J. (2021). Identification of Sorghum (Sorghum bicolor (L.) Moench) Genotypes with Potential for Hydric and Heat Stress Tolerance in Northeastern Mexico. Plants, 10(11), 2265. https://doi.org/10.3390/plants10112265