
Comparative Study of Ovule Development between Wild (Passiflora foetida L.) and Cultivated (P. edulis Sims) Species of Passiflora L. Provide Insights into Its Differential Developmental Patterns

 ,
,
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
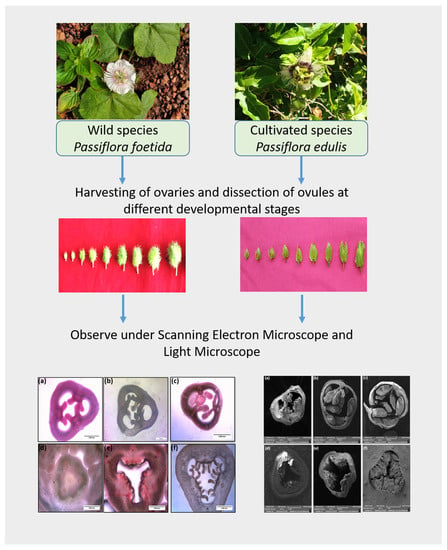
2. Materials and Methods
2.1. Plant Acclimatisation and Growth
2.2. Tissue Harvesting
2.3. Histology and Staining
2.4. Scanning Electron Microscopy
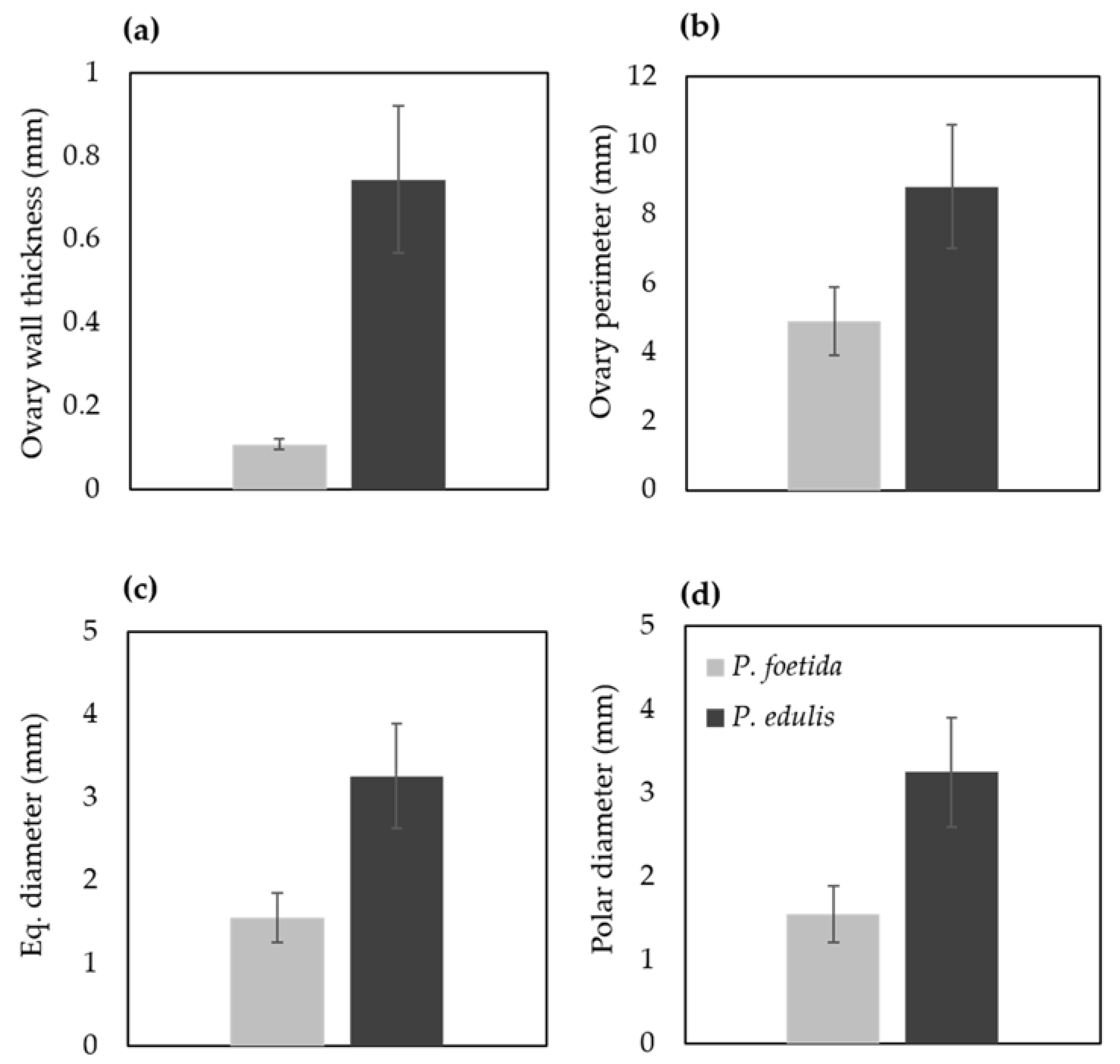
2.5. Measurement of Ovary and Ovule Characteristics
3. Results
3.1. Morphological Differences between P. foetida and P. edulis
3.2. Floral Characterization of Two Species
3.3. Ovule Development in Wild and Cultivated Species of Passiflora
3.4. Pollen Morphology
4. Discussion
4.1. Differences between Wild and Cultivated Species of Passiflora
4.2. Ovule Development in P. foetida and P. edulis
4.3. Ovule Size and Number between P. foetida and P. edulis
4.4. Pollen Morphology
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodriguez-Amaya, D.B. Passion Fruits. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Ed.; Academic Press: Oxford, UK, 2003; pp. 4368–4373. ISBN 978-0-12-227055-0. [Google Scholar]
- Abreu, P.P.; Souza, M.M.; Santos, E.A.; Pires, M.V.; Pires, M.M.; de Almeida, A.-A.F. Passion flower hybrids and their use in the ornamental plant market: Perspectives for sustainable development with emphasis on Brazil. Euphytica 2009, 166, 307–315. [Google Scholar] [CrossRef]
- Ingale, A.G.; Hivrale, A.U. Pharmacological studies of Passiflora sp. and their bioactive compounds. Afr. J. Plant Sci. 2010, 4, 10. [Google Scholar]
- Shah, J.J.; Dave, Y.S. Tendrils of Passiflora foetida: Histogenesis and morphology. Am. J. Bot. 1970, 57, 786–793. [Google Scholar] [CrossRef]
- Simpson, M.G. 8—Diversity and classification of flowering plants: Eudicots. In Plant Systematics, 2nd ed.; Simpson, M.G., Ed.; Academic Press: San Diego, CA, USA, 2010; pp. 275–448. ISBN 978-0-12-374380-0. [Google Scholar]
- Ulmer, T.; MacDougal, J.M. Passiflora: Passionflowers of the World; Timber Press: Portland, OR, USA, 2004; ISBN 0-88192-648-5. [Google Scholar]
- Moraes, A.M.; Milward-de-Azevedo, M.A.; Menini Neto, L.; de Faria, A.P.G. Distribution patterns of Passiflora L. (Passifloraceae s.s.) in the Serra Da Mantiqueira, Southeast Brazil. Braz. J. Bot. 2020, 43, 999–1012. [Google Scholar] [CrossRef]
- Casierra-Posada, F.; Jarma-Orozco, A. Chapter 22—Nutritional composition of Passiflora Species. In Nutritional Composition of Fruit Cultivars; Simmonds, M.S.J., Preedy, V.R., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 517–534. ISBN 978-0-12-408117-8. [Google Scholar]
- Silva, M.A.A.; Souza, M.M.; Silva, G.S.; Melo, C.A.F.; Corrêa, R.X.; Araújo, I.S.; Conceição, L.D.H.C.S. Analysis of transferability of microsatellite primers (SSR) in wild Passiflora species and intraspecific genetic diversity in Passiflora alata. Genet. Mol. Res. 2014, 13, 5908–5918. [Google Scholar] [CrossRef]
- Vianna, L.S.; Pereira, T.N.S.; Santos, E.A.; Viana, A.P.; Pereira, M.G.; Ramos, H.C.C.; Rossi, A.A.B. Research article ISSR and SSR markers for determining genetic relationships among three wild species of Passiflora. Genet. Mol. Res. 2019, 18. [Google Scholar] [CrossRef]
- De Santana, F.C.; Shinagawa, F.B.; Araujo, E.d.S.; Costa, A.M.; Mancini-Filho, J. Chemical composition and antioxidant capacity of Brazilian Passiflora seed oils. J. Food Sci. 2015, 80, C2647–C2654. [Google Scholar] [CrossRef] [PubMed]
- Ramaiya, S.D.; Bujang, J.S.; Zakaria, M.H. Genetic diversity in Passiflora species assessed by morphological and ITS sequence analysis. Sci. World J. 2014, 2014, 598313. [Google Scholar] [CrossRef] [PubMed]
- Rocha, D.; Batista, D.; Faleiro, F.; Rogalski, M.; Ribeiro, L.; Mercadante-Simões, M.; Yockteng, R.; Silva, M.; Soares, W.; Pinheiro, M.; et al. Passiflora spp. Passionfruit. In Biotechnology of Fruit and Nut Crops; Ritz, R., Ed.; CABI: London, UK, 2020; pp. 381–408. ISBN 978-1-78064-827-9. [Google Scholar]
- Miroddi, M.; Calapai, G.; Navarra, M.; Minciullo, P.L.; Gangemi, S. Passiflora incarnata L.: Ethnopharmacology, clinical application, safety and evaluation of clinical trials. J. Ethnopharmacol. 2013, 150, 791–804. [Google Scholar] [CrossRef] [PubMed]
- Porter-Utley, K.E. A revision of Passiflora L. subgenus Decaloba (DC.) Rchb. Supersection Cieca (Medik.) J. M. MacDougal & Feuillet (Passifloraceae). PhytoKeys 2014, 43, 1. [Google Scholar] [CrossRef]
- Sakalem, M.E.; Negri, G.; Tabach, R. Chemical composition of hydroethanolic extracts from five species of the Passiflora genus. Rev. Bras. Farmacogn. 2012, 22, 1219–1232. [Google Scholar] [CrossRef]
- He, X.; Luan, F.; Yang, Y.; Wang, Z.; Zhao, Z.; Fang, J.; Wang, M.; Zuo, M.; Li, Y. Passiflora edulis: An insight into current researches on phytochemistry and pharmacology. Front. Pharmacol. 2020, 11, 617. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.-W.; Lin, K.-H.; Chen, K.-H.; Wu, C.-W.; Chang, Y.-S. Flavonoid compounds and photosynthesis in Passiflora plant leaves under varying light intensities. Plants 2020, 9, 633. [Google Scholar] [CrossRef]
- Patil, A.S.; Lade, B.D.; Paikrao, H.M. A scientific update on Passiflora foetida. Eur. J. Med. Plants 2015, 5, 145–155. [Google Scholar] [CrossRef]
- Song, Y.; Wei, X.-Q.; Li, M.-Y.; Duan, X.-W.; Sun, Y.-M.; Yang, R.-L.; Su, X.-D.; Huang, R.-M.; Wang, H. Nutritional composition and antioxidant properties of the fruits of a chinese wild Passiflora foetida. Molecules 2018, 23, 459. [Google Scholar] [CrossRef] [PubMed]
- Zibadi, S.; Watson, R.R. Passion fruit (Passiflora edulis). Evid. Based Integr. Med. 2004, 1, 183–187. [Google Scholar] [CrossRef]
- Gunathilake, K.D.P.P.; Ranaweera, K.K.D.S.; Rupasinghe, H.P.V. Influence of boiling, steaming and frying of selected leafy vegetables on the in vitro anti-inflammation associated biological activities. Plants 2018, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Scorza, L.C.T.; Hernandes-Lopes, J.; Melo-de-Pinna, G.F.A.; Dornelas, M.C. Expression patterns of Passiflora edulis APETALA1/FRUITFULL homologues shed light onto tendril and corona identities. EvoDevo 2017, 8, 3. [Google Scholar] [CrossRef]
- Tripathi, P.C. Passion fruit. In Horticultural Crops of High Nutraceutical Values; Peter, K.V., Ed.; Brillion Publishing: New Delhi, India, 2018; pp. 245–270. ISBN 978-93-87445-10-9. [Google Scholar]
- Das, M.R.; Hossain, T.; Mia, M.A.B.; Ahmed, J.U.; Karim, A.J.M.S.; Hossain, M.M. Blooming pattern of passion fruit flower (Passiflora edulis Sims.) under diversified flashes. Am. J. Agric. Biol. Sci. 2013, 8. [Google Scholar] [CrossRef][Green Version]
- Dhawan, K.; Dhawan, S.; Sharma, A. Passiflora: A review update. J. Ethnopharmacol. 2004, 94, 1–23. [Google Scholar] [CrossRef]
- Hemingway, C.A.; Christensen, A.R.; Malcomber, S.T. B- and C-Class Gene Expression during corona development of the blue passionflower (Passiflora caerulea, Passifloraceae). Am. J. Bot. 2011, 98, 923–934. [Google Scholar] [CrossRef]
- Patel, S. Morphology and pharmacology of Passiflora edulis: A review. J. Herb. Med. Toxicol. 2009, 3, 1–6. [Google Scholar]
- Vanderplank, J. Passion Flowers, 3rd ed.; MIT Press: Cambridge, MA, USA, 2000; ISBN 978-0-262-72035-9. [Google Scholar]
- Kishore, K.; Pathak, K.A.; Shukla, R.; Bharali, R. Studies on floral biology of passion fruit (Passiflora spp.). Pak. J. Bot. 2010, 42, 21–29. [Google Scholar]
- Das, M.R.; Hossain, T.; Mia, M.A.B.; Ahmed, J.U.; Kariman, A.J.M.S.; Hossain, M.M. Fruit setting behaviour of passion fruit. Am. J. Plant Sci. 2013, 4, 1066–1073. [Google Scholar] [CrossRef]
- de Souza, M.M.; Pereira, T.N.S.; Hoffmann, M.; de Melo, E.J.T.; Louro, R.P. Embryo sac development in yellow passion fruit Passiflora edulis f. flavicarpa (Passifloraceae). Genet. Mol. Biol. 2002, 25, 471–475. [Google Scholar] [CrossRef][Green Version]
- Ickert-Bond, S.M.; Gerrath, J.; Wen, J. Gynoecial structure of Vitales and implications for the evolution of placentation in the Rosids. Int. J. Plant Sci. 2014, 175, 998–1032. [Google Scholar] [CrossRef]
- Endress, P.K. Angiosperm ovules: Diversity, development, evolution. Ann. Bot. 2011, 107, 1465–1489. [Google Scholar] [CrossRef]
- Ferrandiz, C. Fruit structure and diversity. In Encyclopedia of Life Sciences; John Wiley & Sons Ltd.: Chichester, UK, 2011; ISBN 978-0-470-01590-2. [Google Scholar]
- Faleiro, F.G.; Junqueira, N.T.V.; Junghans, T.G.; de Jesus, O.N.; Miranda, D.; Otoni, W.C. Advances in passion fruit (Passiflora spp.) Propagation. Rev. Bras. Frutic. 2019, 41, e155. [Google Scholar] [CrossRef]
- Oliveira, I.S.d.S.; de Souza, V.C.; Santos, S.B.d.C.; Almeida, J.N.d.R.; Henrique, J.d.S.; Costa, P.M.d.A.; Silva, J.H.C.S.; Cassimiro, C.A.L.; de Lucena, C.É.A.; Melo, T.d.S. Floral biology and pollen viability of Passiflora edulis Sims. J. Exp. Agric. Int. 2019, 41, 1–9. [Google Scholar] [CrossRef][Green Version]
- Hopping, M.E. Structure and development of fruit and seeds in Chinese gooseberry (Actinidia chinensis Planch.). N. Z. J. Bot. 1976, 14, 63–68. [Google Scholar] [CrossRef]
- Silva, K.R.; Stützel, T.; Oriani, A. Seed development and its relationship to fruit structure in species of Bromelioideae (Bromeliaceae) with fleshy fruits. Bot. J. Linn. Soc. 2020, 192, 868–886. [Google Scholar] [CrossRef]
- Dauelsberg, P.; Matus, J.T.; Poupin, M.J.; Leiva-Ampuero, A.; Godoy, F.; Vega, A.; Arce-Johnson, P. Effect of pollination and fertilization on the expression of genes related to floral transition, hormone synthesis and berry development in grapevine. J. Plant Physiol. 2011, 168, 1667–1674. [Google Scholar] [CrossRef] [PubMed]
- Kühn, N.; Arce-Johnson, P. Pollination: A key event controlling the expression of genes related to phytohormone biosynthesis during grapevine berry formation. Plant Signal. Behav. 2012, 7, 7–11. [Google Scholar] [CrossRef]
- Shinozaki, Y.; Beauvoit, B.P.; Takahara, M.; Hao, S.; Ezura, K.; Andrieu, M.-H.; Nishida, K.; Mori, K.; Suzuki, Y.; Kuhara, S.; et al. Fruit setting rewires central metabolism via gibberellin cascades. Proc. Natl. Acad. Sci. USA 2020, 117, 23970–23981. [Google Scholar] [CrossRef]
- Li, B.-F.; Yu, S.-X.; Hu, L.-Q.; Zhang, Y.-J.; Zhai, N.; Xu, L.; Lin, W.-H. Simple culture methods and treatment to study hormonal regulation of ovule development. Front. Plant Sci. 2018, 9, 784. [Google Scholar] [CrossRef]
- Cucinotta, M.; Di Marzo, M.; Guazzotti, A.; de Folter, S.; Kater, M.M.; Colombo, L. Gynoecium size and ovule number are interconnected traits that impact seed yield. J. Exp. Bot. 2020, 71, 2479–2489. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, N.; Del Carpio, D.P.; Hofmann, A.; Mizuta, Y.; Kurihara, D.; Higashiyama, T.; Uchida, N.; Torii, K.U.; Colombo, L.; Groth, G.; et al. A peptide pair coordinates regular ovule initiation patterns with seed number and fruit size. Curr. Biol. 2020, 30, 4352–4361.e4. [Google Scholar] [CrossRef] [PubMed]
- Varga, A.; Bruinsma, J. Roles of seeds and auxins in tomato fruit growth. Z. Für Pflanzenphysiol. 1976, 80, 95–104. [Google Scholar] [CrossRef]
- Doi, K.; Nozaki, R.; Takahashi, K.; Iwasaki, N. Effects of the number of seeds per berry on fruit growth characteristics, especially on the duration of stage II in blueberry. Plants 2018, 7, 96. [Google Scholar] [CrossRef]
- Marcelis, L.F.M.; Baan Hofman-Eijer, L.R. Effects of seed number on competition and dominance among fruits in Capsicum annuum L. Ann. Bot. 1997, 79, 687–693. [Google Scholar] [CrossRef]
- Stephenson, A.G.; Devlin, B.; Horton, J.B. The effects of seed number and prior fruit dominance on the pattern of fruit production in Cucurbits pepo (Zucchini squash). Ann. Bot. 1988, 62, 653–661. [Google Scholar] [CrossRef]
- Vidal, M.d.G.; de Jong, D.; Wien, H.C.; Morse, R.A. Pollination and fruit set in pumpkin (Cucurbita pepo) by honey bees. Braz. J. Bot. 2010, 33, 106–113. [Google Scholar] [CrossRef]
- Sonkoly, J.; Vojtkó, E.A.; Tökölyi, J.; Török, P.; Sramkó, G.; Illyés, Z.; Molnár , V.A. Higher seed number compensates for lower fruit set in deceptive orchids. J. Ecol. 2016, 104, 343–351. [Google Scholar] [CrossRef]
- Chiarini, F.E.; Barboza, G.E. Placentation patterns and seed number in fruits of South American Solanum subgen. Leptostemonum (Solanaceae) species. Darwiniana 2007, 45, 163–174. [Google Scholar] [CrossRef]
- Rajan, S.; Yadava, L.P.; Kumar, R.; Saxena, S.K. Direct and indirect effects of seed related characters on number of seed in guava (Psidium guajava L.) Fruits. Sci. Hortic. 2008, 116, 47–51. [Google Scholar] [CrossRef]
- López-Palacios, C.; Peña-Valdivia, C.B.; Reyes-Agüero, J.A.; Aguirre-Rivera, J.R.; Ramírez-Tobías, H.M.; Soto-Hernández, R.M.; Jiménez-Bremont, J.F. Inter- and intra-specific vriation in fruit biomass, number of seeds, and physical characteristics of seeds in Opuntia spp., Cactaceae. Genet. Resour. Crop Evol. 2015, 62, 1205–1223. [Google Scholar] [CrossRef]
- Boavida, L.C.; Vieira, A.M.; Becker, J.D.; Feijo, J.A. Gametophyte interaction and sexual reproduction: How plants make a zygote. Int. J. Dev. Biol. 2004, 49, 615–632. [Google Scholar] [CrossRef]
- Simpson, M.G. 9—Plant morphology. In Plant Systematics, 2nd ed.; Simpson, M.G., Ed.; Academic Press: San Diego, CA, USA, 2010; pp. 451–513. ISBN 978-0-12-374380-0. [Google Scholar]
- Amela García, M.T.; Miguez, M.B.; Gottsberger, G. Pollen: Ovule ratio and its relationship with other reproductive traits in some Passiflora species (Passifloraceae). An. Jardín Botánico Madr. 2014, 71, e009. [Google Scholar] [CrossRef]
- Martarello, N.S.; Gruchowski-Woitowicz, F.C.; Agostini, K. Pollinator efficacy in yellow passion fruit (Passiflora edulis f. flavicarpa Deg., Passifloraceae). Neotrop. Entomol. 2021, 50, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Nadeau, J.A.; Zhang, X.S.; Li, J.; O’Neill, S.D. Ovule development: Identification of stage-specific and tissue-specific cDNAs. Plant Cell 1996, 8, 213–239. [Google Scholar] [CrossRef] [PubMed]
- Bencivenga, S.; Colombo, L.; Masiero, S. Cross talk between the sporophyte and the megagametophyte during ovule development. Sex. Plant Reprod. 2011, 24, 113–121. [Google Scholar] [CrossRef]
- Lazarte, J.E.; Palser, B.F. Morphology, vascular anatomy and embryology of pistillate and staminate flowers of Asparagus officinalis. Am. J. Bot. 1979, 66, 753–764. [Google Scholar] [CrossRef]
- Pinto, S.C.; Mendes, M.A.; Coimbra, S.; Tucker, M.R. Revisiting the female germline and its expanding toolbox. Trends Plant Sci. 2019, 24, 455–467. [Google Scholar] [CrossRef]
- Robinson-Beers, K.; Pruitt, R.E.; Gasser, C.S. Ovule development in wild-type arabidopsis and two female-sterile mutants. Plant Cell 1992, 4, 1237–1249. [Google Scholar] [CrossRef]
- Pérez-Pastrana, J.; Islas-Flores, I.; Bárány, I.; Álvarez-López, D.; Canto-Flick, A.; Canto-Canché, B.; Peña-Yam, L.; Muñoz-Ramírez, L.; Avilés-Viñas, S.; Testillano, P.S.; et al. Development of the ovule and seed of habanero chili pepper (Capsicum chinense Jacq.): Anatomical characterization and immunocytochemical patterns of pectin methyl-esterification. J. Plant Physiol. 2018, 230, 1–12. [Google Scholar] [CrossRef]
- Brzezicka, E.; Kozieradzka-Kiszkurno, M. Female gametophyte development in Sedum sediforme (Jacq.) Pau (Crassulaceae): An anatomical, cytochemical and ultrastructural analysis. Protoplasma 2019, 256, 537–553. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Luo, Q.; Zhang, X.; Zhang, Q.; Cheng, Y. Identification of vital candidate microRNA/mRNA pairs regulating ovule development using high-throughput sequencing in hazel. BMC Dev. Biol. 2020, 20, 13. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Sun, J.; Guo, X.; Wang, K.; Liu, Q.; Liu, Q. Anther and ovule development of Clematis serratifolia (Ranunculaceae)–with new formation types in megaspore and nucellus. PLoS ONE 2020, 15, e0240432. [Google Scholar] [CrossRef]
- Xu, G.; Huang, J.; Lei, S.; Sun, X.; Li, X. Comparative gene expression profile analysis of ovules provides insights into Jatropha curcas L. ovule development. Sci. Rep. 2019, 9, 15973. [Google Scholar] [CrossRef]
- de Moura, S.M.; Rossi, M.L.; Artico, S.; Grossi-de-Sa, M.F.; Martinelli, A.P.; Alves-Ferreira, M. Characterization of floral morphoanatomy and identification of marker genes preferentially expressed during specific stages of cotton flower development. Planta 2020, 252, 71. [Google Scholar] [CrossRef] [PubMed]
- Halbritter, H.; Silvia, U.; Grímsson, F.; Weber, M.; Zetter, R.; Hesse, M.; Buchner, R.; Svojtka, M.; Frosch-Radivo, A. Illustrated Pollen Terminology; Springer International Publishing: Cham, Switzerland, 2018; ISBN 978-3-319-71364-9. [Google Scholar]
- Dokland, T.; Hutmacher, D.W.; Ng, M.M.-L.; Schantz, J.-T. Techniques in Microscopy for Biomedical Applications; Manuals in Biomedical Research; World Scientific: Singapore, 2006; Volume 2, ISBN 978-981-256-434-4. [Google Scholar]
- Fos, M.; Nuez, F.; Garcıa-Martınez, J.L. The gene Pat-2, which induces natural parthenocarpy, alters the gibberellin content in unpollinated tomato ovaries. Plant Physiol. 2000, 122, 471–480. [Google Scholar] [CrossRef]
- Moreno-Sanz, P.; D’Amato, E.; Nebish, A.; Costantini, L.; Grando, M.S. An optimized histological proceeding to study the female gametophyte development in grapevine. Plant Methods 2020, 16, 61. [Google Scholar] [CrossRef]
- Lynch, S.P.; Webster, G.L. A new technique of preparing pollen for scanning electron microscopy. Grana 1975, 15, 127–136. [Google Scholar] [CrossRef]
- Dustmann, J.H.; von der Ohe, K. Scanning electron microscopic studies on pollen from honey. IV. surface pattern of pollen of Sapium sebiferum and Euphorbia spp. (Euphorbiaceae). Apidologie 1993, 24, 59–66. [Google Scholar] [CrossRef][Green Version]
- Pathan, A.K.; Bond, J.; Gaskin, R.E. Sample preparation for SEM of plant surfaces. Mater. Today 2010, 12, 32–43. [Google Scholar] [CrossRef]
- Garcia, M.T.A.; Galati, B.G.; Anton, A.M. Microsporogenesis, microgametogenesis and pollen morphology of Passiflora spp. (Passifloraceae). Bot. J. Linn. Soc. 2002, 139, 383–394. [Google Scholar] [CrossRef][Green Version]
- Kim, H.N. Comparative study of immune-enhancing activity of crude and mannoprotein-free yeast-glucan preparations. J. Microbiol. Biotechnol. 1999, 9, 826–831. [Google Scholar]
- Hartig, S.M. Basic image analysis and manipulation in ImageJ. Curr. Protoc. Mol. Biol. 2013, 102, 14.15.1–14.15.12. [Google Scholar] [CrossRef]
- Mochizuki, J.; Itagaki, T.; Blue, Y.A.; Ito, M.; Sakai, S. Ovule and seed production patterns in relation to flower size variations in actinomorphic and zygomorphic flower species. AoB Plants 2019, 11, 1–8. [Google Scholar] [CrossRef]
- Radhamani, T.R.; Sudarshana, L.; Krishnan, R. Defense and carnivory: Dual role of bracts in Passiflora foetida. J. Biosci. 1995, 20, 657–664. [Google Scholar] [CrossRef]
- Acevedo-Rodríguez, P. Vines and Climbing Plants of Puerto Rico and the Virgin Islands; Department of Botany, National Museum of Natural History: Washington, DC, USA, 2005; Volume 51. [Google Scholar]
- Colombo, L.; Battaglia, R.; Kater, M.M. Arabidopsis ovule development and its evolutionary conservation. Trends Plant Sci. 2008, 13, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Gaiser, J.C.; Robinson-Beers, K.; Gasser, C.S. The Arabidopsis SUPERMAN gene mediates asymmetric growth of the outer integument of ovules. Plant Cell 1995, 7, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Gillaspy, G.; Ben-David, H.; Gruissem, W. Fruits: A developmental perspective. Plant Cell 1993, 5, 1439–1451. [Google Scholar] [CrossRef] [PubMed]
- Padhye, M.D.; Deshpande, B.G. The male and female gametophytes of Passiflora foetida Linn. Proc. Indian Acad. Sci. Sect. B 1960, 52, 124–130. [Google Scholar] [CrossRef]
- Drews, G.N.; Koltunow, A.M.G. The female gametophyte. Arab. Book 2011, 9, e0155. [Google Scholar] [CrossRef]
- Greenway, C.A.; Harder, L.D. Variation in ovule and seed size and associated size–number trade-offs in angiosperms. Am. J. Bot. 2007, 94, 840–846. [Google Scholar] [CrossRef]
- Colle, M.; Weng, Y.; Kang, Y.; Ophir, R.; Sherman, A.; Grumet, R. Variation in cucumber (Cucumis sativus L.) fruit size and shape results from multiple components acting pre-anthesis and post-pollination. Planta 2017, 246, 641–658. [Google Scholar] [CrossRef]
- Wetzstein, H.Y.; Yi, W.; Porter, J.A.; Ravid, N. Flower position and size impact ovule number per flower, fruitset, and fruit size in pomegranate. J. Am. Soc. Hortic. Sci. 2013, 138, 159–166. [Google Scholar] [CrossRef]
- Strelin, M.M.; Aizen, M.A. The interplay between ovule number, pollination and resources as determinants of seed set in a modular plant. PeerJ 2018, 6, e5384. [Google Scholar] [CrossRef]
- Singh, K.; Sharma, Y.P.; Sharma, P.R.; Gairola, S. Pollen morphology and variability of the Rosa L. species of Western Himalaya in India. Genet. Resour. Crop Evol. 2020, 67, 2129–2148. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vijay, A.; Nizam, A.; Radhakrishnan, A.M.; Anju, T.; Kashyap, A.K.; Kumar, N.; Kumar, A. Comparative Study of Ovule Development between Wild (Passiflora foetida L.) and Cultivated (P. edulis Sims) Species of Passiflora L. Provide Insights into Its Differential Developmental Patterns. J. Zool. Bot. Gard. 2021, 2, 502-516. https://doi.org/10.3390/jzbg2030036
Vijay A, Nizam A, Radhakrishnan AM, Anju T, Kashyap AK, Kumar N, Kumar A. Comparative Study of Ovule Development between Wild (Passiflora foetida L.) and Cultivated (P. edulis Sims) Species of Passiflora L. Provide Insights into Its Differential Developmental Patterns. Journal of Zoological and Botanical Gardens. 2021; 2(3):502-516. https://doi.org/10.3390/jzbg2030036
Chicago/Turabian StyleVijay, Archa, Ashifa Nizam, Arun Madasseril Radhakrishnan, Thattantavide Anju, Arun Kumar Kashyap, Nitesh Kumar, and Ajay Kumar. 2021. "Comparative Study of Ovule Development between Wild (Passiflora foetida L.) and Cultivated (P. edulis Sims) Species of Passiflora L. Provide Insights into Its Differential Developmental Patterns" Journal of Zoological and Botanical Gardens 2, no. 3: 502-516. https://doi.org/10.3390/jzbg2030036
APA StyleVijay, A., Nizam, A., Radhakrishnan, A. M., Anju, T., Kashyap, A. K., Kumar, N., & Kumar, A. (2021). Comparative Study of Ovule Development between Wild (Passiflora foetida L.) and Cultivated (P. edulis Sims) Species of Passiflora L. Provide Insights into Its Differential Developmental Patterns. Journal of Zoological and Botanical Gardens, 2(3), 502-516. https://doi.org/10.3390/jzbg2030036