
Histone Deacetylase 3 Regulates Adipocyte Phenotype at Early Stages of Differentiation

 , , and
, , and 
Abstract
:1. Introduction
2. Results
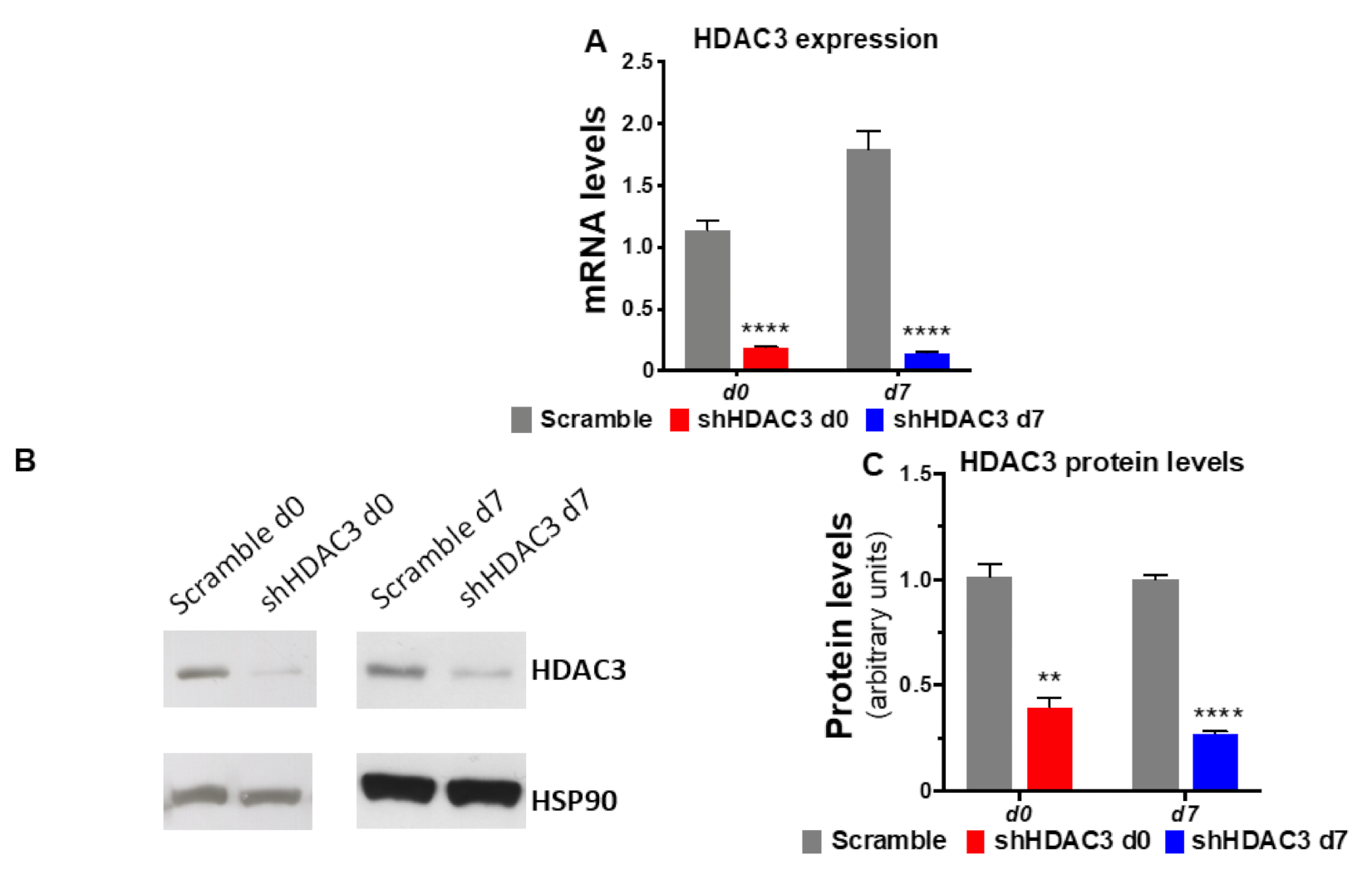
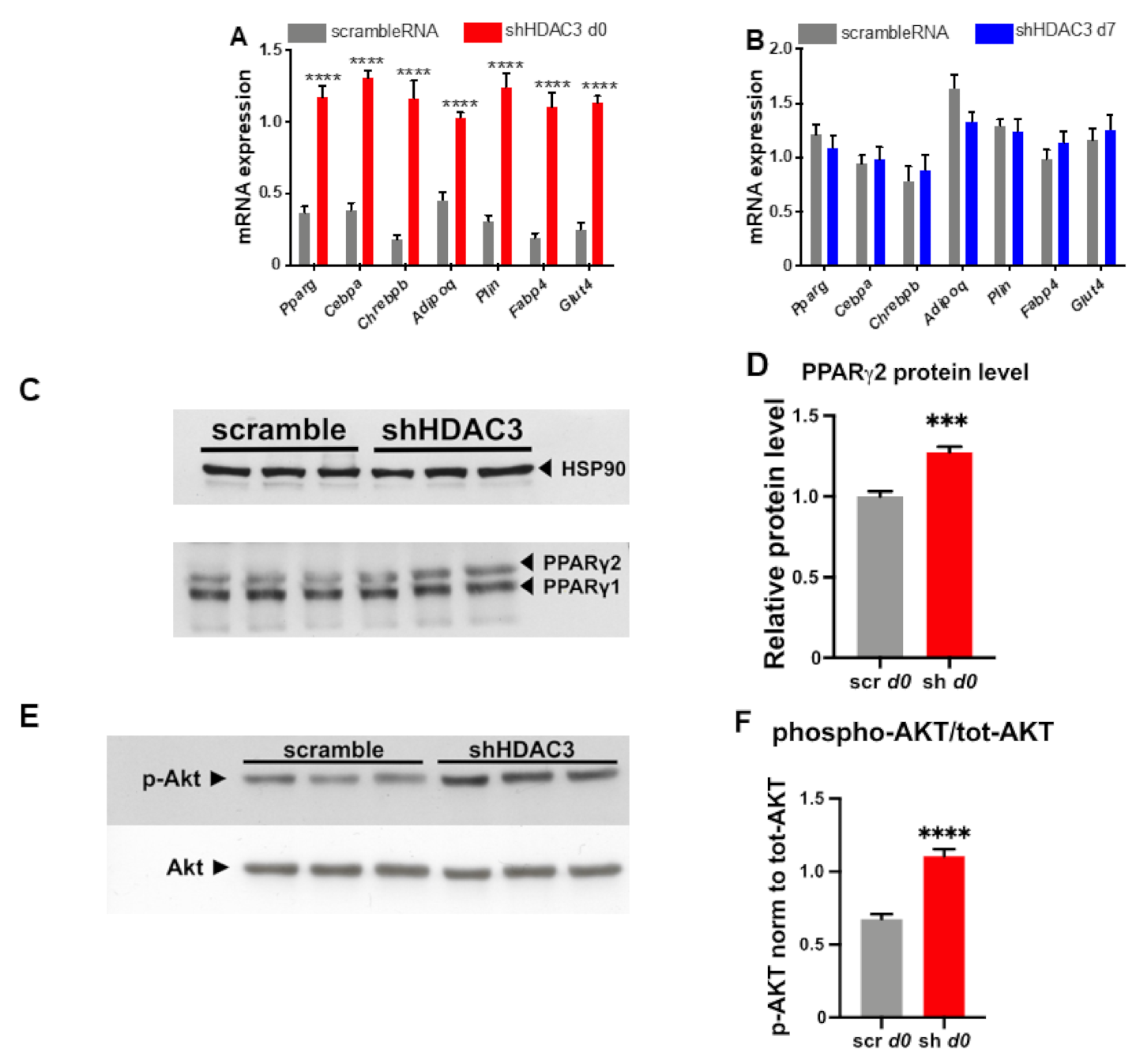
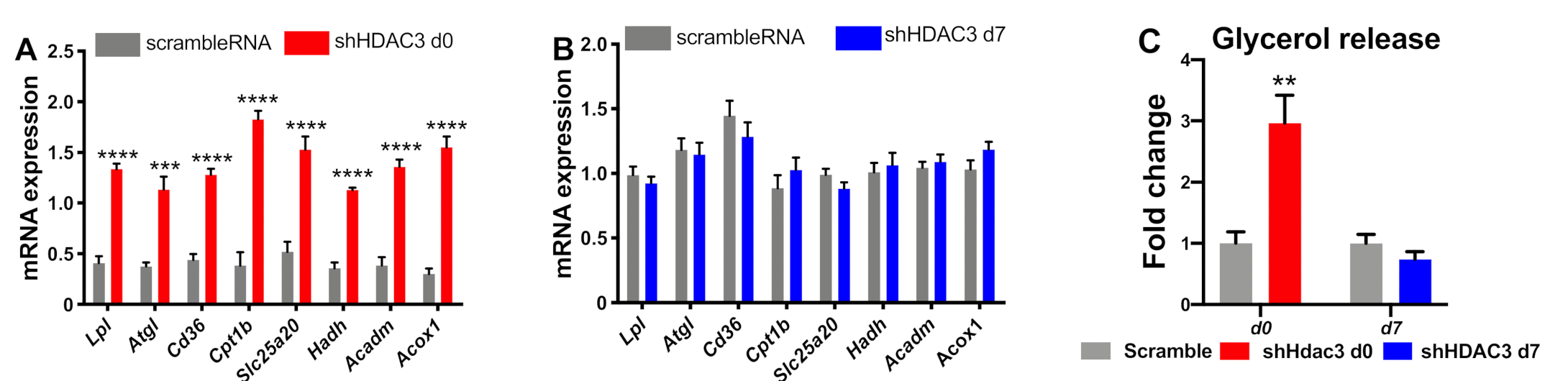
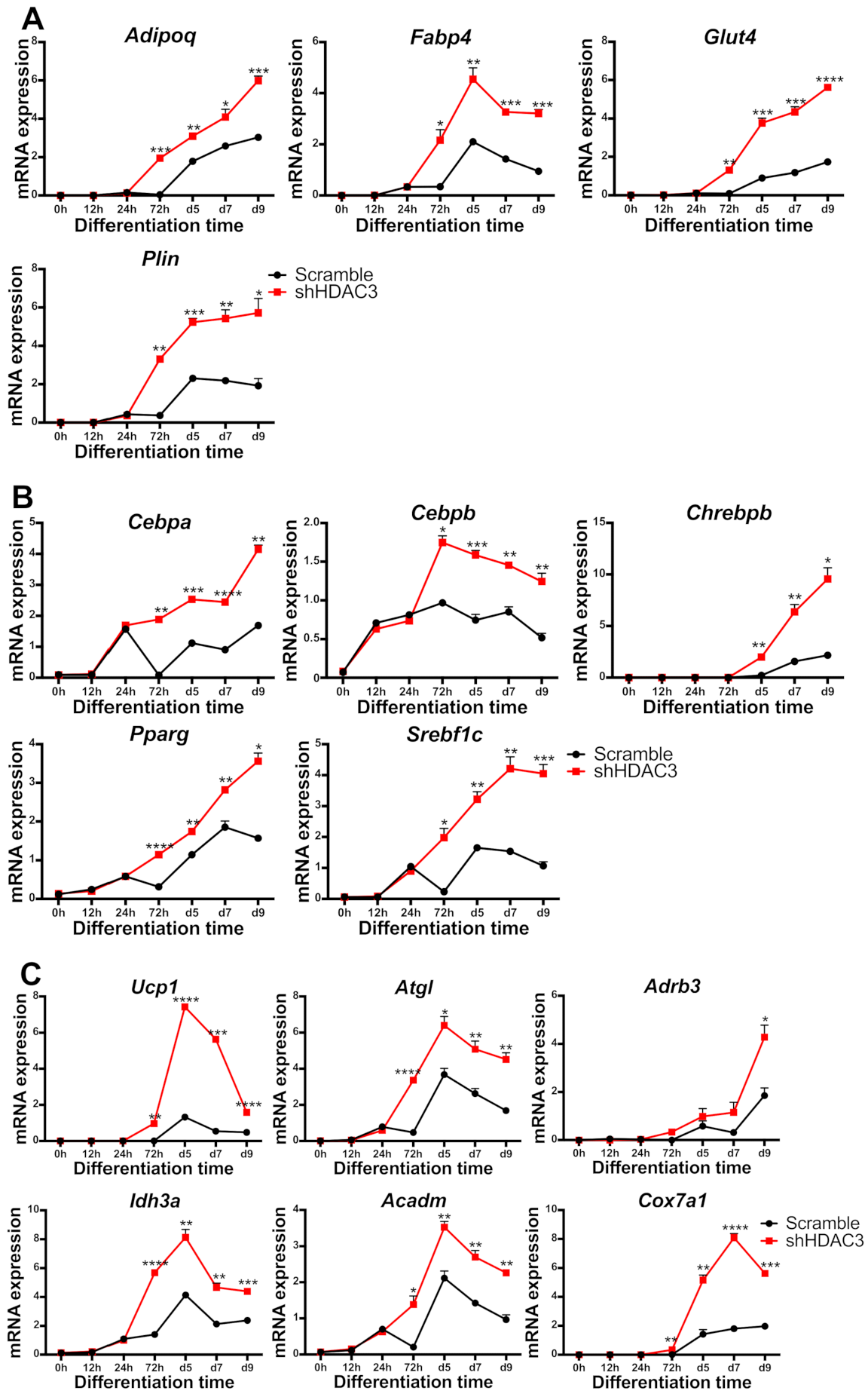
2.1. Early Silencing of HDAC3 Amplifies Adipocyte Differentiation and Functionality
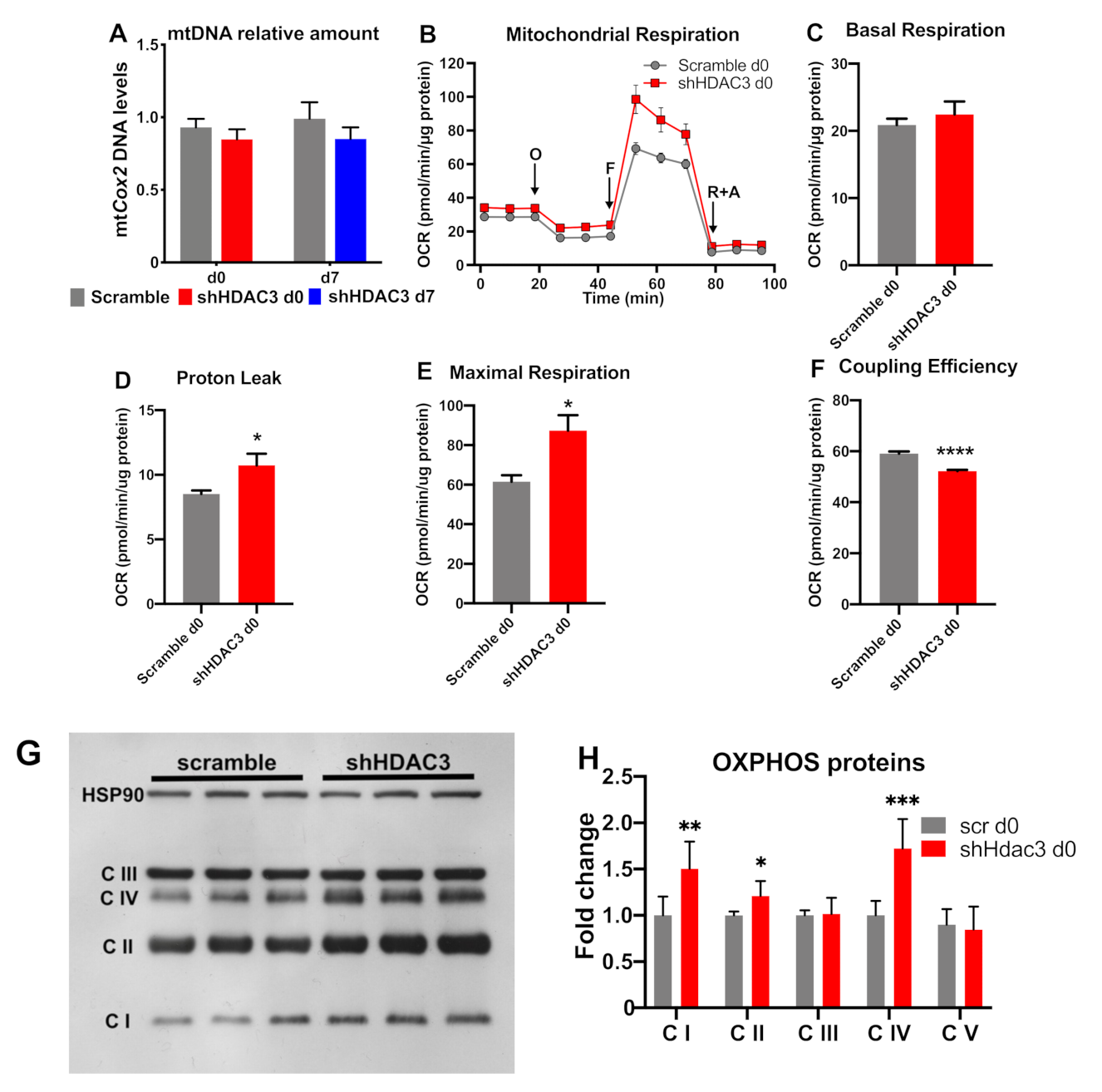
2.2. Early Silencing of HDAC3 Promotes Oxidative Metabolism of Mature Adipocytes
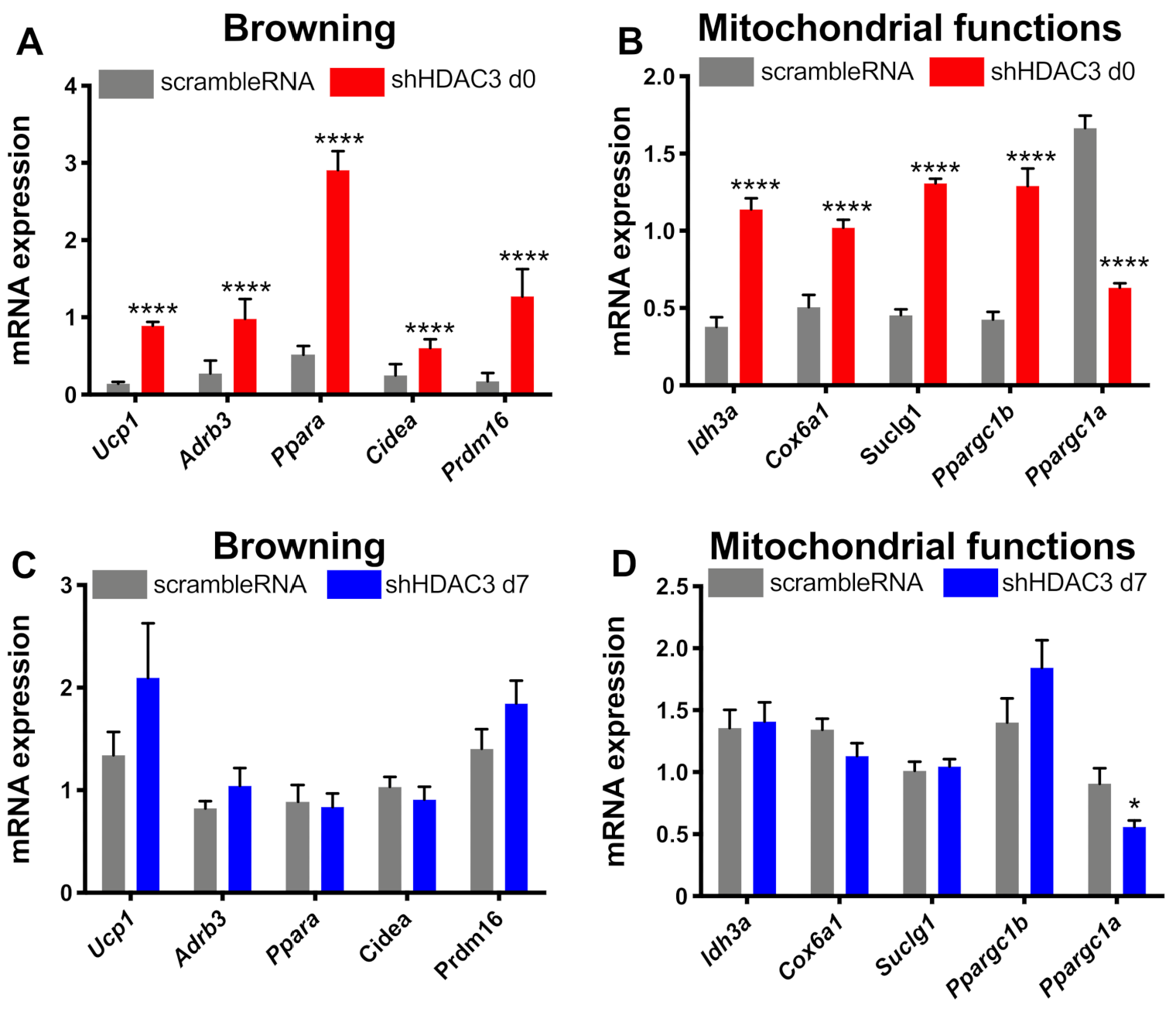
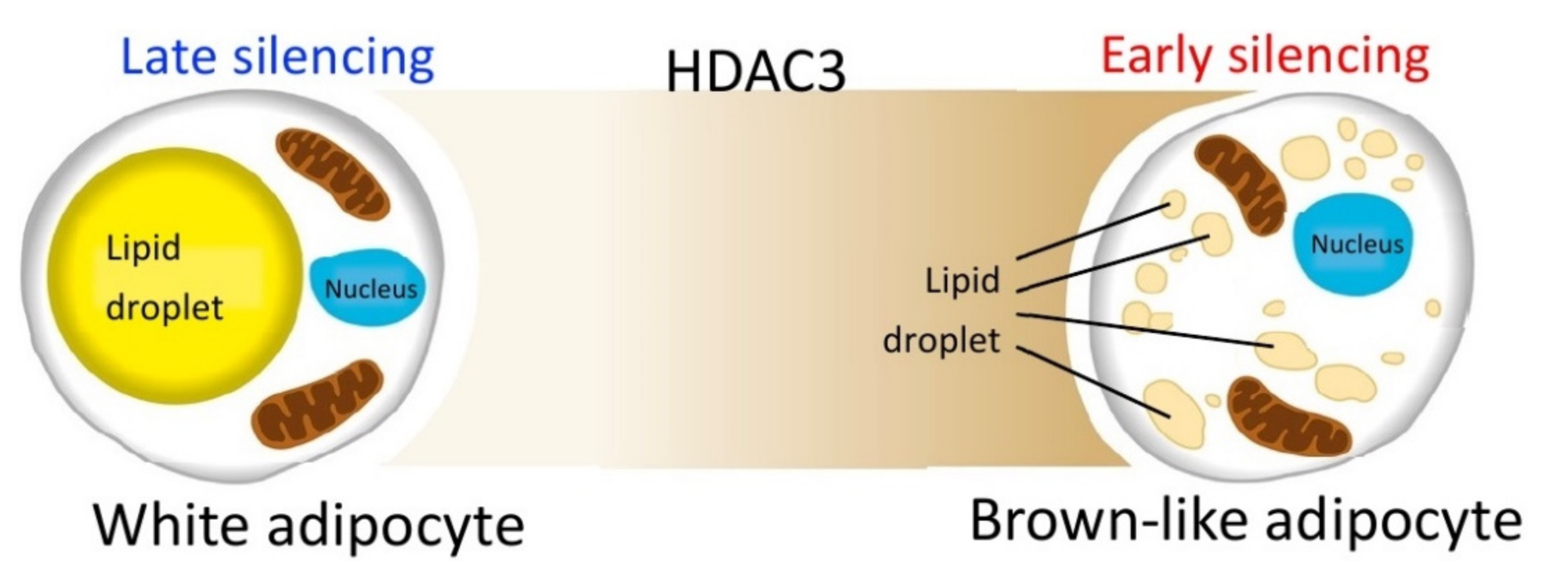
2.3. Early Silencing of HDAC3 Stimulates Adipocyte Browning and Mithocondria Functionality
2.4. Early Silencing of HDAC3 Modulates Adipocyte Pro-Inflammatory Profile
2.5. HDAC3 Silencing in Precursor Cells Affects Gene Expression from Initial Stages of Adipocyte Differentiation
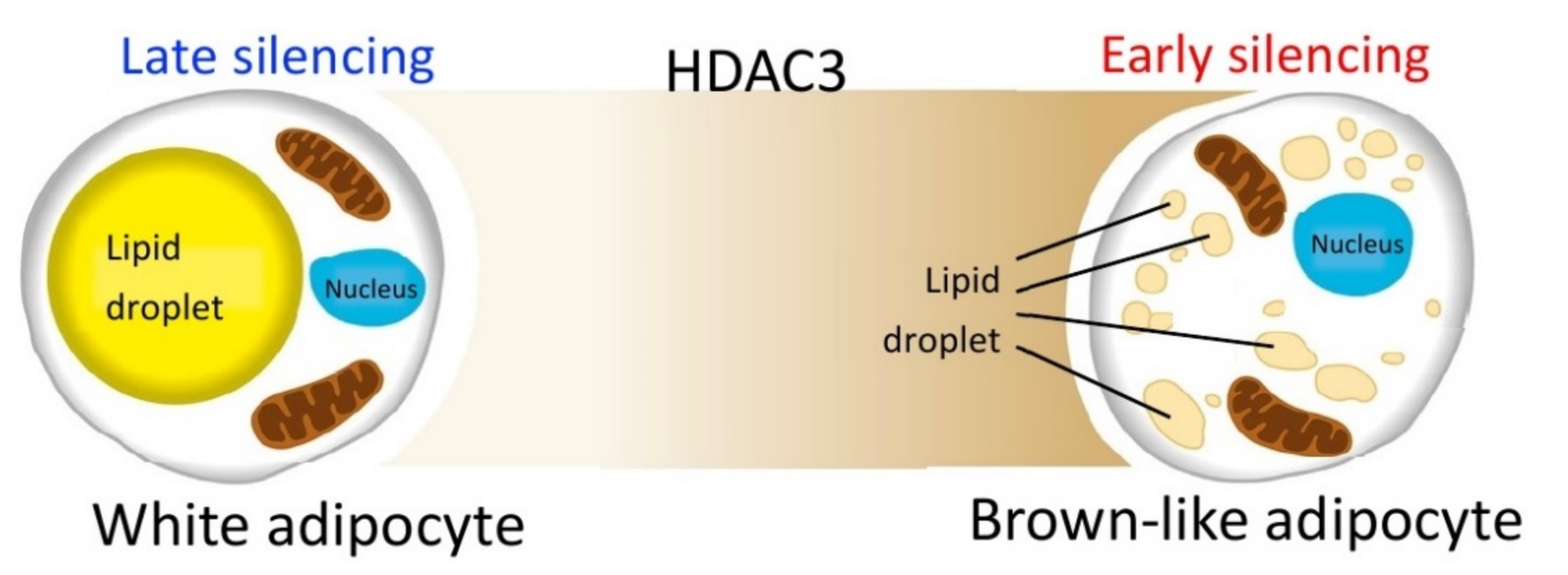
3. Discussion
4. Materials and Methods
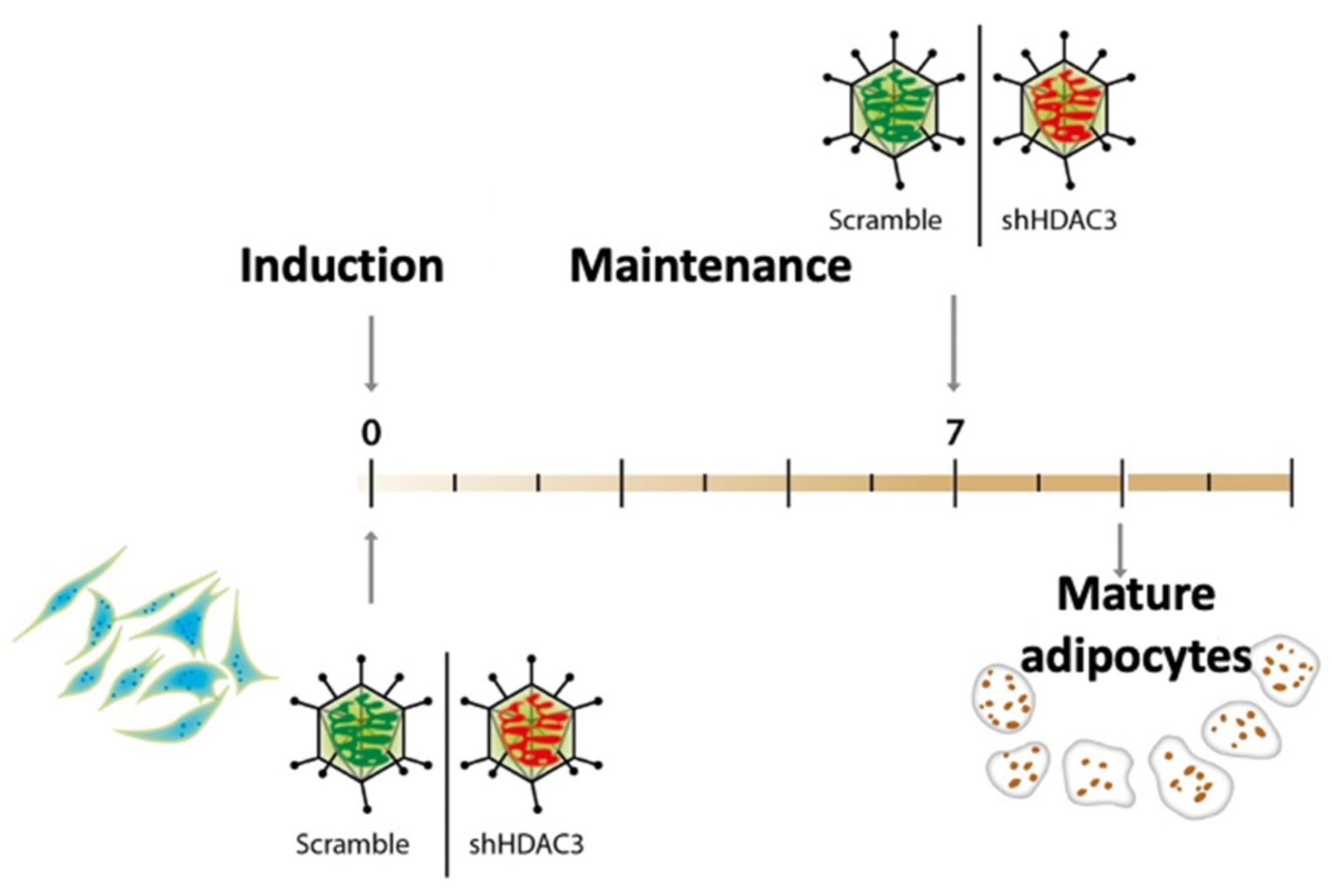
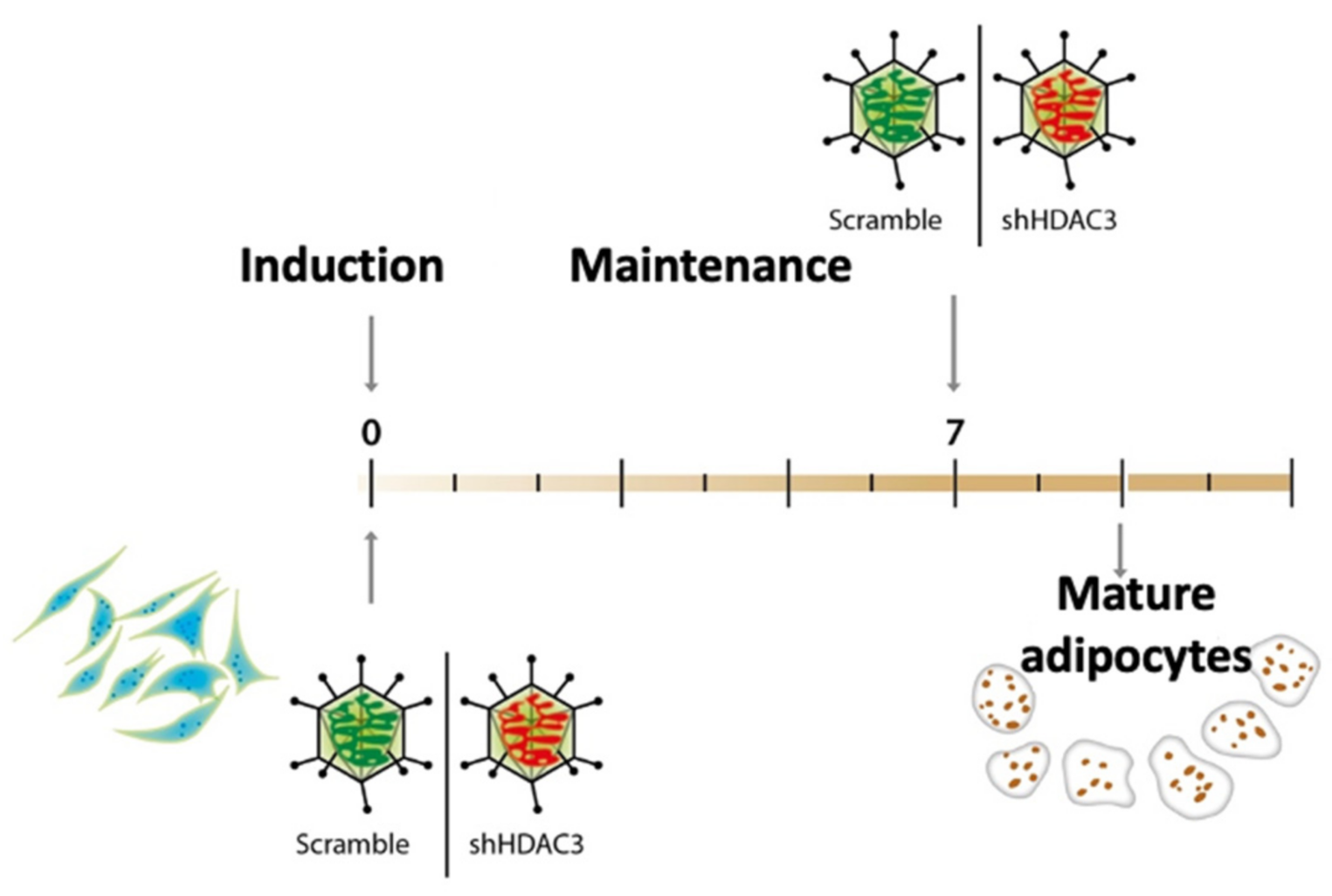
4.1. Cell Culture and Differentiation
4.2. Cell Infection
4.3. Gene Expression
4.4. Lipid Content in Adipocytes
4.5. mtDNA Quantification
4.6. Glycerol Release from Adipocytes
4.7. Measurement of Mitochondrial Respiration
4.8. Protein Analyses and Quantification
4.9. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| HDACs | Histone deacetylases |
| HDAC3 | Histone deacetylase 3 |
| MSCs | Mesenchymal stem cells |
| BMI | Body mass index |
| CREB | cAMP response element binding protein |
| C/EBP | CCAAT/enhancer-binding protein |
| SREBP-1c | Sterol regulatory element binding protein-1c |
| PPARG | Peroxisome proliferator-activated receptor gamma |
| WAT | White adipose tissue |
| BAT | Brown adipose tissue |
| UCP1 | Uncoupling protein 1 |
| ETC | Electron transport chain |
| FCCP | carbonyl cyanide-4-(trifluoromethoxy)phenylhydrazone |
| ChREBPB | Carbohydrate-responsive element-binding protein |
| PLIN | Perilipin |
| FABP4 | Fatty acid binding protein 4 |
| ADIPOQ | Adiponectin |
| GLUT4 | Glucose transporter 4 |
| CD36 | Cluster of differentiation 36 |
| LPL | Lipoprotein lipase |
| ATGL | Adipose triglyceride lipase |
| CPT1B | Carnitine palmitoyl-transferase 1b |
| SLC25A20 | Solute carrier family 25 member 20 |
| HDAH | Hydroxyacyl-coenzyme A dehydrogenase |
| ACADM | Medium-chain acyl-CoA dehydrogenase |
| ACOX1 | Peroxisomal acyl-coenzyme A oxidase 1 |
| ADRB3 | β3 adrenergic receptor |
| PPARA | Peroxisome proliferator activator receptor α |
| CIDEA | Cell death inducing DFFA-like effector α |
| PRDM16 | PR domain containing 16 |
| IDH3A | Isocitrate dehydrogenase subunit α |
| mtCOX2 | Mitochondrial cytochrome c oxidase subunit 2 |
| COX6A1 | Cytochrome c oxidase subunit 6a1 |
| COX7A1 | Cytochrome c oxidase subunit 7a1 |
| SUCLG1 | Succinyl-CoA ligase subunit 1 |
| PPARGC1A | PPARγ coactivator 1α |
| PPARGC1B | PPARγ coactivator 1β |
| MCP1 | Monocyte chemoattractant protein-1 |
| IL6 | Interleukin 6 |
| COL6A1 | Collagen type 6a1 |
| iNOS | Inducible nitric oxide synthase |
| MEST | Mesoderm-specific transcript |
| H3K27 | Lysine 27 Histone 3 |
References
- Caballero, B. Humans against Obesity: Who Will Win? Adv. Nutr. 2019, 10, S4–S9. [Google Scholar] [CrossRef]
- WHO. Obesity and Overweight Key Facts. Available online: http://www.who.int/mediacentre/factsheets/fs311/en/ (accessed on 2 August 2021).
- Cercato, C.; Fonseca, F.A. Cardiovascular risk and obesity. Diabetol. Metab. Syndr. 2019, 11, 74. [Google Scholar] [CrossRef] [PubMed]
- Guilherme, A.; Virbasius, J.V.; Puri, V.; Czech, M.P. Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Anandacoomarasamy, A.; Caterson, I.; Sambrook, P.; Fransen, M.; March, L. The impact of obesity on the musculoskeletal system. Int. J. Obes. 2008, 32, 211–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Morley, T.S.; Kim, M.; Clegg, D.J.; Scherer, P.E. Obesity and cancer—Mechanisms underlying tumour progression and recurrence. Nat. Rev. Endocrinol. 2014, 10, 455–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansari, S.; Haboubi, H.; Haboubi, N. Adult obesity complications: Challenges and clinical impact. Adv. Endocrinal. Metab. 2020, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Russo, L.; Lumeng, C.N. Properties and functions of adipose tissue macrophages in obesity. Immunology 2018, 155, 407–417. [Google Scholar] [CrossRef]
- Zatterale, F.; Longo, M.; Naderi, J.; Raciti, G.A.; Desiderio, A.; Miele, C.; Beguinot, F. Chronic Adipose Tissue Inflammation Linking Obesity to Insulin Resistance and Type 2 Diabetes. Front. Physiol. 2020, 10, 1607. [Google Scholar] [CrossRef] [PubMed]
- Wernstedt Asterholm, I.; Tao, C.; Morley, T.S.; Wang, Q.A.; Delgado-Lopez, F.; Wang, Z.V.; Scherer, P.E. Adipocyte Inflammation Is Essential for Healthy Adipose Tissue Expansion and Remodeling. Cell Metab. 2014, 20, 103–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.A.; Tao, C.; Gupta, R.K.; Scherer, P.E. Tracking adipogenesis during white adipose tissue development, expansion and regeneration. Nat. Med. 2013, 19, 1338–1344. [Google Scholar] [CrossRef]
- Muller, S.; Ader, I.; Creff, J.; Leménager, H.; Achard, P.; Casteilla, L.; Sensebé, L.; Carrière, A.; Deschaseaux, F. Human adipose stromal-vascular fraction self-organizes to form vascularized adipose tissue in 3D cultures. Sci. Rep. 2019, 9, 7250. [Google Scholar] [CrossRef]
- Arner, P.; Bernard, S.; Salehpour, M.; Possnert, G.; Liebl, J.; Steier, P. Dynamics of human adipose lipid turnover in health and metabolic disease. Nature 2011, 478, 110–113. [Google Scholar] [CrossRef] [Green Version]
- Siersbæk, R.; Rabiee, A.; Nielsen, R.; Sidoli, S.; Traynor, S.; Loft, A.; Poulsen, L.L.C.; Rogowska-Wrzesinska, A.; Jensen, O.N.; Mandrup, S. Transcription factor cooperativity in early adipogenic hotspots and super-enhancers. Cell Rep. 2014, 7, 1443–1455. [Google Scholar] [CrossRef] [Green Version]
- Siersbæk, R.; Mandrup, S. Transcriptional networks controlling adipocyte differentiation. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 247–255. [Google Scholar] [CrossRef]
- Lefterova, M.I.; Haakonsson, A.K.; Lazar, M.A.; Mandrup, S. PPARγ and the global map of adipogenesis and beyond. Trends Endocrinol. Metab. 2014, 25, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Giralt, M.; Villarroya, F. White, brown, beige/brite: Different adipose cells for different functions? Endocrinology 2013, 154, 2992–3000. [Google Scholar] [CrossRef] [Green Version]
- Sidossis, L.; Kajimura, S. Brown and beige fat in humans: Thermogenic adipocytes that control energy and glucose homeostasis. J. Clin. Investig. 2015, 125, 478–486. [Google Scholar] [CrossRef]
- Kuryłowicz, A.; Puzianowska-Kuźnicka, M. Induction of adipose tissue browning as a strategy to combat obesity. Int. J. Mol. Sci. 2020, 21, 6241. [Google Scholar] [CrossRef]
- Trayhurn, P.; Beattie, J.H. Physiological role of adipose tissue: White adipose tissue as an endocrine and secretory organ. Proc. Nutr. Soc. 2001, 60, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Park, A. Distinction of white, beige and brown adipocytes derived from mesenchymal stem cells. World J. Stem Cells 2014, 6, 33. [Google Scholar] [CrossRef]
- Inagaki, T.; Sakai, J.; Kajimura, S. Transcriptional and epigenetic control of brown and beige adipose cell fate and function. Nat. Rev. Mol. Cell Biol. 2016, 17, 480–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donohoe, D.R.; Bultman, S.J. Metaboloepigenetics: Interrelationships between energy metabolism and epigenetic control of gene expression. J. Cell Physiol. 2012, 227, 3169–3177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, J.A.; Milagro, F.I.; Claycombe, K.J.; Schalinske, K.L. Epigenetics in Adipose Tissue, Obesity, Weight Loss, and Diabetes. Adv. Nutr. 2014, 5, 71–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohde, K.; Keller, M.; la Cour Poulsen, L.; Blüher, M.; Kovacs, P.; Böttcher, Y. Genetics and epigenetics in obesity. Metabolism 2019, 92, 37–50. [Google Scholar] [CrossRef] [Green Version]
- Ling, C.; Rönn, T. Epigenetics in Human Obesity and Type 2 Diabetes. Cell Metab. 2019, 29, 1028–1044. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, A.; Fiorino, E.; Giudici, M.; Gilardi, F.; Galmozzi, A.; Mitro, N.; Cermenati, G.; Godio, C.; Caruso, D.; De Fabiani, E.; et al. Linking epigenetics to lipid metabolism: Focus on histone deacetylases. Mol. Membr. Biol. 2012, 29, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Galmozzi, A.; Mitro, N.; Ferrari, A.; Gers, E.; Gilardi, F.; Godio, C.; Cermenati, G.; Gualerzi, A.; Donetti, E.; Rotili, D.; et al. Inhibition of class I histone deacetylases unveils a mitochondrial signature and enhances oxidative metabolism in skeletal muscle and adipose tissue. Diabetes 2013, 62, 732–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrari, A.; Fiorino, E.; Longo, R.; Barilla, S.; Mitro, N.; Cermenati, G.; Giudici, M.; Caruso, D.; Mai, A.; Guerrini, U.; et al. Attenuation of diet-induced obesity and induction of white fat browning with a chemical inhibitor of histone deacetylases. Int. J. Obes. 2017, 41, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, A.; Longo, R.; Peri, C.; Coppi, L.; Caruso, D.; Mai, A.; Mitro, N.; De Fabiani, E.; Crestani, M. Inhibition of class I HDACs imprints adipogenesis toward oxidative and brown-like phenotype. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2020, 1865, 158594. [Google Scholar] [CrossRef]
- Ferrari, A.; Longo, R.; Fiorino, E.; Silva, R.; Mitro, N.; Cermenati, G.; Gilardi, F.; Desvergne, B.; Andolfo, A.; Magagnotti, C.; et al. HDAC3 is a molecular brake of the metabolic switch supporting white adipose tissue browning. Nat. Commun. 2017, 8, 93. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.C.B.; Adlanmerini, M.; Hauck, A.K.; Lazar, M.A. Dichotomous engagement of HDAC3 activity governs inflammatory responses. Nature 2020, 584, 286–290. [Google Scholar] [CrossRef]
- Herman, M.A.; Peroni, O.D.; Villoria, J.; Schön, M.R.; Abumrad, N.A.; Blüher, M.; Klein, S.; Kahn, B.B. A novel ChREBP isoform in adipose tissue regulates systemic glucose metabolism. Nature 2012, 484, 333–338. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; An, Y.A.; Kim, M.; Zhang, Z.; Zhao, S.; Zhu, Y.; Asterholm, I.W.; Kusminski, C.M.; Scherer, P.E. Suppressing Adipocyte Inflammation Promotes Insulin Resistance in Mice. Mol. Metab. 2020, 39, 101010. [Google Scholar] [CrossRef]
- Pasarica, M.; Gowronska-Kozak, B.; Burk, D.; Remedios, I.; Hymel, D.; Gimble, J.; Ravussin, E.; Bray, G.A.; Smith, S.R. Adipose tissue collagen VI in obesity. J. Clin. Endocrinol. Metab. 2009, 94, 5155–5162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anunciado-Koza, R.P.; Manuel, J.; Mynatt, R.L.; Zhang, J.; Kozak, L.P.; Koza, R.A. Diet-induced adipose tissue expansion is mitigated in mice with a targeted inactivation of mesoderm specific transcript (Mest). PLoS ONE 2017, 12, e0179879. [Google Scholar] [CrossRef] [Green Version]
- Uldry, M.; Yang, W.; St-Pierre, J.; Lin, J.; Seale, P.; Spiegelman, B.M. Complementary action of the PGC-1 coactivators in mitochondrial biogenesis and brown fat differentiation. Cell Metab. 2006, 3, 333–341. [Google Scholar] [CrossRef] [Green Version]
- Enguix, N.; Pardo, R.; González, A.; López, V.M.; Simó, R.; Kralli, A.; Villena, J.A. Mice lacking PGC-1β in adipose tissues reveal a dissociation between mitochondrial dysfunction and insulin resistance. Mol. Metab. 2013, 2, 215–226. [Google Scholar] [CrossRef]
- Nikonova, L.; Koza, R.A.; Mendoza, T.; Chao, P.-M.; Curley, J.P.; Kozak, L.P. Mesoderm-specific transcript is associated with fat mass expansion in response to a positive energy balance. FASEB J. 2008, 22, 3925–3937. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | 5′→3′ Sequence | |
|---|---|---|
| Hdac3 | Forward | TGTCTCAATGTGCCCTTACG |
| Reverse | CCTAATCGATCACAGCCCAG | |
| Probe | ACTTCTACCAGCCGACGTGCATC | |
| Pparg | Forward | TGTTATGGGTGAAACTCTGGG |
| Reverse | AGAGCTGATTCCGAAGTTGG | |
| Probe | CCCTCGCTGATGCACTGCCTATGA | |
| Cebpa | Forward | AGAGCCGAGATAAAGCCAAAC |
| Reverse | TCATTGTCACTGGTCAACTCC | |
| Probe | AGCACCTTCTGTTGCGTCTCCA | |
| Cebpb | Forward | CCCCGCGTTCATGCA |
| Reverse | CAGTCGGGCTCGTAGT | |
| Probe | ACTTCCATGGGTCTAAAGGCG | |
| Chrebpb | Forward | CTGCAGATCGCGTGGAG |
| Reverse | GCAACTTGAGGCCTTTGAAG | |
| Probe | CAAGCTGGTCTCTCCCAAGTGGAA | |
| Srebf1c | Forward | ATGGATTGCACATTTGAAGACATGCT |
| Reverse | CCTGTGTCCCCTGTCTCAC | |
| Probe | CTTCCCGGGCCTGTTTGACGCCCCCTA | |
| Adipoq | Forward | AGGCATCCCAGGACATC |
| Reverse | CCTGTCATTCCAACATCTCC | |
| Probe | CCTTAGGACCAAGAAGACCTGCATCTC | |
| Plin | Forward | ACAGACACAGAGGGAGAGG |
| Reverse | AGTGTTCTGCACGGTGTG | |
| Probe | AGGAGGAAGAAGAGTCCGAGGCT | |
| Fabp4 | Forward | GGCGTGGAATTCGATGAA |
| Reverse | GCTTGTCACCATCTCGTT | |
| Probe | TGATGCTCTTCACCTTCCTGTCGT | |
| Glut4 | Forward | TGTCGCTGGTTTCTCCAACTG |
| Reverse | CCATACGATCCGCAACATACTG | |
| Probe | ACCTGTAACTTCATTGTCGGCATGGGTTT | |
| Ucp1 | Forward | GAGCTGGTAACATATGACCTC |
| Reverse | GAGCTGACAGTAAATGGCA | |
| Probe | ACAAAATACTGGCAGATGACGTCCC | |
| Ppara | Forward | ACGCATGTGAAGGCTGTAAG |
| Reverse | CACTTGTGAAAACGGCAGTAC | |
| Probe | CGGCTGAAGCTGGTGTACGACAA | |
| Cidea | Forward | CACGCATTTCATGATCTTGG |
| Reverse | CCTGTATAGGTCGAAGGTGA | |
| Probe | TTACTACCCGGTGTCCATTTCTGTCC | |
| Lpl | Forward | GCCATGACAAGTCTCTGAAG |
| Reverse | CTTTCAAACACCCAAACAAGG | |
| Probe | AGTCTGGCTGACACTGGACAAACA | |
| Atgl | Forward | TCGTGTTTCAGACGGAGA |
| Reverse | CACATAGCGCACCCCT | |
| Probe | TGCAGACATTGGCCTGGATGAG | |
| Cd36 | Forward | GCGACATGATTAATGGCACAG |
| Reverse | GATCCGAACACAGCGTAGATAG | |
| Probe | CAACAAAAGGTGGAAAGGAGGCTGC | |
| Cpt1b | Forward | GATGCAGTTCCAGAGAATCC |
| Reverse | CTTGTTCTTGCCAGAGCT | |
| Probe | TCTGCCCACTCTACCCTTCCTC | |
| Slc25a20 | Forward | TCTTTGGGTTTGGTCTGGG |
| Reverse | ATTTGATCCGTTCTCCAGGG | |
| Probe | TCTCCAGAGGATGAACTTAGCTACCCAC | |
| Hadh | Forward | TCTTGACTATGTTGGACTGGATAC |
| Reverse | AAGGACTGGGCTGAAATAAGG | |
| Probe | CTTGGACGGGTGGCATGAAATGG | |
| Acadm | Forward | ACCCAGATCCTAAAGTACCC |
| Reverse | CGAAAGCAATTCCTCTGGTG | |
| Probe | TGGCCCATGTTTAGTTCCTTTTTTCCAA | |
| Acox1 | Forward | TCACGTTTACCCCGGC |
| Reverse | CAAGTACGACACCATACCAC | |
| Probe | CATCAAGAACCTGGCCGTCTGC | |
| Ppargc1b | Forward | GGTGTTCGGTGAGATTGTAGAG |
| Reverse | GTGATAAAACCGTGCTTCTGG | |
| Probe | TCTTTTACTTCTCGTCAGCACCTGGC | |
| Ppargc1a | Forward | CATTTGATGCACTGACAGATGGA |
| Reverse | GTCAGGCATGGAGGAAGGAC | |
| Probe | CCGTGACCACTGACA ACGAGGCC | |
| Idh3a | Forward | ACGGAAGGAGAATACAGTGG |
| Reverse | GTACTCGAAGGCAAACTCTG | |
| Probe | ACCCCATCAACGATCACATGCTCA | |
| mtCox2 | Forward | TGGTGAACTACGACTGCT |
| Reverse | CTGGGATGGCATCAGTTT | |
| Probe | TGGCAGAACGACTCGGTTATCAACT | |
| Cox6a1 | Forward | GTTCGTTGCCTACCCTCAC |
| Reverse | TCTCTTTACTCATCTTCATAGCCG | |
| Probe | ACCATACCCTCTTCCACAACCCTCA | |
| Cox7a1 | Forward | TGTGGCAGAGAAGCAGAAG |
| Reverse | AGCCCAAGCAGTATAAGCAG | |
| Probe | CGACAATGACCTCCCAGTACACTTGA | |
| Suclg1 | Forward | AATGATCCAGCCACAGAAGG |
| Reverse | AGCAATGAAGGACACTACAGG | |
| Probe | AGCATAACTCAGGTCCAAAGGCCAA | |
| Mcp1 | Forward | CTCATGTACCCGCTGTATAG |
| Reverse | TTAGAGCCACGACCATACA | |
| Probe | ATACTGGATGCCGTCTATGTCG | |
| Il6 | Forward | ACGATAGTCAATTCCAGAAACC |
| Reverse | GTTGTCACCAGCATCAGT | |
| Probe | CTTGCAGAGAGGAACTTCATAGC | |
| Col6a1 | Forward | CTGGTGAAGGAGAACTATGCAG |
| Reverse | GTCTAGCAGGATGGTGATGTC | |
| Probe | CCGGGGAGGAGAATGTGATTGGA | |
| Mest | Forward | TTCTGTGTCCATCCCCATTC |
| Reverse | CTGTGGGTAGTGGCTAATGTG | |
| Probe | TGTACAGGAAAACGCTGCCGC | |
| Nos2 | Forward | CCCTCCTGATCTTGTGTTGGA |
| Reverse | CAACCCGAGCTCCTGGAA | |
| Probe | CCATGGAGCATCCCAAGTACGAGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cricrí, D.; Coppi, L.; Pedretti, S.; Mitro, N.; Caruso, D.; De Fabiani, E.; Crestani, M. Histone Deacetylase 3 Regulates Adipocyte Phenotype at Early Stages of Differentiation. Int. J. Mol. Sci. 2021, 22, 9300. https://doi.org/10.3390/ijms22179300
Cricrí D, Coppi L, Pedretti S, Mitro N, Caruso D, De Fabiani E, Crestani M. Histone Deacetylase 3 Regulates Adipocyte Phenotype at Early Stages of Differentiation. International Journal of Molecular Sciences. 2021; 22(17):9300. https://doi.org/10.3390/ijms22179300
Chicago/Turabian StyleCricrí, Dalma, Lara Coppi, Silvia Pedretti, Nico Mitro, Donatella Caruso, Emma De Fabiani, and Maurizio Crestani. 2021. "Histone Deacetylase 3 Regulates Adipocyte Phenotype at Early Stages of Differentiation" International Journal of Molecular Sciences 22, no. 17: 9300. https://doi.org/10.3390/ijms22179300
APA StyleCricrí, D., Coppi, L., Pedretti, S., Mitro, N., Caruso, D., De Fabiani, E., & Crestani, M. (2021). Histone Deacetylase 3 Regulates Adipocyte Phenotype at Early Stages of Differentiation. International Journal of Molecular Sciences, 22(17), 9300. https://doi.org/10.3390/ijms22179300