
Identification of Novel Insulin Resistance Related ceRNA Network in T2DM and Its Potential Editing by CRISPR/Cas9

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Demographic, Clinical Data and Diabetes Laboratory Profile of the Diabetic and Healthy Groups
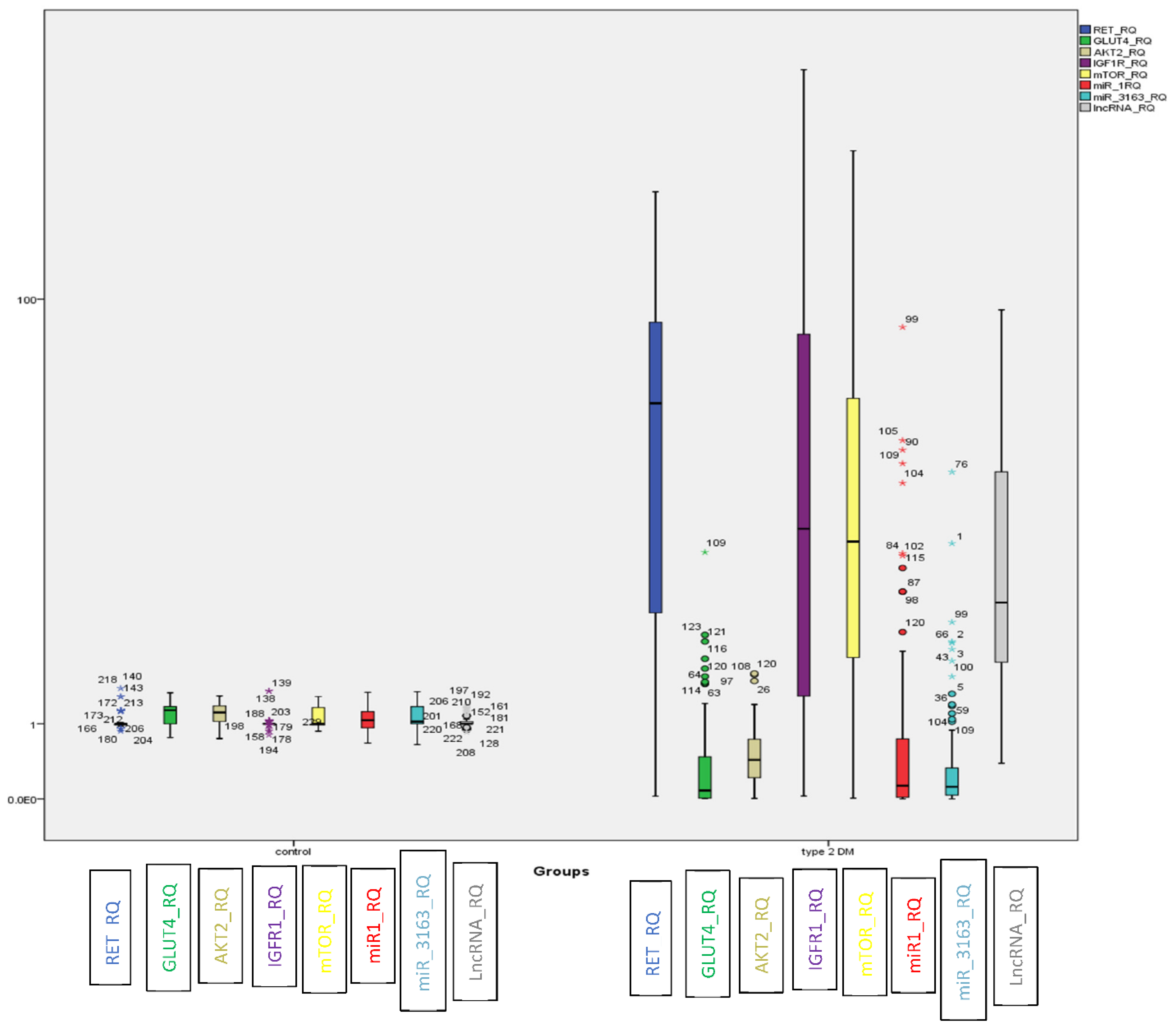
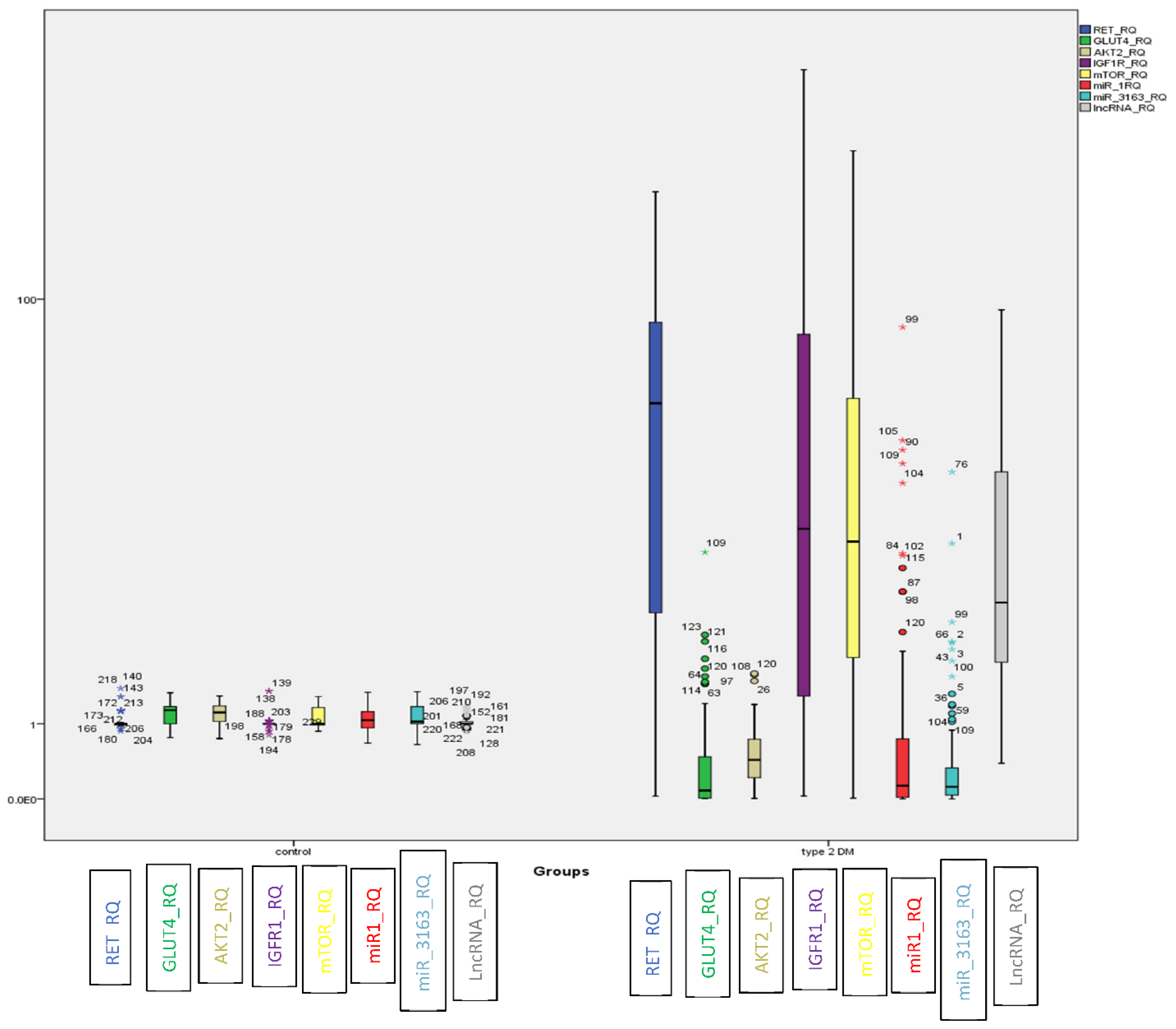
2.2. Evaluation of the Identified ceRNA Panel Expression in the Diabetic and Healthy Groups
2.3. Correlation between Different Partners in the Genetic Network, Glycemic Control and Insulin Resistance in Both Diabetic and Healthy Groups
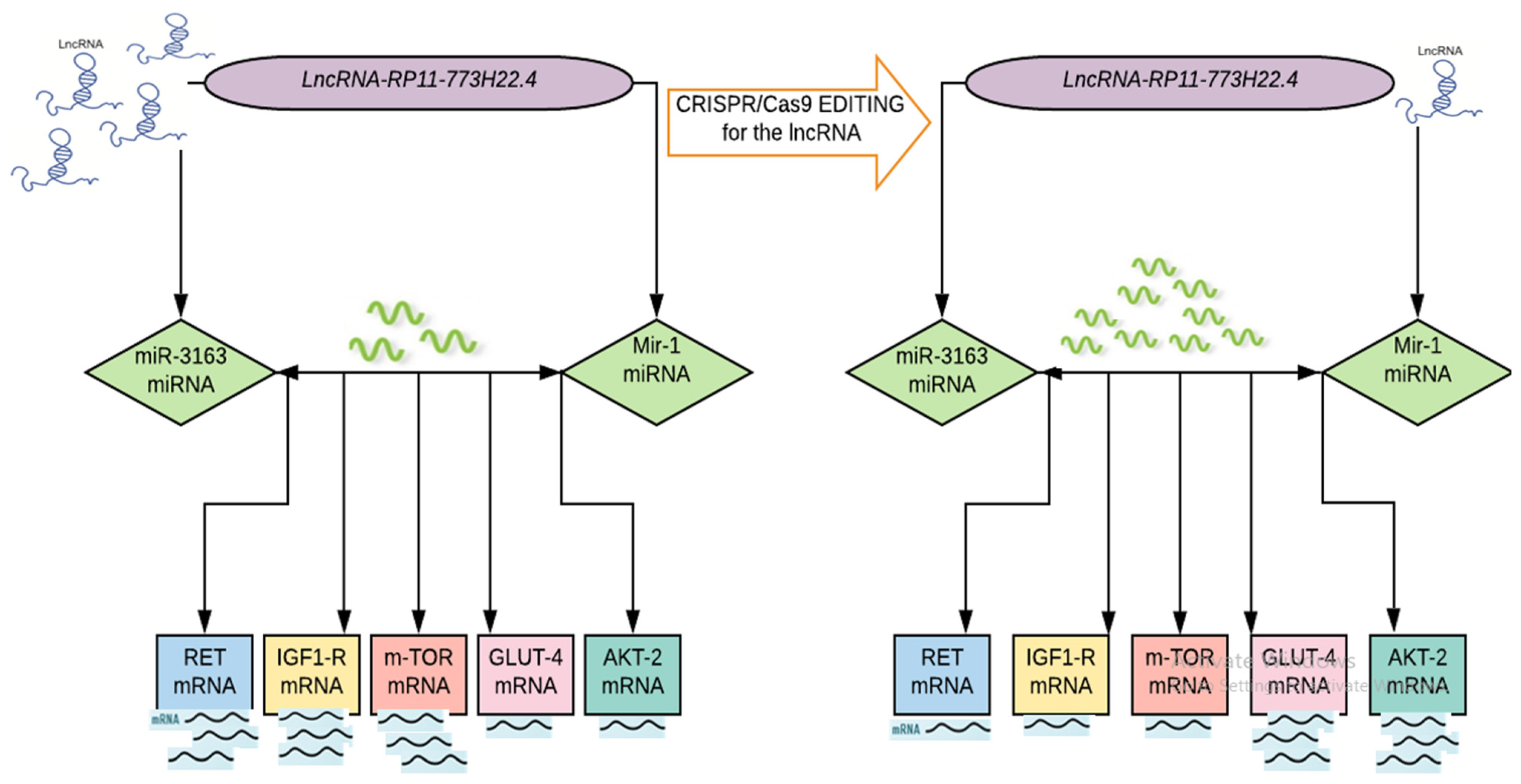
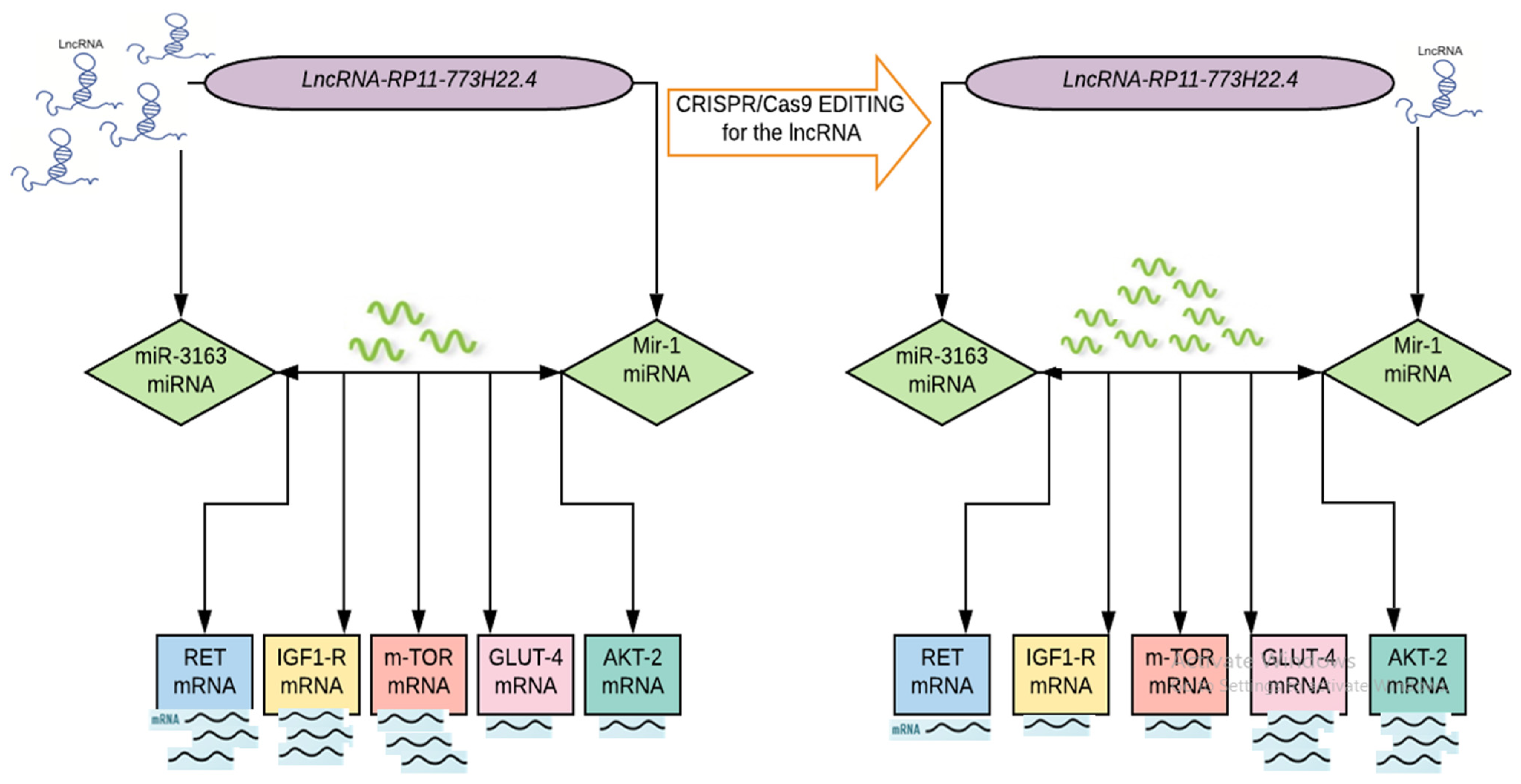
2.4. CRISPR/Cas9 Mediated Knockout of LncRNA-RP11-773H22.4 in Lymphocyte Cell Line Obtained from Diabetic Patients
2.4.1. Effect of CRISPR/Cas9 Editing on Cell Count and Viability
2.4.2. Effect of CRISPR/Cas9 Editing on the Expression of the Identified Genetic Network
2.4.3. Effect of CRISPR/Cas9 Editing on GLUT4 and mTOR Proteins
3. Discussion
4. Material and Methods
4.1. Participants and Samples
4.2. Bioinformatics-Based Selection of Insulin Resistance Related ceRNA Panel
4.3. Validation of the Identified Insulin Resistance Related ceRNA Panel in Human Clinical Samples
4.3.1. Extraction of Total RNA, Including Coding and Non-Coding RNAs from Human PBMCs Samples
4.3.2. Quantitative Real Time-PCR of the Identified Insulin Resistance Related ceRNA Panel
4.4. Validation of the Identified Insulin Resistance related ceRNA Panel and CRISPR/Cas9 Editing of LncRNA-RP11-773H22.4 in Lymphocyte Cell Line
4.4.1. Culture of Human Lymphocytes
4.4.2. Synthesis of gRNAs
4.4.3. Lymphocyte Transfection in a 12-Well Plate Using Lipofectamine CRISPRMAX Reagent (cat. no. CMAX00008, Thermofischer, Waltham, MA, USA)
4.4.4. Immunofluorescence Microscopy to Evaluate the Activity of GLUT4 and mTOR Proteins
4.5. Statistics
4.6. Data and Resource Availability
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Wu, Y.; Ding, Y.; Tanaka, Y.; Zhang, W. Risk Factors Contributing to Type 2 Diabetes and Recent Advances in the Treatment and Prevention. Int. J. Med. Sci. 2014, 11, 1185–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Diabetes Federation. IDF Diabetes Atlas, 9th ed.; International Diabetes Federation: Brussels, Belgium, 2019. [Google Scholar]
- Hegazi, R.; El-Gamal, M.; Abdel-Hady, N.; Hamdy, O. Epidemiology of and Risk Factors for Type 2 Diabetes in Egypt. Ann. Glob. Health 2016, 81, 814–820. [Google Scholar] [CrossRef] [Green Version]
- Sommese, L.; Zullo, A.; Mancini, F.P.; Fabbricini, R.; Soricelli, A.; Napoli, C. Clinical relevance of epigenetics in the onset and management of type 2 diabetes mellitus. Epigenetics 2017, 12, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, S.; Iossa, S.; Venditti, P. Skeletal muscle insulin resistance: Role of mitochondria and other ROS sources. J Endocrinol. 2017, 233, R15–R42. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Li, X.; Zhan, X.; Sun, L.; Gao, J.; Cao, Y.; Qiu, H. Construction of competitive endogenous RNA network reveals regulatory role of long non-coding RNAs in type 2 diabetes mellitus. J. Cell Mol. Med. 2017, 12, 3204–3213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulitsky, I. Evolution to the rescue: Using comparative genomics to understand long non-coding RNAs. Nat. Rev. Genet. 2016, 17, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Sathishkumar, C.; Prabu, P.; Mohan, V.; Balasubramanyam, M. Linking a role of lncRNAs (long non-coding RNAs) with insulin resistance, accelerated senescence, and inflammation in patients with type 2 diabetes. Hum. Genom. 2018, 12, 41. [Google Scholar] [CrossRef]
- Honardoost, M.; Keramati, F.; Arefian, E.; Yeganeh, S.M.; Soleimani, M. Network of three specific microRNAs influence type 2 diabetes through inducing insulin resistance in muscle cell lines. J. Cell. Biochem. 2019, 120, 1532–1538. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Mao, Z.D.; Shi, Y.J.; Qian, Y.; Liu, Z.G.; Yin, X.W. Comprehensive analysis of miRNA-mRNA-lncRNA networks in severe asthma. Epigenomics 2019, 11, 115–131. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Zhang, T.; Qi, L.; Zhou, C.; Wei, J.; Feng, F.; Liu, R.; Sun, C. Integrated analysis of co-expression and ceRNA network identifies five lncRNAs as prognostic markers for breast cancer. J. Cell. Mol. Med. 2019, 23, 8410–8419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.J.; Kang, K.H.; Ju, J.H. CRISPR-Cas9: A promising tool for gene editing on induced pluripotent stem cells. Korean J. Intern. Med. 2017, 32, 42–61. [Google Scholar] [CrossRef] [PubMed]
- De Rosa, S.; Arcidiacono, B.; Chiefari, E.; Brunetti, A.; Indolfi, C.; Foti, D.P. Type 2 Diabetes Mellitus and Cardiovascular Disease: Genetic and Epigenetic Links. Front. Endocrinol. 2018, 9, 2. [Google Scholar] [CrossRef]
- Liu, Q.; Tong, D.; Yuan, W.; Liu, G.; Yuan, G.; Lan, W.; Zhang, D.; Zhang, J.; Huang, Z.; Zhang, Y.; et al. Different RET gene mutation-induced multiple endocrine neoplasia type 2A in 3 Chinese families. Medicine 2017, 96, e5967. [Google Scholar] [CrossRef] [PubMed]
- Paratala, B.S.; Chung, J.H.; Williams, C.B.; Yilmazel, B.; Petrosky, W.; Williams, K.; Schrock, A.B.; Gay, L.M.; Lee, E.; Dolfi, S.C.; et al. RET rearrangements are actionable alterations in breast cancer. Nat. Commun. 2018, 9, 4821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteves, J.V.; Enguita, F.J.; Machado, U.F. MicroRNAs-Mediated Regulation of Skeletal Muscle GLUT4 Expression and Translocation in Insulin Resistance. J. Diabetes Res. 2017, 2017, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Acharya, S.; Xu, J.; Wang, X.; Jain, S.; Wang, H.; Zhang, Q.; Chang, C.-C.; Bower, J.; Arun, B.; Seewaldt, V.; et al. Downregulation of GLUT4 contributes to effective intervention of estrogen receptor-negative/HER2-overexpressing early stage breast disease progression by lapatinib. Am. J. Cancer Res. 2016, 6, 981–995. [Google Scholar] [PubMed]
- Cho, H.; Mu, J.; Kim, J.K.; Thorvaldsen, J.L.; Chu, Q.; Crenshaw, E.B., III; Kaestner, K.H.; Bartolomei, M.S.; Shulman, G.I.; Birnbaum, M.J. Insulin resistance and a diabetes mellitus-like syndrome in mice lacking the protein kinase Akt2 (PKB β). Science 2001, 292, 1728–1731. [Google Scholar] [CrossRef] [PubMed]
- Caruso, M.; Zhang, X.; Ma, D.; Yang, Z.; Qi, Y.; Yi, Z. Novel Endogenous, Insulin-Stimulated Akt2 Protein Interaction Partners in L6 Myoblasts. PLoS ONE 2015, 10, e0140255. [Google Scholar] [CrossRef]
- De Meyts, P.; Whittaker, J. Structural biology of insulin and IGF1 receptors: Implications for drug design. Nat. Rev. Drug Discov. 2002, 1, 769–783. [Google Scholar] [CrossRef]
- Sachdev, D.; Li, S.-L.; Hartell, J.S.; Fujita-Yamaguchi, Y.; Miller, J.S.; Yee, D. A chimeric humanized single-chain antibody against the type I insulin-like growth factor (IGF) receptor renders breast cancer cells refractory to the mitogenic effects of IGFI. Cancer Res. 2003, 63, 627–635. [Google Scholar]
- Das, A.; Reis, F.; Maejima, Y.; Cai, Z.; Ren, J. mTOR Signaling in Cardiometabolic Disease, Cancer, and Aging. Oxidative Med. Cell. Longev. 2017, 2017, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Khan, K.H.; Wong, M.; Rihawi, K.; Bodla, S.; Morganstein, D.; Banerji, U.; Molife, L.R. Hyperglycemia and Phosphatidylinositol 3-Kinase/Protein Kinase B/Mammalian Target of Rapamycin (PI3K/AKT/mTOR) Inhibitors in Phase I Trials: Incidence, Predictive Factors, and Management. Oncology 2016, 21, 855–860. [Google Scholar] [CrossRef] [Green Version]
- Nigi, L.; Grieco, G.E.; Ventriglia, G.; Brusco, N.; Mancarella, F.; Formichi, C.; Dotta, F.; Sebastiani, G. MicroRNAs as Regulators of Insulin Signaling: Research Updates and Potential Therapeutic Perspectives in Type 2 Diabetes. Int. J. Mol. Sci. 2018, 19, 3705. [Google Scholar] [CrossRef] [Green Version]
- Yildirim, S.S.; Akman, D.; Catalucci, D.; Turan, B. Relationship Between Downregulation of miRNAs and Increase of Oxidative Stress in the Development of Diabetic Cardiac Dysfunction: Junctin as a Target Protein of miR-1. Cell Biophys. 2013, 67, 1397–1408. [Google Scholar] [CrossRef]
- Chen, T.; Ding, G.; Jin, Z.; Wagner, M.B.; Yuan, Z. Insulin ameliorates miR-1-induced injury in H9c2 cells under oxidative stress via Akt activation. Mol. Cell. Biochem. 2012, 369, 167–174. [Google Scholar] [CrossRef]
- Feng, B.; Cao, Y.; Chen, S.; Ruiz, M.; Chakrabarti, S. Reprint of: miRNA-1 regulates endothelin-1 in diabetes. Life Sci. 2014, 118, 275–280. [Google Scholar] [CrossRef] [Green Version]
- Su, L.; Han, D.; Wu, J.; Huo, X. Skp2 regulates non-small cell lung cancer cell growth by Meg3 and miR-3163. Tumour Biol. 2016, 37, 3925–3931. [Google Scholar] [CrossRef]
- Jia, M.; Wei, Z.; Liu, P.; Zhao, X. Silencing of ABCG2 by MicroRNA-3163 Inhibits Multidrug Resistance in Retinoblastoma Cancer Stem Cells. J. Korean Med. Sci. 2016, 31, 836–842. [Google Scholar] [CrossRef]
- Chakraborty, C.; Doss, C.G.P.; Bandyopadhyay, S.; Agoramoorthy, G. Influence of miRNA in insulin signaling pathway and insulin resistance: Micro-molecules with a major role in type-2 diabetes. Wiley Interdiscip. Rev. RNA 2014, 5, 697–712. [Google Scholar] [CrossRef]
- Wang, L.; Cho, K.B.; Li, Y.; Tao, G.; Xie, Z.; Guo, B. Long Noncoding RNA (lncRNA)-Mediated Competing Endogenous RNA Networks Provide Novel Potential Biomarkers and Therapeutic Targets for Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 5758. [Google Scholar] [CrossRef] [Green Version]
- Kamel, M.M.; Matboli, M.; Sallam, M.; Montasser, M.F.; Saad, A.S.; El-Tawdi, A.H. Investigation of long noncoding RNAs expression profile as potential serum biomarkers in patients with hepatocellular carcinoma. Transl. Res. 2016, 168, 134–145. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Lefort, C.T.; Kim, M. Human T Lymphocyte Isolation, Culture and Analysis of Migration In Vitro. J. Vis. Exp. 2010, e2017. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Liang, X.; Xie, H.; Kumar, S.; Ravinder, N.; Potter, J.; Jeu, X.D.M.D.; Chesnut, J.D. Improved delivery of Cas9 protein/gRNA complexes using lipofectamine CRISPRMAX. Biotechnol. Lett. 2016. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| Type 2 DM N (%) | Healthy N (%) | χ2(P) (a) | |
|---|---|---|---|
| Sex: Male (87) Female (142) | 47 (38.2%) 76 (61.8%) | 40 (37.7%) 66 (62.3%) | 0.005 (0.941) |
| Smoking: smoker: (102) non-smoker:(117) x-smoker: (10) | 86 (69.9%) 30 (24.4%) 7 (5.7%) | 60 (56.7%) 46 (43.3%) 3 (2.8%) | 0.004 (0.870) |
| Family history of diabetes: Positive: (98) Negative: (131) | 98 (79.7%) 25 (20.3%) | 0 (0%) 106 (100%) | 147.636 (0.000 **) |
| Oral Anti Diabetic Medications (OAD): Metformin: (44) SU: (24) Combined: (12) Not Taking OAD: (149) | 44 (35.8%) 24 (19.5%) 12 (9.8%) 43 (35%) | NA | NA |
| Mean ± SD | Mean ± SD | t(P) (b) | |
| Duration of diabetes (yrs) | 9.38 ± 6.067 | NA | NA |
| Fasting blood glucose (mg/dL) | 197 ± 91.852 | 88.08 ± 14.594 | 12.173 (0.000 **) |
| 2hpp glucose (mg/dL) | 274.79 ± 99.6 | 111.07 ± 16.926 | 16.712 (0.000 **) |
| Systolic Bl.pr. (mmHg) | 135.45 ± 15.59 | 118.02 ± 7.736 | 10.452 (0.000 **) |
| Diastolic Bl.pr. (mmHg) | 88.09 ± 11.969 | 76.56 ± 5.088 | 9.226 (0.000 **) |
| BMI (kg/m2) | 34.05 ± 5.71 | 30.67 ± 4.93 | 2.347 (0.000 **) |
| Total Cholesterol (mg/dL) | 292.46 ± 60.866 | 102.97 ± 24.47 | 30.023 (0.000 **) |
| TGs (mg/dL) | 263.84 ± 75.914 | 90.16 ± 19.62 | 22.766 (0.000 **) |
| HDL (mg/dL) | 31.98 ± 9.6 | 71.18 ± 11.235 | −28.458 (0.000 **) |
| LDL (mg/dL) | 200.77 ± 55.287 | 75.49 ± 17.861 | 22.341 (0.000 **) |
| alb./cr. (mg/mg) | 24.39 ± 4.625 | 13.59 ± 3.349 | 19.944 (0.000 **) |
| Hb A1c (gm%) | 7.91 ± 2.11 | 3.89 ± 1.205 | 17.307 (0.000 **) |
| Mean Rank | Mean Rank | U(P) (c) | |
| Fasting insulin (uIU/mL) | 140.52 | 85.38 | 3379.5 (0.000 **) |
| HOMA-IR | 161.67 | 60.84 | 778 (0.000 **) |
| Type 2DM N (%) | Healthy N (%) | χ2(P) (a) | |
|---|---|---|---|
| HOMA-IR Positive ≥ 2.3 (102) Negative < 2.3 (127) | 102 (82.9%) 21 (17.1%) | 0 (0%) 106 (100%) | 158.501 (0.000 **) |
| LncRNA-RP11-773H22.4 RQ: Positive ≥ 1.2 (124) Negative < 1.2 (105) | 117 (95.1%) 6 (4.9%) | 7 (6.6%) 99 (93.4%) | 179.680 (0.000 **) |
| miR-3163 RQ: Positive ≤ 0.91 (120) Negative > 0.91 (109) | 109 (88.6%) 14 (11.4%) | 11 (10.4%) 95 (89.6%) | 139.734 (0.000 **) |
| miR-1 RQ: Positive ≤0.87 (116) Negative > 0.87(113) | 96 (78%) 27 (22%) | 20 (18.9%) 86 (81.1%) | 79.776 (0.000 **) |
| RET mRNA RQ: Positive ≥ 1.36 (117) Negative < 1.36 (112) | 114 (92.7%) 9 (7.3%) | 3 (2.8%) 103 (97.2%) | 183.952 (0.000 **) |
| mTOR mRNA RQ: Positive ≥ 1.4 (116) Negative < 1.4 (113) | 105 (85.4%) 18 (14.6%) | 11 (10.4%) 95 (89.6%) | 128.085 (0.000 **) |
| IGF1R mRNA RQ: Positive ≥ 1.07 (100) Negative < 1.07 (129) | 98 (79.7%) 25 (20.3%) | 2 (1.9%) 104 (98.1%) | 140.05 (0.000 **) |
| GLUT4 mRNA RQ: Positive ≤ 0.92 (116) Negative > 0.92 (113) | 109 (88.6%) 14 (11.4%) | 7 (6.6%) 99 (93.4%) | 153.210 (0.000 **) |
| AKT2 mRNA RQ: Positive ≤ 0.99 (118) Negative > 0.99 (111) | 113(91.9%) 10 (8.1%) | 5 (4.7%) 101 (95.3%) | 173.143 (0.000 **) |
| lncRNA_RQ | miR_3163_RQ | miR_1RQ | RET_RQ | IGF1R_RQ | mTOR_RQ | GLUT4_RQ | AKT2_RQ | HbA1c | HOMA_IR | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| lncRNA_RQ | Correlation Coefficient (Sig.) | 1.000 | −0.515 (0.000) | −0.366 (0.000) | 0.656 (0.000) | 0.483 (0.000) | 0.469 (0.000) | −0.579 (0.000) | −0.588 (0.000) | 0.651 (0.000) | 0.581 (0.000) |
| miR_3163_RQ | Correlation Coefficient (Sig.) | −0.515 (0.000) | 1.000 | 0.459 (0.000) | −0.475 (0.000) | −0.330 (0.000) | −0.399 (0.000) | 0.455 (0.000) | 0.505 (0.000) | −0.576 (0.000) | −0.555 (0.000) |
| miR_1RQ | Correlation Coefficient (Sig.) | −0.366 (0.000) | 0.459 (0.000) | 1.000 | −0.457 (0.000) | −0.290 (0.000) | −0.259 (0.000) | 0.377 (0.000) | 0.412 (0.000) | −0.362 (0.000) | −0.378 (0.000) |
| RET_RQ | Correlation Coefficient (Sig.) | 0.656 (0.000) | −0.475 (0.000) | −0.457 (0.000) | 1.000 | 0.694 (0.000) | 0.440 (0.000) | −0.491 (0.000) | −0.615 (0.000) | 0.562 (0.000) | 0.543 (0.000) |
| IGF1R_RQ | Correlation Coefficient (Sig.) | 0.483 (0.000) | −0.330 (0.000) | −0.290 (0.000) | 0.694 (0.000) | 1.000 | 0.392 (0.000) | −0.362 (0.000) | −0.420 (0.000) | 0.462 (0.000) | 0.296 (0.000) |
| mTOR_RQ | Correlation Coefficient (Sig.) | 0.469 (0.000) | −0.399 (0.000) | −0.259 (0.000) | 0.440 (0.000) | 0.392 (0.000) | 1.000 | −0.421 (0.000) | −0.488 (0.000) | 0.547 (0.000) | 0.415 (0.000) |
| GLUT4_RQ | Correlation Coefficient (Sig.) | −0.579 (0.000) | 0.455 (0.000) | 0.377 (0.000) | −0.491 (0.000) | −0.362 (0.000) | −0.421 (0.000) | 1.000 | 0.523 (0.000) | −0.517 (0.000) | −0.573 (0.000) |
| AKT2_RQ | Correlation Coefficient (Sig.) | −0.588 (0.000) | 0.505 (0.000) | 0.412 (0.000) | −0.615 (0.000) | −0.420 (0.000) | −0.488 (0.000) | 0.523 (0.000) | 1.000 | −0.614 (0.000) | −0.608 (0.000) |
| HbA1c | Correlation Coefficient (Sig.) | 0.651 (0.000) | −0.576 (0.000) | −0.362 (0.000) | 0.562 (0.000) | 0.462 (0.000) | 0.547 (0.000) | −0.517 (0.000) | −0.614 (0.000) | 1.000 | 0.611 (0.000) |
| HOMA_IR | Correlation Coefficient (Sig.) | 0.581 (0.000) | −0.555 (0.000) | −0.378 (0.000) | 0.543 (0.000) | 0.296 (0.000) | 0.415 (0.000) | −0.573 (0.000) | −0.608 (0.000) | 0.611 (0.000) | 1.000 |
| Lymphocytes’ Cell Lines | U(P) (a) | ||
|---|---|---|---|
| T2DM Lymphocytes without Editing | T2DM Lymphocytes with CRISPR Editing | ||
| Mean Rank | Mean Rank | ||
| LncRNA-RP11-773H22.4 RQ | 14 | 5 | 0.000 (0.000 **) |
| miR-3163 miRNA RQ | 5 | 14 | 0.000 (0.001 **) |
| miR-1 miRNA RQ | 5 | 14 | 0.000 (0.001 **) |
| RET mRNA RQ | 14 | 5 | 0.000 (0.000 **) |
| IGF1R mRNA RQ | 14 | 5 | 0.000 (0.000 **) |
| mTOR mRNA RQ | 14 | 5 | 0.000 (0.000 **) |
| GLUT-4 mRNA RQ | 5 | 14 | 0.000 (0.000 **) |
| AKT2 mRNA RQ | 5 | 14 | 0.000 (0.000 **) |
| lncRNA_RQ | miR_3163_RQ | miR_1 RQ | RET_RQ | IGF1R_RQ | mTOR_RQ | GLUT4RQ | AKT2_RQ | ||
|---|---|---|---|---|---|---|---|---|---|
| lncRNA_RQ | Correlation Coefficient (Sig.) | 1.000 | −0.931 (0.000) | −0.983 (0.000) | 0.914 (0.000) | 0.931 (0.000) | 0.879 (0.000) | −0.897 (0.000) | −0.879 (0.000) |
| miR_3163_RQ | Correlation Coefficient (Sig.) | −0.931 (0.000) | 1.000 | 0.965 (0.000) | −0.948 (0.000) | −0.965 (0.000) | −0.931 (0.000) | 0.913 (0.000) | 0.948 (0.000) |
| miR_1RQ | Correlation Coefficient (Sig.) | −0.983 (0.000) | 0.965 (0.000) | 1.000 | −0.931 (0.000 | −0.948 (0.000) | −0.897 (0.000) | 0.879 (0.000) | 0.897 (0.000) |
| RET_RQ | Correlation Coefficient (Sig.) | 0.914 (0.000) | −0.948 (0.000) | −0.931 (0.000) | 1.000 | 0.983 (0.000) | 0.983 (0.000) | −0.919 (0.000) | −0.931 (0.000) |
| IGF1R_RQ | Correlation Coefficient (Sig.) | 0.931 (0.000) | −0.965 (0.000) | −0.948 (0.000) | 0.983 (0.000) | 1.000 | 0.948 (0.000) | −0.931 (0.000) | −0.948 (0.000) |
| mTOR_RQ | Correlation Coefficient (Sig.) | 0.879 (0.000) | −0.931 (0.000) | −0.897 (0.000) | 0.983 (0.000) | 0.948 (0.000) | 1.000 | −0.931 (0.000) | −0.948 (0.000) |
| GLUT4RQ | Correlation Coefficient (Sig.) | −0.897 (0.000) | 0.913 (0.000) | 0.879 (0.000) | −0.914 (0.000) | −0.931 (0.000) | −0.931 (0.000) | 1.000 | 0.983 (0.000) |
| AKT2_RQ | Correlation Coefficient (Sig.) | −0.879 (0.000) | 0.948 (0.000) | 0.897 (0.000) | −0.931 (0.000) | −0.948 (0.000) | −0.948 (0.000) | 0.983 (0.000) | 1.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matboli, M.; Kamel, M.M.; Essawy, N.; Bekhit, M.M.; Abdulrahman, B.; Mohamed, G.F.; Eissa, S. Identification of Novel Insulin Resistance Related ceRNA Network in T2DM and Its Potential Editing by CRISPR/Cas9. Int. J. Mol. Sci. 2021, 22, 8129. https://doi.org/10.3390/ijms22158129
Matboli M, Kamel MM, Essawy N, Bekhit MM, Abdulrahman B, Mohamed GF, Eissa S. Identification of Novel Insulin Resistance Related ceRNA Network in T2DM and Its Potential Editing by CRISPR/Cas9. International Journal of Molecular Sciences. 2021; 22(15):8129. https://doi.org/10.3390/ijms22158129
Chicago/Turabian StyleMatboli, Marwa, Marwa Mostafa Kamel, Nada Essawy, Meram Mohamed Bekhit, Basant Abdulrahman, Ghada F. Mohamed, and Sanaa Eissa. 2021. "Identification of Novel Insulin Resistance Related ceRNA Network in T2DM and Its Potential Editing by CRISPR/Cas9" International Journal of Molecular Sciences 22, no. 15: 8129. https://doi.org/10.3390/ijms22158129
APA StyleMatboli, M., Kamel, M. M., Essawy, N., Bekhit, M. M., Abdulrahman, B., Mohamed, G. F., & Eissa, S. (2021). Identification of Novel Insulin Resistance Related ceRNA Network in T2DM and Its Potential Editing by CRISPR/Cas9. International Journal of Molecular Sciences, 22(15), 8129. https://doi.org/10.3390/ijms22158129