
Impact of Long-Term Manure and Sewage Sludge Application to Soil as Organic Fertilizer on the Incidence of Pathogenic Microorganisms and Antibiotic Resistance Genes

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples Collection
2.2. DNA Isolation and 16S rRNA Amplicons Generation
2.3. Data Processing and Analysis
2.4. qPCR Assays
2.5. Statistical Analysis of ARGs Abundance
3. Results and Discussion
3.1. Microbiological Safety of the Organic Fertilizers
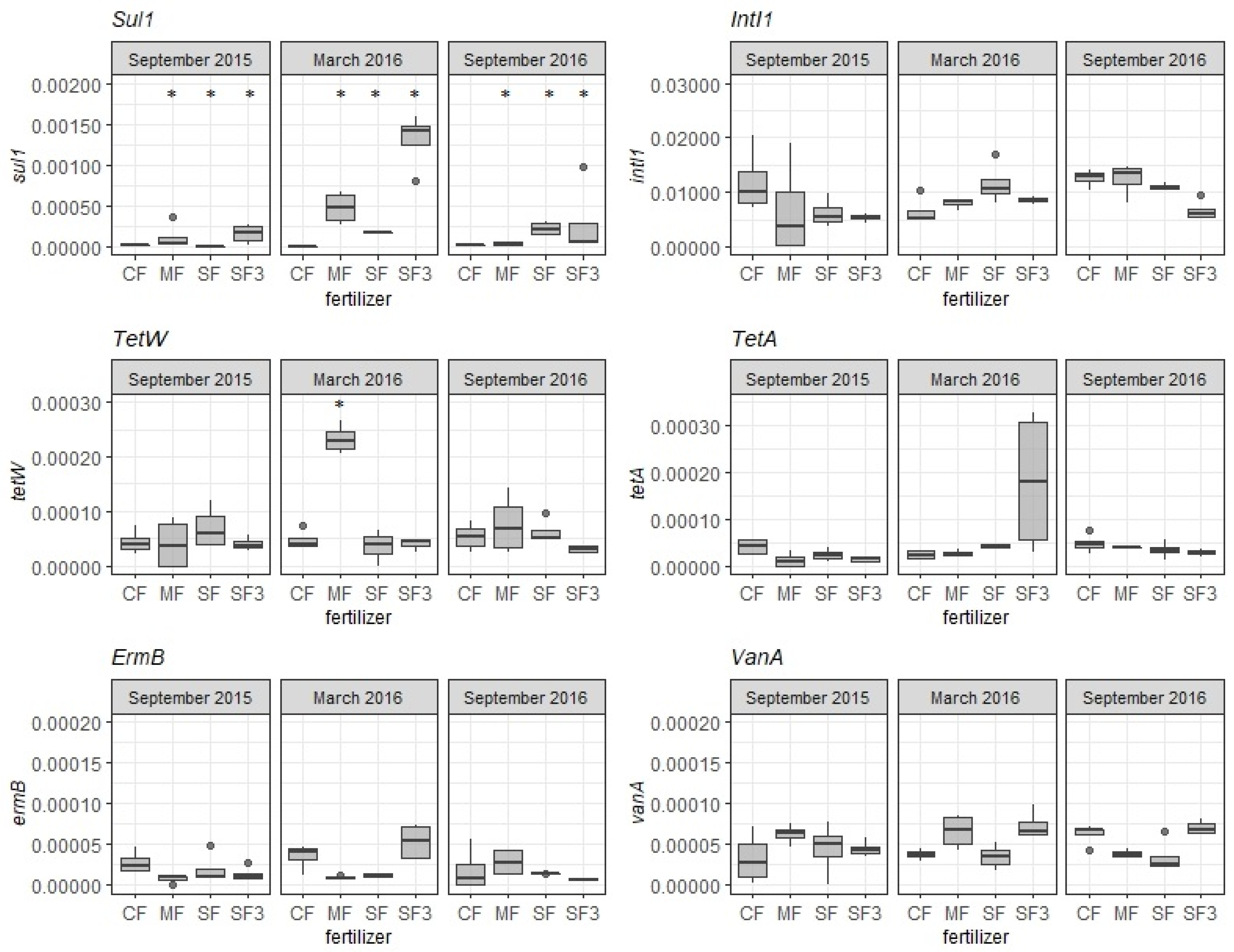
3.2. Abundance of ARGs in the Organic Fertilizers
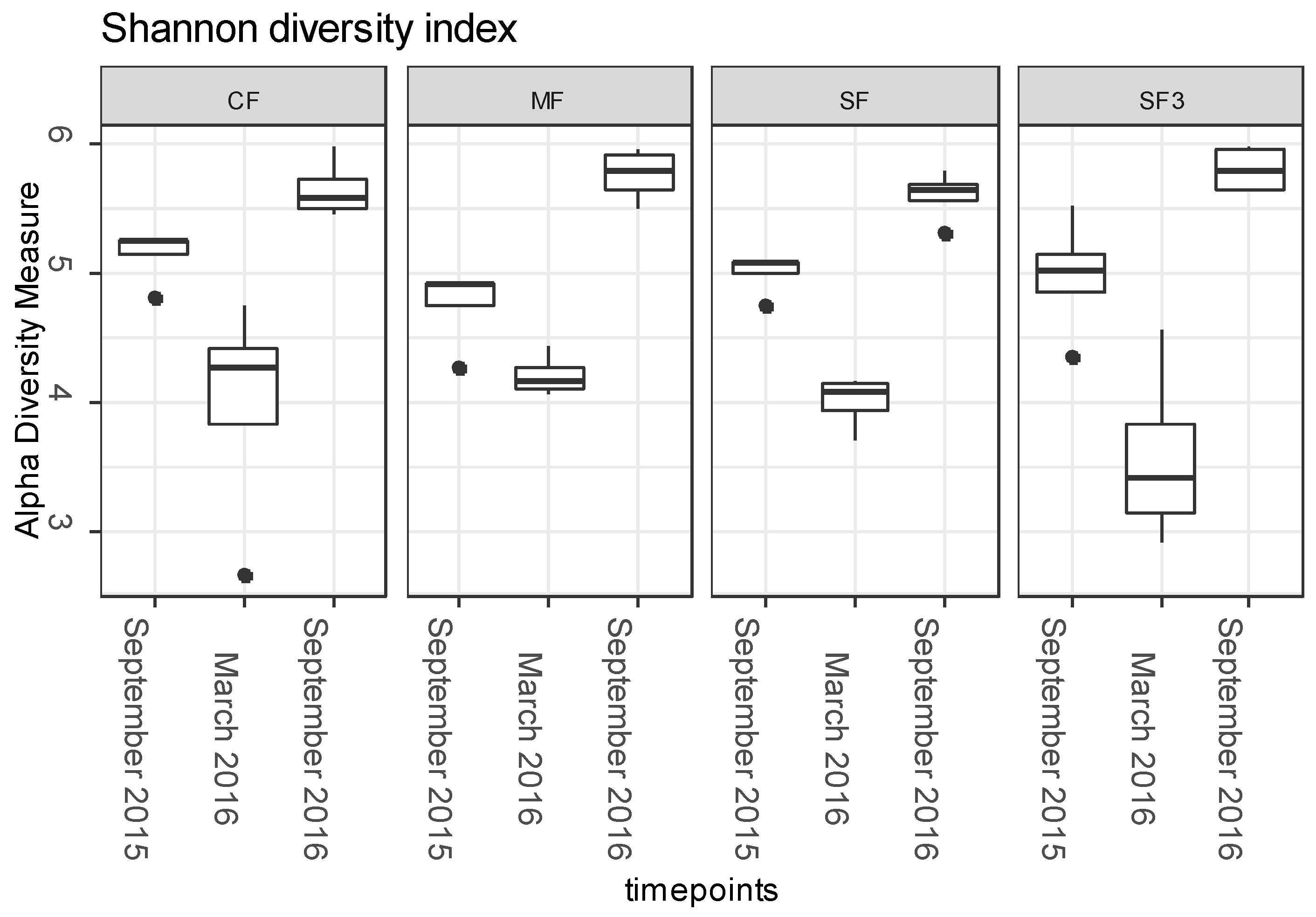
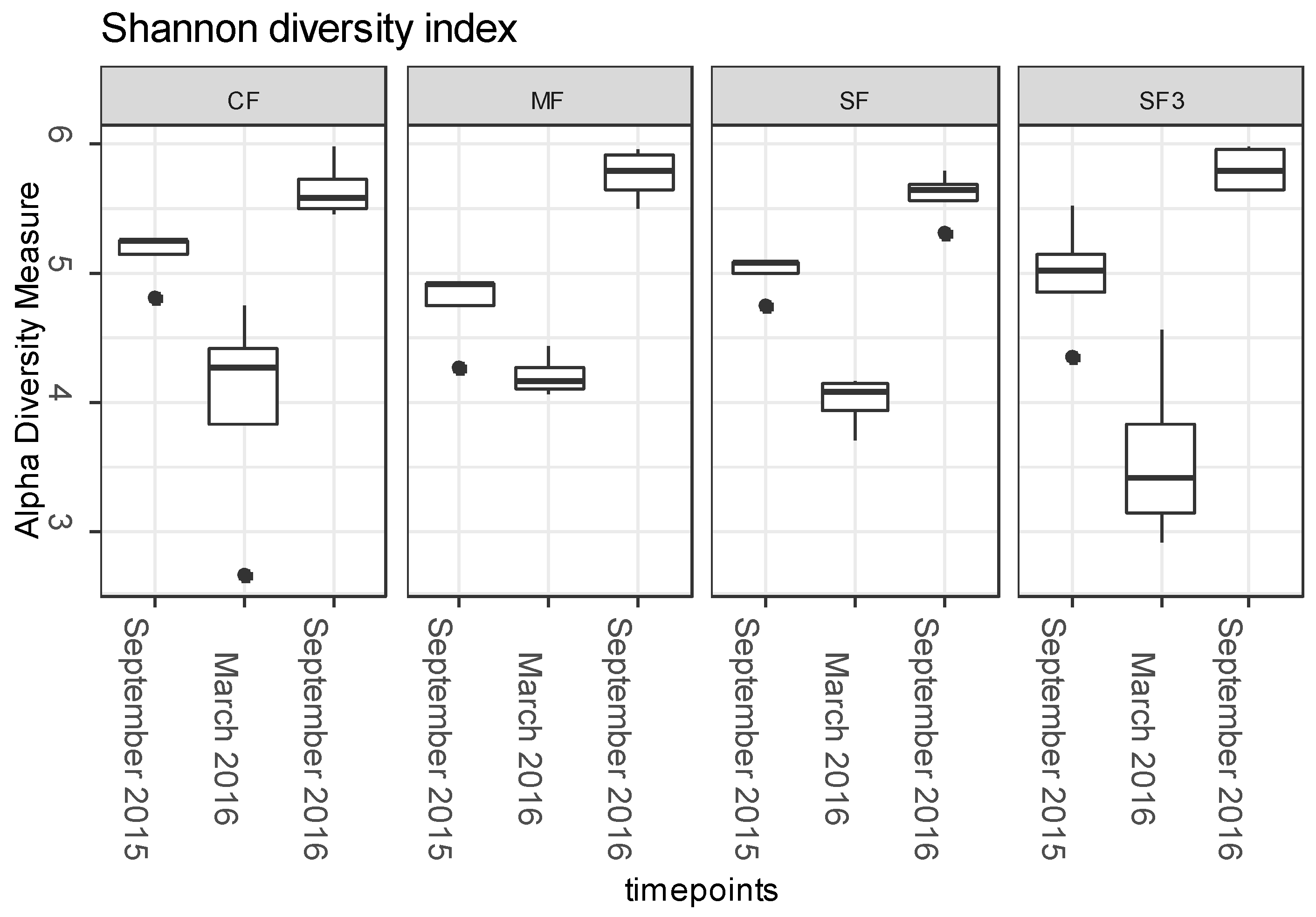
3.3. ARGs Fate in Fertilized Soils
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.T.; Shen, H.; He, X.H.; Thomas, B.; Lupwayi, N.Z.; Hao, X.Y.; Thomas, M.C.; Shi, X.J. Fertilization Shapes Bacterial Community Structure by Alteration of Soil pH. Front. Microbiol. 2017, 8, 1325. [Google Scholar] [CrossRef] [Green Version]
- Sorinolu, A.J.; Tyagi, N.; Kumar, A.; Munir, M. Antibiotic resistance development and human health risks during wastewater reuse and biosolids application in agriculture. Chemosphere 2021, 265, 129032. [Google Scholar] [CrossRef]
- Gómez-Sagasti, M.T.; Hernández, A.; Artetxe, U.; Garbisu, C.; Becerril, J.M. How Valuable Are Organic Amendments as Tools for the Phytomanagement of Degraded Soils? The Knowns, Known Unknowns, and Unknowns. Front. Sustain. Food Syst. 2018, 2, 68. [Google Scholar] [CrossRef] [Green Version]
- Eurostat. Sewage Sludge Production and Disposal from Urban Wastewater. 2017. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php?title=File:Sewage_sludge_disposal_from_urban_wastewater_treatment,_by_treatment_method,_2017_(%25_of_total_dry_mass).png (accessed on 5 May 2021).
- Czech Statistical Office, C.S. Consumption of Fertilisers in the Crop Year. 2018. Available online: https://vdb.czso.cz/vdbvo2/faces/index.jsf?page=vystup-objekt&pvo=ZEM11#w= (accessed on 5 May 2021).
- Czech Statistical Office, C.S. Production and Disposal of Wastewater Digested Sludge. 2018. Available online: https://www.czso.cz/csu/czso/3-environment-v440y0ba3f (accessed on 5 May 2021).
- Collivignarelli, M.C.; Abba, A.; Frattarola, A.; Miino, M.C.; Padovani, S.; Katsoyiannis, I.; Torretta, V. Legislation for the Reuse of Biosolids on Agricultural Land in Europe: Overview. Sustainability 2019, 11, 6015. [Google Scholar] [CrossRef] [Green Version]
- EUR-Lex. Closing the Loop—An EU Action Plan for the Circular Economy. 2015. Available online: https://eur-lex.europa.eu/resource.html?uri=cellar:8a8ef5e8-99a0-11e5-b3b7-01aa75ed71a1.0012.02/DOC_1&format=PDF (accessed on 5 May 2021).
- Urra, J.; Alkorta, I.; Garbisu, C. Potential Benefits and Risks for Soil Health Derived From the Use of Organic Amendments in Agriculture. Agronomy 2019, 9, 542. [Google Scholar] [CrossRef] [Green Version]
- Stiborova, H.; Kolar, M.; Vrkoslavova, J.; Pulkrabova, J.; Hajslova, J.; Demnerova, K.; Uhlik, O. Linking toxicity profiles to pollutants in sludge and sediments. J. Hazard. Mater. 2017, 321, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.M.; Cummins, E. Evaluation of “Classic” and Emerging Contaminants Resulting from the Application of Biosolids to Agricultural Lands: A Review. Hum. Ecol. Risk Assess. Int. J. 2015, 21, 492–513. [Google Scholar] [CrossRef]
- Lee, W.; Park, S.H.; Kim, J.; Jung, J.Y. Occurrence and removal of hazardous chemicals and toxic metals in 27 industrial wastewater treatment plants in Korea. Desalin. Water Treat. 2015, 54, 1141–1149. [Google Scholar] [CrossRef]
- Rizzo, L.; Manaia, C.; Merlin, C.; Schwartz, T.; Dagot, C.; Ploy, M.C.; Michael, I.; Fatta-Kassinos, D. Urban wastewater treatment plants as hotspots for antibiotic resistant bacteria and genes spread into the environment: A review. Sci. Total Environ. 2013, 447, 345–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiborova, H.; Bacakova, A.; Musilova, L.; Demnerova, K. From Natural Phenomenon of Antibiotic Resistance in the Environment to Emergence of Multiresistant Strains. Chem. Listy 2018, 112, 833–839. [Google Scholar]
- Checcucci, A.; Trevisi, P.; Luise, D.; Modesto, M.; Blasioli, S.; Braschi, I.; Mattarelli, P. Exploring the Animal Waste Resistome: The Spread of Antimicrobial Resistance Genes Through the Use of Livestock Manure. Front. Microbiol. 2020, 11, 9. [Google Scholar] [CrossRef]
- Pazda, M.; Kumirska, J.; Stepnowski, P.; Mulkiewicz, E. Antibiotic resistance genes identified in wastewater treatment plant systems—A review. Sci. Total Environ. 2019, 697, 134023. [Google Scholar] [CrossRef]
- Chen, C.Q.; Pankow, C.A.; Oh, M.; Heath, L.S.; Zhang, L.Q.; Du, P.; Xia, K.; Pruden, A. Effect of antibiotic use and composting on antibiotic resistance gene abundance and resistome risks of soils receiving manure-derived amendments. Environ. Int. 2019, 128, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Zhang, W.; Yang, L.X.; Stedtfeld, R.D.; Peng, A.P.; Gu, C.; Boyd, S.A.; Li, H. Antibiotic resistance genes and bacterial communities in cornfield and pasture soils receiving swine and dairy manures. Environ. Pollut. 2019, 248, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Han, X.-M.; Hu, H.-W.; Chen, Q.-L.; Yang, L.-Y.; Li, H.-L.; Zhu, Y.-G.; Li, X.-Z.; Ma, Y.-B. Antibiotic resistance genes and associated bacterial communities in agricultural soils amended with different sources of animal manures. Soil Biol. Biochem. 2018, 126, 91–102. [Google Scholar] [CrossRef]
- Cycoń, M.; Mrozik, A.; Piotrowska-Seget, Z. Antibiotics in the Soil Environment—Degradation and Their Impact on Microbial Activity and Diversity. Front. Microbiol. 2019, 10, 338. [Google Scholar] [CrossRef]
- Pei, M.K.; Zhang, B.; He, Y.L.; Su, J.Q.; Gin, K.; Lev, O.; Shen, G.X.; Hu, S.Q. State of the art of tertiary treatment technologies for controlling antibiotic resistance in wastewater treatment plants. Environ. Int. 2019, 131, 105026. [Google Scholar] [CrossRef]
- Schluter, A.; Szczepanowski, R.; Puhler, A.; Top, E.M. Genomics of IncP-1 antibiotic resistance plasmids isolated from wastewater treatment plants provides evidence for a widely accessible drug resistance gene pool. Fems Microbiol. Rev. 2007, 31, 449–477. [Google Scholar] [CrossRef]
- Andersson, D.I.; Hughes, D. Microbiological effects of sublethal levels of antibiotics. Nat. Rev. Microl. 2014, 12, 465–478. [Google Scholar] [CrossRef]
- WHO. WHO Report on Surveillance of Antibiotic Consumption 2016–2018. 2018. Available online: https://www.who.int/medicines/areas/rational_use/oms-amr-amc-report-2016-2018/en/ (accessed on 5 May 2021).
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [Green Version]
- Klein, E.Y.; Van Boeckel, T.P.; Martinez, E.M.; Pant, S.; Gandra, S.; Levin, S.A.; Goossens, H.; Laxminarayan, R. Global increase and geographic convergence in antibiotic consumption between 2000 and 2015. Proc. Natl. Acad. Sci. USA 2018, 115, E3463–E3470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiborova, H.; Wolfram, J.; Demnerova, K.; Macek, T.; Uhlik, O. Bacterial community structure in treated sewage sludge with mesophilic and thermophilic anaerobic digestion. Folia Microbiol. 2015, 60, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Goberna, M.; Simón, P.; Hernández, M.T.; García, C. Prokaryotic communities and potential pathogens in sewage sludge: Response to wastewaster origin, loading rate and treatment technology. Sci. Total Environ. 2018, 615, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Ramos, T.M.; Jay-Russell, M.T.; Millner, P.D.; Shade, J.; Misiewicz, T.; Sorge, U.S.; Hutchinson, M.; Lilley, J.; Pires, A.F.A. Assessment of Biological Soil Amendments of Animal Origin Use, Research Needs, and Extension Opportunities in Organic Production. Front. Sustain. Food Syst. 2019, 3. [Google Scholar] [CrossRef] [Green Version]
- Clements, D.P.; Bihn, E.A. Chapter 16—The Impact of Food Safety Training on the Adoption of Good Agricultural Practices on Farms. In Safety and Practice for Organic Food; Biswas, D., Micallef, S.A., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 321–344. [Google Scholar]
- Sharma, M.; Millner, P.D.; Hashem, F.; Vinyard, B.T.; East, C.L.; Handy, E.T.; White, K.; Stonebraker, R.; Cotton, C.P. Survival of Escherichia coli in Manure-Amended Soils Is Affected by Spatiotemporal, Agricultural, and Weather Factors in the Mid-Atlantic United States. Appl. Environ. Microbiol. 2019, 85, e02392-18. [Google Scholar] [CrossRef] [Green Version]
- Ingham, S.C.; Losinski, J.A.; Andrews, M.P.; Breuer, J.E.; Breuer, J.R.; Wood, T.M.; Wright, T.H. Escherichia coli contamination of vegetables grown in soils fertilized with noncomposted bovine manure: Garden-scale studies. Appl. Environ. Microbiol. 2004, 70, 6420–6427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FDA. Multistate Outbreak of E. coli O157:H7 Infections Linked to Romaine Lettuce (Final Update). 2018. Available online: https://www.cdc.gov/ecoli/2018/o157h7-04-18/index.html (accessed on 26 November 2020).
- FDA. Outbreak Investigation of E. coli: Romaine from Salinas, California (November 2019). Available online: https://www.fda.gov/food/outbreaks-foodborne-illness/outbreak-investigation-e-coli-romaine-salinas-california-november-2019 (accessed on 26 November 2020).
- Gu, G.; Strawn, L.K.; Oryang, D.O.; Zheng, J.; Reed, E.A.; Ottesen, A.R.; Bell, R.L.; Chen, Y.; Duret, S.; Ingram, D.T.; et al. Agricultural Practices Influence Salmonella Contamination and Survival in Pre-harvest Tomato Production. Front. Microbiol. 2018, 9, 2451. [Google Scholar] [CrossRef] [Green Version]
- Kracmarova, M.; Karpiskova, J.; Uhlik, O.; Strejcek, M.; Szakova, J.; Balik, J.; Demnerova, K.; Stiborova, H. Microbial Communities in Soils and Endosphere of Solanum tuberosum L. and their Response to Long-Term Fertilization. Microorganisms 2020, 8, 1377. [Google Scholar] [CrossRef]
- Kracmarova, M.; Kratochvilova, H.; Uhlik, O.; Strejcek, M.; Szakova, J.; Cerny, J.; Tlustos, P.; Balik, J.; Demnerova, K.; Stiborova, H. Response of soil microbes and soil enzymatic activity to 20 years of fertilization. Agronomy 2020, 10, 1542. [Google Scholar] [CrossRef]
- Fraraccio, S.; Strejcek, M.; Dolinova, I.; Macek, T.; Uhlik, O. Secondary compound hypothesis revisited: Selected plant secondary metabolites promote bacterial degradation of cis-1,2-dichloroethylene (cDCE). Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. A language and environment for statistical computing in R Foundation for Statistical Computing. In Vienna, Austria. Available online: https://_wwwR-projectorg/ (accessed on 26 November 2020).
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.L.; McGarrell, D.M.; Sun, Y.N.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; Sankaran, K.; Fukuyama, J.A.; McMurdie, P.J.; Holmes, S.P. Bioconductor Workflow for Microbiome Data Analysis: From raw reads to community analyses. F1000Research 2016, 5, 1492. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.B.F.G.; Kindt, R.; Legendre, P.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.5-6. 2019. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 9 May 2021).
- Xu, J.; Xu, Y.; Wang, H.M.; Guo, C.S.; Qiu, H.Y.; He, Y.; Zhang, Y.; Li, X.C.; Meng, W. Occurrence of antibiotics and antibiotic resistance genes in a sewage treatment plant and its effluent-receiving river. Chemosphere 2015, 119, 1379–1385. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate—A practical and powerful approach to multiple testing. J. R. Stat. Soc. Series B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot 2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Schommer, V.A.; Wenzel, B.M.; Daroit, D.J. Anaerobic co-digestion of swine manure and chicken feathers: Effects of manure maturation and microbial pretreatment of feathers on methane production. Renew. Energy 2020, 152, 1284–1291. [Google Scholar] [CrossRef]
- Heinonen-Tanski, H.; Mohaibes, M.; Karinen, P.; Koivunen, J. Methods to reduce pathogen microorganisms in manure. Livest. Sci. 2006, 102, 248–255. [Google Scholar] [CrossRef]
- Chen, Z.; Jiang, X. Microbiological Safety of Chicken Litter or Chicken Litter-Based Organic Fertilizers: A Review. Agriculture 2014, 4, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Usman, K.; Khan, S.; Ghulam, S.; Khan, M.U.; Khan, N.; Khan, M.A.; Khalil, S.K. Sewage Sludge: An Important Biological Resource for Sustainable Agriculture and Its Environmental Implications. Am. J. Plant Sci. 2012, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.L.; Ortiz, R.; Steele, T.W.J.; Stuckey, D.C. Toxicants inhibiting anaerobic digestion: A review. Biotechnol. Adv. 2014, 32, 1523–1534. [Google Scholar] [CrossRef]
- López, A.; Rodríguez-Chueca, J.; Mosteo, R.; Gómez, J.; Ormad, M.P. Microbiological quality of sewage sludge after digestion treatment: A pilot scale case of study. J. Clean. Prod. 2020, 254, 120101. [Google Scholar] [CrossRef]
- Piterina, A.V.; Bartlett, J.; Pembroke, T.J. Evaluation of the removal of indicator bacteria from domestic sludge processed by Autothermal Thermophilic Aerobic Digestion (ATAD). Int. J. Environ. Res. Public Health 2010, 7, 3422–3441. [Google Scholar] [CrossRef] [Green Version]
- Lloret, E.; Pastor, L.; Martínez-Medina, A.; Blaya, J.; Pascual, J.A. Evaluation of the removal of pathogens included in the Proposal for a European Directive on spreading of sludge on land during autothermal thermophilic aerobic digestion (ATAD). Chem. Eng. J. 2012, 198, 171–179. [Google Scholar] [CrossRef]
- Abdallah, A.M.; Rashid, M.; Adroub, S.A.; Arnoux, M.; Ali, S.; van Soolingen, D.; Bitter, W.; Pain, A. Complete Genome Sequence of Mycobacterium phlei Type Strain RIVM601174. J. Bacteriol. 2012, 194, 3284–3285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghielmetti, G.; Friedel, U.; Scherrer, S.; Sarno, E.; Landolt, P.; Dietz, O.; Hilbe, M.; Zweifel, C.; Stephan, R. Non-tuberculous Mycobacteria isolated from lymph nodes and faecal samples of healthy slaughtered cattle and the abattoir environment. Transbound. Emerg. Dis. 2018, 65, 711–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahansepas, A.; Aghazadeh, M.; Rezaee, M.A.; Hasani, A.; Sharifi, Y.; Aghazadeh, T.; Mardaneh, J. Occurrence of Enterococcus faecalis and Enterococcus faecium in Various Clinical Infections: Detection of Their Drug Resistance and Virulence Determinants. Microb. Drug Resist. 2018, 24, 76–82. [Google Scholar] [CrossRef]
- Gu, G.Y.; Strawn, L.K.; Zheng, J.; Reed, E.A.; Rideout, S.L. Diversity and Dynamics of Salmonella enterica in Water Sources, Poultry Litters, and Field Soils Amended With Poultry Litter in a Major Agricultural Area of Virginia. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Alegbeleye, O.O.; Singleton, I.; Sant’Ana, A.S. Sources and contamination routes of microbial pathogens to fresh produce during field cultivation: A review. Food Microbiol. 2018, 73, 177–208. [Google Scholar] [CrossRef]
- Maffei, D.F.; Batalha, E.Y.; Landgraf, M.; Schaffner, D.W.; Franco, B.D.G.M. Microbiology of organic and conventionally grown fresh produce. Braz. J. Microbiol. 2016, 47, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.M.; Zhang, S.W.; Yuan, Z.W. Adoption of solid organic waste composting products: A critical review. J. Clean. Prod. 2020, 272, 122712. [Google Scholar] [CrossRef]
- Tran, D.T.Q.; Bradbury, M.I.; Van Ogtrop, F.F.; Bozkurt, H.; Jones, B.J.; McConchie, R. Environmental Drivers for Persistence of Escherichia coli and Salmonella in Manure-Amended Soils: A Meta-Analysis. J. Food Prot. 2020, 83, 1268–1277. [Google Scholar] [CrossRef]
- Zhang, Y.; Gu, A.Z.; Cen, T.Y.; Li, X.Y.; He, M.; Li, D.; Chen, J.M. Sub-inhibitory concentrations of heavy metals facilitate the horizontal transfer of plasmid-mediated antibiotic resistance genes in water environment. Environ. Pollut. 2018, 237, 74–82. [Google Scholar] [CrossRef]
- Li, J.; Phulpoto, I.A.; Zhang, G.; Yu, Z. Acceleration of emergence of E. coli antibiotic resistance in a simulated sublethal concentration of copper and tetracycline co-contaminated environment. AMB Express 2021, 11, 14. [Google Scholar] [CrossRef]
- Stalder, T.; Barraud, O.; Casellas, M.; Dagot, C.; Ploy, M.-C. Integron Involvement in Environmental Spread of Antibiotic Resistance. Front. Microbiol. 2012, 3, 119. [Google Scholar] [CrossRef] [Green Version]
- An, X.-L.; Chen, Q.-L.; Zhu, D.; Zhu, Y.-G.; Gillings, M.R.; Su, J.-Q. Impact of Wastewater Treatment on the Prevalence of Integrons and the Genetic Diversity of Integron Gene Cassettes. Appl. Environ. Microbiol. 2018, 84, e02766-17. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wang, P.L.; Yang, Q.X. Occurrence and diversity of antibiotic resistance in untreated hospital wastewater. Sci. Total Environ. 2018, 621, 990–999. [Google Scholar] [CrossRef]
- Conde-Cid, M.; Nunez-Delgado, A.; Fernandez-Sanjurjo, M.J.; Alvarez-Rodriguez, E.; Fernandez-Calvino, D.; Arias-Estevez, M. Tetracycline and Sulfonamide Antibiotics in Soils: Presence, Fate and Environmental Risks. Processes 2020, 8, 1479. [Google Scholar] [CrossRef]
- Granados-Chinchilla, F.; Rodríguez, C. Tetracyclines in Food and Feedingstuffs: From Regulation to Analytical Methods, Bacterial Resistance, and Environmental and Health Implications. J. Anal. Methods Chem. 2017, 2017, 1315497. [Google Scholar] [CrossRef]
- Ibrahim, M.; Ahmad, F.; Yaqub, B.; Ramzan, A.; Imran, A.; Afzaal, M.; Mirza, S.A.; Mazhar, I.; Younus, M.; Akram, Q.; et al. Current trends of antimicrobials used in food animals and aquaculture. In Antibiotics and Antimicrobial Resistance Genes in the Environment; Elsevier: Amsterdam, The Netherlands, 2020; pp. 39–69. [Google Scholar] [CrossRef]
- Taylor, P.; Reeder, R. Antibiotic use on crops in low and middle-income countries based on recommendations made by agricultural advisors. CABI Agric. Biosci. 2020, 1, 1–14. [Google Scholar] [CrossRef]
- Antunes, P.; Machado, J.; Sousa, J.C.; Peixe, L. Dissemination of sulfonamide resistance genes (sul1, sul2, and sul3) in Portuguese Salmonella enterica strains and relation with integrons. Antimicrob. Agents Chemother. 2005, 49, 836–839. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Shin, S.G.; Jang, H.M.; Kim, Y.B.; Lee, J.; Kim, Y.M. Characterization of antibiotic resistance genes in representative organic solid wastes: Food waste-recycling wastewater, manure, and sewage sludge. Sci. Total Environ. 2017, 579, 1692–1698. [Google Scholar] [CrossRef]
- Jiang, H.; Cheng, H.; Liang, Y.; Yu, S.; Yu, T.; Fang, J.; Zhu, C. Diverse Mobile Genetic Elements and Conjugal Transferability of Sulfonamide Resistance Genes (sul1, sul2, and sul3) in Escherichia coli Isolates From Penaeus vannamei and Pork From Large Markets in Zhejiang, China. Front. Microbiol. 2019, 10, 1787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domínguez, M.; Miranda, C.D.; Fuentes, O.; de la Fuente, M.; Godoy, F.A.; Bello-Toledo, H.; González-Rocha, G. Occurrence of Transferable Integrons and sul and dfr Genes Among Sulfonamide-and/or Trimethoprim-Resistant Bacteria Isolated From Chilean Salmonid Farms. Front. Microbiol. 2019, 10, 748. [Google Scholar] [CrossRef]
- Fyfe, C.; Grossman, T.H.; Kerstein, K.; Sutcliffe, J. Resistance to macrolide antibiotics in public health pathogens. Cold Spring Harb. Perspect. Med. 2016, 6, a025395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijesekara, P.N.K.; Kumbukgolla, W.W.; Jayaweera, J.A.A.S.; Rawat, D. Review on Usage of Vancomycin in Livestock and Humans: Maintaining Its Efficacy, Prevention of Resistance and Alternative Therapy. Vet. Sci. 2017, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Murray, R.; Tien, Y.-C.; Scott, A.; Topp, E. The impact of municipal sewage sludge stabilization processes on the abundance, field persistence, and transmission of antibiotic resistant bacteria and antibiotic resistance genes to vegetables at harvest. Sci. Total Environ. 2019, 651, 1680–1687. [Google Scholar] [CrossRef]
- Heuer, H.; Smalla, K. Manure and sulfadiazine synergistically increased bacterial antibiotic resistance in soil over at least two months. Environ. Microbiol. 2007, 9, 657–666. [Google Scholar] [CrossRef]
- Heuer, H.; Schmitt, H.; Smalla, K. Antibiotic resistance gene spread due to manure application on agricultural fields. Curr. Opin. Microbiol. 2011, 14, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Feng, K.; Li, S.; Zhang, Y.; Chen, H.; Yin, H.; Xu, M.; Deng, Y. Exploring abundance, diversity and variation of a widespread antibiotic resistance gene in wastewater treatment plants. Environ. Int. 2018, 117, 186–195. [Google Scholar] [CrossRef]
- Schwarz, J.; Thiele-Bruhn, S.; Eckhardt, K.U.; Schulten, H.R. Sorption of Sulfonamide Antibiotics to Soil Organic Sorbents: Batch Experiments with Model Compounds and Computational Chemistry. ISRN Soil Sci. 2012, 2012, 159189. [Google Scholar] [CrossRef] [Green Version]
- Sukul, P.; Lamshöft, M.; Zühlke, S.; Spiteller, M. Sorption and desorption of sulfadiazine in soil and soil-manure systems. Chemosphere 2008, 73, 1344–1350. [Google Scholar] [CrossRef]
- Tien, Y.-C.; Drury, C.; Reynolds, D.; Topp, E. Enrichment of antibiotic resistance genes in soil receiving composts derived from swine manure, yard wastes, or food wastes, and evidence for multi-year persistence of swine Clostridium spp. Can. J. Microbiol. 2017, 64, 201–208. [Google Scholar] [CrossRef]
- Cerqueira, F.; Matamoros, V.; Bayona, J.M.; Berendonk, T.U.; Elsinga, G.; Hornstra, L.M.; Pina, B. Antibiotic resistance gene distribution in agricultural fields and crops. A soil-to-food analysis. Environ. Res. 2019, 177, 108608. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Hu, H.W.; Chen, Q.L.; Singh, B.K.; Yan, H.; Chen, D.L.; He, J.Z. Transfer of antibiotic resistance from manure-amended soils to vegetable microbiomes. Environ. Intern. 2019, 130, 104912. [Google Scholar] [CrossRef]
- Zhu, Y.G.; Johnson, T.A.; Su, J.Q.; Qiao, M.; Guo, G.X.; Stedtfeld, R.D.; Hashsham, S.A.; Tiedje, J.M. Diverse and abundant antibiotic resistance genes in Chinese swine farms. Proc. Natl. Acad. Sci. USA 2013, 110, 3435–3440. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.Y.; Li, G.X.; Chang, J.L.; Kong, Y.L.; Jiang, T.; Wang, J.N.; Yuan, J. Enrichment of antibiotic resistance genes after sheep manure aerobic heap composting. Bioresour. Technol. 2021, 323, 124620. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Chen, Z.; Zhang, F.; Liu, Y.; Yan, T. Abundance, diversity and mobility potential of antibiotic resistance genes in pristine Tibetan Plateau soil as revealed by soil metagenomics. FEMS Microbiol. Ecol. 2020, 96, fiaa172. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Shen, M.; Zhang, Y.; Zhu, D.; Wang, H.; Zhao, Y.; Kang, Y. The changes in antibiotic resistance genes during 86 years of the soil ripening process without anthropogenic activities. Chemosphere 2021, 266, 128985. [Google Scholar] [CrossRef] [PubMed]
- Szekeres, E.; Chiriac, C.M.; Baricz, A.; Szőke-Nagy, T.; Lung, I.; Soran, M.L.; Rudi, K.; Dragos, N.; Coman, C. Investigating antibiotics, antibiotic resistance genes, and microbial contaminants in groundwater in relation to the proximity of urban areas. Environ. Pollut. 2018, 236, 734–744. [Google Scholar] [CrossRef]
- Sabri, N.A.; Schmitt, H.; Van der Zaan, B.; Gerritsen, H.W.; Zuidema, T.; Rijnaarts, H.H.M.; Langenhoff, A.A.M. Prevalence of antibiotics and antibiotic resistance genes in a wastewater effluent-receiving river in the Netherlands. J. Environ. Chem. Eng. 2020, 8, 102245. [Google Scholar] [CrossRef]
- Jackson, C.R.; Fedorka-Cray, P.J.; Barrett, J.B.; Ladely, S.R. Effects of Tylosin Use on Erythromycin Resistance in Enterococci Isolated from Swine. Appl. Environ. Microbiol. 2004, 70, 4205–4210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.; Nawaz, M.; Khan, S.; Steele, R. Detection and characterization of erythromycin-resistant methylase genes in Gram-positive bacteria isolated from poultry litter. Appl. Microbiol. Biotechnol. 2002, 59, 377–381. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| ARG | Manure 2012 | Manure 2015 | Sludge 2012 | Sludge 2015 |
|---|---|---|---|---|
| tetA/16S-rRNA | 1.46 × 10−2 ± 4.90 × 10−4 | 2.69 × 10−2 ± 1.69 × 10−2 | 1.19 × 10−2 ± 1.47 × 10−3 | 2.06 × 10−3 ± 1.18 × 10−3 |
| tetW/16S-rRNA | 3.59 × 10−3 ± 4.43 × 10−4 | 4.93 × 10−2 ± 6.36 × 10−3 | 6.36 × 10−3 ± 1.06 × 10−3 | 4.42 × 10−4 ± 2.91 × 10−5 |
| sul1/16S-rRNA | 7.97 × 10−3 ± 2.02 × 10−3 | 2.69 × 10−2 ± 1.69 × 10−2 | 2.59 × 10−3 ± 2.36 × 10−3 | 3.02 × 10−2 ± 9.55 × 10−3 |
| sul2/16S-rRNA | 2.60 × 10−4 ± 1.16 × 10−5 | 4.56 × 10−4 ± 6.94 × 10−5 | 2.19 × 10−4 ± 6.83 × 10−5 | 2.08 × 10−4 ± 3.07 × 10−5 |
| ermB/16S-rRNA | 5.53 × 10−6 ± 2.25 × 10−6 | 9.35 × 10−4 ± 5.57 × 10−4 | 2.24 × 10−3 ± 8.60 × 10−4 | 1.35 × 10−2 ± 3.45 × 10−3 |
| vanA/16S-rRNA | 2.05 × 10−5 ± 1.92 × 10−5 | 2.29 × 10−5 ± 1.01 × 10−5 | 2.02 × 10−3 ± 3.31 × 10−3 | 3.44 × 10−6 ± 3.00 × 10−6 |
| Intl1/16S-rRNA | 3.51 × 10−2 ± 1.82 × 10−3 | 2.96 × 10−1 ± 3.2 × 10−2 | 4.19 × 10−1 ± 1.69 × 10−1 | 5.30 × 10−2 ± 2.67 × 10−2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stiborova, H.; Kracmarova, M.; Vesela, T.; Biesiekierska, M.; Cerny, J.; Balik, J.; Demnerova, K. Impact of Long-Term Manure and Sewage Sludge Application to Soil as Organic Fertilizer on the Incidence of Pathogenic Microorganisms and Antibiotic Resistance Genes. Agronomy 2021, 11, 1423. https://doi.org/10.3390/agronomy11071423
Stiborova H, Kracmarova M, Vesela T, Biesiekierska M, Cerny J, Balik J, Demnerova K. Impact of Long-Term Manure and Sewage Sludge Application to Soil as Organic Fertilizer on the Incidence of Pathogenic Microorganisms and Antibiotic Resistance Genes. Agronomy. 2021; 11(7):1423. https://doi.org/10.3390/agronomy11071423
Chicago/Turabian StyleStiborova, Hana, Martina Kracmarova, Tereza Vesela, Marta Biesiekierska, Jindrich Cerny, Jiri Balik, and Katerina Demnerova. 2021. "Impact of Long-Term Manure and Sewage Sludge Application to Soil as Organic Fertilizer on the Incidence of Pathogenic Microorganisms and Antibiotic Resistance Genes" Agronomy 11, no. 7: 1423. https://doi.org/10.3390/agronomy11071423
APA StyleStiborova, H., Kracmarova, M., Vesela, T., Biesiekierska, M., Cerny, J., Balik, J., & Demnerova, K. (2021). Impact of Long-Term Manure and Sewage Sludge Application to Soil as Organic Fertilizer on the Incidence of Pathogenic Microorganisms and Antibiotic Resistance Genes. Agronomy, 11(7), 1423. https://doi.org/10.3390/agronomy11071423