
The Young Stages of the Cannonball Jellyfish (Stomolophus sp. 2) from the Central Gulf of California (Mexico)


1
Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México (UNAM), Mexico City 04510, Mexico
2
Centro de Investigaciones Biológicas del Noroeste S.C (CIBNOR), Guaymas 85460, Mexico
3
Departamento de Zoologia, Instituto de Biociências, Universidade de São Paulo, Rua do Matão 101 trav. 14, São Paulo 05508-090, SP, Brazil
4
Centro de Biologia Marinha, Universidade de São Paulo, Rodovia Dr Manoel Hipólito do Rego km 131.5, São Sebastião 11612-109, SP, Brazil
*
Author to whom correspondence should be addressed.
Diversity 2021, 13(6), 229; https://doi.org/10.3390/d13060229
Submission received: 27 April 2021
/
Revised: 8 May 2021
/
Accepted: 10 May 2021
/
Published: 24 May 2021
(This article belongs to the Special Issue Patterns and Ecology of Jellyfish in Marine Environment)
Abstract
:Exploitation of the cannonball jellyfish (Stomolophus sp. 2) is increasing in Mexico and USA due to successful fisheries associated with seasonal blooms in coastal areas. Previously, it was proposed that such blooms could be identified by recognizing the presence of young stages in the water. In our work, we aim to describe the young stages (ephyra and metaephyra) found in the Las Guásimas lagoon, Sonora, Mexico. The description of specimens is based on photographs, drawings, and morphological measurements aimed at helping in the early detection of blooming events.
1. Introduction
In Western countries, jellyfish are usually associated with bad memories, mostly due to the stings these marine animals can inflict on humans [1,2,3,4]; however, in Eastern countries, jellyfish are synonymous with a fine dish or delicacy [5,6,7,8], and eventually have been associated with many medicinal properties [9]. The use of jellyfish as food has long been recognized in many places. It is only recently that Western countries have recognized its importance as an economic product [10,11,12]. In many places, jellyfish fisheries have been developed, and then declined [13], due to different factors (e.g., fluctuation of stocks or lower interest of fishery companies), but some countries are fortunate to have certain valuable species in their territorial waters. Among the most valued features are the thickness and weight of the gelatinous part [5,9,11]. The rhizostome genus Stomolophus L. Agassiz, 1860 morphologically embraces such features and is widely distributed on both sides of the American continents [14,15].
Historically, five species of Stomolophus have been described (S. agaricus, S. chunii, S. collaris, S. fritillaria, and S. meleagris) [15,16,17,18]. However, most records have mention only S. meleagris, with some of those possibly based on misidentifications [16,19,20]. Scyphozoan systematics currently considers only two of the species to be valid (S. meleagris and S. fritillarius), both from the Atlantic Ocean [15,21]. A recent study [22] revealed cryptic diversity within this jellyfish genus, hypothesizing five undescribed species from the Pacific Ocean. As mentioned by Gómez-Daglio and Dawson [22] and Getino-Mamet et al. [23], the existence of different species within the genus Stomolophus is an ongoing investigation that is not yet entirely conclusive, nor has it reached a broad scientific consensus. Although Gómez-Daglio and Dawson [22] proposed the existence of different species, the holotypes mentioned in their research have not yet been described; in addition, the distribution frequency of significant genetic distances that the authors reported only detected two discontinuities [24].
Jellyfish of the genus Stomolophus are commercially exploited by many different countries in the Americas [11]. In Mexico, especially in the Sonora region, there are favorable conditions for conducting jellyfish fisheries. Species in the area present marked seasonality (winter and spring) and can only be exploited during a few months [10]. Little is known about the polyp, planula, and ephyra stages of the jellyfish S. meleagris. Despite their commercial importance, much less is known about their distribution and settlement areas, which is information that is more than desirable for management purposes, as they are a reservoir for subsequent outbreaks, although this is still difficult to prove [25,26].
The identification of the early stage (ephyrae) can provide evidence for forecasting jellyfish blooms [27,28], which is importance for effective management of all aspects of the fishery.
Previous studies have described the ephyra stage of S. meleagris of some Atlantic populations [16,29,30], however, for the Pacific region, there is only a Panamanian study [31]. Although the species have been mentioned in several studies in the Gulf of California [24,32,33,34,35,36], little is known about how to recognize and identify the young medusae.
The goals of this work were to identify the young stage (ephyrae) found in Las Guásimas Bay and to describe the different developmental stages of the ephyrae. We highlight that, in the study area, the only adult jellyfish species found is Stomolophus sp. 2, which is heavily fished during the season.
2. Materials and Methods
The study was conducted in Las Guásimas lagoon, Sonora, Mexico (Figure 1) (27°49′–55′ N 110°29′–45′ W). The lagoon has an area of 51 km2, with an opening of 2 km to the Gulf of California. There is seasonal variation in temperatures (winter/summer) from 17 to 34 °C, with a salinity of 36 to 41 ups.
We performed superficial zooplankton tows (250 microns) every month from 2014 to 2016, inside and outside the lagoon. The samples were preserved in 4% formaldehyde solution (buffered with sodium borate) the samples were inspected and all ephyrae were sorted. All individuals were measured following Straehler-Pohl and Jarms [27], Straehler-Pohl et al. [37], and Holst [28] (Figure 2). Total body diameter (TBD), central disk diameter (CDD), total marginal lappet length (TMLL), lappet stem length (LStL), rhopalial lappet length (RLL), as well as the body proportions, i.e., CDD/TBD × 100, RLL/TMLL × 100, TMLL/TBD × 100, and LStL/TMLL × 100 were calculated (Figure 2).
We also observed the shape of the “rhopalial lappet”, “rhopalial canal”, and “velar canal” of every specimen to record the differences in each stage. We additionally observed the gastrovascular system; the development of rhopaliar and velar canals; and changes in mouth, oral arms, and scapulets morphology in order to distinguish the different developmental stages of the ephyrae.
3. Results
Ephyrae were only found during the winter and spring (November to May). We examined a total of 28 samples and discovered 154 specimens of Stomolophus sp. 2. They were sorted in three different stages, which are described below.
3.1. Features of the Ephyrae of Stomolophus sp. 2
Ephyrae have eight pairs of rhopalial lappets which are lancet-shaped (sometimes sword-like, broadly oval, or breadknife-shaped). The rhopalia arise from the base of the rhopalial canal and have a small peduncle; there is a statocyst but no ocellus. The canal is divided, and each branch enters each of the lappets. The mouth has four lips, and the central stomach has three to four gastric filaments. In the earliest stage, the size varies from 0.6 to 1.7 mm (TBD), from 0.2 to 0.9 mm (CDD) (39–51% of TBD), and the proportion of the lappets (LStT/RLL-to-TMLL) are 50% each (Table 1).
Although we mentioned that the ephyrae had eight pairs of lappets, we found individuals with six or nine pairs. The shape of the lappets also varied (cf. Straehler-Pohl and Jarms [27]) and we found four types; the lancet-shaped lappets (65.8% for Stage I and 49.4% for Stage II) were predominant. The shape of the velar canal presented five different forms, i.e., “table-like”, “triangular”, “rhombic”, “rounded”, and “round spoon” with the most abundant being the “round spoon” in Stage I. All specimens were transparent in color.
3.2. Different Stages of the Development of Stomolophus sp. 2
In Stage I (Figure 3), the smallest and youngest form are found, due to its lower complexity. The gastric system is very simple and poorly developed, with only three to four gastric filaments that sometimes reach the outside through the small manubrium. The rhopalial lappets are lancet-like, and the rhopalial canals are also shortly bifurcated and lancet shaped. The mouth is small and rounded, there is little distinction of the mouth lips, and there are no traces of scapulets.
In Stage II (Figure 4), the gastric cavity is a somewhat more developed. The velar canals are anchor-like (growing to the sides and connecting with the rhopaliar ones), increasing the area. Velar lappets start to develop. Scapulets start to develop on the manubrium above the mouth arms, looking like small hollow tube-like protrusions. The mouth develops and the lips tend to form a cross-shaped contour with well-developed papillae on the margin.
In Stage III (Figure 5), the internal gastric space increases in complexity, and it is easy to see all the connections of velar and rhopaliar canals. Between the rhopaliar and velar canals, it is possible to see additional canals forming, with a centripetal growing pattern. The umbrella also increases in size, and the connection of the central stomach with the radial canals (either velar or rhopaliar) is more evident. The mouth lips enlarge and branch, and the distance between them increases, the papillae (digitata) can be seen on both sides of the lips. The scapulets are more developed, with branches and papillae (digitata) developing.
4. Discussion
Detecting early jellyfish stages is an important step for recognizing possible jellyfish blooms which can have ecological and commercial impacts, causing some problems or benefits to local populations. In NW Mexico, it is quite relevant to detect young jellyfish stages because when the species Stomolophus sp. 2 (cannonball jellyfish) is blooming, it is exploited economically by local fishermen, and any early warnings can provide time enough for the local people to organize themselves and be ready when the bloom season starts. In general, the presence of ephyrae extends over the winter and spring seasons, during which time the presence of adult organisms can be observed.
Although there are some previous descriptions of ephyrae of the species S. meleagris, there are no detailed measurements, or body proportions, or even details of the gastric cavity development. Such descriptions are exclusively based on Atlantic [29,30] or Panamanian specimens [31] which seem to be a different species [22,23]. It is important to state that our data refer to the species occurring in the central area of the Gulf of California, comprising the blue variety recognized as Stomolophus sp. 2.
It is widely known that wild collected gelatinous animals suffer due to the collecting techniques, and the same applies to young ephyrae [27,28]. However, in this study, although we collected some damaged specimens, we were fortunate to obtain some individuals in relatively good shape that allowed us to provide a detailed description of the species. It was possible to describe several characteristics that are important for their recognition (lappets shape and gastric system), in very good conditions.
Even though we found four different shapes of the rhopaliar lappets, the lancet-like shape was the most common, found in 66% of the specimens inspected. Although some authors [27] have mentioned that environmental conditions (either in the sea or in lab conditions) can alter the morphology, the natural phenotypic plasticity in the species is not completely discounted. In other rhizostome species, the lancet-like shape of the ephyrae lappets was also observed, i.e., Phyllorhiza punctata, Chrysaora fuscescens, and C. lactea [27,37].
For the identification of medusae, body proportions are usually considered instead of absolute size, because such measurements tend to vary in specimens of the same species due to the nutritional condition and age of the polyps [27,38,39]. For instance, total diameters (TBD) vary with temperature [27,40] with differences ranging from 0.4 to 1.5–2.5 mm in P. punctata in temperatures of 25–28 °C. Additionally, in another example with the species C. hysoscella [27], the total diameter varied from 1.6 to 3.4 mm with temperatures of 5, 10, 15, or 22 °C. In our case, the ephyrae used for the description in this study were collected in temperatures from 18 to 22 °C (although they were found in a larger spectrum of 17.7–28.9 °C) with a TBD from 6.5 to 1.9 mm, opposite to what was expected, as both measurements were found in 21.2 °C. The body proportion CDD (related to TBD) was 39% in 23 °C and 51% in 21 °C; such variations were also reported for other species such as Mastigias papua and P. punctata when changing from 25 to 28 °C [27]. Similar results were found in Aurelia, which in the polyp stage releases ephyrae with larger CDD sizes at high temperatures (28 °C) as compared with at lower temperatures (20 °C) [40]. Thus, these results reinforce that temperature is an important factor in the development of scyphozoan species [40]. The proportions LStL and RLL (related to TMLL) on average both presented 50% as was observed in Rhizostoma octopus, M. papua, and Cassiopea (but we should highlight that our data showed up to 33% variation). Similar ephyrae of the Pelagiidae, Cyaneidae, and Aureliinae presented a ratio of RLL to TMLL of 58–60%, 56–57%, and 42–48%, respectively, [37] as compared with this study of 47–65%.
In this study, we only found transparent organisms, without any type of coloration. Transparent stains with brown color and dark red pigments in the sensory organs have previously been reported for ephyrae of S. meleagris [31]. However, these were organisms found in South Carolina in the Atlantic, while our specimens were from the Pacific Ocean.
Concerning the gastric system development, the Stage I ephyrae have a simple cavity without any connections between the radial (rhopaliar and velar) canals. Later on, in Stage II ephyrae, the velar canals develop and start to produce lateral branches that reach the rhopaliar canals, thus, forming a ring canal. The velar lappets start to form and continue to enlarge. In Stage III, ephyrae show a more developed canal system with new radial canals between the rhopaliar and velar canals.
Following the development of the gastric system is quite important because, in adult medusae, the identification is mostly based on this feature. In younger stages, determining gastric development brings greater reliability to ephyra identification [27,41,42].
Although, at first glance, the ephyrae of Stomolophus sp. 2 described by us have some similarities to other species such as A. aurita and C. hysocella; our samples allowed us to perform quite a good identification and also to provide proper descriptions of the different developmental stages. In general, the growth stages are related to the age of the animals awith a sequential series of changes, highlighting a general pattern of development. Further comparative studies would provide more data and contribute to more accurate identification of ephyrae from many different areas.
Author Contributions
Conceptualization, L.C.G.-S., J.L.-M. and A.C.M.; data curation, L.C.G.-S., J.L.-M. and A.C.M.; formal analysis L.C.G.-S. and A.C.M.; funding acquisition, J.L.-M.; investigation, L.C.G.-S., J.L.-M. and A.C.M.; project administration, J.L.-M. and A.C.M.; resources, J.L.-M. and A.C.M.; supervision, J.L.-M. and A.C.M.; visualization, L.C.G.-S., J.L.-M. and A.C.M.; writing—original draft, L.C.G.-S., J.L.-M. and A.C.M.; writing—review & editing, L.C.G.-S., J.L.-M. and A.C.M. All authors have read and agreed to the published version of the manuscript.
Funding
ACM was supported by the FAPESP (2015/21007-9) and the CNPq (309440/2019-0). This investigation was financially supported by the SEMARNAT-2018-1-A3-S-77965 project. This is a contribution from NP-BioMar USP.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The ephyra samples and the data obtained from them are available in the Fisheries laboratory of the Guaymas Unit of the Centro de Investigaciones Biológicas del Noroeste, S.C. in Mexico (https://www.cibnor.gob.mx/unidades-foraneas/guaymas).
Acknowledgments
We thank CIBNOR, SC. and Instituto de Biociências (Universidade de São Paulo) for providing facilities for this project. We also thank Eloisa Herrera Valdivia, Rufino Morales Azpeitia, Francisco Álvarez Tello, Edgar Gamero-Mora, and Gisele R. Tiseo for help in their labs. We also thank the referees for their comments, suggestions, and corrections that improved our manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Purcell, J.E.; Uye, S.I.; Lo, W.T. Anthropogenic causes of jellyfish blooms and their direct consequences for humans: A review. Mar. Ecol. Prog. Ser. 2007, 350, 153–174. [Google Scholar] [CrossRef]
- Brotz, L. Changing Jellyfish Populations: Trends in Large Marine Ecosystems. Master’s Thesis, The University of British Columbia, Vancouver, BC, Canada, 2011. [Google Scholar] [CrossRef]
- Kim, D.; Seo, J.; Yoon, W.; Suh, Y. Estimating the economic damage caused by jellyfish to fisheries in Korea. Fish. Sci. 2012, 78, 1147–1152. [Google Scholar] [CrossRef]
- Purcell, J.E. Jellyfish and Ctenophore Blooms Coincide with Human Proliferations and Environmental Perturbations. Ann. Rev. Mar. Sci. 2012, 4, 209–235. [Google Scholar] [CrossRef]
- Hsieh, Y.-H.P.; Rudloe, J. Potential of utilizing jellyfish as food in Western countries. Trends Food Sci. Technol. 1994, 5, 225–229. [Google Scholar] [CrossRef]
- Omori, M.; Nakano, E. Jellyfish fisheries in southeast Asia. Hydrobiologia 2001, 451, 19–26. [Google Scholar] [CrossRef]
- Li, J.; Hsieh, Y.-H.P. Traditional Chinese food technology and cuisine. Asia Pac. J. Clin. Nutr. 2004, 13, 147–155. [Google Scholar] [PubMed]
- Kitamura, M.; Omori, M. Synopsis of edible jellyfishes collected from Southeast Asia, with notes on jellyfish fisheries. Plankton Benthos Res. 2010, 5, 106–118. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, Y.-H.P.; Leong, F.-M.; Rudloe, J. Jellyfish as food. Hydrobiologia 2001, 451, 11–17. [Google Scholar] [CrossRef]
- López-Martínez, J.; Álvarez-Tello, J. The jellyfish fishery in Mexico. Agric. Sci. 2013, 4, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Brotz, L.; Schiariti, A.; López-Martínez, J.; Álvarez-Tello, J.; Peggy Hsieh, Y.H.; Jones, R.P.; Quiñones, J.; Dong, Z.; Morandini, A.C.; Preciado, M.; et al. Jellyfish fisheries in the Americas: Origin, state of the art, and perspectives on new fishing grounds. Rev. Fish Biol Fish. 2017, 27, 1–29. [Google Scholar] [CrossRef]
- Torri, L.; Tuccillo, F.; Bonelli, S.; Piraino, S.; Leone, A. The Attitudes of Italian Consumers towards Jellyfish as Novel Food. Food Qual. Prefer. 2020, 79, 103782. Available online: http://www.sciencedirect.com/science/article/pii/S0950329318310346 (accessed on 27 August 2019). [CrossRef]
- Dong, Z.; Liu, D.; Keesing, J.K. Contrasting Trends in Populations of Rhopilema esculentum and Aurelia aurita in Chinese Waters. In Jellyfish Blooms; Pitt, K., Lucas, C., Eds.; Springer: Dordrecht, Holland, 2014; pp. 207–218. [Google Scholar] [CrossRef]
- Kramp, P.L. Zoogeographical studies on Rhizostomeae. Vidensk. Medd. Fra Dansk Naturh. Foren. 1970, 133, 7–30. [Google Scholar]
- Jarms, G.; Morandini, A.C. World Atlas of Jellyfish; Dölling und Galitz Verlag: Hamburg, Germany, 2019; p. 816. [Google Scholar]
- Mayer, A.G. Medusae of the world III: The Scyphomedusae. Publ. Carnegie Inst. Wash. 1910, 109, 499–735. [Google Scholar]
- Kramp, P.L. A revision of Ernst Haeckel’s determinations of a collection of medusae belonging to the Zoological Museum of Copenhagen. Deep Sea Res. 1955, 3, 149–168. [Google Scholar]
- Kramp, P.L. Synopsis of the medusae of the world. J. Mar. Biol. Assoc. UK 1961, 40, 7–469. [Google Scholar] [CrossRef]
- Uchida, T. Some medusae of the central Pacific. J. Fac. Sci. Hokkaido Univ. Ser. 6 Zool. 1947, 9, 297–319. [Google Scholar]
- Mianzan, H.W.; Cornelius, P.F.S. Cubomedusae and Scyphomedusae. In South Atlantic Zooplankton; Boltovskoy, D., Ed.; Backhuys Publishers: Leiden, Holland, 1999; Volume 1, pp. 513–559. [Google Scholar]
- Banha, T.N.S.; Morandini, A.C.; Rosário, R.P.; Martinelli Filho, J.E. Scyphozoan jellyfish (Cnidaria, Medusozoa) from Amazon coast: Distribution, temporal variation and length–weight relationship. J. Plankton Res. 2020, 42, 767–778. [Google Scholar] [CrossRef]
- Gómez-Daglio, L.; Dawson, M.N. Species richness of jellyfishes (Scyphozoa: Discomedusae) in the Tropical Eastern Pacific: Missed taxa, molecules, and morphology match in a biodiversity hotspot. Invertebr. Syst. 2017, 31, 635–663. [Google Scholar] [CrossRef]
- Getino Mamet, L.N.; Gómez-Daglio, L.; García-De Léon, F.J. High genetic differentiation in the edible cannonball jellyfish (Cnidaria: Scyphozoa: Stomolophus spp.) from the Gulf of California, Mexico. Fish. Res. 2019, 219, 105328. [Google Scholar] [CrossRef]
- Lopez-Martinez, J.; Arzola-Sotelo, E.A.; Nevarez-Martinez, M.O.; Alvarez-Tello, F.J.; Morales-Bojorquez, E. Modeling growth on the cannonball jellyfish Stomolophus meleagris based on a multi-model inference approach. Hydrobiologia 2020, 847, 1399–1422. [Google Scholar] [CrossRef]
- Boero, F.; Bouillon, J.; Gravili, C.; Miglietta, M.; Parsons, T.; Piraino, S. Gelatinous plankton: Irregularities rule the world (sometimes). Mar. Ecol. Prog. Ser. 2008, 18, 299–310. [Google Scholar] [CrossRef]
- Gibbons, M.; Boero, F.; Brotz, L. We should not assume that fishing jellyfish will solve our jellyfish problem. ICES J. Mar. Sci. 2016, 73, 1012–1018. [Google Scholar] [CrossRef] [Green Version]
- Straehler-Pohl, I.; Jarms, G. Identification key for young ephyrae: A first step for early detection of jellyfish blooms. Hydrobiologia 2010, 645, 3–21. [Google Scholar] [CrossRef]
- Holst, S. Morphology and development of benthic and pelagic life stages of North Sea jellyfish (Scyphozoa, Cnidaria) with special emphasis on the identification of ephyra stages. Mar. Biol. 2012, 159, 2707–2722. [Google Scholar] [CrossRef]
- Calder, D.R. Life history of the cannonball jellyfish, Stomolophus meleagris L. Agassiz, 1860 (Scyphozoa, Rhizostomida). Biol. Bull. 1982, 162, 149–162. [Google Scholar] [CrossRef]
- Gómez-Aguirre, S. Larva éfira y diferenciación de Stomolophus meleagris (Scyphozoa Rhizostomeae) en plancton de lagunas costeras de Tabasco, México. An. Inst. Biol. Univ. Nac. Autón. México Ser. Zool. 1991, 62, 383–389. [Google Scholar]
- Stiasny, G. Papers from Dr. Th. Mortensen’s Pacific Expedition 1914-16. XII. Zur Kenntnis der Entwicklung von Stomolophus meleagris L. Agassiz. Vidensk Medd Fra Dansk Naturh Foren 1922, 73, 499–511. [Google Scholar]
- Burrola-Sánchez, M.S.; López-Martínez, J.; Padilla-Arredondo, G.; Urias-Laborin, D.; Padilla-Serrato, J.G. Influencia de los procesos costeros sobre la distribución de la medusa bola de cañón Stomolophus melegris (Agassiz, 1860) en el Golfo de California. In La Variabilidad Ambiental y Las Pesquerías De México; López-Martínez, J., Ed.; Comisión Nacional de Acuacultura y Pesca: Mazatlán, México, 2008; pp. 156–177. [Google Scholar]
- López-Martínez, J.; Rodríguez-Romero, J. Primer registro de la asociación del jurelillo negro Hemicaranx zelotes Gilbert (Pisces: Carangidae) con la medusa bala de cañón Stomolophus meleagris Agassiz (Scyphozoa: Rhizostomatidae) en Bahía de Kino, Golfo de California. Hidrobiológica 2008, 18, 173–176. [Google Scholar]
- Nevárez-López, C.A.; Váldez-Holguín, J.E.; Hernández-Saavedra, N.Y. Caracterización Genética de los fenotipos de la medusa “Bola de (Stomolophus meleagris, L. AGASSIZ 1862) en las Guásimas, Sonora. In Biotecnología Marina; XII Congreso Nacional de Biotecnología y Bioingeniería y VII Simposio Internacional de Producción de Alcoholes y Levaduras: Acapulco, Guerrero, México, 2009. [Google Scholar]
- Carvalho-Saucedo, L.; García-Domínguez, F.; Rodriguez-Jaramillo, C.; López-Martínez, J. Variación lipídica en los ovocitos de la medusa Stomolophus meleagris (Scyphozoa: Rhizostomeae), durante el desarrollo gonádico, en la laguna Las Guásimas, Sonora, México. Rev. Biol. Trop. 2010, 58, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Álvarez-Tello, J.; López-Martínez, J.; Lluch-Cota, D. Trophic spectrum and feeding pattern of cannonball jellyfish Stomolophus meleagris (Agassiz, 1862) from central Gulf of California. J. Mar. Biol. Assoc. UK 2015, 6, 1217–1227. [Google Scholar] [CrossRef]
- Straehler-Pohl, I.; Widmer, C.L.; Morandini, A.C. Characterizations of juvenile stages of some semaeostome Scyphozoa (Cnidaria), with recognition of a new family (Phacellophoridae). Zootaxa 2011, 2741, 1–37. [Google Scholar] [CrossRef]
- Russell, F.S. The Medusae of the British Isles II, Pelagic Scyphozoa with a Supplement to the First Volume on Hydromedusae; Cambridge University Press: Cambridge, UK, 1970; pp. 1–284. [Google Scholar]
- Fu, Z.; Shibata, M.; Makake, R.; Ikeda, H.; Uye, S. Body size reduction under starvation, and the point of no return, in ephyrae of the moon jellyfish Aurelia aurita. Mar. Ecol. Prog. Ser. 2014, 510, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Chiaverano, L.M.; Graham, W.M. Morphological plasticity in Aurelia polyps, with subsequent effects on asexual fecundity and morphology of young medusae. Mar. Ecol. Prog. Ser. 2017, 582, 79–92. [Google Scholar] [CrossRef]
- Morandini, A.C.; Silveira, F.L.; Jarms, G. The life cycle of Chrysaora lactea Eschscholtz, 1829 (Cnidaria, Scyphozoa) with notes on the scyphistoma stage of three other species. Hydrobiologia 2004, 530/531, 347–354. [Google Scholar] [CrossRef]
- Widmer, C.L. Life cycle of Chrysaora fuscescens (Cnidaria: Scyphozoa) and a key to sympatric ephyrae. Pac. Sci. 2008, 62, 71–82. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Collecting area, Las Guásimas lagoon, Sonora, Mexico, and central Gulf of California.
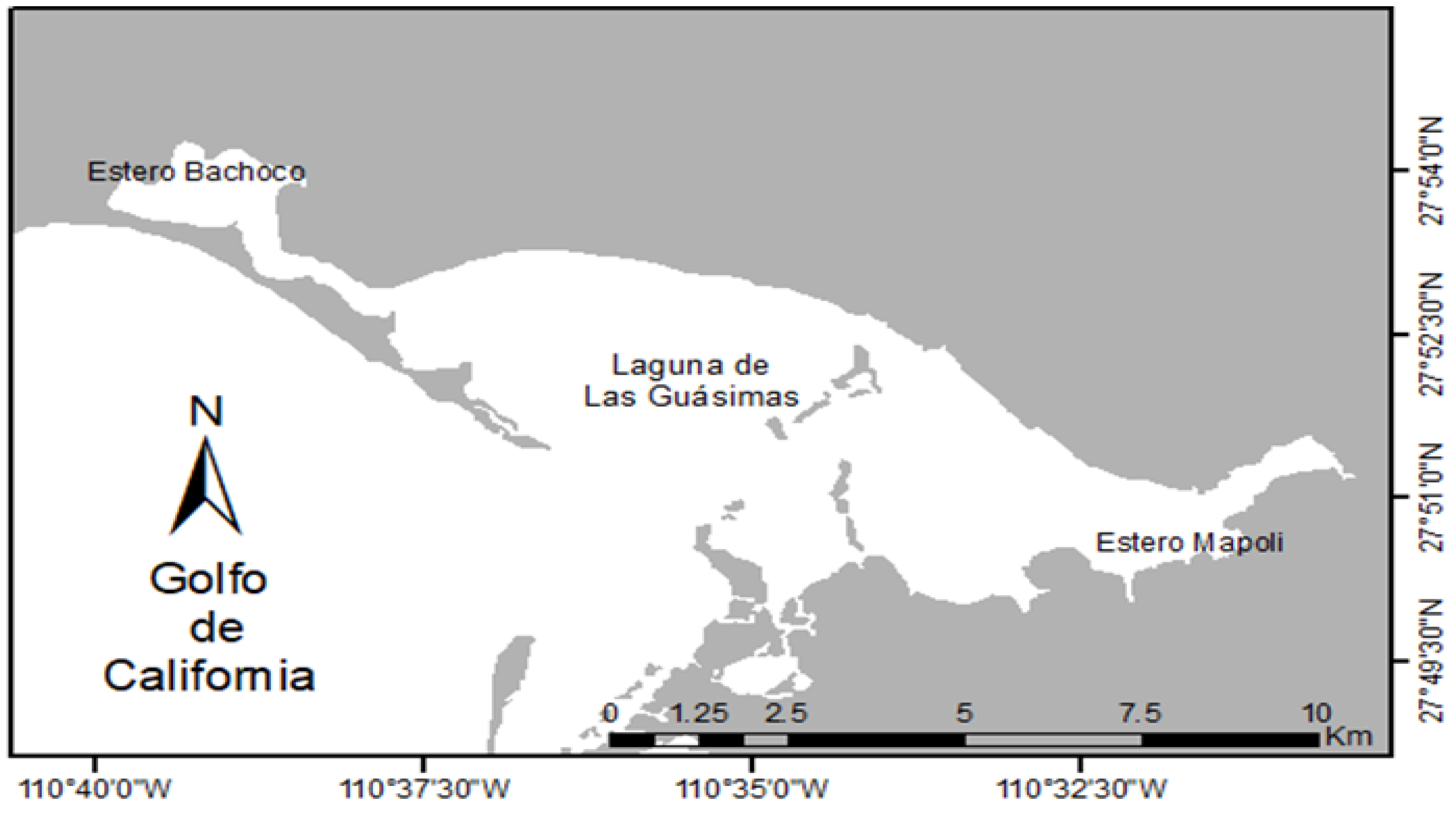
Figure 2.
Ephyrae of Stomolophus sp. 2, and measurements taken following Straehler-Pohl and Jarms [27] and Straehler-Pohl et al. [37].
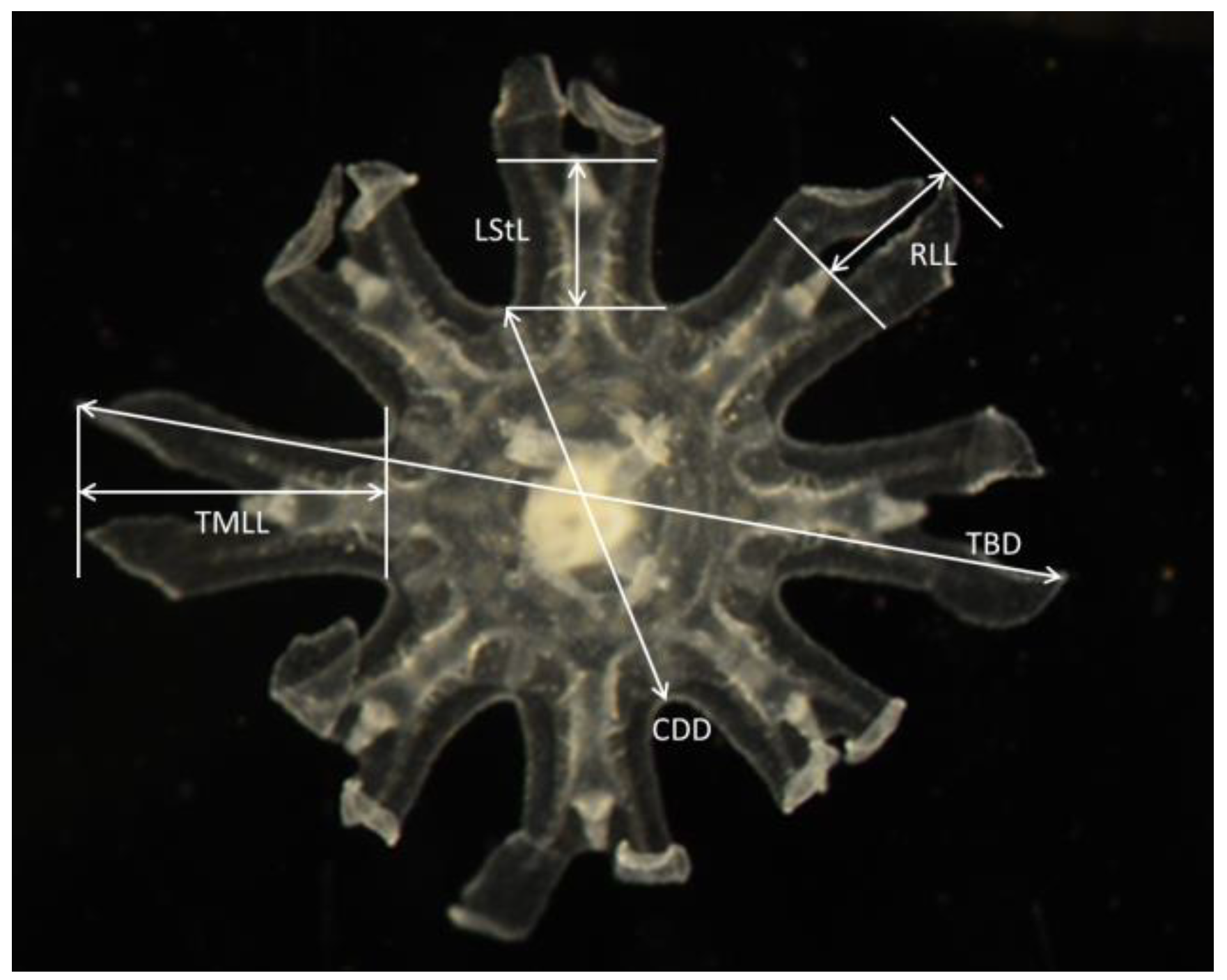
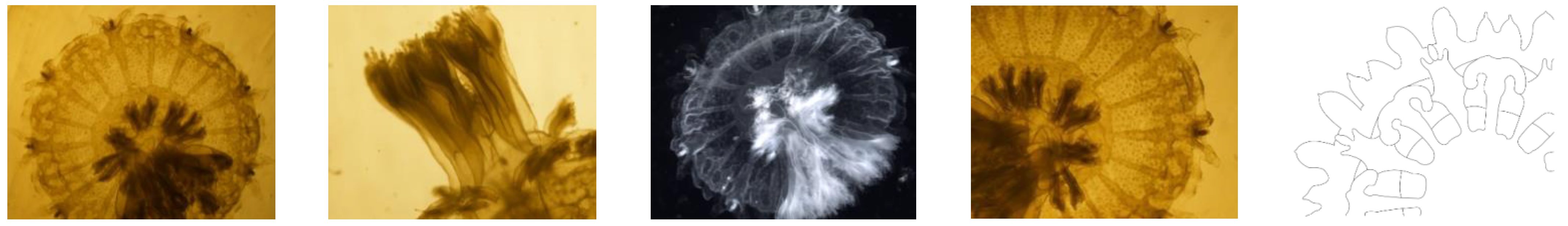
Figure 3.
Stage I Ephyrae (Stomolophus sp. 2). Without scapulets, mouth poorly developed (only mouth lips distinguishable, without papillae or faintly developed), gastric system poorly developed with only velar and rhopaliar canals, four gastric filaments, lancet-shaped lappets. No velar lappets.
Figure 3.
Stage I Ephyrae (Stomolophus sp. 2). Without scapulets, mouth poorly developed (only mouth lips distinguishable, without papillae or faintly developed), gastric system poorly developed with only velar and rhopaliar canals, four gastric filaments, lancet-shaped lappets. No velar lappets.
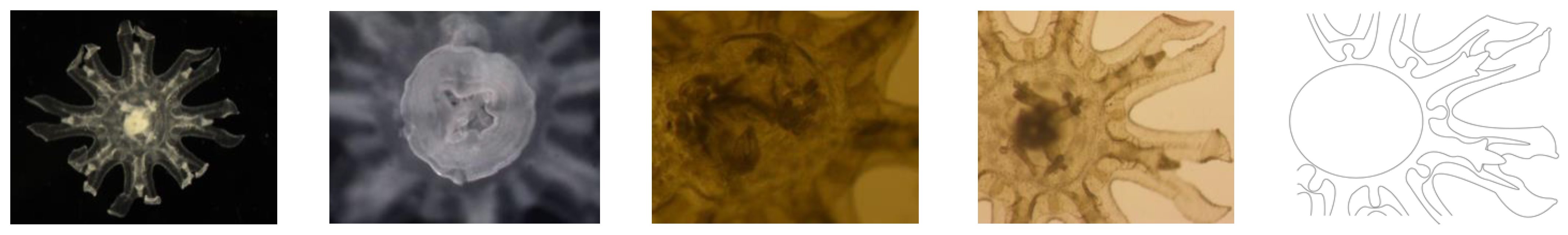
Figure 4.
Stage II Ephyrae (Stomolophus sp. 2). Scapulets starting to develop. Mouth increasing in size and with papillae at the margin. Gastric system developing with connection of rhopaliar and velar canals forming a ring canal at the rim. Rhopaliar lappets are still lancet shaped, but becoming more rhomboid, velar lappets appearing between rhopaliar ones.
Figure 4.
Stage II Ephyrae (Stomolophus sp. 2). Scapulets starting to develop. Mouth increasing in size and with papillae at the margin. Gastric system developing with connection of rhopaliar and velar canals forming a ring canal at the rim. Rhopaliar lappets are still lancet shaped, but becoming more rhomboid, velar lappets appearing between rhopaliar ones.
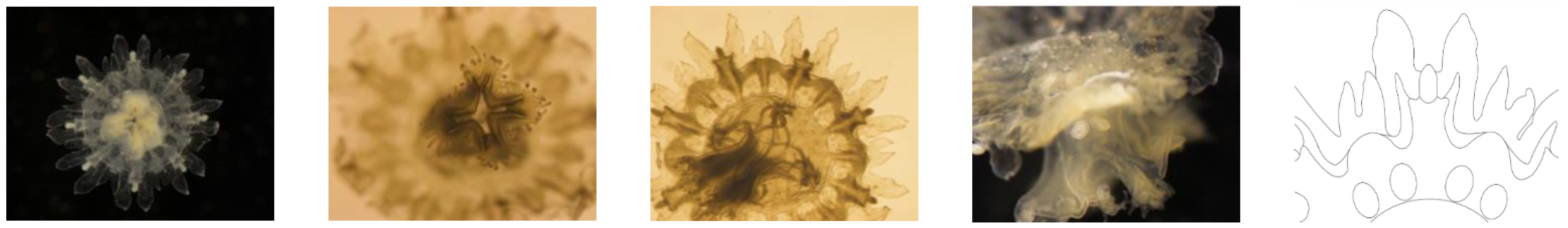
Figure 5.
Stage III Ephyrae (Stomolophus sp. 2). Increased canal system, stomach connected to the margin via radial canals (rhopaliar and velar ones) that are connected to each other by a ring canal; additional radial canals developing centripetally. All marginal lappets becoming rounder and wider. Mouth lips elongating and already branched, with papillae (digitata) at the inner and outer rim.
Figure 5.
Stage III Ephyrae (Stomolophus sp. 2). Increased canal system, stomach connected to the margin via radial canals (rhopaliar and velar ones) that are connected to each other by a ring canal; additional radial canals developing centripetally. All marginal lappets becoming rounder and wider. Mouth lips elongating and already branched, with papillae (digitata) at the inner and outer rim.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Proportions of body sizes in ephyrae of Stomolophus sp. 2. CDD, central disk diameter; TBD, total body diameter; TMLL, total marginal lappet length; RLL, rhopalial lappet length; LStL, lappet stem length.
Table 1.
Proportions of body sizes in ephyrae of Stomolophus sp. 2. CDD, central disk diameter; TBD, total body diameter; TMLL, total marginal lappet length; RLL, rhopalial lappet length; LStL, lappet stem length.
| Specimen | CDD/TBD × 100 | TMLL/TBD × 100 | RLL/TMLL × 100 | LStL/TMLL × 100 |
|---|---|---|---|---|
| 1 | 41.66 | 29.58 | 60.46 | 50.55 |
| 2 | 48.41 | 29.30 | 51.93 | 52.80 |
| 3 | 37.71 | 33.72 | 54.70 | 49.91 |
| 4 | 54.29 | 26.87 | 53.59 | 57.13 |
| 5 | 53.54 | 29.67 | 50.33 | 46.63 |
| 6 | 44.78 | 26.45 | 65.66 | 68.67 |
| 7 | 45.05 | 32.57 | 53.52 | 57.78 |
| 8 | 48.01 | 30.56 | 51.57 | 48.43 |
| 9 | 39.46 | 29.84 | 47.18 | 42.91 |
| 10 | 48.75 | 27.87 | 62.19 | 48.06 |
| 11 | 41.58 | 32.42 | 57.94 | 47.84 |
| 12 | 49.19 | 30.98 | 55.57 | 46.78 |
| 13 | 42.32 | 28.39 | 52.66 | 45.94 |
| 14 | 44.24 | 28.61 | 57.89 | 42.83 |
| 15 | 38.31 | 32.18 | 54.68 | 43.53 |
| 16 | 45.68 | 27.98 | 57.20 | 49.69 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gómez-Salinas, L.C.; López-Martínez, J.; Morandini, A.C. The Young Stages of the Cannonball Jellyfish (Stomolophus sp. 2) from the Central Gulf of California (Mexico). Diversity 2021, 13, 229. https://doi.org/10.3390/d13060229
AMA Style
Gómez-Salinas LC, López-Martínez J, Morandini AC. The Young Stages of the Cannonball Jellyfish (Stomolophus sp. 2) from the Central Gulf of California (Mexico). Diversity. 2021; 13(6):229. https://doi.org/10.3390/d13060229
Chicago/Turabian StyleGómez-Salinas, Laura Cristina, Juana López-Martínez, and André Carrara Morandini. 2021. "The Young Stages of the Cannonball Jellyfish (Stomolophus sp. 2) from the Central Gulf of California (Mexico)" Diversity 13, no. 6: 229. https://doi.org/10.3390/d13060229
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.