
Functional Connectome of the Human Brain with Total Correlation





Abstract
1. Introduction
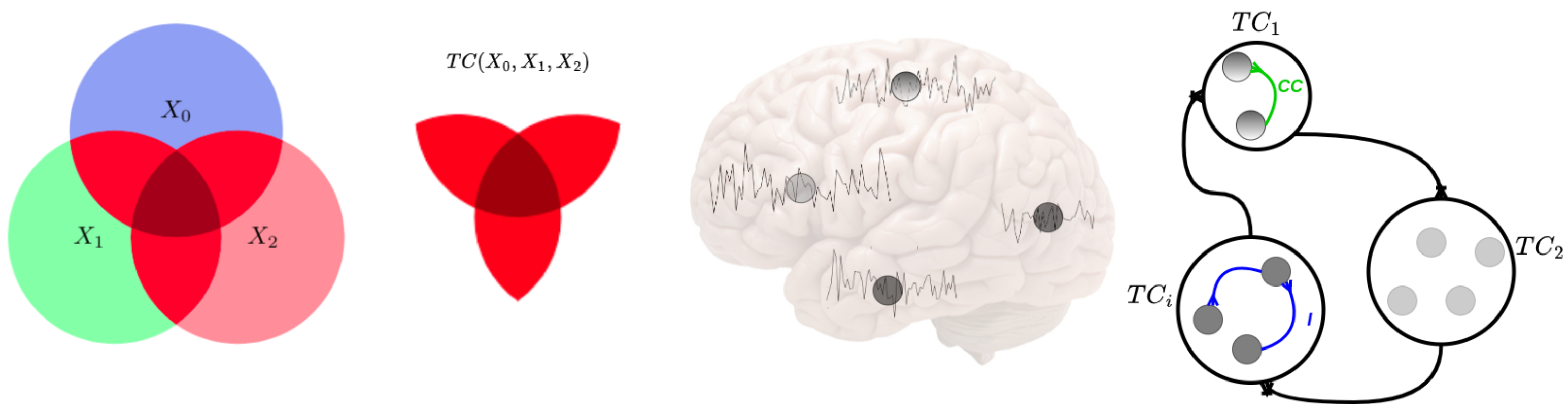
2. Total Correlation as Neural Connectivity Descriptor
2.1. Definitions and Preliminaries
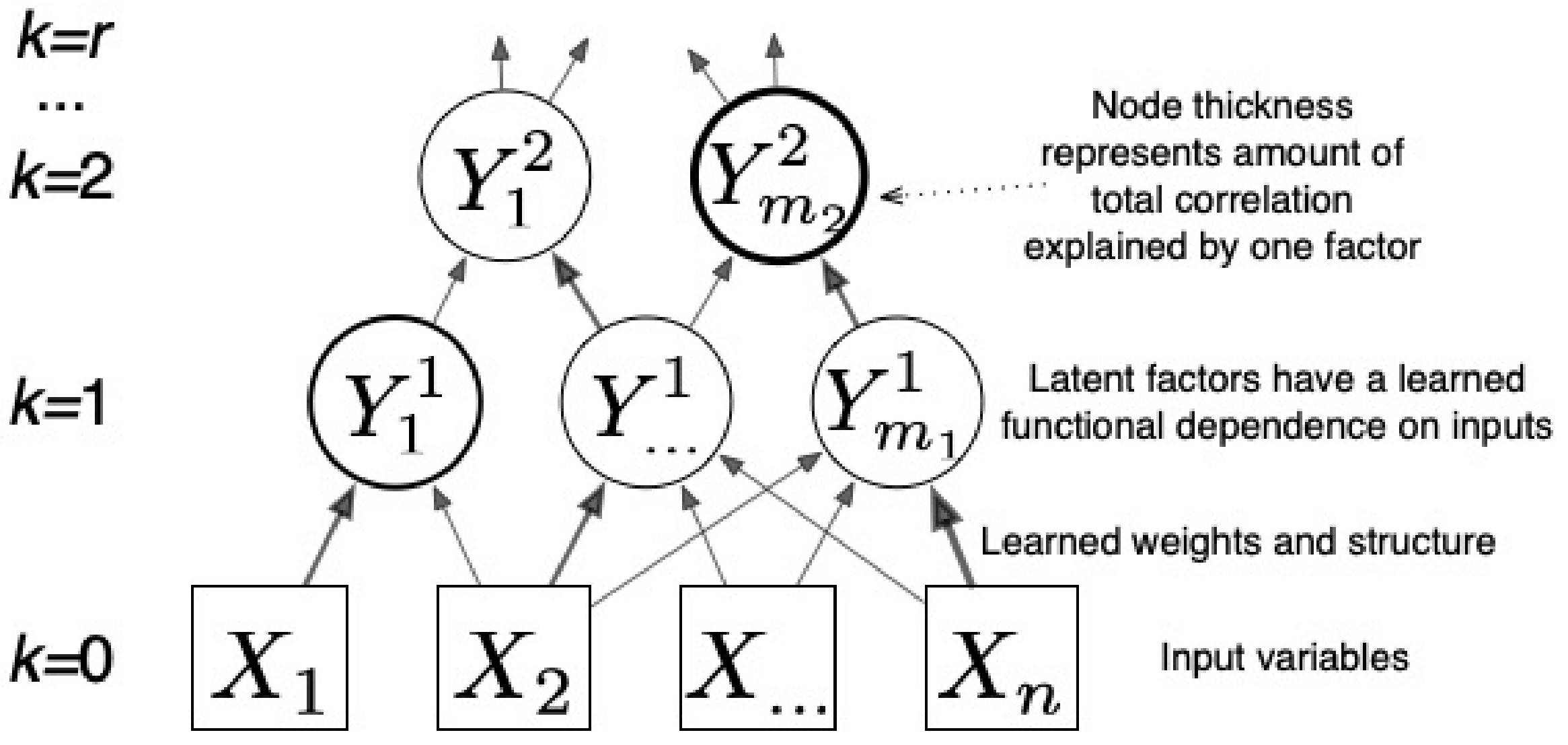
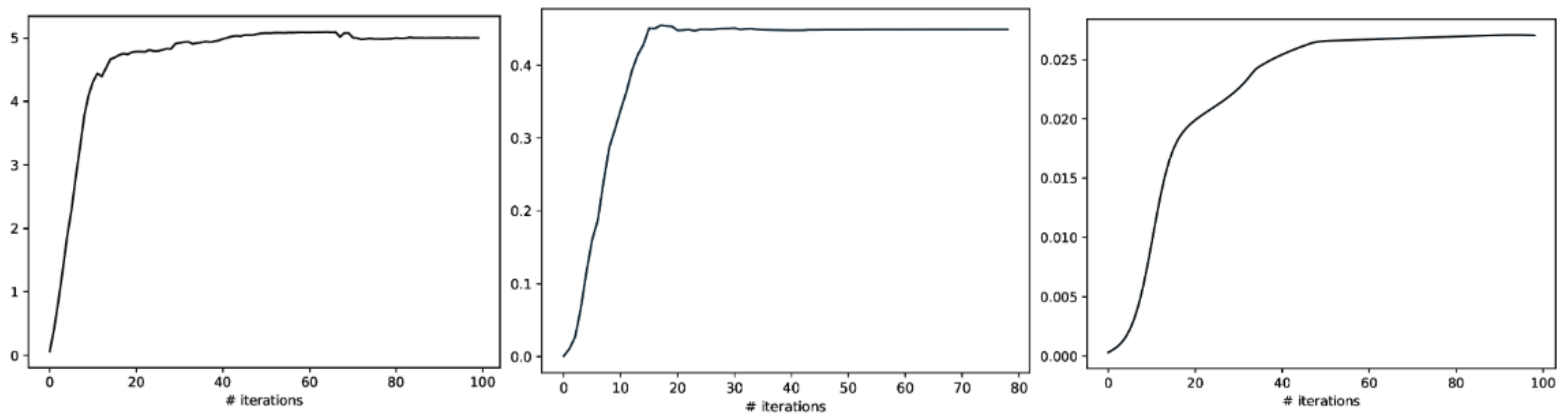
2.2. Total Correlation Estimated from CorEx
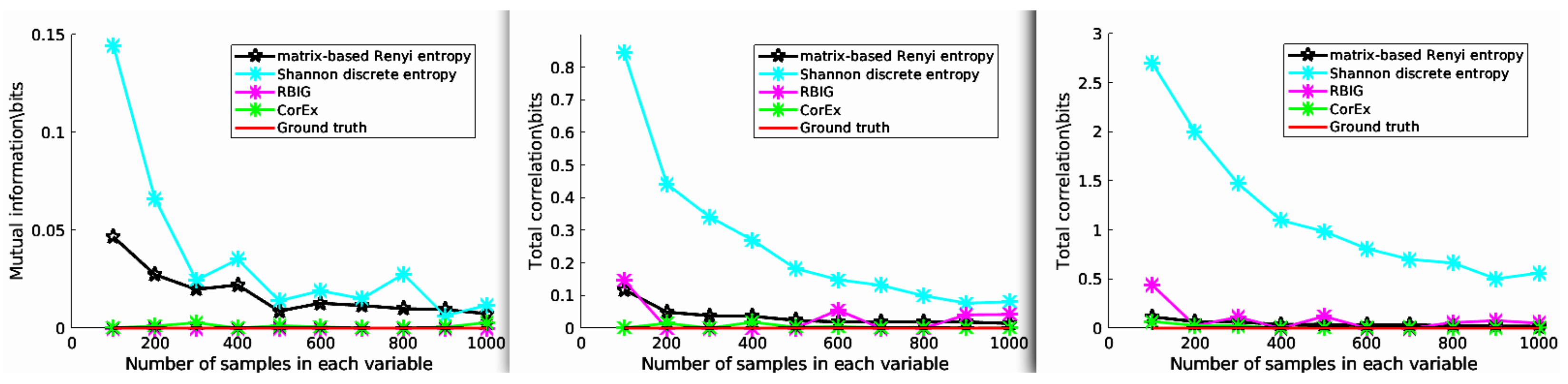
3. Experiment 1: Total Correlation for Independent Mixtures
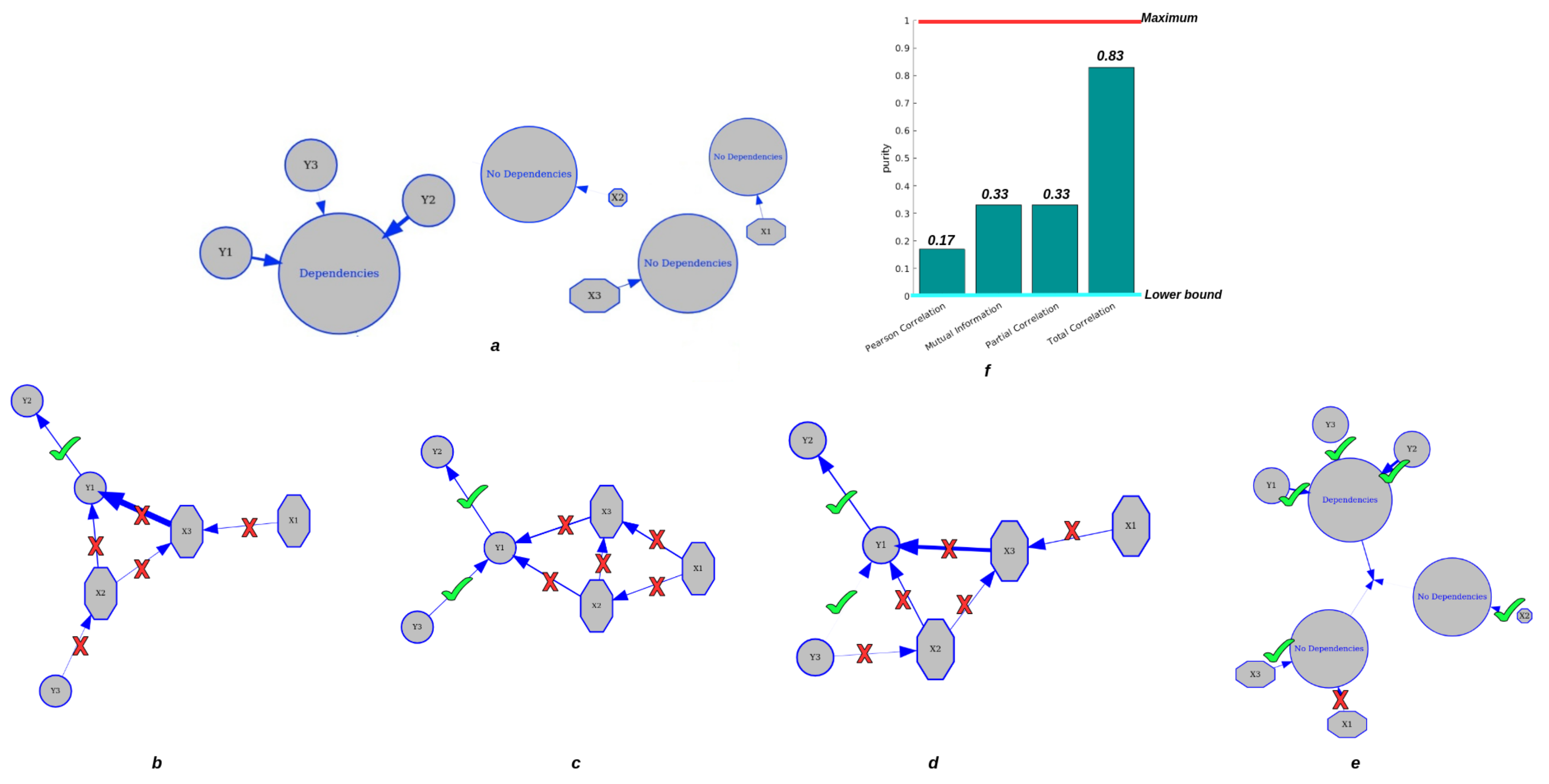
4. Experiment 2: Clustering by Total Correlation for Dependent and Independent Mixtures
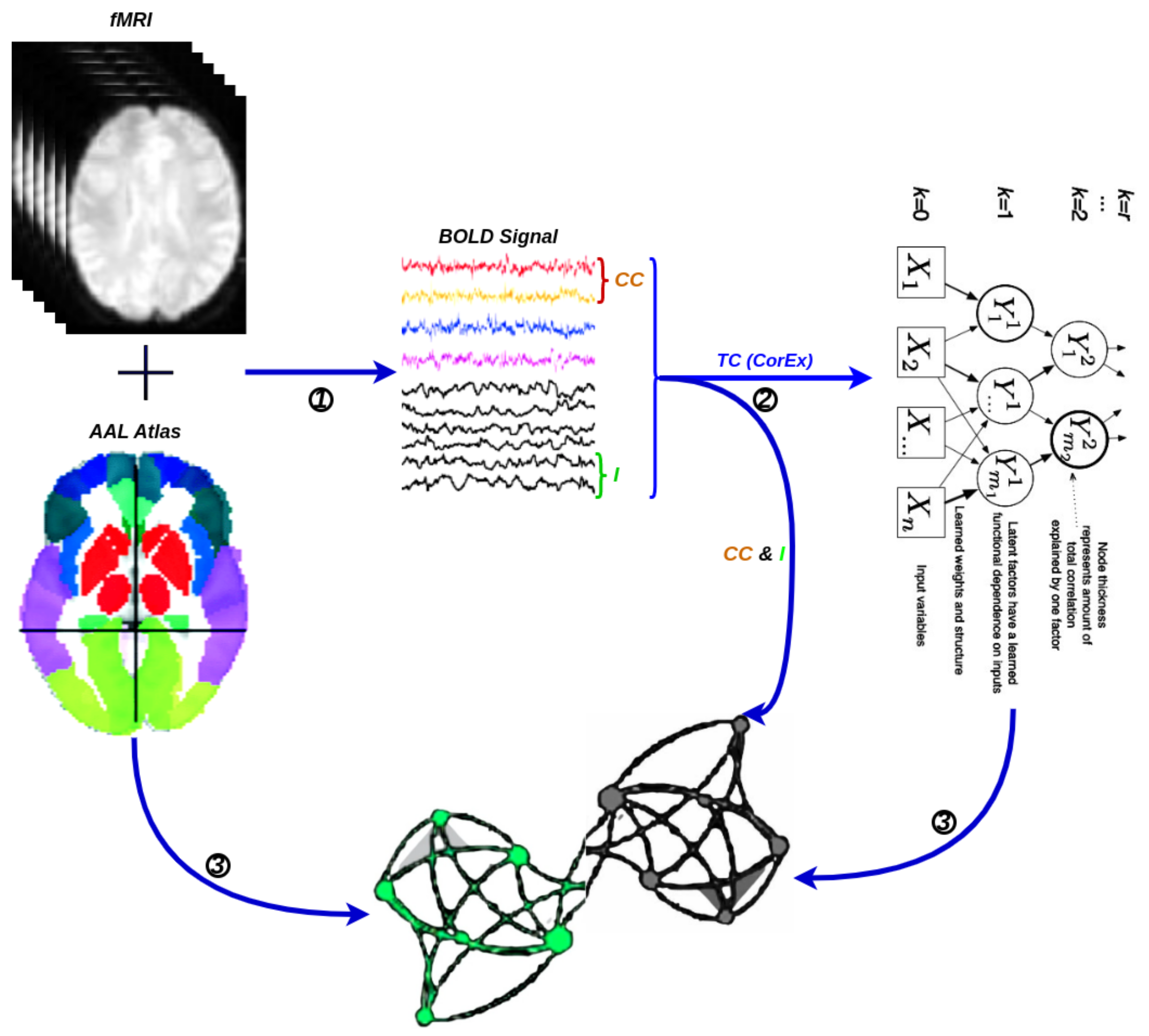
5. Experiment 3: Brain Functional Connectivity Analysis Using Total Correlation
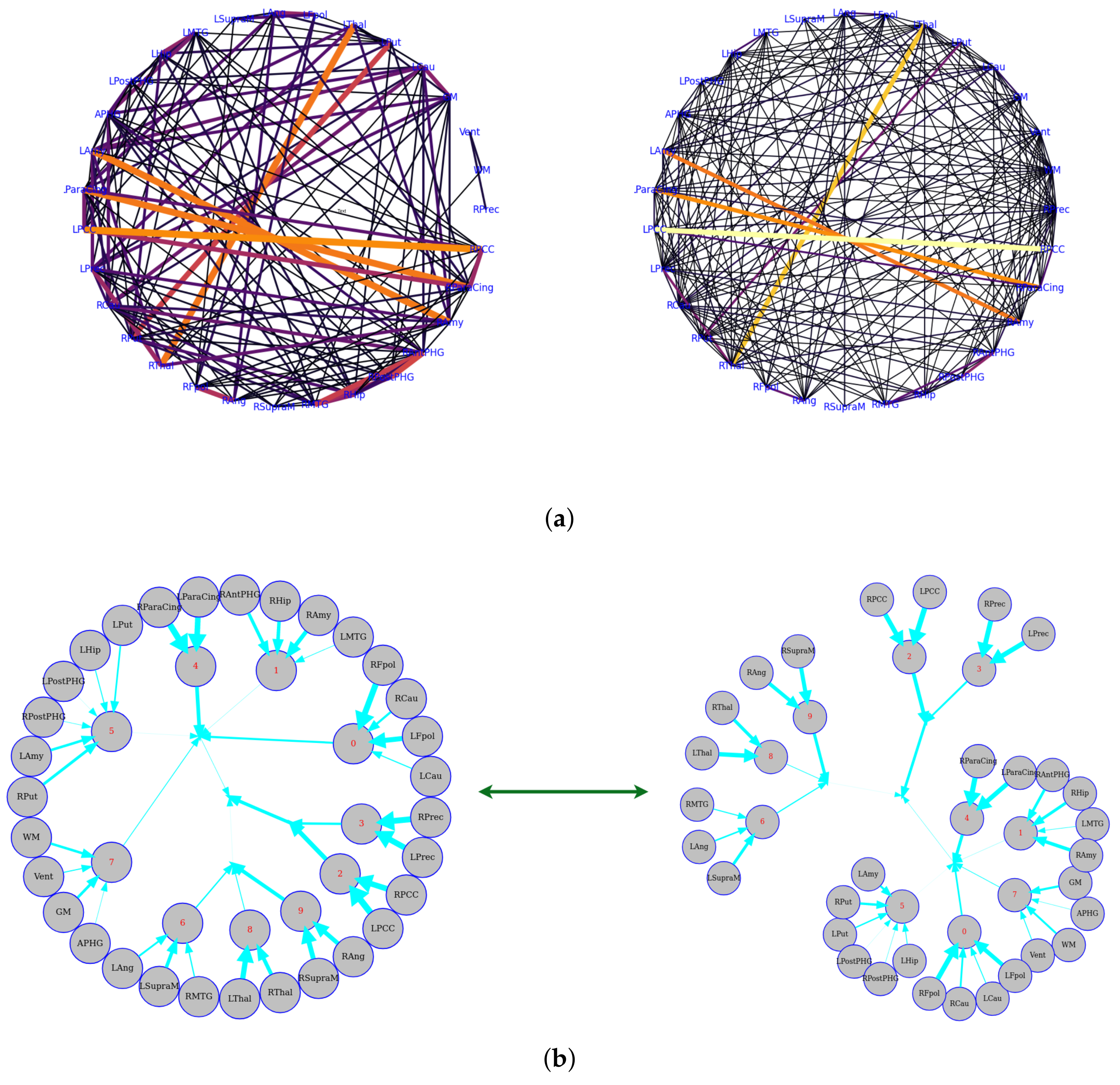
5.1. First Total-Correlation-Based Clustering Example from fMRI Data
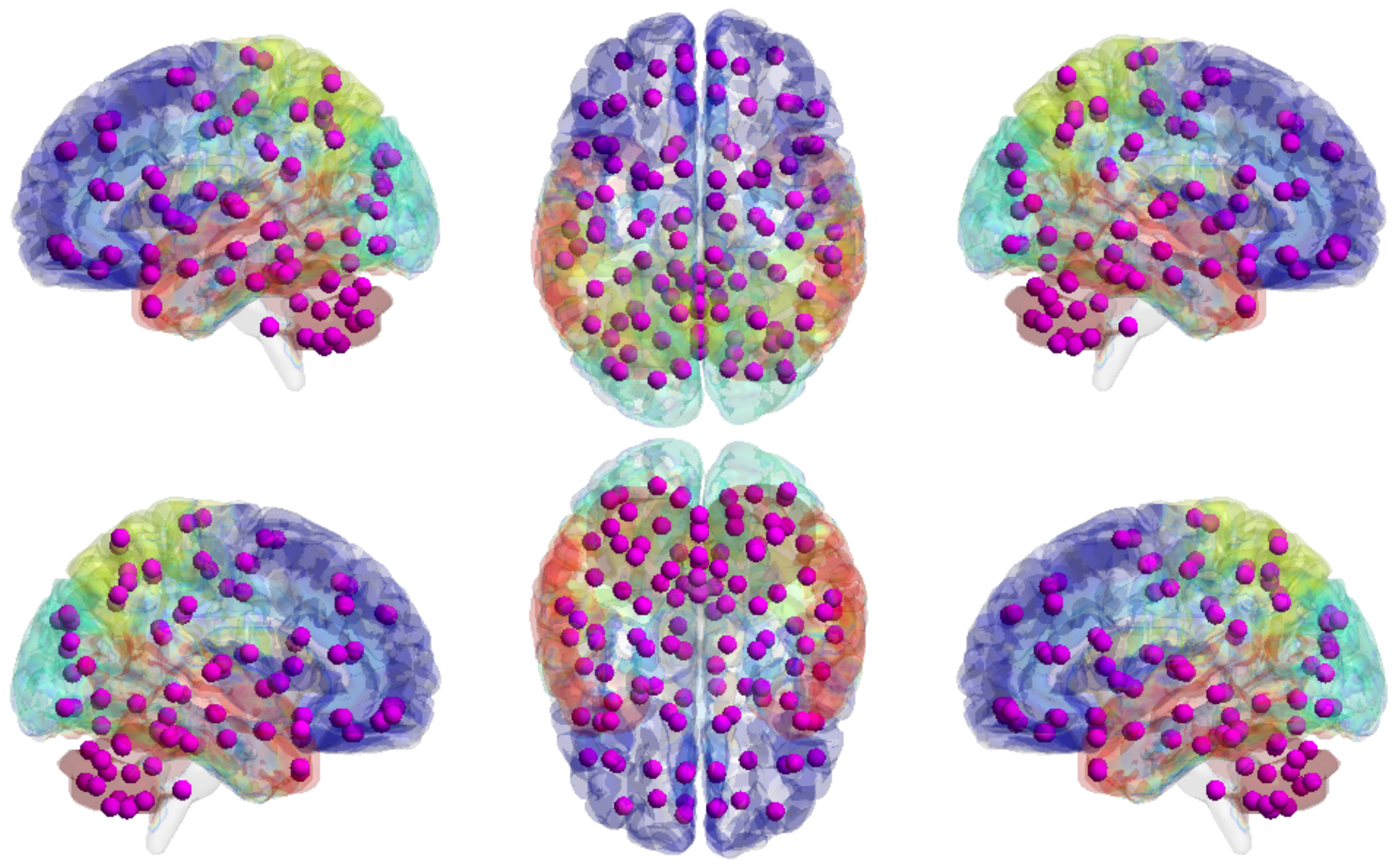
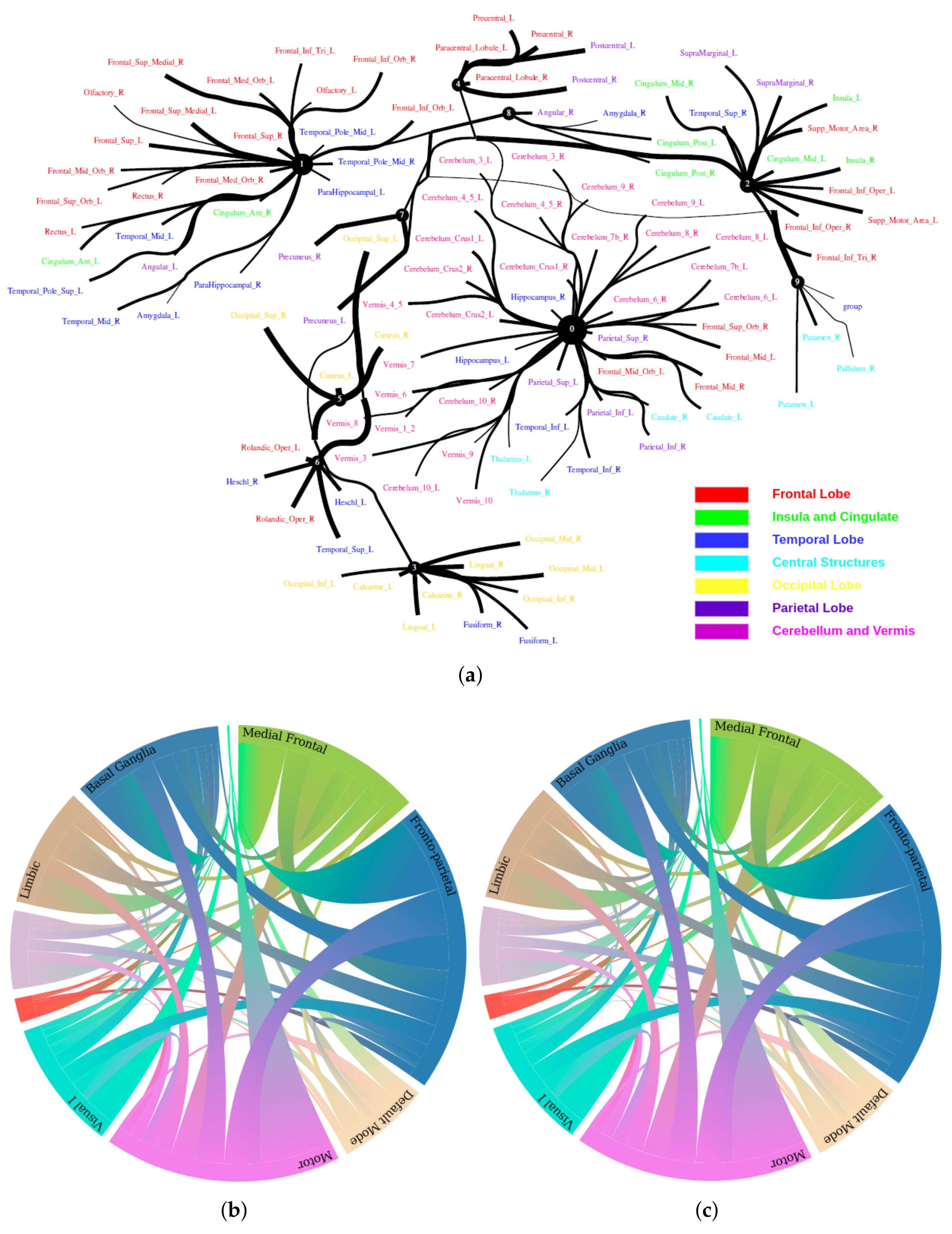
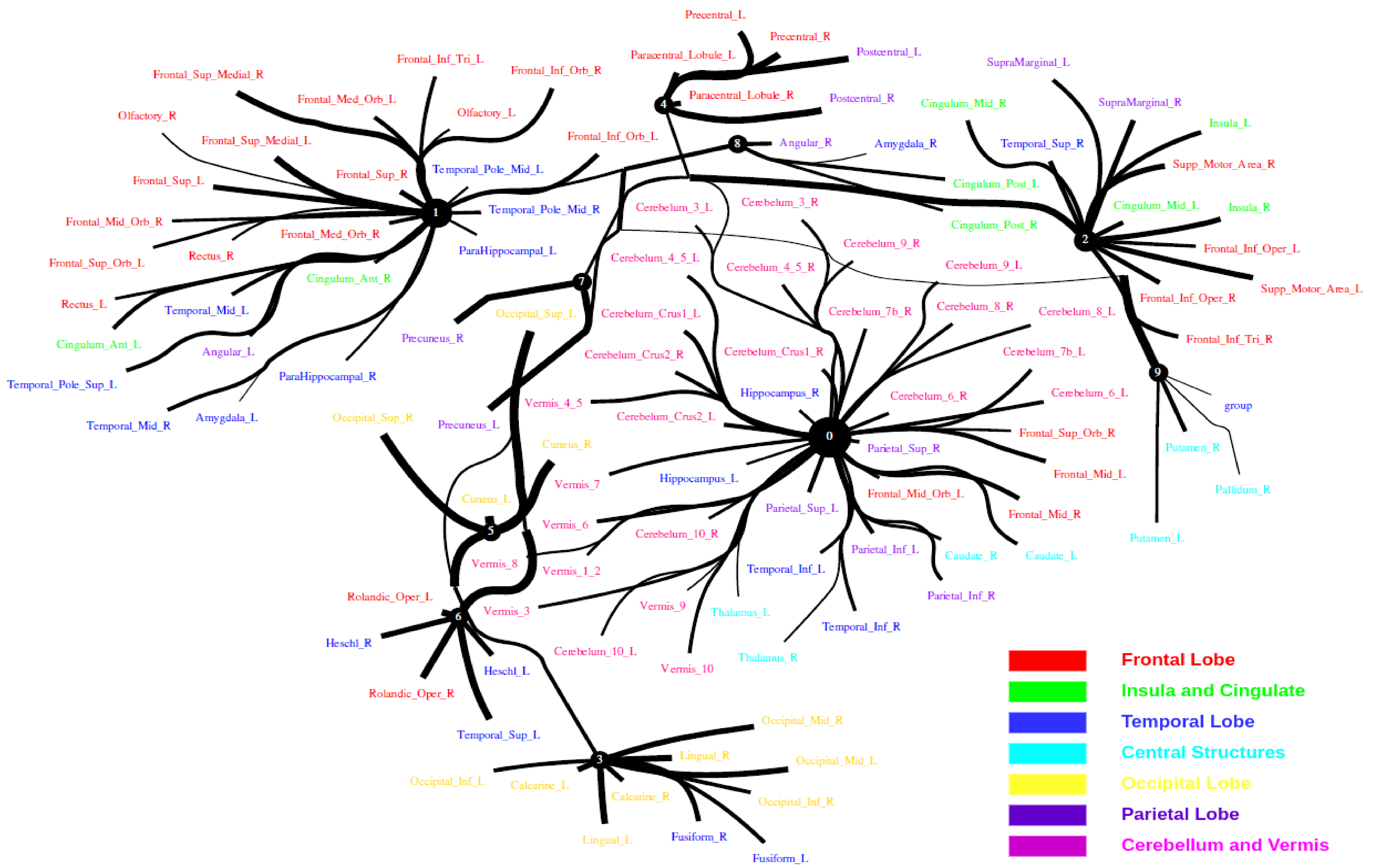
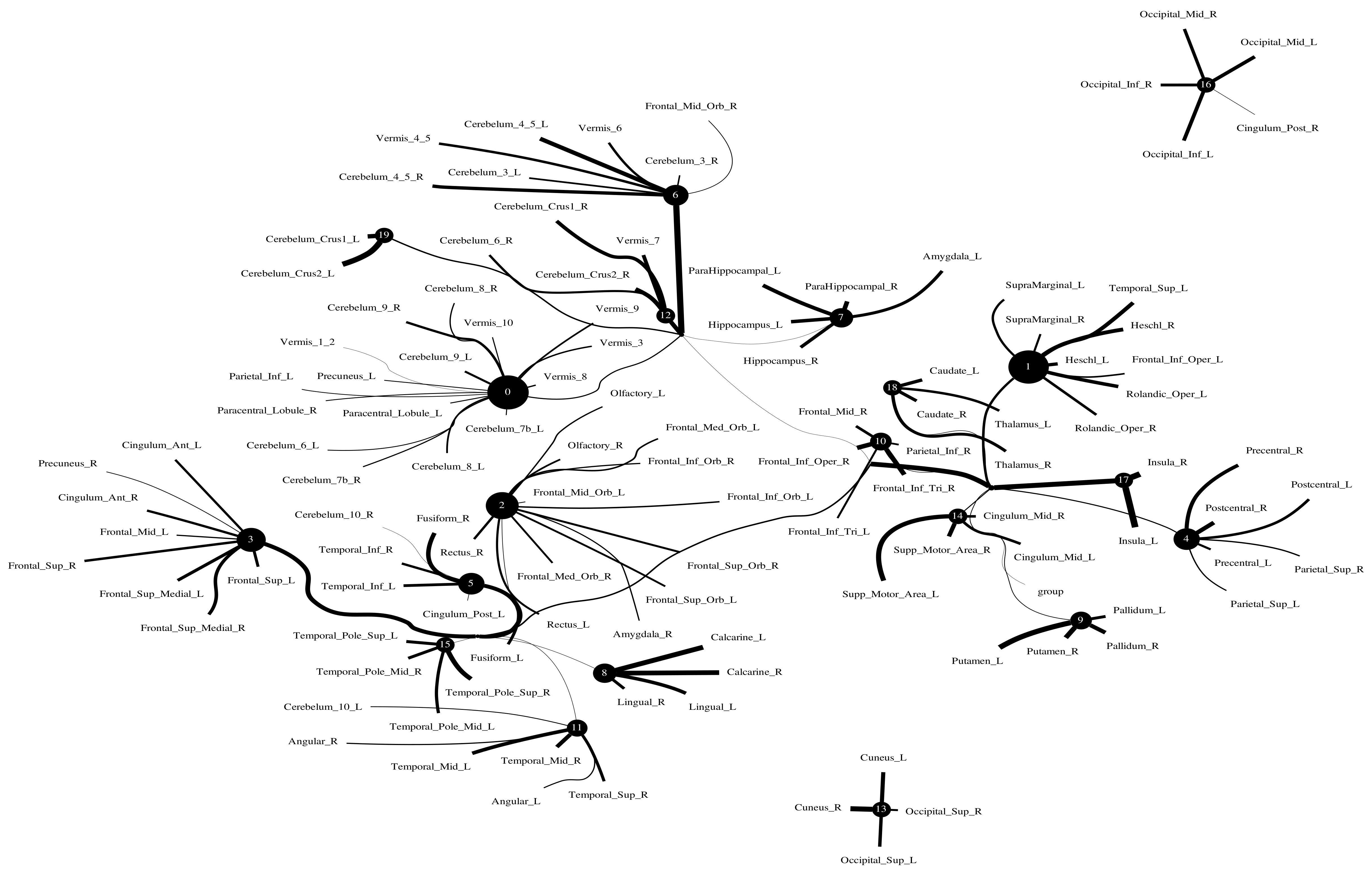
5.2. Large-Scale Connectome with Resting-State fMRI
5.2.1. A Selection of Pre-Defined Atlas
5.2.2. Time Series Signals Extraction
5.2.3. HCP900

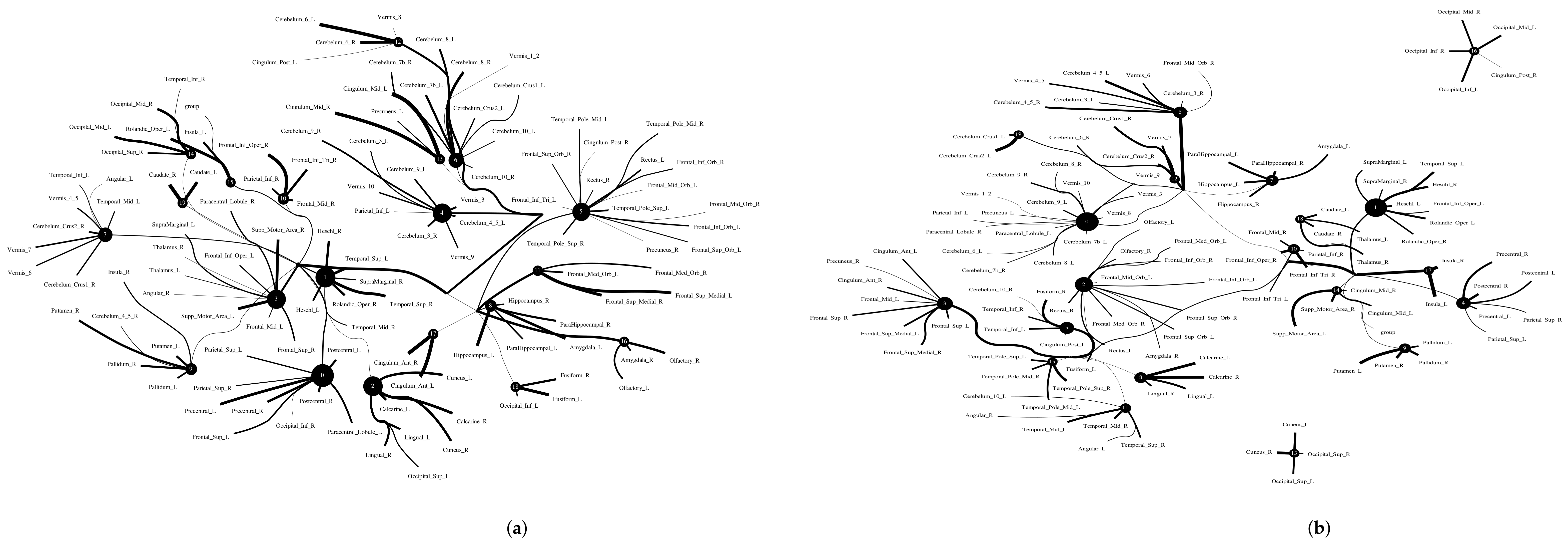
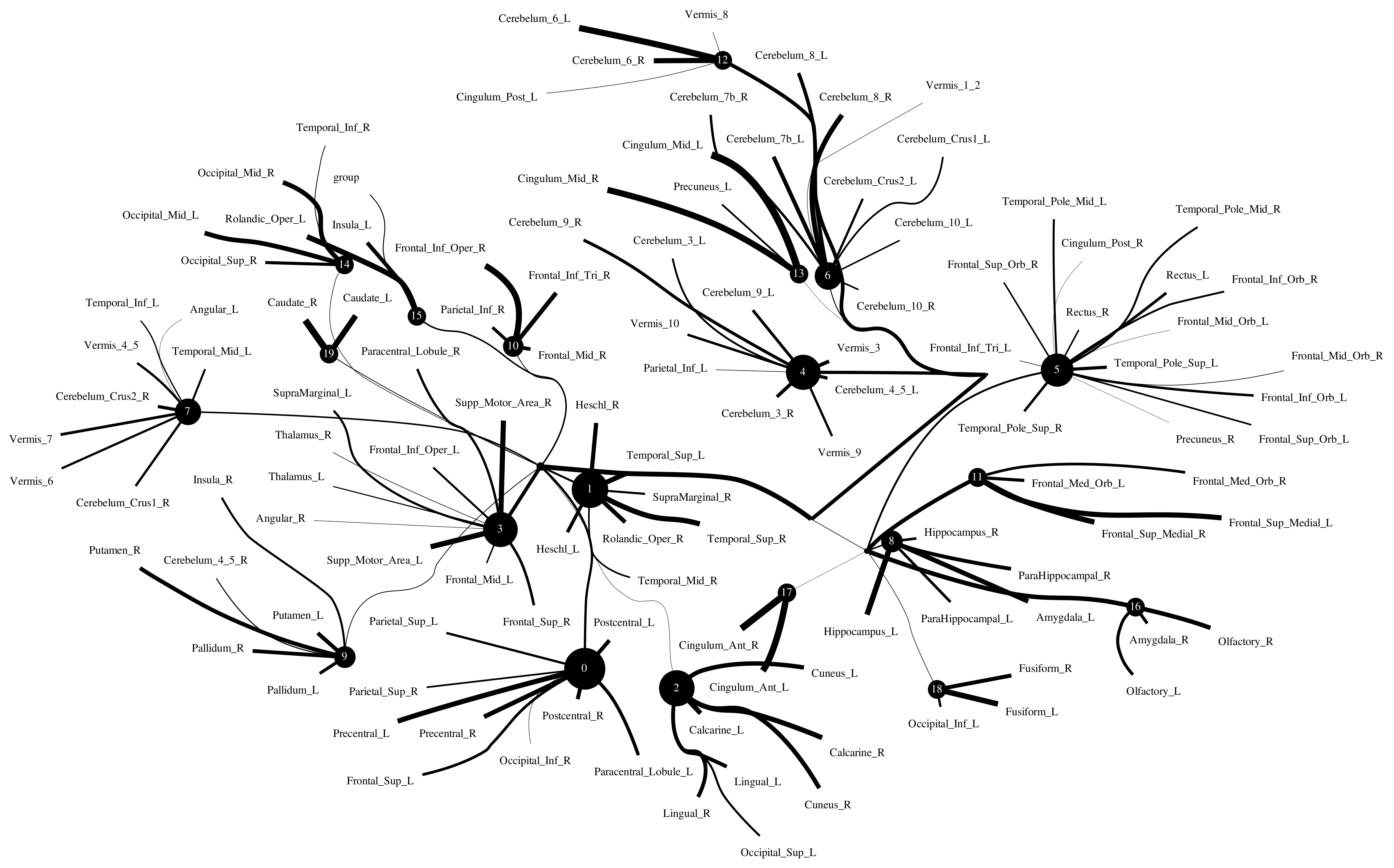
5.2.4. Computational Psychiatry Applications with ACPI
6. Discussion
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| TC | Total Correlation |
| CorEx | Correlation Explanation |
| CC | Linear Correlation |
| I | Mutual Information |
| VAEs | Variational Autoencoders |
| fMRI | functional Magnetic Resonance Imaging |
| BOLD | Blood-Oxygen-Level-Dependent Imaging |
| DCM | Dynamic Causal Modeling |
| GLM | General Linear Model |
| ROI | Region Of Interest |
| HCP | Human Connectome Project |
| MTA | Multimodal Treatment of Attention Deficit Hyperactivity Disorder |
| GNNs | Graph Neural Networks |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brain Area | AAL Regions | AAL Index No. |
|---|---|---|
| Precentral gyrus | 1, 2 | |
| Superior frontal gyrus, dorsolateral | 3, 4 | |
| Superior frontal gyrus, orbital part | 5, 6 | |
| Middle frontal gyrus | 7, 8 | |
| Middle frontal gyrus, orbital part | 9, 10 | |
| Inferior frontal gyrus, opercular part | 11, 12 | |
| Inferior frontal gyrus, triangular part | 13, 14 | |
| Frontal Lobe | Inferior frontal gyrus, orbital part | 15, 16 |
| Rolandic operculum | 17, 18 | |
| Smentary motor area | 19, 20 | |
| Olfactory cortex | 21, 22 | |
| Superior frontal gyrus, medial | 23, 24 | |
| Superior frontal gyrus, medial orbital | 25, 26 | |
| Gyrus rectus | 27, 28 | |
| Paracentral lobule | 69, 70 | |
| Insula | 29, 30 | |
| Insula and | Anterior cingulate and paracingulate gyri | 31, 32 |
| Cingulate | Median cingulate and paracingulate gyri | 33, 34 |
| Posterior cingulate gyrus | 35, 36 | |
| Hippocampus | 37, 38 | |
| Parahippocampal gyrus | 39, 40 | |
| Amygdala | 41, 42 | |
| Fusiform gyrus | 55, 56 | |
| Temporal | Heschl gyrus | 79, 80 |
| Lobe | Superior temporal gyrus | 81, 82 |
| Temporal pole: superior temporal gyrus | 83, 84 | |
| Middle temporal gyrus | 85, 86 | |
| Temporal pole: middle temporal gyrus | 87, 88 | |
| Inferior temporal gyrus | 89, 90 | |
| Caudate nucleus | 71, 72 | |
| Central | Lenticular nucleus, putamen | 73, 74 |
| Structures | Lenticular nucleus, pallidum | 75, 76 |
| Thalamus | 77, 78 | |
| Calcarine fissure and surrounding cortex | 43, 44 | |
| Cuneus | 45, 46 | |
| Occipital | Lingual gyrus | 47, 48 |
| Lobe | Superior occipital gyrus | 49, 50 |
| Middle occipital gyrus | 51, 52 | |
| Inferior occipital gyrus | 53, 54 | |
| Postcentral gyrus | 57, 58 | |
| Superior parietal gyrus | 59, 60 | |
| Parietal | Inferior parietal, but supramarginal and angular gyri | 61, 62 |
| Lobe | Supramarginal gyrus | 63, 64 |
| Angular gyrus | 65, 66 | |
| Precuneus | 67, 68 | |
| Cerebellum Crus 1 | 91, 92 | |
| Cerebellum Crus 2 | 93, 94 | |
| Cerebellum 3 | 95, 96 | |
| Cerebellum 4, 5 | 97, 98 | |
| Cerebellum 6 | 99, 100 | |
| Cerebellum 7b | 101, 102 | |
| Cerebellum 8 | 103, 104 | |
| Cerebellum 9 | 105, 106 | |
| Cerebellum and Vermis | Cerebellum 10 | 107, 108 |
| Vermis 1, 2 | 109 | |
| Vermis 3 | 110 | |
| Vermis 4, 5 | 111 | |
| Vermis 6 | 112 | |
| Vermis 7 | 113 | |
| Vermis 8 | 114 | |
| Vermis 9 | 115 | |
| Vermis 10 | 116 |
References
- Friston, K. Functional and effective connectivity: A review. Brain Connect. 2011, 1, 13–36. [Google Scholar] [CrossRef] [PubMed]
- Porta, A.; Faes, L.; Bari, V.; Marchi, A.; Bassani, T.; Nollo, G.; Perseguini, N.M.; Milan, J.; Minatel, V.; Borghi-Silva, A.; et al. Effect of age on complexity and causality of the cardiovascular control: Comparison between model-based and model-free approaches. PLoS ONE 2014, 9, e89463. [Google Scholar] [CrossRef]
- Heuvel, M.; Pol, H. Exploring the brain network: A review on resting-state fmri functional connectivity. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 2010, 20, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Sporns, O.; Tononi, G.; Kötter, R. The human connectome: A structural description of the human brain. PLoS Comput. Biol. 2005, 1, e42. [Google Scholar] [CrossRef] [PubMed]
- Bastos, A.; Schoffelen, J.-M. A tutorial review of functional connectivity analysis methods and their interpretational pitfalls. Front. Syst. Neurosci. 2016, 9, 1. [Google Scholar] [CrossRef]
- Lizier, J.T.; Heinzle, J.; Horstmann, A.; Haynes, J.; Prokopenko, M. Multivariate information-theoretic measures reveal directed information structure and task relevant changes in fMRI connectivity. J. Comput. Neurosci. 2011, 30, 85–107. [Google Scholar] [CrossRef]
- Piasini, E.; Panzeri, S. Information theory in neuroscience. Entropy 2019, 21, 62. [Google Scholar] [CrossRef]
- Ince, R.; Giordano, B.; Kayser, C.; Rousselet, G.; Gross, J.; Schyns, P. A statistical framework for neuroimaging data analysis based on mutual information estimated via a gaussian copula. Hum. Brain Mapp. 2016, 38, 11. [Google Scholar] [CrossRef]
- Dimitrov, A.; Lazar, A.; Victor, J. Information theory in neuroscience. J. Comput. Neurosci. 2011, 30, 1–5. [Google Scholar] [CrossRef]
- Borst, A.; Theunissen, F. Information theory and neural coding. Nat. Neurosci. 1999, 2, 947–957. [Google Scholar] [CrossRef]
- Tkacik, G.; Marre, O.; Mora, T.; Amodei, D.; Berry, M., II; Bialek, W. The simplest maximum entropy model for collective behavior in a neural network. J. Stat. Mech. Theory Exp. 2012, 2013, 7. [Google Scholar]
- Gomez-Villa, A.; Bertalmio, M.; Malo, J. Visual information flow in Wilson-Cowan networks. J. Neurophysiol. 2020, 123, 2249–2268. [Google Scholar] [CrossRef] [PubMed]
- Malo, J. Spatio-chromatic information available from different neural layers via gaussianization. J. Math. Neurosci. 2020, 10, 18. [Google Scholar] [CrossRef] [PubMed]
- Malo, J. Information flow in biological networks for color vision. Entropy 2022, 24, 1442. [Google Scholar] [CrossRef]
- Farahani, F.; Karwowski, W.; Lighthall, N. Application of graph theory for identifying connectivity patterns in human brain networks: A systematic review. Front. Neurosci. 2019, 13, 585. [Google Scholar] [CrossRef]
- Sporns, O. Graph theory methods: Applications in brain networks. Dialogues Clin. Neurosci. 2018, 20, 111–121. [Google Scholar] [CrossRef]
- Rosas, F.; Mediano, P.A.M.; Ugarte, M.; Jensen, H.J. An information-theoretic approach to self-organisation: Emergence of complex interdependencies in coupled dynamical systems. Entropy 2018, 20, 793. [Google Scholar] [CrossRef]
- Rosas, F.E.; Mediano, P.A.M.; Gastpar, M.; Jensen, H.J. Quantifying high-order interdependencies via multivariate extensions of the mutual information. Phys. Rev. E 2019, 100, 32305. [Google Scholar] [CrossRef]
- Tononi, G.; Edelman, G. Consciousness and complexity. Science 1999, 282, 1846–1851. [Google Scholar] [CrossRef]
- Pereda, E.; Quian, R.; Bhattacharya, J. Nonlinear multivariate analysis of neurophysiological signals. Prog. Neurobiol. 2005, 77, 1–37. [Google Scholar] [CrossRef]
- Chai, B.; Walther, D.B.; Beck, D.M.; Fei-Fei, L. Exploring functional connectivity of the human brain using multivariate information analysis. In Proceedings of the 22nd International Conference on Neural Information Processing Systems, Vancouver, BC, Canada, 7–10 December 2009; Curran Associates Inc.: Red Hook, NY, USA, 2009; pp. 270–278. [Google Scholar]
- Wang, Z.; Alahmadi, A.; Zhu, D.; Li, T. Brain functional connectivity analysis using mutual information. In Proceedings of the 2015 IEEE Global Conference on Signal and Information Processing (GlobalSIP), Orlando, FL, USA, 14–16 December 2015; pp. 542–546. [Google Scholar]
- Jomaa, M.E.S.H.; Colominas, M.; Jrad, N.; Bogaert, P.V.; Humeau-Heurtier, A. A new mutual information measure to estimate functional connectivity: Preliminary study. In Proceedings of the Conference proceedings: Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Berlin, Germany, 23–27 July 2019; pp. 640–643. [Google Scholar]
- Li, Q. Functional connectivity inference from fmri data using multivariate information measures. Neural Netw. 2022, 146, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Steeg, G.V.; Malo, J. Functional connectivity in visual areas from Total Correlation. arXiv 2022. Available online: https://arxiv.org/abs/2208.05770 (accessed on 11 August 2022).
- Watanabe, S. Information theoretical analysis of multivariate correlation. IBM J. Res. Dev. 1960, 4, 66–82. [Google Scholar] [CrossRef]
- Studeny, M.; Vejnarova, J. The multi-information function as a tool for measuring stochastic dependence. In Learning in Graphical Models; Springer: Dordrecht, The Netherlands; pp. 261–298.
- Laparra, V.; Camps-Valls, G.; Malo, J. Iterative gaussianization: From ICA to random rotations. IEEE Trans. Neural Netw. 2011, 22, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Laparra, V.; Johnson, E.; Camps, G.; Santos, R.; Malo, J. Information theory measures via multidimensional gaussianization. arXiv Stats. Mach. Learn. 2022. Available online: https://arxiv.org/abs/2010.03807 (accessed on 25 November 2020).
- Essen, D.V.; Smith, S.; Barch, D.; Behrens, T.; Yacoub, E.; Ugurbil, K. The wu-minn human connectome project: An overview. NeuroImage 2013, 80, 62–79. [Google Scholar] [CrossRef]
- Essen, D.C.; Ugurbil, K.; Auerbach, E.; Barch, D.; Behrens, T.E.J.; Bucholz, R.; Chang, A.; Chen, L.; Corbetta, M.; Curtiss, S.; et al. The human connectome project: A data acquisition perspective. NeuroImage 2012, 62, 2222–2231. [Google Scholar] [CrossRef]
- Steeg, G.V.; Galstyan, A. Discovering structure in high-dimensional data through correlation explanation. Adv. Neural Inf. Process. Syst. 2014, 577. [Google Scholar]
- Steeg, G.V.; Galstyan, A. Maximally informative hierarchical representations of high-dimensional data. In AISTATS’15; PMLR: San Diego, CA, USA, 2015. [Google Scholar]
- Cover, T.M.; Thomas, J.A. Elements of Information Theory (Wiley Series in Telecommunications and Signal Processing); Wiley-Interscience: Hoboken, NJ, USA, 2006. [Google Scholar]
- Kraskov, A.; Stögbauer, H.; Grassberger, P. Estimating mutual information. Phys. Rev. E 2004, 69, 66138. [Google Scholar] [CrossRef] [PubMed]
- Fabila-Carrasco, J.S.; Tan, C.; Escudero, J. Permutation entropy for graph signals. IEEE Trans. Signal Inf. Process. Over Netw. 2022, 8, 288–300. [Google Scholar] [CrossRef]
- Lyu, S.; Simoncelli, E.P. Nonlinear Extraction of Independent Components of Natural Images Using Radial Gaussianization. Neural Comput. 2009, 21, 1485–1519. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Brekelmans, R.; Steeg, G.V.; Galstyan, A. Auto-encoding correlation explanation. In Proceedings of the 22nd International Conference on AI and Statistics (AISTATS), Naha, Japan, 16–18 April 2019. [Google Scholar]
- Yu, S.; Giraldo, L.G.S.; Jenssen, R.; Principe, J.C. Multivariate extension of matrix-based rényi’s α-order entropy functional. IEEE Trans. Pattern Anal. Mach. Intell. 2019, 42, 2960–2966. [Google Scholar] [CrossRef] [PubMed]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Steeg, G.V. Unsupervised learning via Total Correlation explanation. In IJCAI; Artificial Intelligence Organization: Melbourne, Australia, 2017. [Google Scholar]
- Steeg, G.V.; Harutyunyan, H.; Moyer, D.; Galstyan, A. Fast structure learning with modular regularization. Adv. Neural Inf. Process. Syst. 2019, 32, 15593–15603. [Google Scholar]
- Manning, C.D.; Raghavan, P.; Schütze, H. Introduction to Information Retrieval; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Tzourio-Mazoyer, N.; Landeau, B.; Crivello, P.D.F.F.; Etard, O.N.D.; Delcroix, N.; Mazoyer, B.; Marc, J. Automated anatomical labeling of activations in spm using a macroscopic anatomical parcellation of the mni mri single-subject brain. NeuroImage 2002, 15, 273–289. [Google Scholar] [CrossRef] [PubMed]
- Behan, B.; Connolly, G.; Datwani, S.; Doucet, M.; Ivanovic, J.; Morioka, R.; Stone, A.; Watts, R.; Smyth, B.; Garavan, H. Response inhibition and elevated parietal-cerebellar correlations in chronic adolescent cannabis users. Neuropharmacology 2013, 84, 6. [Google Scholar] [CrossRef]
- Bubl, E.; van Elst, L.T.; Gondan, M.; Ebert, D.; Greenlee, M. Vision in depressive disorder. World J. Biol. Psychiatry Off. J. World Fed. Soc. Biol. Psychiatry 2007, 10, 377–384. [Google Scholar] [CrossRef]
- Zhang, R.; Volkow, N. Brain default-mode network dysfunction in addiction. NeuroImage 2019, 200, 313–331. [Google Scholar] [CrossRef]
- Giedd, J.; Keshavan, M.; Paus, T. Why do many psychiatric disorders emerge during adolescence? Nat. Rev. Neurosci. 2008, 9, 947–957. [Google Scholar]
- Medina, K.; Hanson, K.; Dager, A.; Cohen-Zion, M.; Nagel, B.; Tapert, S. Neuropsychological functioning in adolescent marijuana users: Subtle deficits detectable after a month of abstinence. J. Int. Neuropsychol. Soc. JINS 2007, 13, 807–820. [Google Scholar] [CrossRef]
- Poline, J.-B.; Brett, M. The general linear model and fmri: Does love last forever? NeuroImage 2012, 62, 871–880. [Google Scholar] [CrossRef] [PubMed]
- Dowdle, L.T.; Ghose, G.; Chen, C.C.C.; Ugurbil, K.; Yacoub, E.; Vizioli, L. Statistical power or more precise insights into neuro-temporal dynamics? assessing the benefits of rapid temporal sampling in fmri. Prog. Neurobiol. 2021, 207, 102171. [Google Scholar] [CrossRef] [PubMed]
- Marreiros, A.; Stephan, K.; Friston, K. Dynamic causal modeling. Scholarpedia 2010, 5, 9568. [Google Scholar] [CrossRef]
- Porta, A.; Faes, L. Wiener–granger causality in network physiology with applications to cardiovascular control and neuroscience. Proc. IEEE 2016, 104, 282–309. [Google Scholar] [CrossRef]
- Welling, M.; Kipf, T.N. Semi-supervised classification with graph convolutional networks. In Proceedings of the (ICLR 2017), Toulon, France, 24–26 April 2017. [Google Scholar]
- Tantardini, M.; Ieva, F.; Tajoli, L.; Piccardi, C. Comparing methods for comparing networks. Sci. Rep. 2019, 9, 17557. [Google Scholar] [CrossRef]
- Cui, H.; Dai, W.; Zhu, Y.; Li, X.; He, L.; Yang, C. Brainnnexplainer: An interpretable graph neural network framework for brain network based disease analysis. arXiv 2021, arXiv:2107.05097. [Google Scholar]
- Zheng, K.; Yu, S.; Li, B.; Jenssen, R.; Chen, B. Brainib: Interpretable brain network-based psychiatric diagnosis with graph information bottleneck. arXiv 2022, arXiv:2205.03612. [Google Scholar]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Steeg, G.V.; Yu, S.; Malo, J. Functional Connectome of the Human Brain with Total Correlation. Entropy 2022, 24, 1725. https://doi.org/10.3390/e24121725
Li Q, Steeg GV, Yu S, Malo J. Functional Connectome of the Human Brain with Total Correlation. Entropy. 2022; 24(12):1725. https://doi.org/10.3390/e24121725
Chicago/Turabian StyleLi, Qiang, Greg Ver Steeg, Shujian Yu, and Jesus Malo. 2022. "Functional Connectome of the Human Brain with Total Correlation" Entropy 24, no. 12: 1725. https://doi.org/10.3390/e24121725
APA StyleLi, Q., Steeg, G. V., Yu, S., & Malo, J. (2022). Functional Connectome of the Human Brain with Total Correlation. Entropy, 24(12), 1725. https://doi.org/10.3390/e24121725