
Goal Directedness, Chemical Organizations, and Cybernetic Mechanisms




Abstract
:1. Introduction
2. Goal Directedness
3. Modeling Goal-Oriented Systems Using Reaction Networks
3.1. Closed and Semi-Self-Maintaining Subnetworks
3.2. Goal-Oriented Systems Are Organizations
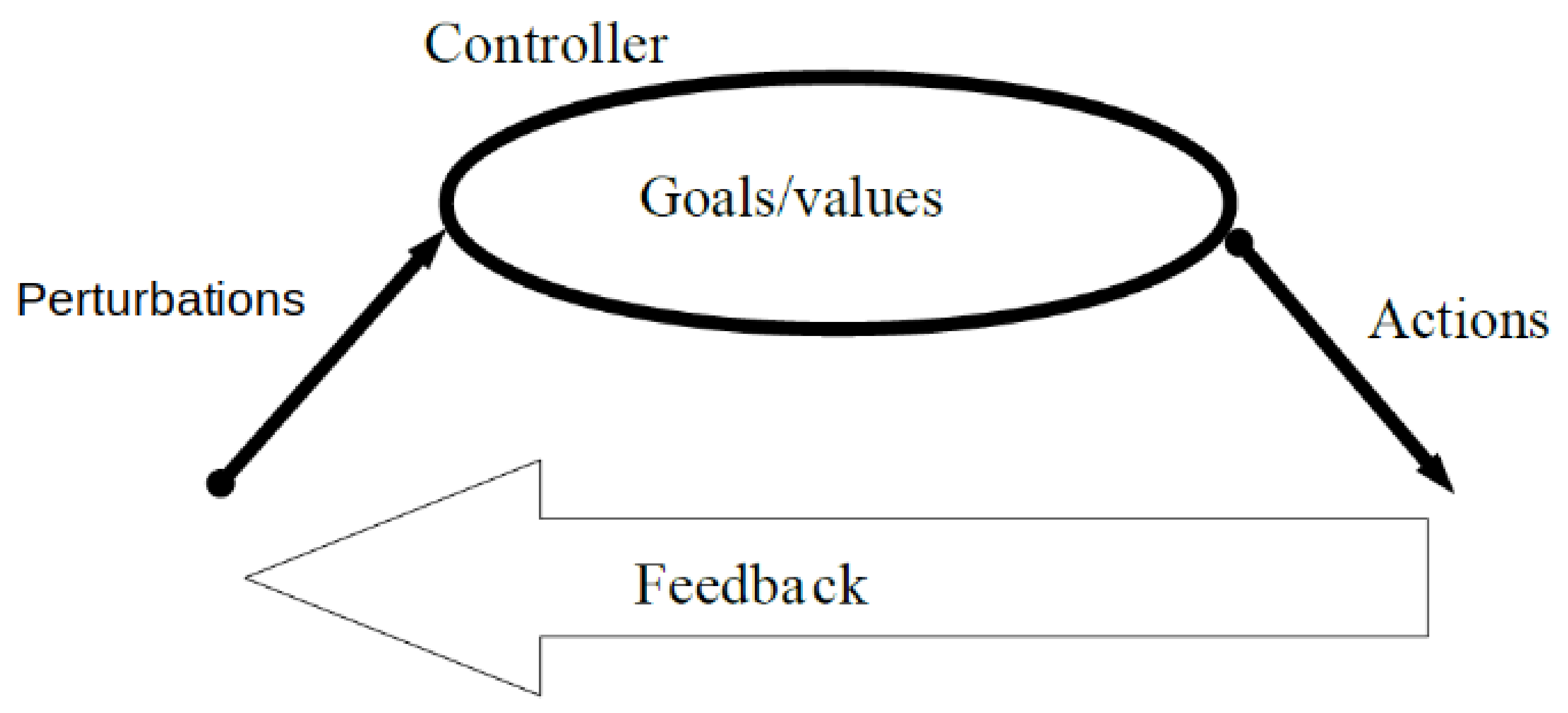
3.3. Three Types of Change for Perturbations, Actions, and Goals
4. Goal Directedness Examples
4.1. Direct Negative Feedback of a Species on Itself
4.2. Cycle of Mutual Production
4.3. One Cycle with Stable Ratio Influencing Another Part, Giving Stable State
4.4. Catalyst Influencing the Self-Maintenance and Over-Producibility
4.5. An Example Incorporating Structural Perturbations
5. Discussion
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Deacon, T.; Sherman, J. The physical origins of purposive systems. Adv. Conscious. Res. 2007, 71, 3. [Google Scholar]
- Rosenblueth, A.; Wiener, N.; Bigelow, J. Behavior, purpose and teleology. Philos. Sci. 1943, 10, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Heylighen, F.; Joslyn, C. Cybernetics and Second-Order Cybernetics. In Encyclopedia of Physical Science and Technology; Elsevier: Amsterdam, The Netherlands, 2003; pp. 155–169. [Google Scholar] [CrossRef] [Green Version]
- Ashby, W. Design for a Brain: The Origin of Adaptive Behaviour; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Luhmann, N. The Differentiation of Society; Columbia University Press: New York, NY, USA, 1982. [Google Scholar]
- Heylighen, F.; Rosseel, E.; Demeyere, F. Self-Steering and Cognition in Complex Systems: Toward a NEW Cybernetics; CRC Press: Boca Raton, FL, USA, 1990; Volume 22. [Google Scholar]
- Razeto-Barry, P. Autopoiesis 40 years later. A review and a reformulation. Orig. Life Evol. Biosph. 2012, 42, 543–567. [Google Scholar] [CrossRef]
- Maturana, H.R.; Varela, F.J. Autopoiesis and Cognition: The Realization of the Living; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1991; Volume 42. [Google Scholar]
- Hordijk, W.; Hein, J.; Steel, M. Autocatalytic sets and the origin of life. Entropy 2010, 12, 1733–1742. [Google Scholar] [CrossRef]
- Kauffman, S. At Home in the Universe: The Search for the Laws of Self-Organization and Complexity; Oxford University Press: Oxford, UK, 1996. [Google Scholar]
- Dittrich, P.; Di Fenizio, P.S. Chemical organisation theory. Bull. Math. Biol. 2007, 69, 1199–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veloz, T.; Razeto-Barry, P. Reaction networks as a language for systemic modeling: Fundamentals and examples. Systems 2017, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Hordijk, W.; Steel, M.; Dittrich, P. Autocatalytic sets and chemical organizations: Modeling self-sustaining reaction networks at the origin of life. New J. Phys. 2018, 20, 015011. [Google Scholar] [CrossRef] [Green Version]
- Rubin, S.; Veloz, T.; Maldonado, P. Beyond planetary-scale feedback self-regulation: Gaia as an autopoietic system. Biosystems 2021, 199, 104314. [Google Scholar] [CrossRef]
- Kreyssig, P.; Wozar, C.; Peter, S.; Veloz, T.; Ibrahim, B.; Dittrich, P. Effects of small particle numbers on long-term behaviour in discrete biochemical systems. Bioinformatics 2014, 30, i475–i481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beer, S. What is cybernetics? Kybernetes 2002, 31, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Trestman, M. Goal-Directedness, Behavior and Evolution: A Philosophical Investigation; University of California: Davis, CA, USA, 2010. [Google Scholar]
- Kruglanski, A.W.; Chernikova, M.; Babush, M.; Dugas, M.; Schumpe, B.M. The architecture of goal systems: Multifinality, equifinality, and counterfinality in means—end relations. In Advances in Motivation Science; Elsevier: Amsterdam, The Netherlands, 2015; Volume 2, pp. 69–98. [Google Scholar]
- Biehl, M.; Polani, D. Apparent actions and apparent goal-directedness. In Artificial Life Conference Proceedings 13; MIT Press: Cambridge, MA, USA, 2015; p. 511. [Google Scholar]
- Heylighen, F. Complexity and Self-Organization. In Encyclopedia of Library and Information Science, 4th ed.; CRC Press: Boca Raton, FL, USA, 2008; pp. 1034–1043. [Google Scholar]
- Ramstead, M.J.D.; Badcock, P.B.; Friston, K.J. Answering Schrödinger’s question: A free-energy formulation. Phys. Life Rev. 2018, 24, 1–16. [Google Scholar] [CrossRef]
- Heylighen, F. The science of self-organization and adaptivity. Encycl. Life Support Syst. 2001, 5, 253–280. [Google Scholar]
- Shinar, G.; Feinberg, M. Structural sources of robustness in biochemical reaction networks. Science 2010, 327, 1389–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falkowski, P.G.; Fenchel, T.; Delong, E.F. The microbial engines that drive Earth’s biogeochemical cycles. Science 2008, 320, 1034–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashby, W.R. Principles of the Self-Organizing System. In Facets of Systems Science; Springer US: Boston, MA, USA, 1991; pp. 521–536. [Google Scholar] [CrossRef]
- Dittrich, P.; Winter, L. Reaction networks as a formal mechanism to explain social phenomena. In Proceedings of the Fourth International Workshop on Agent-Based Approaches in Economics and Social Complex Systems (AESCS 2005), Tokyo, Japan, 9–13 July 2005; pp. 9–13. [Google Scholar]
- Strogatz, S.H. Nonlinear Dynamics and Chaos: With Applications to Physics, Biology, Chemistry, and Engineering; Westview Press: Boulder, CO, USA, 2014. [Google Scholar]
- Peter, S.; Dittrich, P. On the relation between organizations and limit sets in chemical reaction systems. Adv. Complex Syst. 2011, 14, 77–96. [Google Scholar] [CrossRef]
- Peter, S.; Veloz, T.; Dittrich, P. Feasibility of Organizations-A Refinement of Chemical Organization Theory with Application to P Systems. In Proceedings of the Eleventh International Conference on Membrane Computing (CMC11), Jena, Germany, 24–27 August 2010; p. 369. [Google Scholar]
- Centler, F.; Kaleta, C.; di Fenizio, P.S.; Dittrich, P. Computing chemical organizations in biological networks. Bioinformatics 2008, 24, 1611–1618. [Google Scholar] [CrossRef] [Green Version]
- Centler, F.; Kaleta, C.; Speroni di Fenizio, P.; Dittrich, P. A parallel algorithm to compute chemical organizations in biological networks. Bioinformatics 2010, 26, 1788–1789. [Google Scholar] [CrossRef] [Green Version]
- Veloz, T.; Razeto-Barry, P. Reaction networks as a language for systemic modeling: On the study of structural changes. Systems 2017, 26, 30. [Google Scholar] [CrossRef] [Green Version]
- Speroni di Fenizio, P. The lattice of chemical organisations. In Artificial Life Conference Proceedings 13; MIT Press: Cambridge, MA, USA, 2015; pp. 242–248. [Google Scholar]
- Veloz, T.; Bassi, A.; Maldonado, P.; Razeto, P. On the existence of synergies and the separability of closed reaction networks. In International Symposium on Molecular Logic and Computational Synthetic Biology; Springer: Berlin/Heidelberg, Germany, 2018; pp. 105–120. [Google Scholar]
- Feinberg, M.; Horn, F.J. Dynamics of open chemical systems and the algebraic structure of the underlying reaction network. Chem. Eng. Sci. 1974, 29, 775–787. [Google Scholar] [CrossRef]
- Carbonaro, N.J.; Thorpe, I.F. Using Structural Kinetic Modeling To Identify Key Determinants of Stability in Reaction Networks. J. Phys. Chem. A 2017, 121, 4982–4992. [Google Scholar] [CrossRef]
- Orth, J.D.; Thiele, I.; Palsson, B.Ø. What is flux balance analysis? Nat. Biotechnol. 2010, 28, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Joshi, B.; Shiu, A. A survey of methods for deciding whether a reaction network is multistationary. Math. Model. Nat. Phenom. 2015, 10, 47–67. [Google Scholar] [CrossRef] [Green Version]
- Deng, J.; Jones, C.; Feinberg, M.; Nachman, A. On the steady states of weakly reversible chemical reaction networks. arXiv 2011, arXiv:1111.2386. [Google Scholar]
- Feinberg, M. Lectures on chemical reaction networks. In Notes of Lectures Given at the Mathematics Research Center; University of Wisconsin: Madison, WI, USA, 1979. [Google Scholar]
- Angeli, D. On modularity and persistence of chemical reaction networks. In Proceedings of the 2008 47th IEEE Conference on Decision and Control, Cancun, Mexico, 9–11 December 2008; pp. 2650–2655. [Google Scholar] [CrossRef]
- Prescott, T.P.; Papachristodoulou, A. Layered decomposition for the model order reduction of timescale separated biochemical reaction networks. J. Theor. Biol. 2014, 356, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Ravasz, E.; Somera, A.L.; Mongru, D.A.; Oltvai, Z.N.; Barabási, A.L. Hierarchical organization of modularity in metabolic networks. Science 2002, 297, 1551–1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Change | Perturbation (in Phase Space) | Action | Goal |
|---|---|---|---|
| State | another point | causal effect | end-state(s) |
| Process | possible directions | dynamic control | end-behavior |
| Structural | dimension | rule control | end-structure |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Busseniers, E.; Veloz, T.; Heylighen, F. Goal Directedness, Chemical Organizations, and Cybernetic Mechanisms. Entropy 2021, 23, 1039. https://doi.org/10.3390/e23081039
Busseniers E, Veloz T, Heylighen F. Goal Directedness, Chemical Organizations, and Cybernetic Mechanisms. Entropy. 2021; 23(8):1039. https://doi.org/10.3390/e23081039
Chicago/Turabian StyleBusseniers, Evo, Tomas Veloz, and Francis Heylighen. 2021. "Goal Directedness, Chemical Organizations, and Cybernetic Mechanisms" Entropy 23, no. 8: 1039. https://doi.org/10.3390/e23081039
APA StyleBusseniers, E., Veloz, T., & Heylighen, F. (2021). Goal Directedness, Chemical Organizations, and Cybernetic Mechanisms. Entropy, 23(8), 1039. https://doi.org/10.3390/e23081039