Mutual Information and Information Gating in Synfire Chains

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
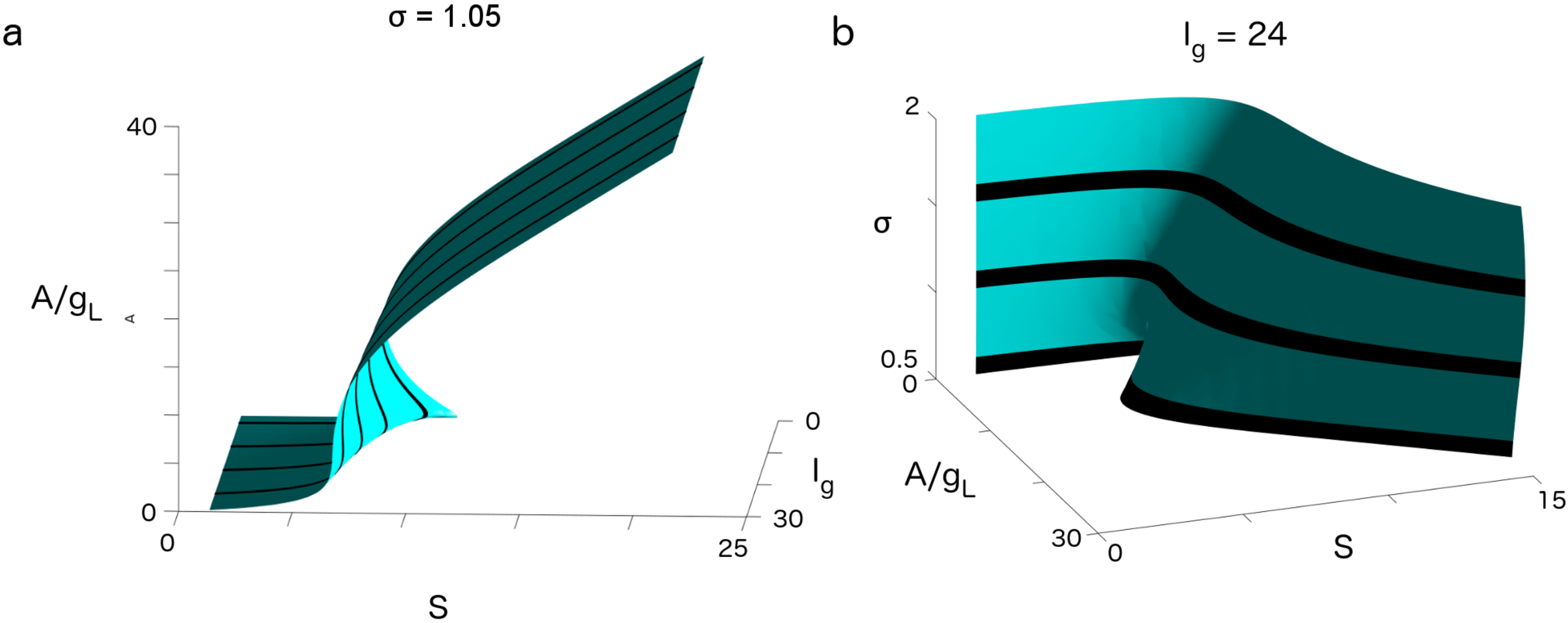
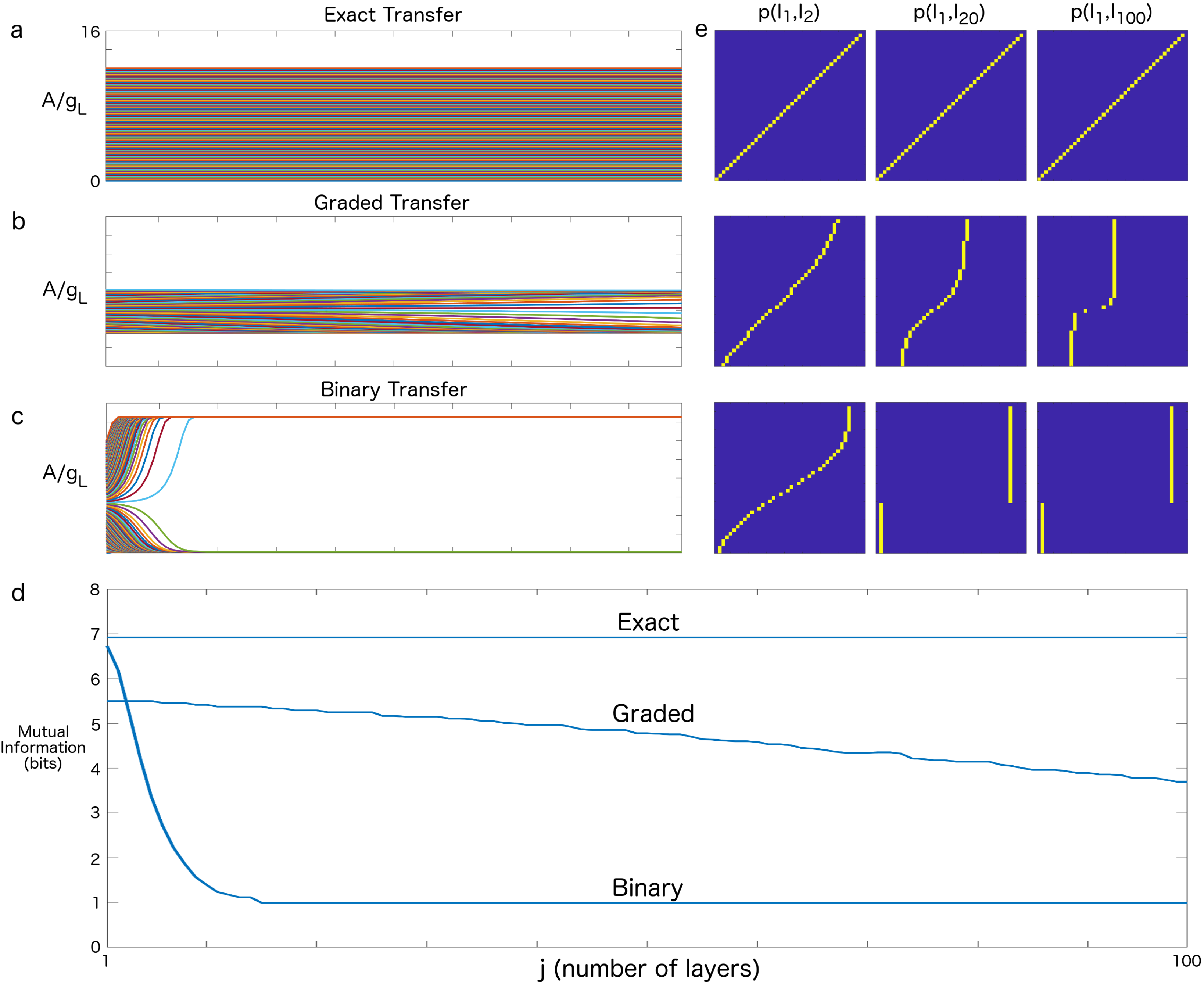
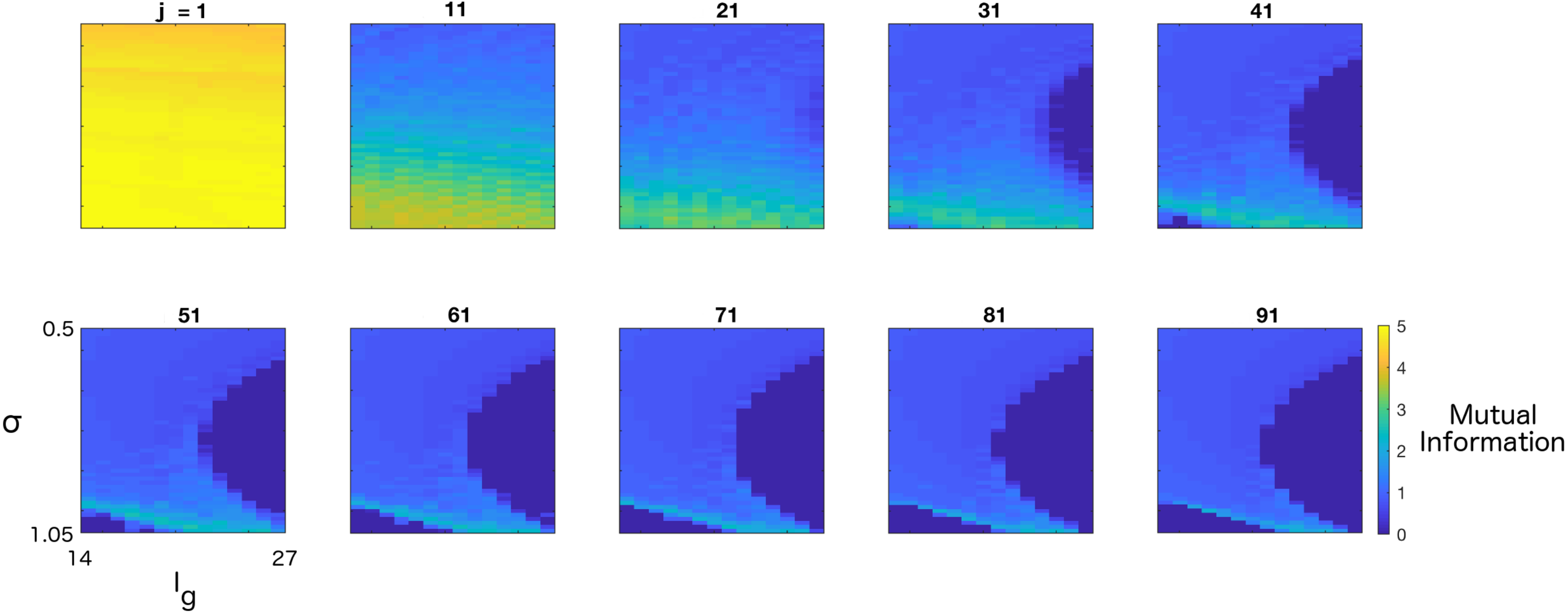
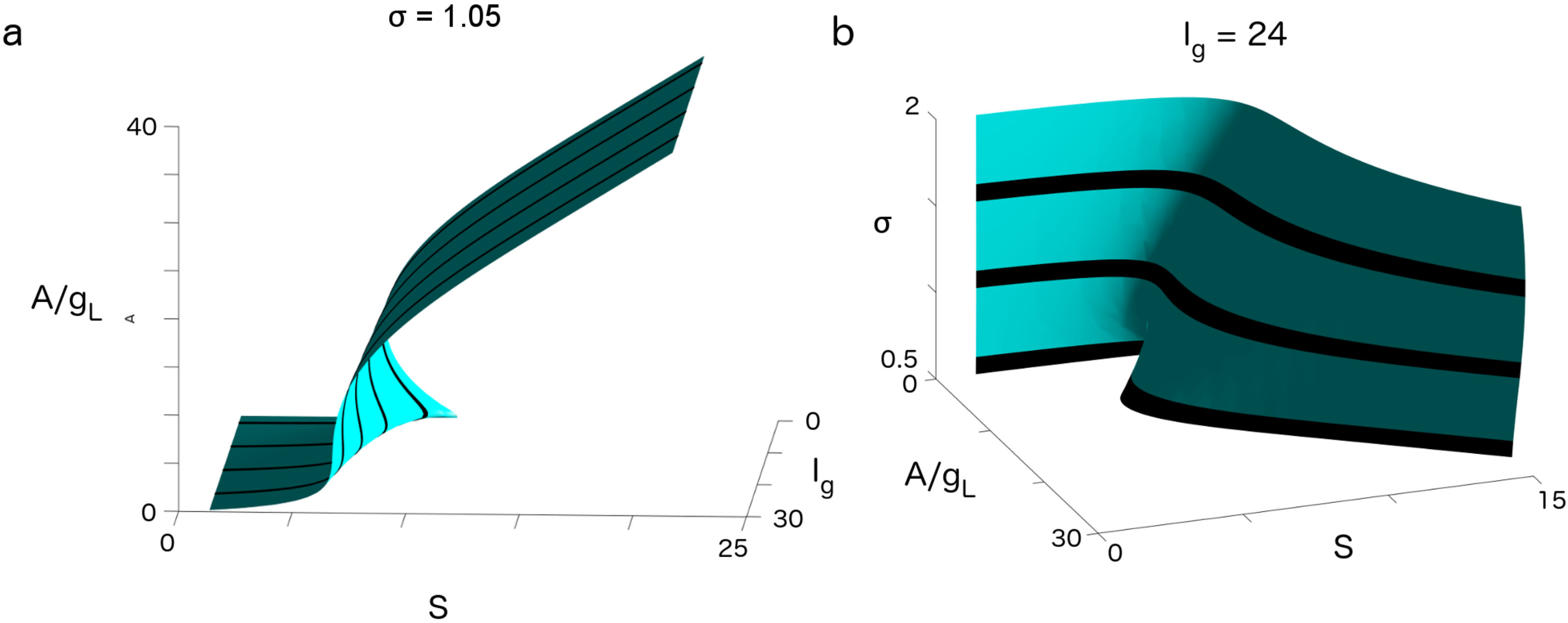
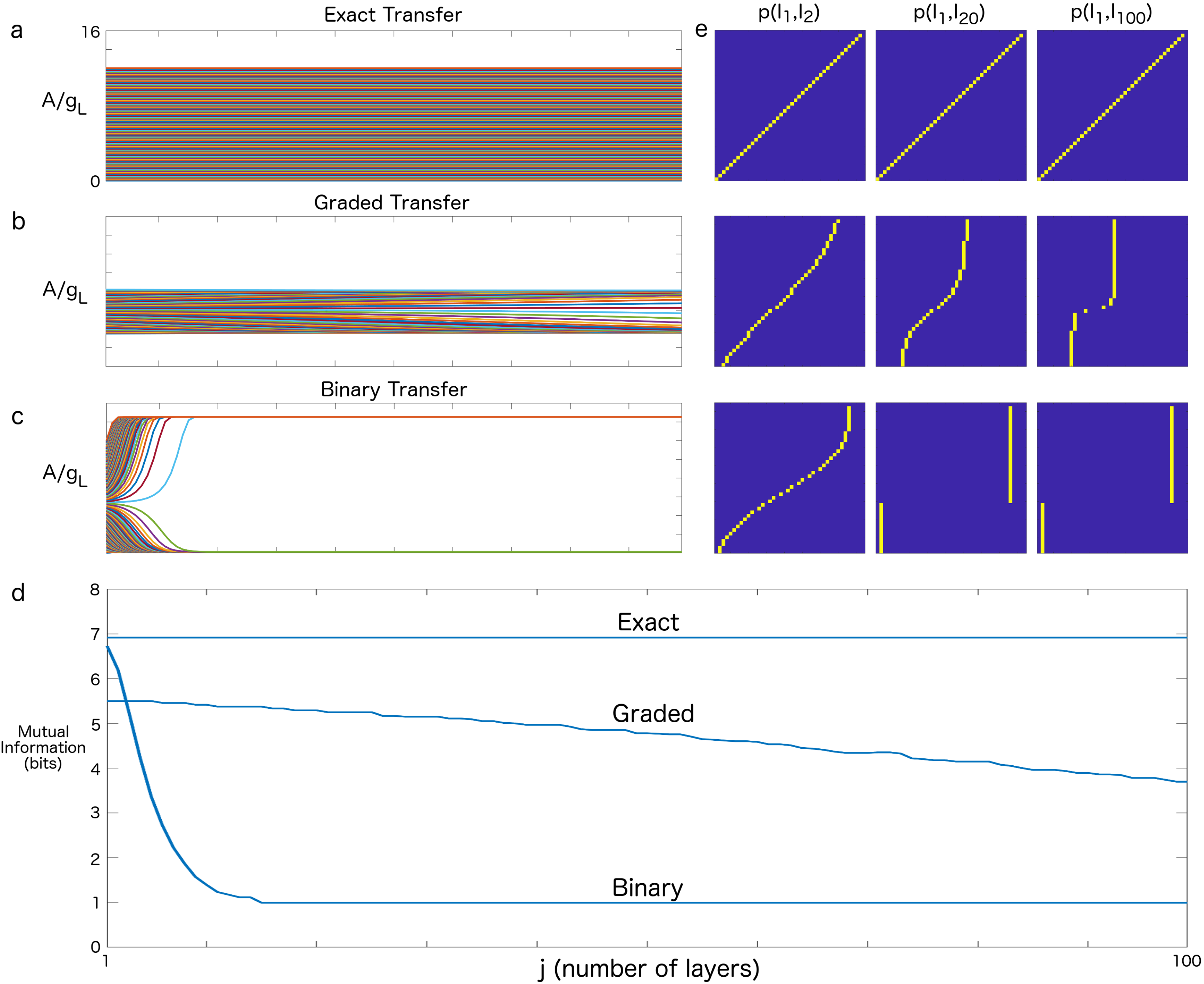
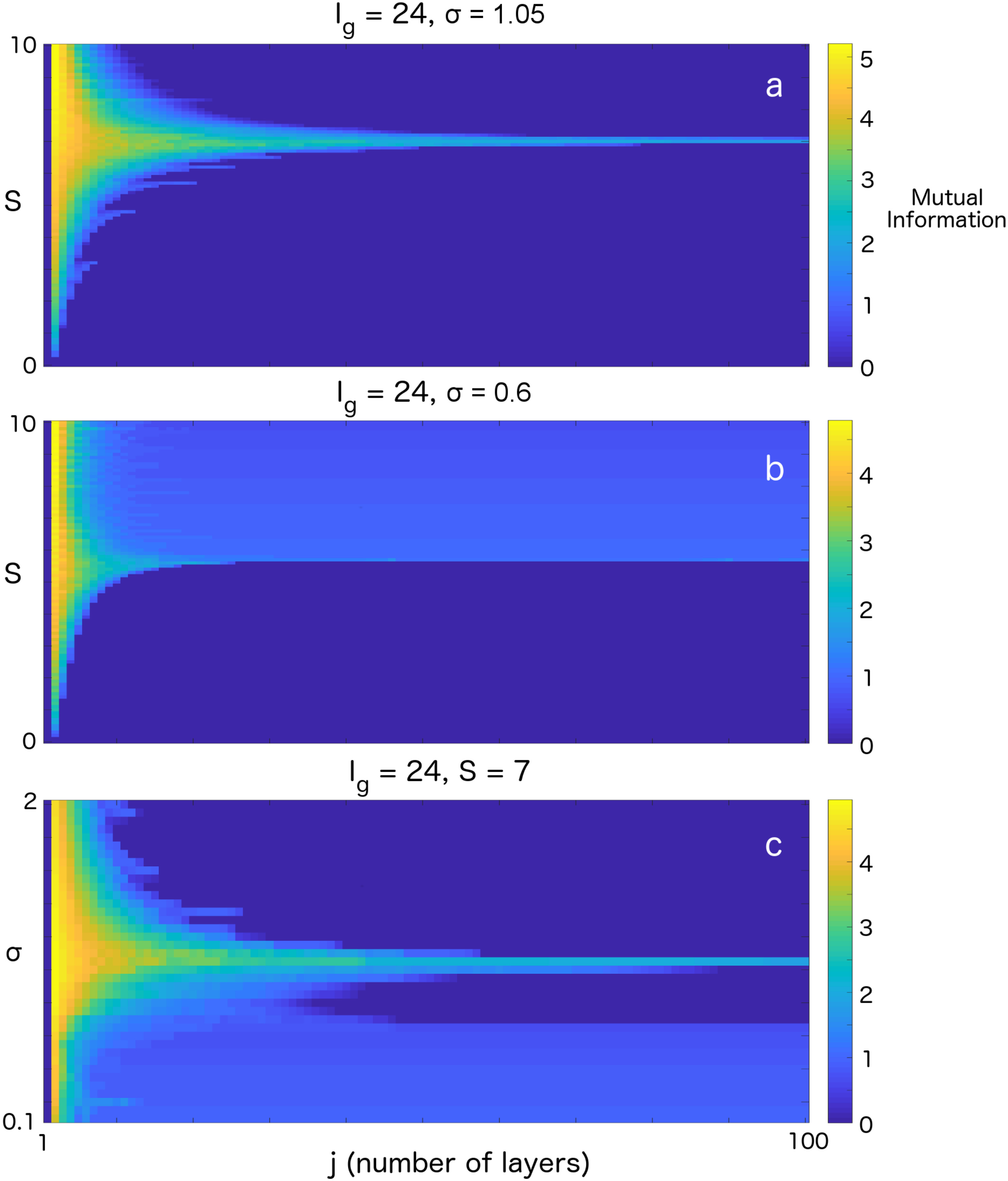
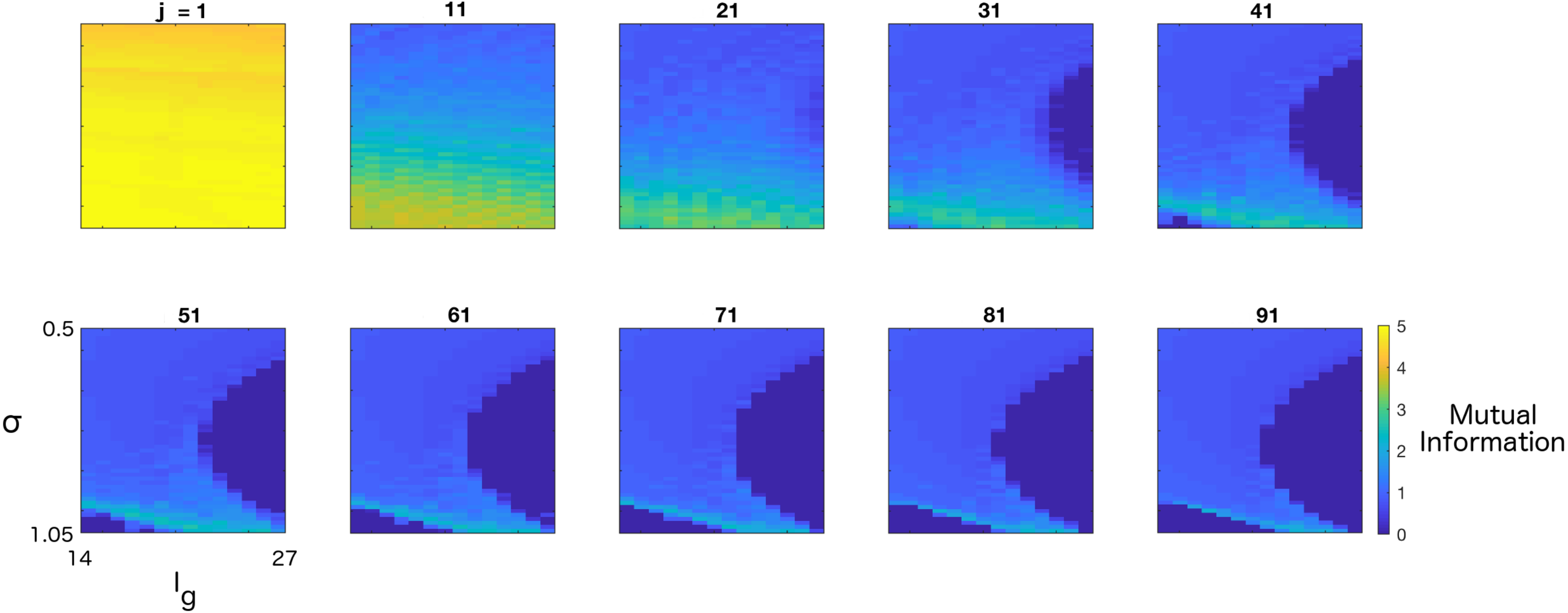
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gray, C.; König, P.; Engel, A.; Singer, W. Oscillatory responses in cat visual cortex exhibit inter-columnar synchronization which reflects global stimulus properties. Nature 1989, 338, 334–337. [Google Scholar] [CrossRef] [PubMed]
- Livingstone, M. Oscillatory firing and interneuronal correlations in squirrel monkey striate cortex. J. Neurophysiol. 1996, 66, 2467–2485. [Google Scholar] [CrossRef] [PubMed]
- Bragin, A.; Jandó, G.; Nádasdy, Z.; Hetke, J.; Wise, K.; Buzsáki, G. Gamma (40–100 Hz) oscillation in the hippocampus of the behaving rat. J. Neurosci. 1995, 15, 47–60. [Google Scholar] [PubMed]
- Brosch, M.; Budinger, E.; Scheich, H. Stimulus-related gamma oscillations in primate auditory cortex. J. Neurophysiol. 2002, 87, 2715–2725. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.; Oostenveld, R.; Peeters, M.; Fries, P. Tactile spatial attention enhances gamma-band activity in somatosensory cortex and reduces low-frequency activity in parieto-occipital areas. J. Neurosci. 2006, 26, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Pesaran, B.; Pezaris, J.; Sahani, M.; Mitra, P.; Andersen, R. Temporal structure in neuronal activity during working memory in macaque parietal cortex. Nat. Neurosci. 2002, 5, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Sohal, V.S.; Zhang, F.; Yizhar, O.; Deisseroth, K. Parvalbumin neurons and gamma rhythms enhance cortical circuit performance. Nature 2009, 459, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Popescu, A.; Popa, D.; Paré, D. Coherent gamma oscillations couple the amygdala and striatum during learning. Nat. Neurosci. 2009, 12, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Bair, W.; Koch, C. Temporal precision of spike trains in extrastriate cortex of the behaving macaque monkey. Neural Comput. 1996, 8, 1185–1202. [Google Scholar] [CrossRef] [PubMed]
- Butts, D.A.; Weng, C.; Jin, J.; Yeh, C.I.; Lesica, N.A.; Alonso, J.M.; Stanley, G.B. Temporal precision in the neural code and the timescales of natural vision. Nature 2007, 449, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Varga, C.; Golshani, P.; Soltesz, I. Frequency-invariant temporal ordering of interneuronal discharges during hippocampal oscillations in awake mice. Proc. Natl. Acad. Sci. USA 2012, 109, E2726–E2734. [Google Scholar] [CrossRef] [PubMed]
- Reyes, A.D. Synchrony-dependent propagation of firing rate in iteratively constructed networks in vitro. Nat. Neurosci. 2003, 6, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.P.; Spencer, D.; Fellous, J.M.; Sejnowski, T.J. Synchrony of thalamocortical inputs maximizes cortical reliability. Science 2010, 328, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Sornborger, A.T.; Wang, Z.; Tao, L. A mechanism for graded, dynamically routable current propagation in pulse-gated synfire chains and implications for information coding. J. Comput. Neurosci. 2015, 39, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Sornborger, A.; Tao, L. Graded, dynamically routable information processing with synfire-gated synfire chains. PLoS Comput. Biol. 2016, 12, e1004979. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Sornborger, A.; Tao, L. A pulse-gated, predictive neural circuit. In Proceedings of the 50th Asilomar Conference on Signals, Systems and Computers, Pacific Grove, CA, USA, 6–9 November 2016. [Google Scholar]
- Xiao, Z.; Zhang, J.; Sornborger, A.; Tao, L. Cusps enable line attractors for neural computation. Phys. Rev. E 2017, 96, 052308. [Google Scholar] [CrossRef] [PubMed]
- Abeles, M. Role of the cortical neuron: Integrator or coincidence detector? Isr. J. Med. Sci. 1982, 18, 83–92. [Google Scholar] [PubMed]
- König, P.; Engel, A.; Singer, W. Integrator or coincidence detector? The role of the cortical neuron revisited. Trends Neurosci. 1996, 19, 130–137. [Google Scholar] [CrossRef]
- Fries, P. A mechanism for cognitive dynamics: Neuronal communication through neuronal coherence. Trends Cogn. Sci. 2005, 9, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Diesmann, M.; Gewaltig, M.O.; Aertsen, A. Stable propagation of synchronous spiking in cortical neural networks. Nature 1999, 402, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Rubin, J.; Terman, D. High frequency stimulation of the subthalamic nucleus eliminates pathological thalamic rhythmicity in a computational model. J. Comput. Neurosci. 2004, 16, 211–235. [Google Scholar] [CrossRef] [PubMed]
- Van Rossum, M.C.; Turrigiano, G.G.; Nelson, S.B. Fast propagation of firing rates through layered networks of noisy neurons. J. Neurosci. 2002, 22, 1956–1966. [Google Scholar] [PubMed]
- Kistler, W.; Gerstner, W. Stable propagation of activity pulses in populations of spiking neurons. Neural Comput. 2002, 14, 987–997. [Google Scholar] [CrossRef] [PubMed]
- Litvak, V.; Sompolinsky, H.; Segev, I.; Abeles, M. On the transmission of rate code in long feedforward networks with excitatory-inhibitory balance. J. Neurosci. 2003, 23, 3006–3015. [Google Scholar] [PubMed]
- Kumar, A.; Rotter, S.; Aertsen, A. Conditions for propagating synchronous spiking and asynchronous firing rates in a cortical network model. J. Neurosci. 2008, 28, 5268–5280. [Google Scholar] [CrossRef] [PubMed]
- Nemenman, I.; Lewen, G.D.; Bialek, W.; de Ruyter van Steveninck, R.R. Neural coding of natural stimuli: Information at sub-millisecond resolution. PLoS Comput. Biol. 2008, 4, e1000025. [Google Scholar] [CrossRef] [PubMed]
- Moldakarimov, S.; Bazhenov, M.; Sejnowski, T.J. Feedback stabilizes propagation of synchronous spiking in cortical neural networks. Proc. Natl. Acad. Sci. USA 2015, 112, 2545–2550. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Rotter, S.; Aertsen, A. Spiking activity propagation in neuronal networks: Reconciling different perspectives on neural coding. Nat. Rev. Neurosci. 2010, 11, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Cannon, J. Analytical calculation of mutual information between weakly coupled poisson-spiking neurons in models of dynamically gated communication. Neural Comput. 2017, 29, 118–145. [Google Scholar] [CrossRef] [PubMed]
- Shannon, C. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Cover, T.; Thomas, J. Elements of Information Theory, 2nd ed.; Wiley: Hoboken, NJ, USA, 2006. [Google Scholar]
- Rieke, F.; Warland, D.; de Ruyter van Steveninck, R.; Bialek, W. Spikes: Exploring the Neural Code; The MIT Press: Cambridge, MA, USA, 1997. [Google Scholar]
- Paninski, L. Estimation of entropy and mutual information. Neural Comput. 2003, 15, 1191–1253. [Google Scholar] [CrossRef]
- Victor, J.D. Binless strategies for estimation of information from neural data. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2002, 66, 051903. [Google Scholar] [CrossRef] [PubMed]
- Nemenman, I.; Bialek, W.; de Ruyter van Steveninck, R. Entropy and information in neural spike trains: progress on the sampling problem. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2004, 69, 056111. [Google Scholar] [CrossRef] [PubMed]
- Shapira, A.H.; Nelken, I. Binless Estimation of Mutual Information in Metric Spaces. In Spike Timing: Mechanisms and Function; DiLorenzo, P., Victor, J., Eds.; CRC Press: Boca Raton, FL, USA, 2013; Chapter 5; pp. 121–135. [Google Scholar]
- Lopes-dos Santos, V.; Panzeri, S.; Kayser, C.; Diamond, M.E.; Quian Quiroga, R. Extracting information in spike time patterns with wavelets and information theory. J. Neurophysiol. 2015, 113, 1015–1033. [Google Scholar] [CrossRef] [PubMed]
- Shew, W.L.; Plenz, D. The functional benefits of criticality in the cortex. Neuroscientist 2013, 19, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Shew, W.L.; Yang, H.; Yu, S.; Roy, R.; Plenz, D. Information capacity and transmission are maximized in balanced cortical networks with neuronal avalanches. J. Neurosci. 2011, 31, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.J.; Sejnowski, T.J. An information-maximization approach to blind separation and blind deconvolution. Neural Comput. 1995, 7, 1129–1159. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.J.; Sejnowski, T.J. The “independent components” of natural scenes are edge filters. Vis. Res. 1997, 37, 3327–3338. [Google Scholar] [CrossRef]
- Yarrow, S.; Challis, E.; Series, P. Fisher and Shannon information in finite neural populations. Neural Comput. 2012, 24, 1740–1780. [Google Scholar] [CrossRef] [PubMed]
- Quiroga, R.Q.; Panzeri, S. Extracting information from neuronal populations: Information theory and decoding approaches. Nat. Rev. Neurosci. 2009, 10, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Fries, P.; Reynolds, J.; Rorie, A.; Desimone, R. Modulation of oscillatory neuronal synchronization by selective visual attention. Science 2001, 291, 1560–1563. [Google Scholar] [CrossRef] [PubMed]
- Salinas, E.; Sejnowski, T. Correlated neuronal activity and the flow of neural information. Nat. Rev. Neurosci. 2001, 2, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Csicsvari, J.; Jamieson, B.; Wise, K.; Buzsáki, G. Mechanisms of gamma oscillations in the hippocampus of the behaving rat. Neuron 2003, 37, 311–322. [Google Scholar] [CrossRef]
- Womelsdorf, T.; Schoffelen, J.; Oostenveld, R.; Singer, W.; Desimone, R.; Engel, A.; Fries, P. Modulation of neuronal interactions through neuronal synchronization. Science 2007, 316, 1609–1612. [Google Scholar] [CrossRef] [PubMed]
- Colgin, L.; Denninger, T.; Fyhn, M.; Hafting, T.; Bonnevie, T.; Jensen, O.; Moser, M.; Moser, E. Frequency of gamma oscillations routes flow of information in the hippocampus. Nature 2009, 462, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Luczak, A.; McNaughton, B.L.; Harris, K.D. Packet-based communication in the cortex. Nat. Rev. Neurosci. 2015, 16, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Yuste, R.; MacLean, J.; Smith, J.; Lansner, A. The cortex as a central pattern generator. Nat. Rev. Neurosci. 2005, 6, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Hubel, D.; Wiesel, T. Receptive fields and functional architecture in two non striate visual areas (18 and 19) of the cat. J. Neurophysiol. 1965, 28, 229–289. [Google Scholar] [CrossRef] [PubMed]
- DeWeese, M.; Wehr, M.; Zador, A. Binary coding in auditory cortex. J. Neurosci. 2003, 23, 7940–7949. [Google Scholar] [PubMed]
- Olshausen, B.; Field, D. Sparse coding of sensory inputs. Curr. Opin. Neurobiol. 2004, 14, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Hromádka, T.; DeWeese, M.; Zador, A. Sparse representation of sounds in the unanesthetized auditory cortex. PLoS Biol. 2008, 6, e16. [Google Scholar] [CrossRef] [PubMed]
- Piet, A.T.; Erlich, J.C.; Kopec, C.D.; Brody, C.D. Rat Prefrontal Cortex Inactivations during Decision Making Are Explained by Bistable Attractor Dynamics. Neural Comput. 2017, 29, 2861–2886. [Google Scholar] [CrossRef] [PubMed]
- Brody, C.D.; Hernandez, A.; Zainos, A.; Romo, R. Timing and neural encoding of somatosensory parametric working memory in macaque prefrontal cortex. Cereb. Cortex 2003, 13, 1196–1207. [Google Scholar] [CrossRef] [PubMed]
- Seung, H.S. How the brain keeps the eyes still. Proc. Natl. Acad. Sci. USA 1996, 93, 13339–13344. [Google Scholar] [CrossRef] [PubMed]
- Koulakov, A.A.; Raghavachari, S.; Kepecs, A.; Lisman, J.E. Model for a robust neural integrator. Nat. Neurosci. 2002, 5, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Goldman, M. Memory without feedback in a neural network. Neuron 2008, 61, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Brody, C.D.; Romo, R.; Kepecs, A. Basic mechanisms for graded persistent activity: Discrete attractors, continuous attractors, and dynamic representations. Curr. Opin. Neurobiol. 2003, 13, 204–211. [Google Scholar] [CrossRef]
- Major, G.; Tank, D. Persistent neural activity: Prevalence and mechanisms. Curr. Opin. Neurobiol. 2004, 14, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Aksay, E.; Olasagasti, I.; Mensh, B.D.; Baker, R.; Goldman, M.S.; Tank, D.W. Functional dissection of circuitry in a neural integrator. Nat. Neurosci. 2007, 10, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Mante, V.; Sussillo, D.; Shenoy, K.V.; Newsome, W.T. Context-dependent computation by recurrent dynamics in prefrontal cortex. Nature 2013, 503, 78–84. [Google Scholar] [CrossRef] [PubMed]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, Z.; Wang, B.; Sornborger, A.T.; Tao, L. Mutual Information and Information Gating in Synfire Chains. Entropy 2018, 20, 102. https://doi.org/10.3390/e20020102
Xiao Z, Wang B, Sornborger AT, Tao L. Mutual Information and Information Gating in Synfire Chains. Entropy. 2018; 20(2):102. https://doi.org/10.3390/e20020102
Chicago/Turabian StyleXiao, Zhuocheng, Binxu Wang, Andrew T. Sornborger, and Louis Tao. 2018. "Mutual Information and Information Gating in Synfire Chains" Entropy 20, no. 2: 102. https://doi.org/10.3390/e20020102
APA StyleXiao, Z., Wang, B., Sornborger, A. T., & Tao, L. (2018). Mutual Information and Information Gating in Synfire Chains. Entropy, 20(2), 102. https://doi.org/10.3390/e20020102