Is Cetacean Intelligence Special? New Perspectives on the Debate


Abstract
1. Introduction
2. Materials and Methods
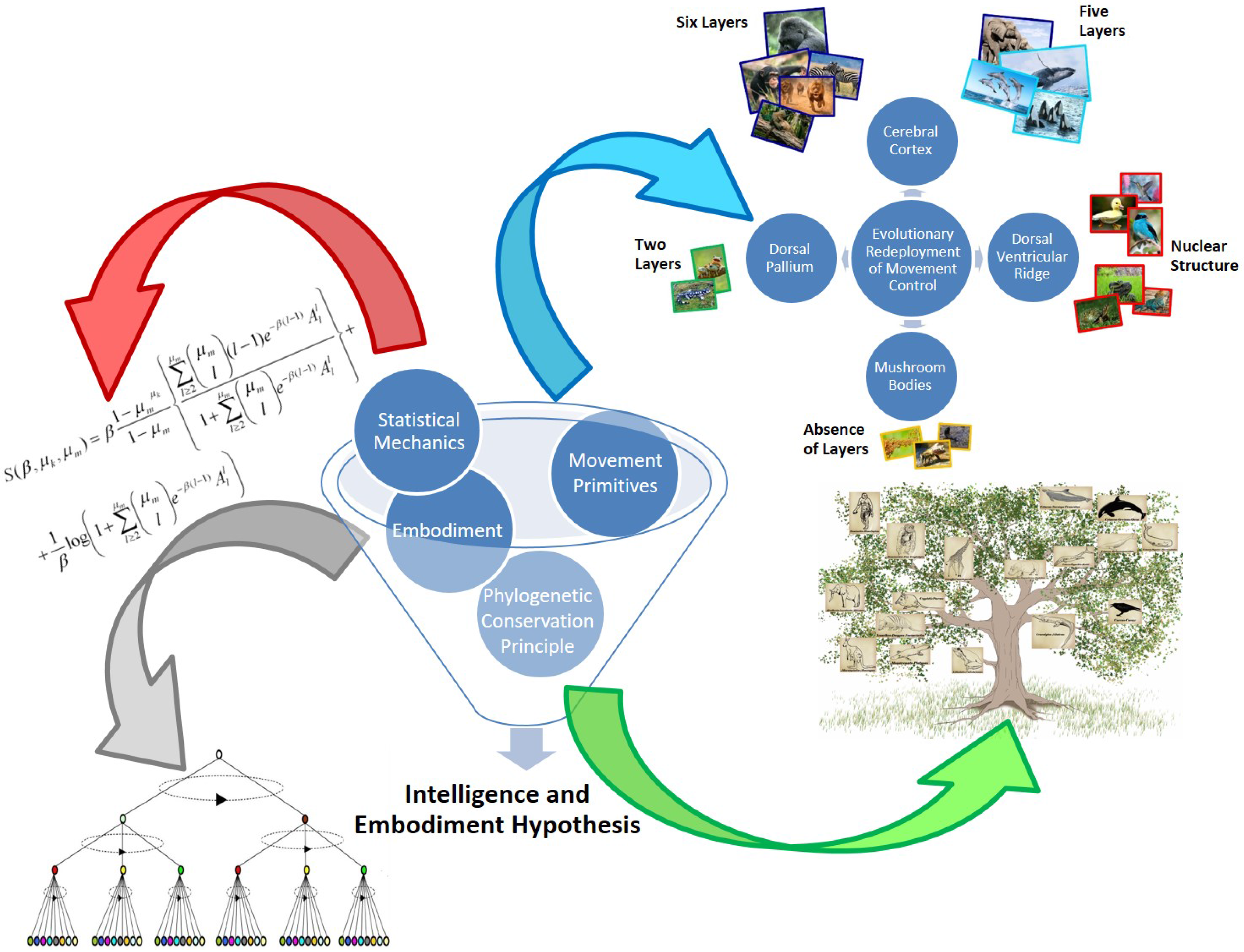
2.1. Hypothesis Background
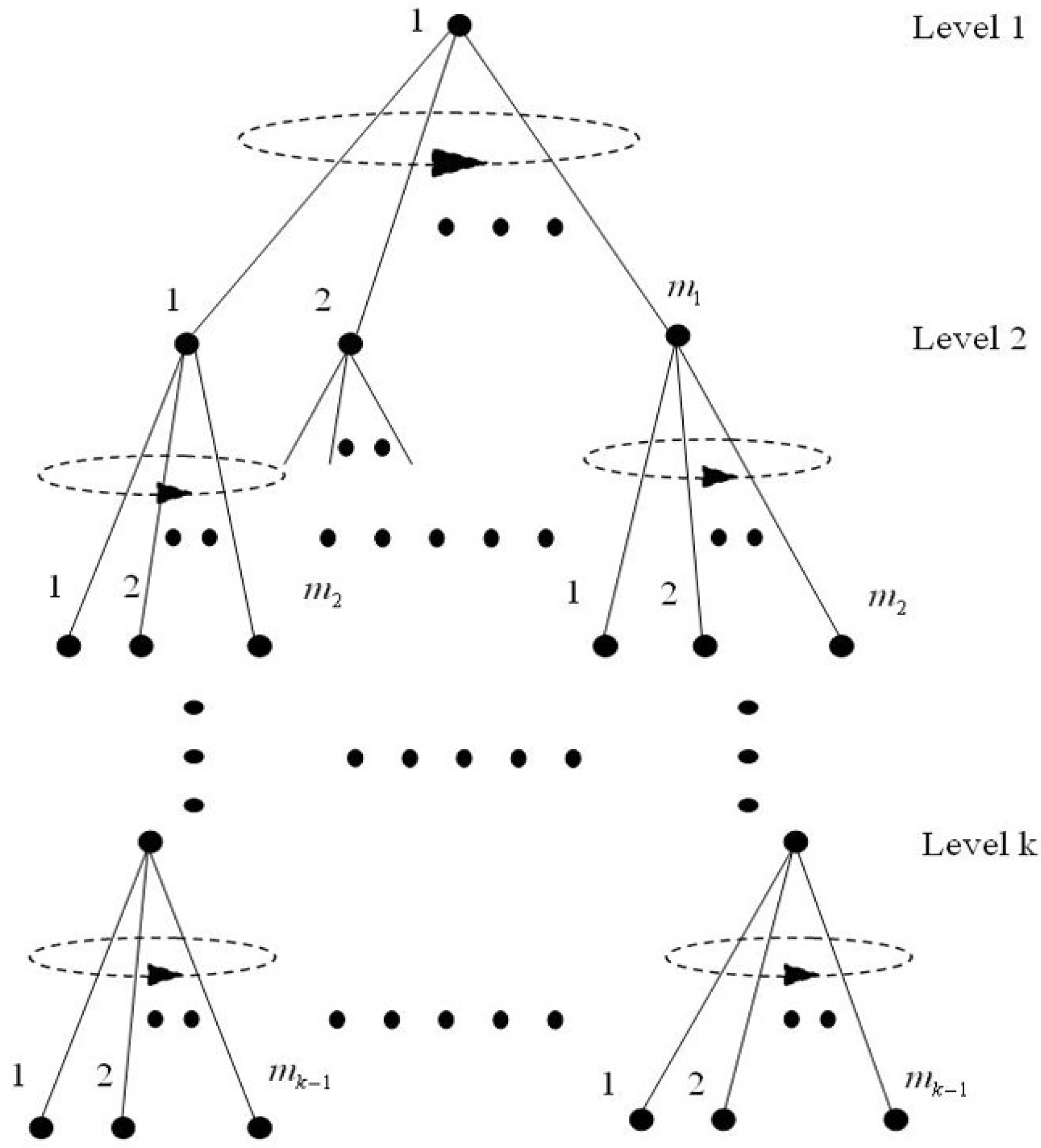
2.2. Statistical Mechanics
2.3. The Principal Actor of Energy Consumption in the Brain
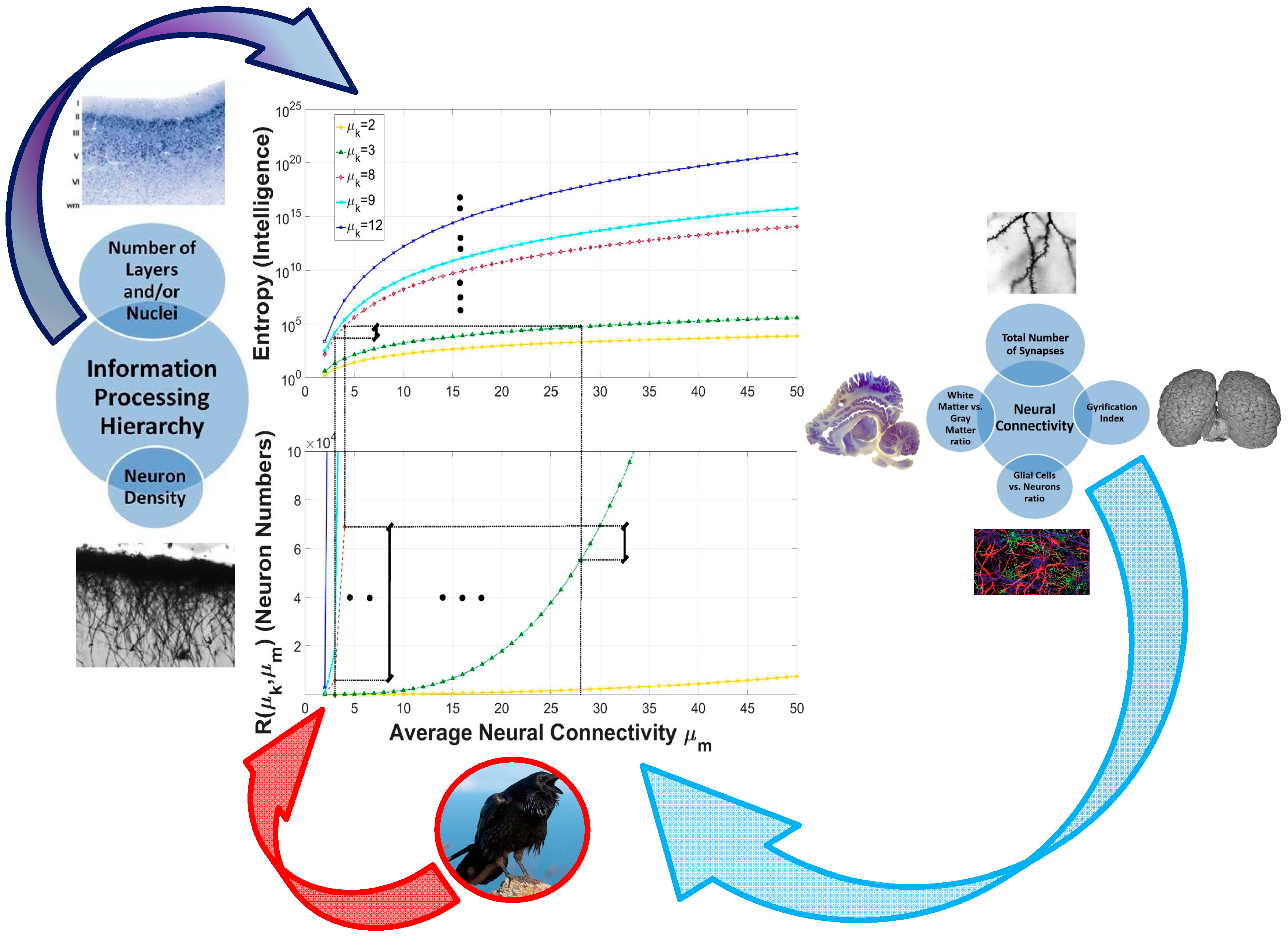
2.4. Neurophysiologic Indicators of Neural Connectivity
2.5. An Extension of the Mathematical Model of the Hypothesis
- These studies demonstrated that the pattern of functional connectivity between cortical areas was consistent with a small-world network architecture. This is of particular interest since this architecture is mainly characterized by being modular and hierarchical. Furthermore, these studies also revealed that as a result of the constraints imposed by the structural connectivity in the functional interactions, the topological parameters are generally conserved between structural and functional networks.
- The patterns of functional connectivity between cortical regions undergo spontaneous fluctuations and are highly responsive to perturbations (i.e., sensory input or cognitive tasks) on a timescale of hundreds of milliseconds. However, these rapid reconfigurations do not affect the stability of global topological characteristics. In other words, on longer timescales of seconds to minutes (i.e., in equilibrium from a statistical mechanics point of view), correlations between spontaneous fluctuations in brain activity form functional networks that are particularly robust.
Physical Interpretation of Equation
- If the average connectivity between symbols is fixed to any given value, for example, then increasing the average number of information processing levels from to not only increases the average number of symbols encoded, i.e., , but also the associated entropy values . It is important to remember that if the connectivity is fixed, then the differences in the information-content in the structures varies exponentially when different numbers of information-processing levels are considered [28].
- Similarly, if the average number of information processing levels is fixed to , then if the average connectivity is increased, for example, from to , then the average number of symbols encoded and the associated entropy values also increases, i.e., and . However, in this case the values of the free energy (in absolute value) are also higher (i.e., ).
- Higher entropy values are always linked to structures encoding a higher number of symbols independently of the parameterization of and . For example, if then . Similarly, if then .
- For all , if the average connectivity is fixed to any particular value, for example, , there always exists a value for the average connectivity , such as the values that are reached by the entropy function in these points, i.e., and respectively, that is identical (or very close because the entropy is an integer function), i.e., , then the following inequalities hold: and . In other words, the average number of representations is higher for the point in the space of parameters with the highest number of information-processing levels, i.e., ) in this case, but at the same time the values of the free energy are lower compared to the point in the space of parameters with a lower number of information-processing levels but a higher average connectivity, i.e., ).
3. Results
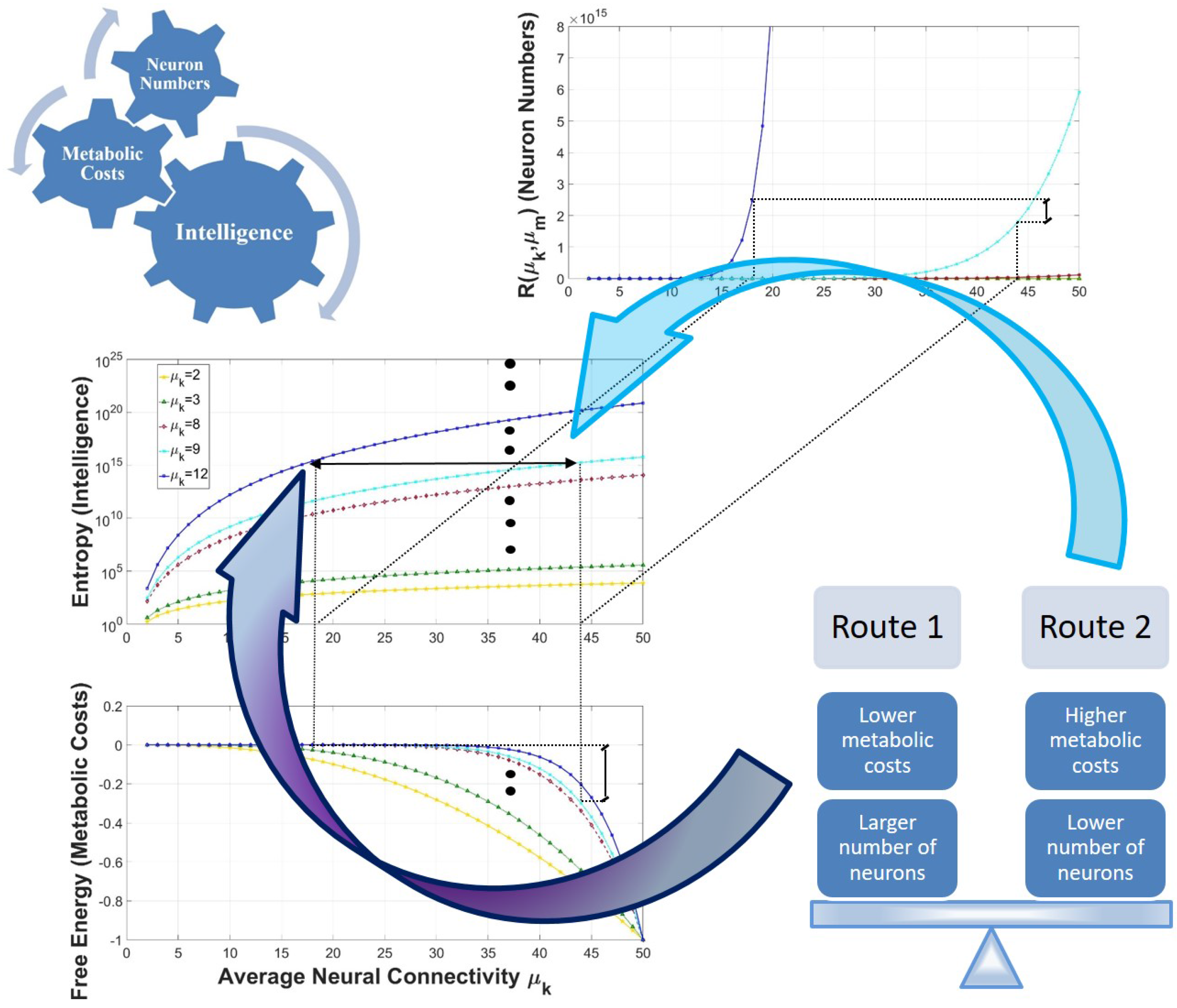
3.1. An Equation Relating Intelligence, Energetic Requirements, and Neuron Numbers
3.2. The Limiting Factors of Intelligence: Neuron Numbers and Energetic Requirements
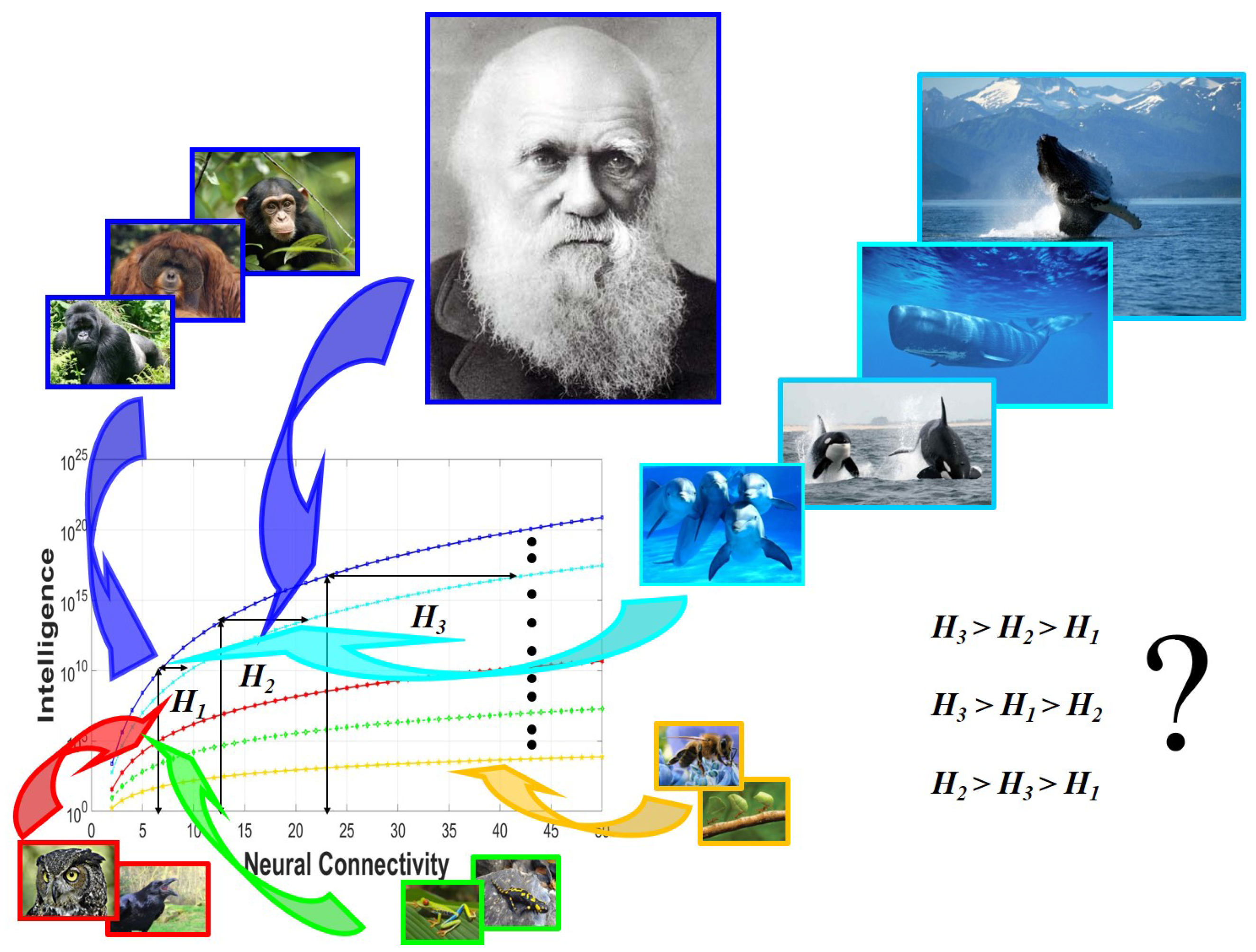
3.3. Two Routes for the Attainment of Higher Levels of Intelligence
3.4. The Primary Role of Connectivity in Intelligence
4. Discussion
4.1. Implications for Cetacean Cognition
4.2. Brain Evolution: The Loss of Layer IV in the Cerebral Cortex
Terrestrial Mammals Lacking Internal Granular Layer IV in the Cerebral Cortex
4.3. Brain Evolution: Large Brains, Thinner Cortices, and an Extreme Gyrencephaly
5. Conclusions
Supplementary Materials
Acknowledgments
Conflicts of Interest
Abbreviations
| RMR | Resting Metabolic Rate |
| DVR | Dorsal Ventricular Ridge |
| NPD | Neuron Packing Density |
References
- Butti, C.; Raghanti, M.A.; Sherwood, C.C.; Hof, P.R. The neocortex of cetaceans: Cytoarchitecture and comparison with other aquatic and terrestrial species. Ann. N. Y. Acad. Sci. 2011, 1225, 7–58. [Google Scholar] [CrossRef] [PubMed]
- Manger, P.R. An examination of cetacean brain structure with a novel hypothesis correlating thermogenesis to the evolution of a big brain. Biol. Rev. 2006, 81, 293–338. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, N.; Pakkenberg, B. Total neocortical cell number in the Mysticete brain. Anat. Rec. 2007, 290, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Hof, P.R.; Van Der Gucht, E. Structure of the cerebral cortex of the Humpback whale, Megaptera Novaeangliae (Cetacean, Mysticety, Balaenopteridae). Anat. Rec. A 2007, 290, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Kojima, T. On the brain of the sperm whale (Physeter catodon L.). Sci. Rep. Whales Res. Inst. Tokyo 1951, 6, 49–72. [Google Scholar]
- Ridgway, S.H.; Hanson, A.C. Sperm Whales and Killer Whales with the Largest Brain of All Toothed Whales Show Extreme Differences in Cerebellum. Brain Behav. Evol. 2014, 83, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Lyamin, O.I.; Manger, P.R.; Ridgway, S.H.; Mukhameov, L.M.; Siegel, J.M. Cetacean sleep: An unusual form of mammalian sleep. Neurosci. Biobehav. Rev. 2008, 32, 1451–1484. [Google Scholar] [CrossRef] [PubMed]
- Ridgway, S.H. The central nervous system of the bottlenose dolphin. In The Bottlenose Dolphin; Leatherwood, S., Reeves, R.R., Eds.; Academic Press: New York, NY, USA, 1990; pp. 69–97. [Google Scholar]
- Isler, K.; van Schaik, P. Metabolic costs of brain size evolution. Biol. Lett. 2006, 2, 557–560. [Google Scholar] [CrossRef]
- Mota, B.; Herculano-Houzel, S. How the cortex gets its folds and inside-out connectivity-driven model for the scaling of mammalian cortical folding. Front. Neuroanat. 2012. [Google Scholar] [CrossRef] [PubMed]
- Prothero, J.W.; Sundsten, J.W. Folding of the cerebral cortex in mammals. Brain Behav. Evol. 1984, 24, 152–167. [Google Scholar] [CrossRef] [PubMed]
- Herculano-Houzel, S. Not All Brains Are Made the Same: New Views on Brain Scaling in Evolution. Brain Behav. Evol. 2011, 78, 22–36. [Google Scholar] [CrossRef] [PubMed]
- Striedter, G.F. Principles of Brain Evolution; Sinauer Associates: Sunderland, MA, USA, 2005. [Google Scholar]
- Connor, R.C. Dolphin social intelligence: Complex alliance relationships in bottlenose dolphins and a consideration of selective environments for extreme brain size evolution in mammals. Philos. Trans. R. Soc. B 2007, 362, 587–602. [Google Scholar]
- Oelschläger, H.H.A.; Kemp, B. Ontogenesis of the sperm whale brain. J. Comp. Neurol. 1998, 399, 210–228. [Google Scholar] [CrossRef]
- Ridgway, S.H.; Au, W.W.L. Hearing and echolocation: Dolphin. In Elsevier’s Encyclopedia of Neuroscience, 2nd ed.; Adelman, G., Smith, B.H., Eds.; Elsevier: New York, NY, USA, 1999; pp. 858–862. [Google Scholar]
- Oelschläger, H.H.A.; Oelschläger, J. Brain. In Encyclopedia of Marine Mammals; Wursig, B., Thewissen, J.G.M., Eds.; Academic Press: San Diego, CA, USA, 2008; pp. 134–149. [Google Scholar]
- Güntürkun, O. Is dolphin cognition special? Brain Behav. Evol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Huggenberger, S. The size and complexity of dolphin brains a paradox? J. Mar. Biol. Assoc. UK 2008, 88, 1103–1108. [Google Scholar] [CrossRef]
- Manger, P.R. Questioning the interpretation of behavioral observations of cetaceans: Is there really support for a special intellectual status for this mammalian order? Neuroscience 2013, 250, 664–696. [Google Scholar] [CrossRef] [PubMed]
- Marino, L. Dolphin cognition. Curr. Biol. 2004, 14, R910–R911. [Google Scholar] [CrossRef] [PubMed]
- Marino, L.; Butti, C.; Connor, R.C.; Fordyce, R.E.; Herman, L.M.; Hof, P.R.; Lefevre, L.; Lusseau, D.; McCowan, B.; Nimchincky, E.A.; et al. A claim in search of evidence: Reply to Manger’s thermogenesis hypothesis of cetacean brain structure. Biol. Rev. Camb. Philos. 2008, 83, 417–440. [Google Scholar] [CrossRef] [PubMed]
- Patzke, N.; Spocter, M.A.; Karlsson, K.A.; Bertelsen, M.F.; Haagensen, M.; Chawana, R.; Streicher, S.; Kaswera, C.; Gilissen, E.; Alagaili, A.N.; et al. In contrast to many other mammals, cetaceans have relatively small hippocampi that appear to lack adult neurogenesis. Brain Struct. Funct. 2013, 220, 361–383. [Google Scholar] [CrossRef] [PubMed]
- Hart, B.L.; Hart, L.A.; Pinter-Wollman, N. Large brains and cognition: Where do elephants fit in? Neurosci. Biobehav. R. 2008, 32, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Mann, J.; Patterson, E.M. Tool use by aquatic animals. Philos. Trans. R. Soc. B 2013, 368. [Google Scholar] [CrossRef] [PubMed]
- Zentall, T.R. Animal intelligence. In The Cambridge Handbook of Intelligence; Sternberg, R.J., Kaufman, S.B., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 309–327. [Google Scholar]
- Chittka, L.; Rossiter, S.J.; Skorupski, P.; Fernando, C. What is comparable in comparative cognition? Philos. Trans. R. Soc. B 2012, 367, 2677–2685. [Google Scholar] [CrossRef] [PubMed]
- Chinea, A.; Korutcheva, E. Intelligence and embodiment: A statistical mechanics approach. Neural Netw. 2013, 40, 52–72. [Google Scholar] [CrossRef] [PubMed]
- Ridgway, S.H.; Carln, P.C.; Van Alstyne, K.R.; Hanson, A.C.; Tarpley, R.J. Comparison of Dolphins’ Body and Brain Measurements with Four other Groups of Cetaceans Reveals Great Diversity. Brain Behav. Evol. 2016, 88, 235–257. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, H.S.; Pakkenberg, B.; Dam, M.; Dietz, R.; Sonne, C.; Mikkelsen, B.; Eriksen, N. Quantitative relationships in delphinid neocortex. Front. Neuroanat. 2014, 8, 132. [Google Scholar] [CrossRef] [PubMed]
- Hecht-Nielsen, R. Confabulation Theory: The Mechanism of Thought; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Tomer, R.; Denes, A.S.; Tessmar-Raible, K.; Arendt, D. Profiling by image registration reveals common origin of annelid mushroom bodies and vertebrate pallium. Cell 2010, 142, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Dugas-Ford, J.; Rowell, J.J.; Ragsdale, C.W. Cell-type homologies and the origins of the neocortex. Proc. Natl. Acad. Sci. USA 2012, 109, 16974–16979. [Google Scholar] [CrossRef] [PubMed]
- Karten, H.J. Vertebrate brains and evolutionary connectomics: On the origins of the mammalian ‘neocortex’. Philos. Trans. R. Soc. B 2015, 370, 60. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.; Hobbs, J.N.; Burroni, J.; Siegelmann, H. The global landscape of cognition:hierarchical aggregation as an organizational principle of human cortical networks and functions. Sci. Rep. 2015, 5, 18112. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, A.; Woolley, S.M.N. Coding principles in the avian brain. Proc. Natl. Acad. Sci. USA 2015, 112, 3517–3522. [Google Scholar] [CrossRef] [PubMed]
- Felleman, D.J.; Van Essen, D.C. Distributed hierarchical processing in the primate cerebral cortex. Cereb. Cortex 1991, 1. [Google Scholar] [CrossRef]
- Salazar, R.F.; Dotson, N.M.; Bressler, S.L.; Gray, C.M. Content Specific Fronto-Parietal Synchronization during Visual Working Memory. Science 2012, 338, 1097–1100. [Google Scholar] [CrossRef] [PubMed]
- Churchland, P.; Sejnowski, T. The Computational Brain; MIT Press: Cambridge, MA, USA, 1992. [Google Scholar]
- Chandler, D. Introduction to Modern Statistical Mechanics; Oxford University Press: Oxford, UK, 1987. [Google Scholar]
- Reichl, L.E. A Modern Course in Statistical Physics; Wiley: New York, NY, USA, 1998. [Google Scholar]
- Hermann, C. Statistical Physics (Including Applications to Condensed Matter); Springer: New York, NY, USA, 2005. [Google Scholar]
- Howarth, C.; Gleeson, P.; Attwell, D. Updated energy budgets for neural computation in the neocortex and cerebellum. J. Cerebr. Blood Flow Metab. 2012, 32, 1222–1232. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, S.; Stimpson, C.D.; Bauernfeind, A.L.; Schapiro, S.J.; Baze, W.B.; McArthur, M.J.; Bronson, E.; Hopkins, W.D.; Semendeferi, K.; Jacobs, B.; et al. Dendritic morphology of pyramidal neurons in the chimpanzee neocortex: Regional specializations and comparison to humans. Cereb. Cortex 2013, 23, 2429–2436. [Google Scholar]
- Butti, C.; Janeway, C.M.; Townshend, C.; Wicinski, B.A.; Reidenberg, J.S.; Ridgway, S.H.; Sherwood, C.C.; Hof, P.R.; Jacobs, B. The neocortex of cetartiodactyls: I. A comparative Golgi analysis of neuronal morphology in the bottlenose dolphin (Tursiops truncatus), the minke whale (Balaenoptera acutorostrata), and the humpback whale (Megaptera novaeangliae). Brain Struct. Funct. 2015, 220, 3339–3368. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, B.; Schall, M.; Prather, M.; Kapler, E.; Driscoll, L.; Baca, S.; Jacobs, J.; Ford, K.; Wainwright, M.; Treml, M. Regional dendritic and spine variation in human cerebral cortex: A quantitative golgi study. Cereb. Cortex 2001, 11, 558–571. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, B.; Harland, T.; Kennedy, D.; Schall, M.; Wicinski, B.; Butti, C.; Hof, P.R.; Sherwood, C.C.; Manger, P.R. The neocortex of cetartiodactyls. II. Neuronal morphology of the visual and motor cortices in the giraffe (Giraffa camelopardalis). Brain Struct. Funct. 2015, 220, 2851–2872. [Google Scholar] [CrossRef] [PubMed]
- Nieuwenhuys, R.; Ten Donkelaar, H.J.; Nicholson, C. The Central Nervous System of Vertebrates; Springer: Berlin, Germany, 1998; Volume 3. [Google Scholar]
- Van Essen, D.C. A tension-based theory of morphogenesis and compact wiring in the central nervous system. Nature 1997, 385, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Harrison, K.H.; Hof, P.; Wang, S.S.H. Scaling laws in the mammalian neocortex: Does form provides clues to function? J. Neurocytol. 2002, 31, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Ventura-Antunes, L.; Mota, B.; Herculano-Houzel, S. Different scaling of white matter volume, cortical connectivity, and gyrification across rodent and primate brains. Front. Neuroanat. 2013, 7. [Google Scholar] [CrossRef]
- Herculano-Houzel, S.; Manger, P.R.; Kaas, J.H. Brain scaling in mammalian evolution as a consequence of concerted and mosaic changes in number of neurons and average neuronal size. Front. Neuroanat. 2014, 8, 77. [Google Scholar] [CrossRef] [PubMed]
- Goodman, M.; Sterner, K.N.; Islam, M.; Uddin, M.; Sherwood, C.C.; Hof, P.R.; Hou, Z.C.; Lipovich, L.; Jia, H.; Grossman, L.I.; et al. Phylogenetic analyses reveal convergent patterns of adaptive evolution in elephants and human ancestries. Proc. Natl. Acad. Sci. USA 2009, 106, 20824–20829. [Google Scholar] [CrossRef] [PubMed]
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Bedny, M.; Caramazza, A. Perception, action, and word meanings in the human brain: The case from action verbs. Ann. N. Y. Acad. Sci. 2011, 1224, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Pullvermüller, F.; Huss, M.; Kheriff, F.; Moscoso del Prado Martin, F.; Hauk, O. Motor cortex maps articulatory features of speech sounds. Proc. Natl. Acad. Sci. USA 2006, 103, 7865–7870. [Google Scholar] [CrossRef] [PubMed]
- Pullvermüller, F. Semantic embodiment, disembodiment or misembodiment? In search of meaning in modules and neuron circuits. Brain Lang. 2013, 127, 86–103. [Google Scholar] [CrossRef] [PubMed]
- Herculano-Houzel, S.; Collins, C.E.; Wong, P.; Kaas, J.H. The basic nonuniformity of the cerebral cortex. Proc. Natl. Acad. Sci. USA 2010, 105, 12593–12598. [Google Scholar] [CrossRef] [PubMed]
- Nieuwenhuys, R.; Voogd, J.; Van Huijzen, C. The Human Central Nervous System; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Wang, Y.; Brzozowska-Prechtl, A.; Karten, H.J. Laminar and columnar auditory cortex in avian brain. Proc. Natl. Acad. Sci. USA 2010, 107, 12676–12681. [Google Scholar] [CrossRef] [PubMed]
- Ahumada-Galleguillos, P.; Fernandez, M.; Marin, G.J.; Letelier, J.C.; Mpodozis, J. Anatomical Organization of the Visual Dorsal Ventricular Ridge in the Chick (Gallus gallus): Layers and Columns in the Avian Pallium. J. Comp. Neurol. 2015, 406, 329–345. [Google Scholar] [CrossRef] [PubMed]
- Husband, S.A.; Shimizu, T. Efferent projections of the ectostriatum in the pigeon Columba livia. J. Comp. Neurol. 1999, 406, 329–345. [Google Scholar] [CrossRef]
- Ulinsky, P. Dorsal Ventricular Ridge: A Treatise on Forebrain Organization in Reptiles and Birds; Wiley: New York, NY, USA, 1983. [Google Scholar]
- Manger, P.R.; Slutsky, D.A.; Molnar, Z. Visual subdivisions of the dorsal ventricular ridge of the iguana (Iguana iguana) as determined by electrophysiologic mapping. J. Comp. Neurol. 2002, 453, 226–246. [Google Scholar] [CrossRef] [PubMed]
- McGowen, M.R.; Grossman, L.I.; Wildman, D.E. Dolphin genome provides evidence for adaptive evolution of nervous system genes and molecular rate slowdown. Philos. Trans. R. Soc. B 2012, 279, 3643–3651. [Google Scholar] [CrossRef] [PubMed]
- Byrne, R.W.; Whiten, A. Machiavellian Intelligence: Social Expertise and the Evolution of the Intellect in Monkeys, Apes and Humans; Oxford University Press: Oxford, UK, 1988. [Google Scholar]
- Olkowicz, S.; Kocourek, M.; Lucan, R.K.; Portes, M.; Fitch, W.T.; Herculano-Houzel, S.; Nemec, P. Birds have primate-like numbers of neurons in the forebrain. Proc. Natl. Acad. Sci. USA 2016, 113, 7255–7260. [Google Scholar] [CrossRef] [PubMed]
- Schoenemann, P.T.; Sheehan, M.J.; Glotzer, D.L. Prefrontal white matter volume is disproportionately larger in humans than in other primates. Nat. Rev. Neurosci. 2005, 8, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Schoenemann, P.T. Evolution of the size and functional areas of the human brain. Annu. Rev. Anthropol. 2006, 35, 379–406. [Google Scholar] [CrossRef]
- Rilling, J.K.; Insel, T.R. The primate neocortex in comparative perspective using magnetic resonance imaging. J. Hum. Evol. 1999, 37, 191–223. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, B.; Lubs, J.; Hannan, M.; Anderson, K.; Butti, C.; Sherwood, C.C.; Hof, P.R.; Manger, P.R. Neuronal morphology in the African elephant (Loxodonta africana) neocortex. Brain Struct. Funct. 2011, 215, 273–298. [Google Scholar] [CrossRef] [PubMed]
- Douglas-Fields, R. Map the other brain. Nature 2013, 501, 25–26. [Google Scholar] [CrossRef]
- Herculano-Houzel, S.; Kaas, J.H. Gorilla and orangutan brains conform to the primate cellular scaling rules: implications for human evolution. Brain Behav. Evol. 2011, 77, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Herculano-Houzel, S.; Avelino-de-Souza, K.; Neves, K.; Porfírio, J.; Messeder, D.; Mattos Feijó, L.; Maldonado, J.; Manger, P. The elephant brain in numbers. Front. Neuroanat. 2014, 8. [Google Scholar] [CrossRef]
- Bianchi, S.; Stimpson, C.D.; Duka, T.; Larsen, M.D.; Janssen, W.G.M.; Collins, Z.; Bauernfeind, A.L.; Schapiro, S.J.; Baze, W.B.; McArthur, M.J.; et al. Synaptogenesis and development of pyramidal neuron dendritic morphology in the chimpanzee neocortex resembles humans. Proc. Natl. Acad. Sci. USA 2013, 110, 10395–10401. [Google Scholar] [CrossRef] [PubMed]
- Tomasello, M.; Call, J. The Gestural Communication of Apes and Monkeys; Lawrence Erlbaum Associates: Mahwah, NJ, USA, 2007. [Google Scholar]
- Costa, D.P.; Williams, T.M. Marine Mammals Energetics. In The Biology of Marine Mammals; Reynold, J., Twiss, J., Eds.; The Smithsonian Institution Press: Washington, DC, USA, 2000; pp. 176–211. [Google Scholar]
- Hakeem, A.Y.; Hof, P.R.; Sherwood, C.C.; Switzer, R.C.; Rasmussen, L.R.L.; Allman, J.M. Brain of the African elephant (Loxodonta africana): Neuroanatomy from magnetic resonance images. Anat. Rec. 2005, 287, 1117–1127. [Google Scholar] [CrossRef] [PubMed]
- Hof, P.R.; Chanis, R.; Marino, L. Cortical Complexity in Cetacean Brains. Anat. Rec. 2005, 287, 1142–1152. [Google Scholar] [CrossRef] [PubMed]
- McKay, G.M. Behavior and Ecology of the Asiatic Elephant in Southeastern Ceylon; Smithsonian Institution, Government Printing Office: Washington, DC, USA, 1973.
- Manger, P.R.; Prowse, M.; Haagensen, M.; Hemingway, J. A quantitative analysis of neocortical gyrencephaly in African elephants (Loxodonta Africana) and six species of cetaceans: comparisons to other mammals. J. Comp. Neurol. 2012, 520, 2430–2439. [Google Scholar] [CrossRef] [PubMed]
- Mink, J.W.; Blumenschine, R.J.; Adams, D.B. Ratio of central nervous system to body metabolism in vertebrates: Its constancy and functional basis. Am. J. Physiol. 1981, 241, 203–212. [Google Scholar]
- Leonard, W.R.; Snodgrass, J.J.; Robertson, M.L. Effects of brain evolution on human nutrition and metabolism. Annu. Rev. Nutr. 2007, 27, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Lockyer, C. All creatures great and smaller: A study in cetacean life history energetics. J. Mar. Biol. Assoc. UK 2007, 87, 1035–1045. [Google Scholar] [CrossRef]
- Sherwood, C.C.; Stimpson, C.D.; Raghanti, M.A.; Wildman, D.E.; Uddin, M.; Grossman, L.I.; Goodman, M.; Redmond, J.C.; Bonar, C.J.; Erwin, J.M. Evolution of increased glia-neuron ratios in the human frontal cortex. Proc. Natl. Acad. Sci. USA 2006, 103, 13606–13611. [Google Scholar] [CrossRef] [PubMed]
- Herman, L.M. Body and self in dolphins. Conscious. Cogn. 2012, 21, 526–545. [Google Scholar] [CrossRef] [PubMed]
- Xitco, J.M.; Gory, J.D.; Kuczaj, S.A. Spontaneous pointing by bottlenose dolphins (Tursiops truncatus). Anim. Cogn. 2001, 4, 115–123. [Google Scholar] [CrossRef]
- Visser, I.N.; Smith, T.G.; Bullock, I.D.; Green, G.D.; Carlsson, O.G.L.; Imberti, S. Antartica peninsula killer whales (Orcinus orca) hunt seals and penguin on floating ice. Mar. Mamm. Sci. 2008, 24, 225–234. [Google Scholar] [CrossRef]
- Foote, A.D.; Griffin, R.M.; Howitt, D.; Larsson, L.; Miller, P.J.O.; Hoelzel, A.R. Killer whales are capable of vocal learning. Biol. Lett. 2006, 2, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Baird, R.W. The killer whale: Foraging specializations and group hunting. In Cetacean Societies: Field Studies of Dolphins and Whales; Mann, J., Conno, R.C., Tyack, P.L., Whitehead, H., Eds.; University of Chicago Press: Chicago, IL, USA, 2000. [Google Scholar]
- Whitehead, H.; Rendell, L. The Cultural Lives of Whales and Dolphins; University of Chicago Press: Chicago, IL, USA, 2015. [Google Scholar]
- Montgomery, S.H.; Geisler, J.H.; McGowen, M.R.; Fox, C.; Marino, L.; Gatesy, J. The Evolutionary History of Cetacean Brain and Body Size. Evolution 2013, 67, 3339–3353. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, F.S.; Szabo, A.S.; Pack, A.; Nahmens, J. The social structure of bubble net feeding whales in SE Alaska. In Proceedings of the 20th Biennal Conference on the Biology of Marine Mammals, Dunedin, New Zealand, 9 December 2013. [Google Scholar]
- Stimpert, A.K.; Au, W.W.; Parks, S.E.; Hurst, T.; Wiley, D.N. Common humpback whale (Megaptera novaeangliae) sound types for passive acoustic monitoring. J. Acoust. Soc. Am. 2011, 129, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Fournet, M.; Szabo, A. Vocal repertoire of southeast Alaska humpback whales (Megaptera novaeangliae). J. Acoust. Soc. Am. 2013, 134, 3988. [Google Scholar] [CrossRef]
- Pitman, R.L.; Deecke, V.B.; Gabriele, C.M.; Srinivasan, M.; Black, N.; Denkinger, J.; Durban, J.W.; Mathews, E.A.; Matkin, D.R.; Neilson, J.L.; et al. Humpback whales interfering when mammal-eating killer whales attack other species: Mobbing behavior and interspecific altruism? Mar. Mamm. Sci. 2016. [Google Scholar] [CrossRef]
- Whitehead, H. Sperm Whales: Social Evolution in the Ocean; University of Chicago Press: Chicago, IL, USA, 2003. [Google Scholar]
- Roth, G.; Dicke, U. Evolution of the brain and intelligence. Trends Cogn. Sci. 2005, 9, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Dicke, U.; Roth, G. Neuronal Factors Determining High Intelligence. Philos. Trans. R. Soc. Lond. B 2016, 371. [Google Scholar] [CrossRef] [PubMed]
- Nettelbeck, T. Basic processes of intelligence. In The Cambridge Handbook of Intelligence; Sternberg, R.J., Kaufman, S.B., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 371–388. [Google Scholar]
- Nettelbeck, T.; Wilson, C. A cross-sequential analysis of developmental differences in speed of visual information processing. J. Exp. Child Psychol. 1985, 40. [Google Scholar] [CrossRef]
- Sheppard, L.D.; Vernon, P.A. Intelligence and speed of information-processing: A review of 50 years of research. Personal. Individ. Differ. 2008, 44, 535–551. [Google Scholar] [CrossRef]
- Howard, C.V.; Reid, M.G. Unbiased Stereology: Three-Dimensional Measurement in Microscopy; BIOS: Oxford, UK, 1998; ISBN 1-85996-071-5. [Google Scholar]
- Azevedo, F.A.C.; Carvalho, L.R.B.; Grinberg, L.T.; Farfel, J.M.; Ferretti, R.E.L.; Leite, R.E.P.; Filho, W.J.; Lent, R.; Herculano-Houzel, S. Equal Numbers of Neuronal and Nonneuronal Cells Make the Human Brain an Isometrically Scaled-up Primate Brain. J. Comp. Neurol. 2009, 513, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Pakkenberg, B.; Gundersen, H.J. Neocortical neuron number in humans: Effect of sex and age. J. Comp. Neurol. 1997, 384, 312–320. [Google Scholar] [CrossRef]
- Witelson, S.F.; Kigar, D.L.; Harvey, T. The exceptional brain of Albert Einstein. Lancet 1999, 353, 2149–2153. [Google Scholar] [CrossRef]
- Chittka, L.; Niven, J. Are bigger brains better? Curr. Biol. 2009, 19, R995–R1008. [Google Scholar] [CrossRef] [PubMed]
- Irving, L.; Scholander, P.F.; Grinnell, S.W. The respiration of the porpoise, Tursiops truncatus. J. Cell. Comp. Physiol. 1941, 17, 145–168. [Google Scholar] [CrossRef]
- Hampton, I.F.G.; Whittow, G.C.; Szekercezes, J.; Rutherford, S. Heat transfer and body temperature in the Atlantic bottlenosed dolphin, Tursiops truncatus. Int. J. Biometeorol. 1971, 15, 247–253. [Google Scholar] [CrossRef] [PubMed]
- De Graaf, A.S. Anatomical Aspects of the Cetacean Brain Stem; Von Gorcum: Assen, The Netherlands, 1967; pp. 1–169. [Google Scholar]
- Ridgway, S.H.; Bullock, T.H.; Carder, T.A.; Seeley, R.L.; Woods, D.; Galanbos, R. Auditory brainstem response in dolphins. Proc. Natl. Acad. Sci. USA 1981, 78, 1943–1947. [Google Scholar] [CrossRef]
- Ridgway, S.H. Neural time and movement time in choice of whistle or pulse burst responses to different auditory stimuli by dolphins. J. Acoust. Soc. Am. 2011, 129, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.H.; Shultz, J.R.; Burish, M.J.; Harrison, K.H.; Hof, P.R.; Towns, L.C.; Wagers, M.W.; Wyatt, K.D. Functional trade-offs in white matter axonal scaling. J. Neurosci. 2008, 28, 4047–4056. [Google Scholar] [CrossRef] [PubMed]
- Gusnard, D.A.; Raichle, M.E. Searching for a Baseline: Functional Imaging and the Resting Human Bran. Nat. Rev. Neurosci. 2001, 2, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.L.; Cox, M.M. Lehninger Principles of Biochemistry, 6th ed.; Macmillan Learning Publishers: London, UK, 2012. [Google Scholar]
- Belanger, M.; Allaman, I.; Magistretti, P. Brain Energy Metabolism: Focus on Astrocyte-Neuron Metabolism Cooperation. Cell 2011, 14. [Google Scholar] [CrossRef] [PubMed]
- Pellerin, L.; Magistretti, P.J. Food for Thought: Challenging the Dogmas. J. Cereb. Blood Flow Metab. 2003, 23, 1282–1286. [Google Scholar] [CrossRef] [PubMed]
- Pellerin, L.; Magistretti, P.J. Neuroenergetics: Calling up astrocytes to satisfy hungry neurons. Neuroscientist 2004, 10, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Pellerin, L.; Bouzier-Store, A.K.; Aubert, A.; Serres, S.; Merle, M.; Costalat, R.; Magistretti, P.J. Activity-dependent regulation of energy metabolism by astrocytes: An update. Glia 2007, 55, 1251–1262. [Google Scholar]
- Marino, L.; Sol, D.; Toren, K.; Lefebvre, L. Does Diving Limit rain Size in Cetaceans? Mar. Mamm. Sci. 2006, 22, 413–425. [Google Scholar] [CrossRef]
- Ridgway, S.H.; Houser, D.; Finneran, J.J.; Carder, D.A.; Keogh, M.; Van Bonn, W.; Smith, C.R.; Scadeng, M.; Mattrey, R.; Hoh, C. Functional Imaging of Dolphin Brain Metabolism and Blood Flow. J. Exp. Biol. 2014, 209, 2902–2910. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.M.; Friedl, W.A.; Haun, J.E. The physiology of bottlenose dolphins (Tursiops truncatus): Heart rate, metabolic rate and plasma kactate concentrations during exercise. J. Exp. Biol. 1993, 179, 31–46. [Google Scholar] [PubMed]
- Wyss, M.T.; Jolivet, R.; Buck, A.; Magistretti, P.J.; Weber, B. In Vivo Evidence for Lactate as a Neuronal Energy Source. J. Neurosci. 2011, 31, 7477–7485. [Google Scholar] [CrossRef] [PubMed]
- Herculano-Houzel, S.; Lent, R. Isotropic fractionator: A simple, rapid method for the quantification of total cell and neuron numbers in the brain. J. Neurosci. 2005, 25, 2518–2521. [Google Scholar] [CrossRef] [PubMed]
- Gundersen, H.J.; Jensen, E.B. The Efficiency of systematic sampling in stereology and its prediction. J. Microsc. 1987, 147, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Gundersen, H.J.; Jensen, E.B.; Kiêu, K.; Nielsen, J. The Efficiency of systematic sampling in stereology reconsidered. J. Microsc. 1999, 193, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Wallöe, S.; Eriksen, N.; Dabelsteen, T.; Pakkenberg, B. A Neurological comparative study of the harp seal (Pagophylus groenlandicus) and harbour porpoise (Phocoena phocoena). Anat. Rec. 2010, 293, 2129–2135. [Google Scholar] [CrossRef] [PubMed]
- Kazu, R.S.; Maldonado, J.; Mota, B.; Manger, P.R.; Herculano-Houzel, S. Cellular scaling rules for the brain of Artiodactyla include a highly folded cortex with few neurons. Front. Neuroanat. 2014, 8, 128. [Google Scholar] [CrossRef] [PubMed]
- Herculano-Houzel, S. The Human Advantage: A New Understanding of How Our Brain Became Remarkable; MIT Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Venn-Watson, S.; Carlin, K.; Ridgway, S. Dolphins as animal models for type 2 diabetes: Sustained, post-prandial hyperglycemia and hyperinsulinemia. Gen. Comp. Endocrinol. 2011, 170, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Mirceta, S.; Signore, A.V.; Burns, J.M.; Cossins, A.R.; Campbell, K.L.; Berenbrink, M. Evolution of Mammalian Diving Capacity Traced by Myoglobin Net Surface Charge. Science 2013, 340, 1234192. [Google Scholar] [CrossRef] [PubMed]
- Marino, L.; Connor, R.C.; Fordyce, E.R.; Herman, L.M.; Hof, P.R.; Lefebvre, L.; Lusseau, D.; McCowan, B.; Nimchinsky, E.A.; Pack, A.A.; et al. Cetaceans Have Complex Brains for Complex Cognition? PLoS Biol. 2007, 22, 413–425. [Google Scholar]
- Herrmann, E.; Call, J.; Hernandez-LLoreda, M.V.; Hare, B.; Tomasello, M. Humans Have Evolved Specialized Skills of Social Cognition: The Cultural Intelligence Hypothesis. Science 2007, 317, 1360–1366. [Google Scholar] [CrossRef] [PubMed]
- van Schaik, C.P.; Burkart, J.M. Social Learning and evolution: The cultural intelligence hypothesis. Philos. Trans. R. Soc. Lond. Biol. Sci. 2011, 366, 1008–1026. [Google Scholar] [CrossRef] [PubMed]
- Zentall, T.R. Animal memory: The role of instructions. Learn. Motiv. 1997, 28, 248–267. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primates and the African Elephant | Orangutan | Gorilla | Chimpanzee | Human | Elephant |
| Cortical Gyrification Index | 2.29 | 2.07 | 2.19 | 2.57 | 3.89 |
| Cortical Surface () | 530 | 2275 | 6300 | ||
| Cortical Glia–neuron ratio | 0.98 | 1.21 | 1.2 | 1.68–1.78 | 4.7 |
| Cortical White matter-Gray matter ratio | 0.488 | 0.618 | 0.617 | 0.71 | 0.78 |
| Cortical Neuron Number (millions) | 8900 | 9100 | 9000 | 16,300 | 5600 |
| Brain weight (g) | 370 | 509.2 | 420 | 1500 | 4783 |
| Basal Metabolic Rate (kcal/day) | 569.1 | 581.9 | 1400 | 65,000 | |
| Brain Metabolic Rate (kcal/day) | 63 | 77 | 280 | 1200 | |
| Cetaceans | Harbor Porpoise | Bottlenose Dolphin | Pilot Whale | Killer Whale | Minke Whale |
| Cortical Gyrification Index | 5.63 | 5.70 | |||
| Cortical Surface () | 3745 | 5800 | 13,629 | 5900 | |
| Cortical Glia–neuron ratio | 2.34 | 2–3.1 | 3.41 | 7.67 | |
| Cortical White matter-Gray matter ratio | 0.66 | 0.76 | 0.74 | ||
| Cortical Neuron Number (millions) | 14,900 | 37,200 | 12,800 | ||
| Brain weight (g) | 540 | 1550 | 2679 | 6622 | 2228 |
| Basal Metabolic Rate (kcal/day) | 2389 | 9000–10,000 | 40,000 | 90,000–110,000 | |
| Brain Metabolic Rate (kcal/day) | 48 | 429–477 | 800 | 1800–2200 |
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chinea, A. Is Cetacean Intelligence Special? New Perspectives on the Debate. Entropy 2017, 19, 543. https://doi.org/10.3390/e19100543
Chinea A. Is Cetacean Intelligence Special? New Perspectives on the Debate. Entropy. 2017; 19(10):543. https://doi.org/10.3390/e19100543
Chicago/Turabian StyleChinea, Alejandro. 2017. "Is Cetacean Intelligence Special? New Perspectives on the Debate" Entropy 19, no. 10: 543. https://doi.org/10.3390/e19100543
APA StyleChinea, A. (2017). Is Cetacean Intelligence Special? New Perspectives on the Debate. Entropy, 19(10), 543. https://doi.org/10.3390/e19100543