Ribozyme Activity of RNA Nonenzymatically Polymerized from 3′,5′-Cyclic GMP

Abstract
:
1. Introduction
2. Experimental
2.1. Materials
2.1.1. Oligonucleotides
2.1.2. Enzymes
2.1.3. Ribonucleases
2.2. Methods
2.2.1. Polymerization of 3′,5′-cyclic GMP
2.2.2. Terminal Labelling of the Material Polymerized from Cyclic Nucleotides
2.2.3. The Ligation Following Intermolecular Cleavage (LIC)
2.3. MALDI-ToF Mass Spectrometry
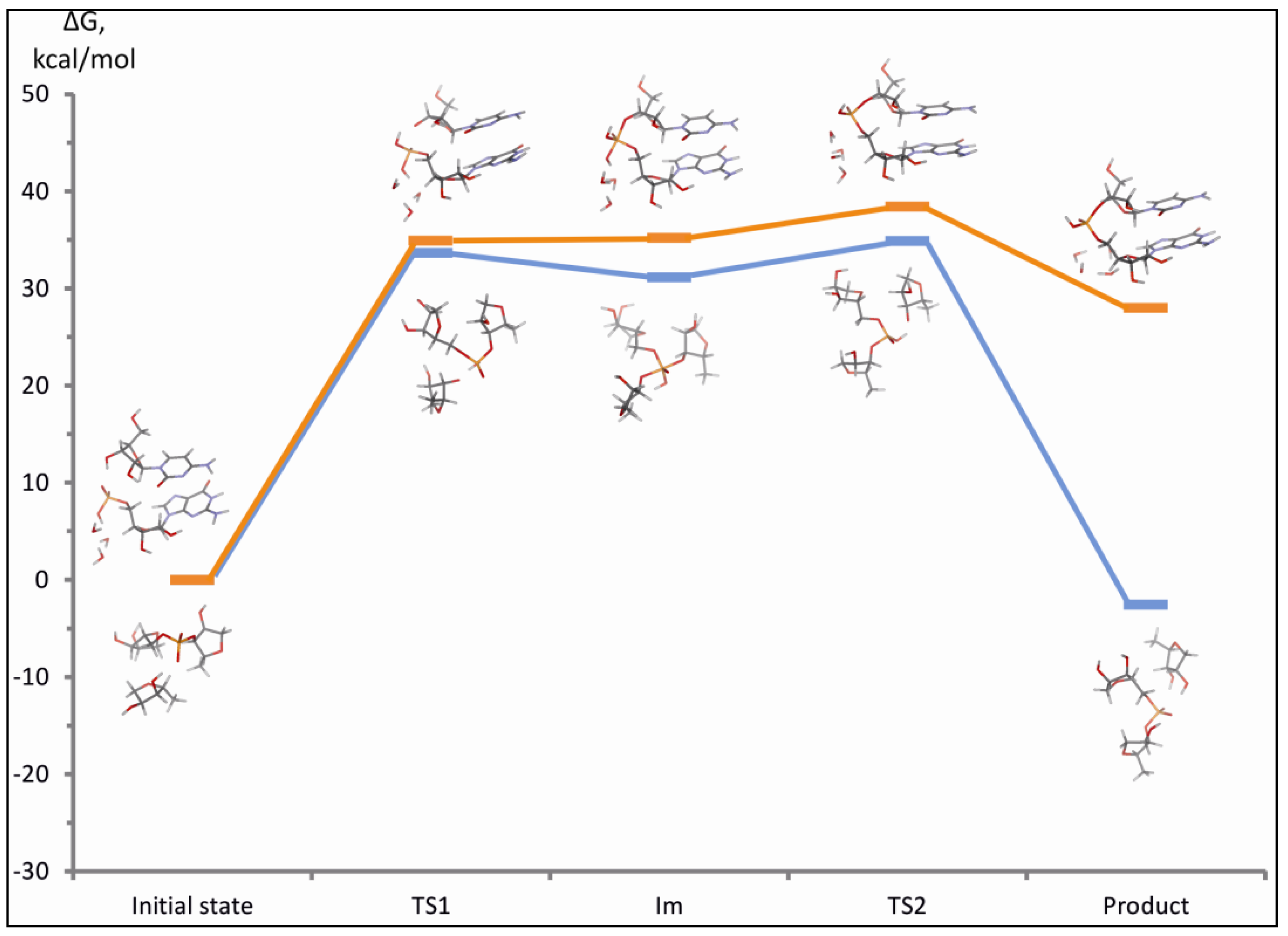
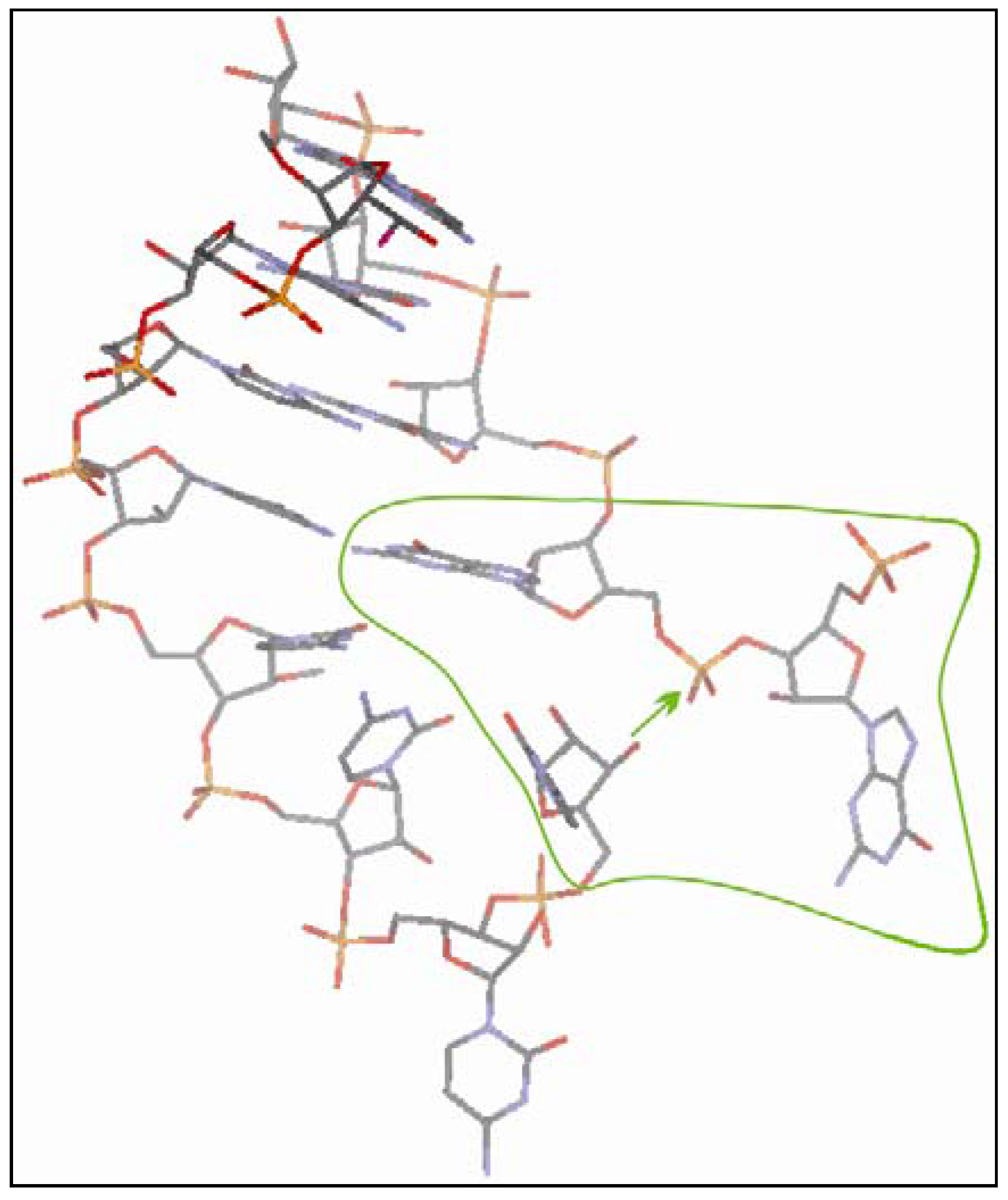
2.4. Computational Details
3. Results and Discussion
3.1. The Experimental Plan
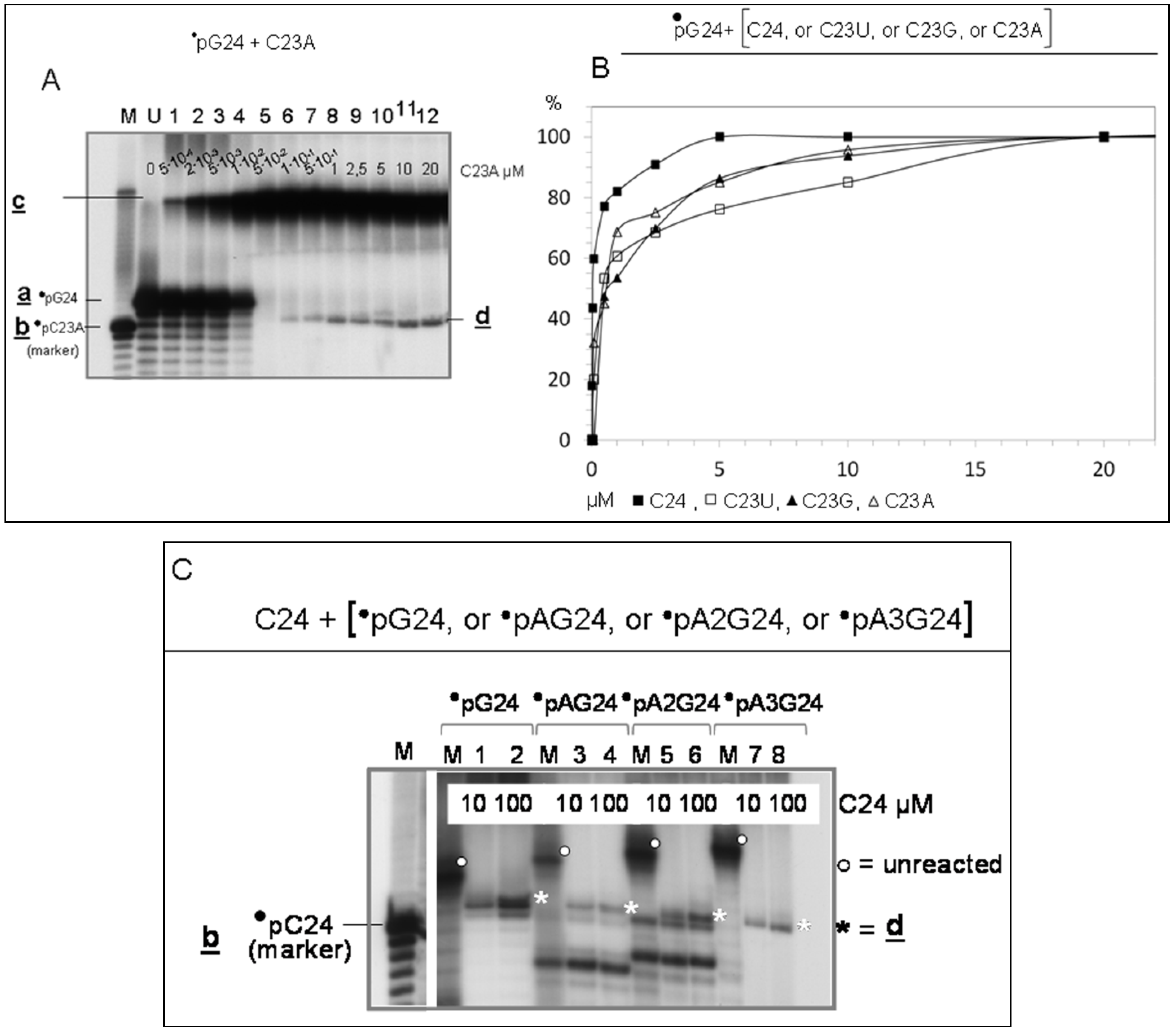
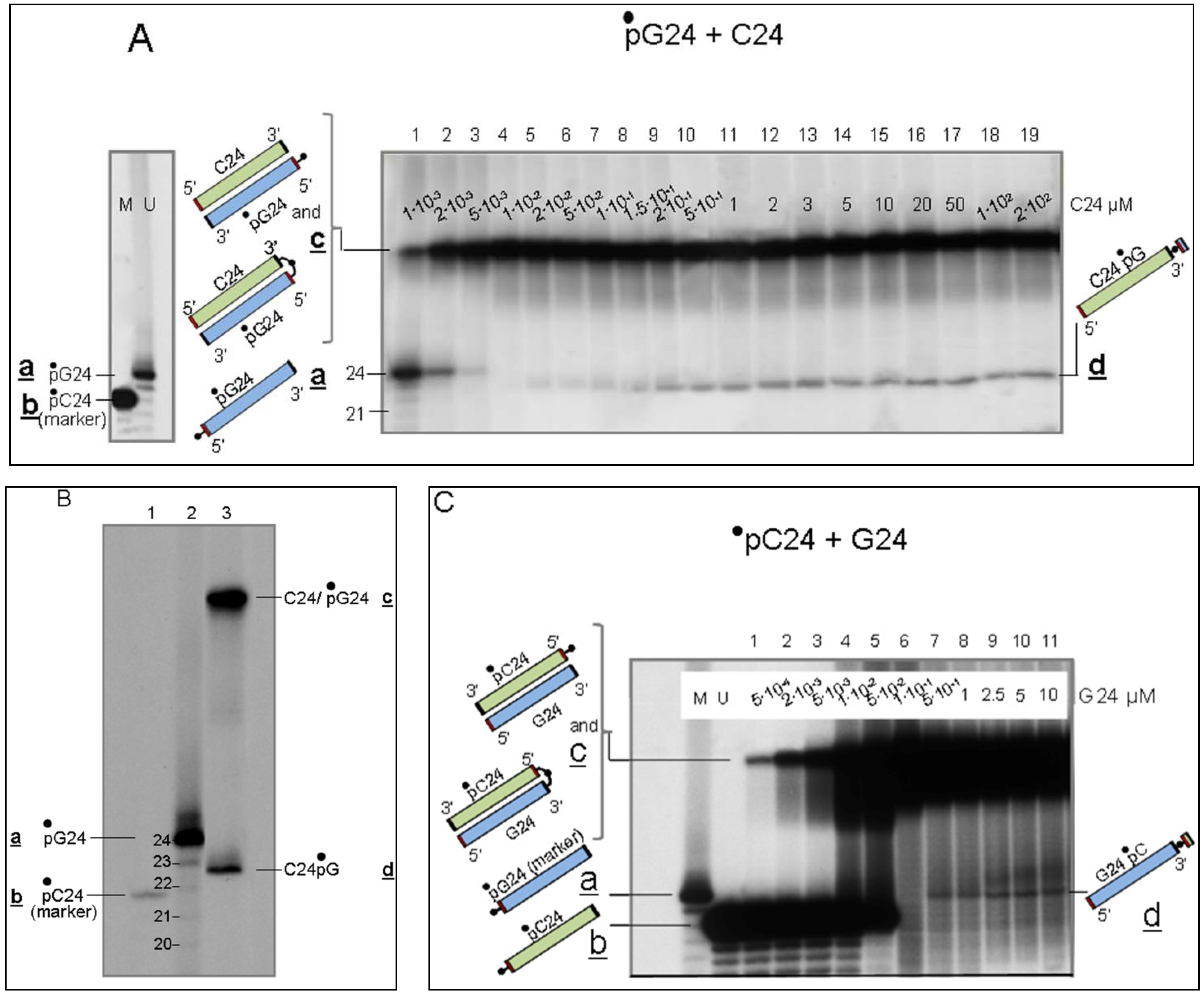
3.2. Detection of the LIC Reaction
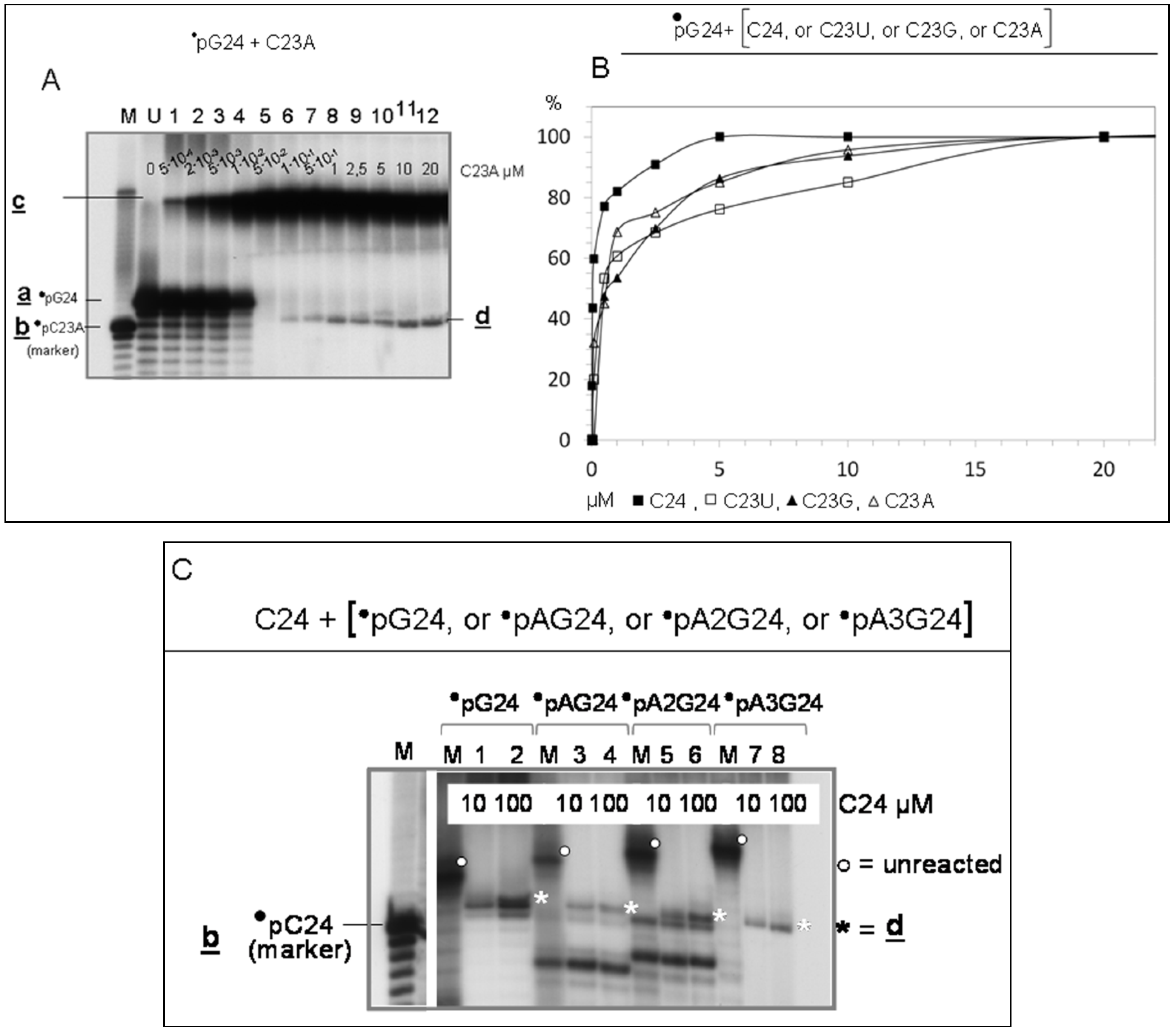
3.2.1. Interaction of Sequence-Complementary Oligonucleotides (G24 + C24) Results in the Production of a New Molecular Species
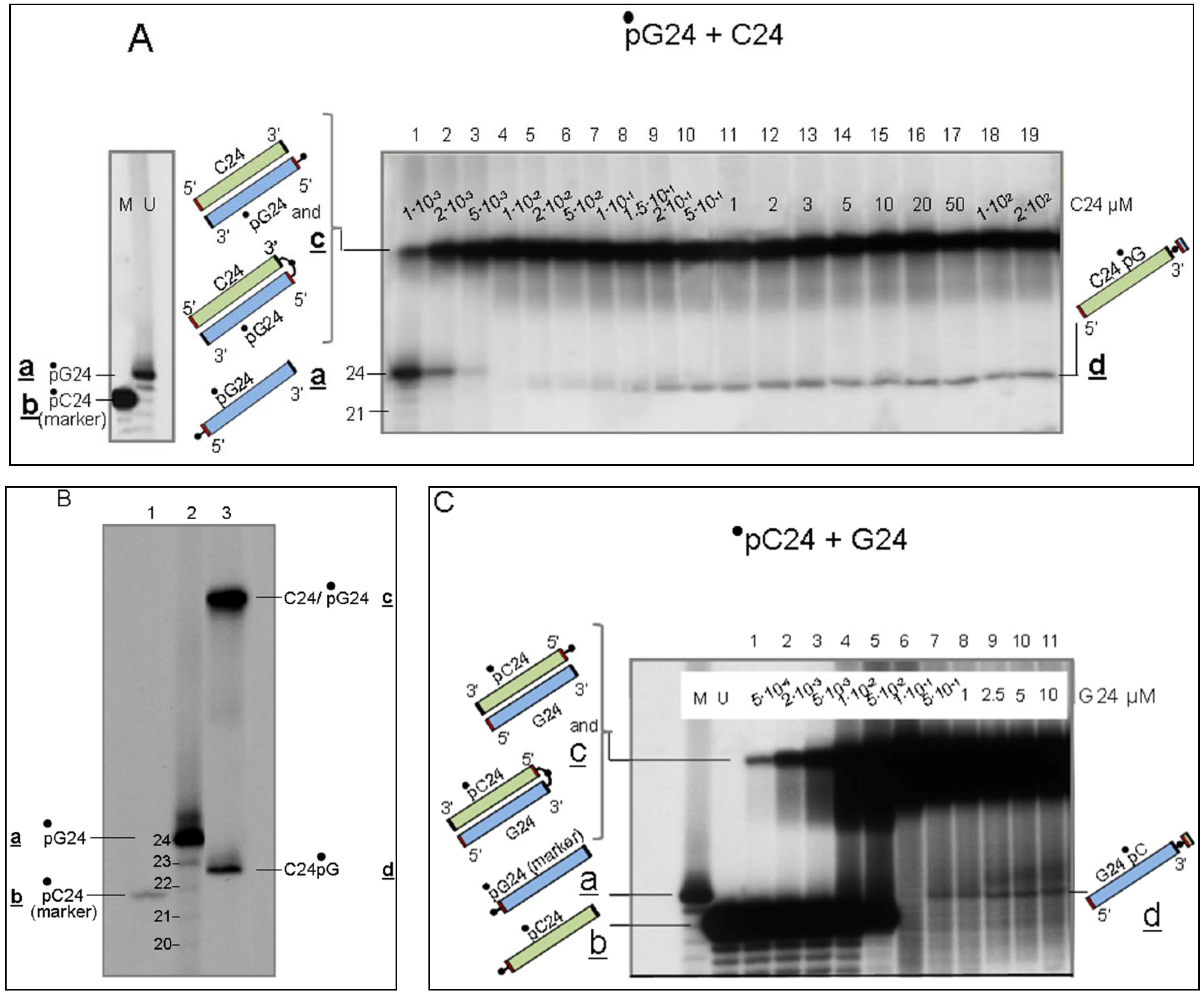
3.2.2. Characterization of Molecules d, Product of Reaction LIC
- (i)
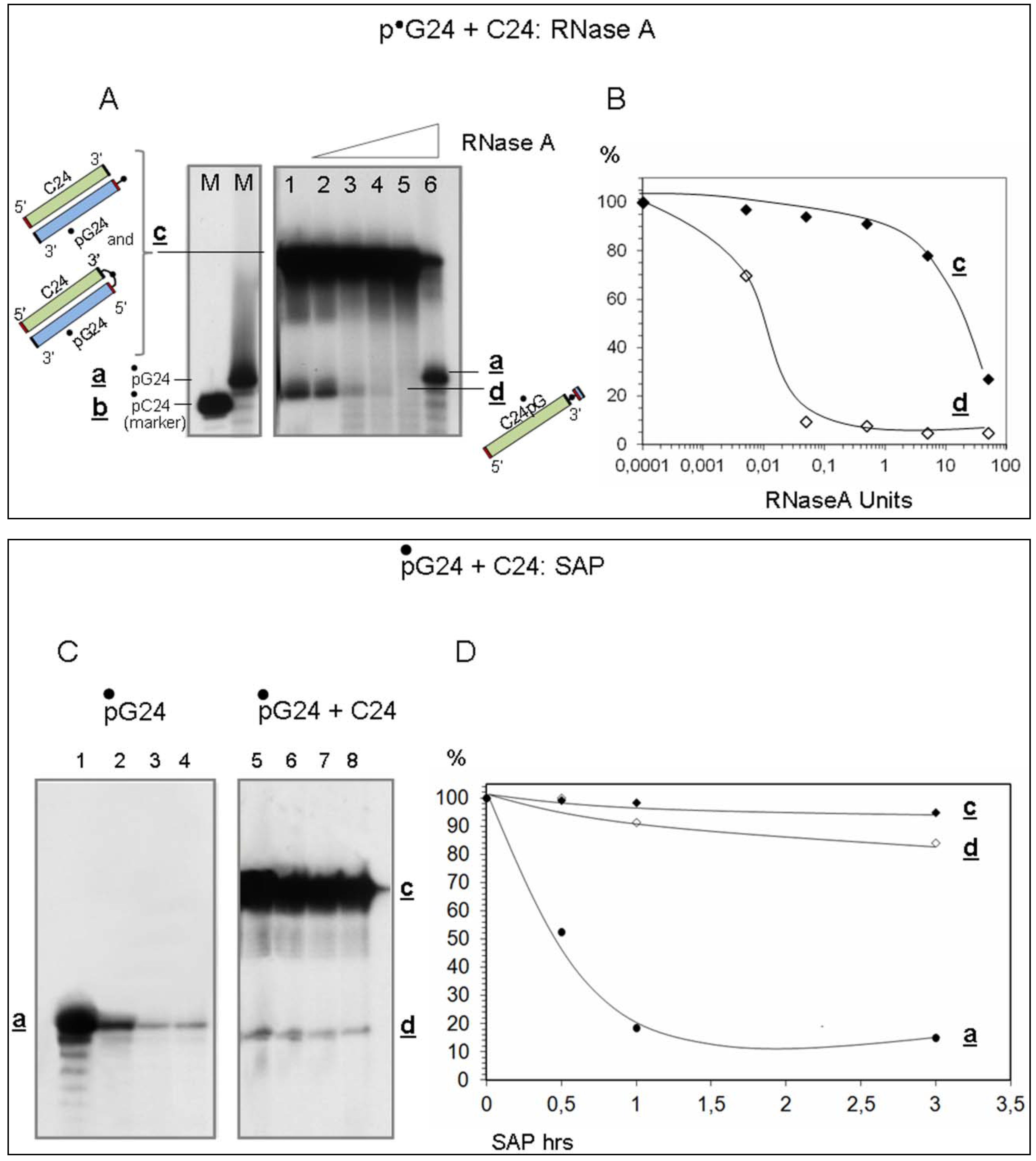
- Evidence that d is almost entirely made of cytosines is provided by specific RNaseA analyses (Figure 2). Given that the C24 oligomer is introduced in the assay as unphosphorylated unlabeled molecule, the acquired phosphate is necessarily derived from the 5′ 32P labelled G24 oligomer (•pG24).
- (ii)
- The 3′ position of the acquired phosphate group is established by 5′ specific exonuclease (Shrimp Alkaline Phosphatase) treatment (Figure 2C,D). Validation of this result is provided by an hybridization competition assay (Supplementary Item #3).
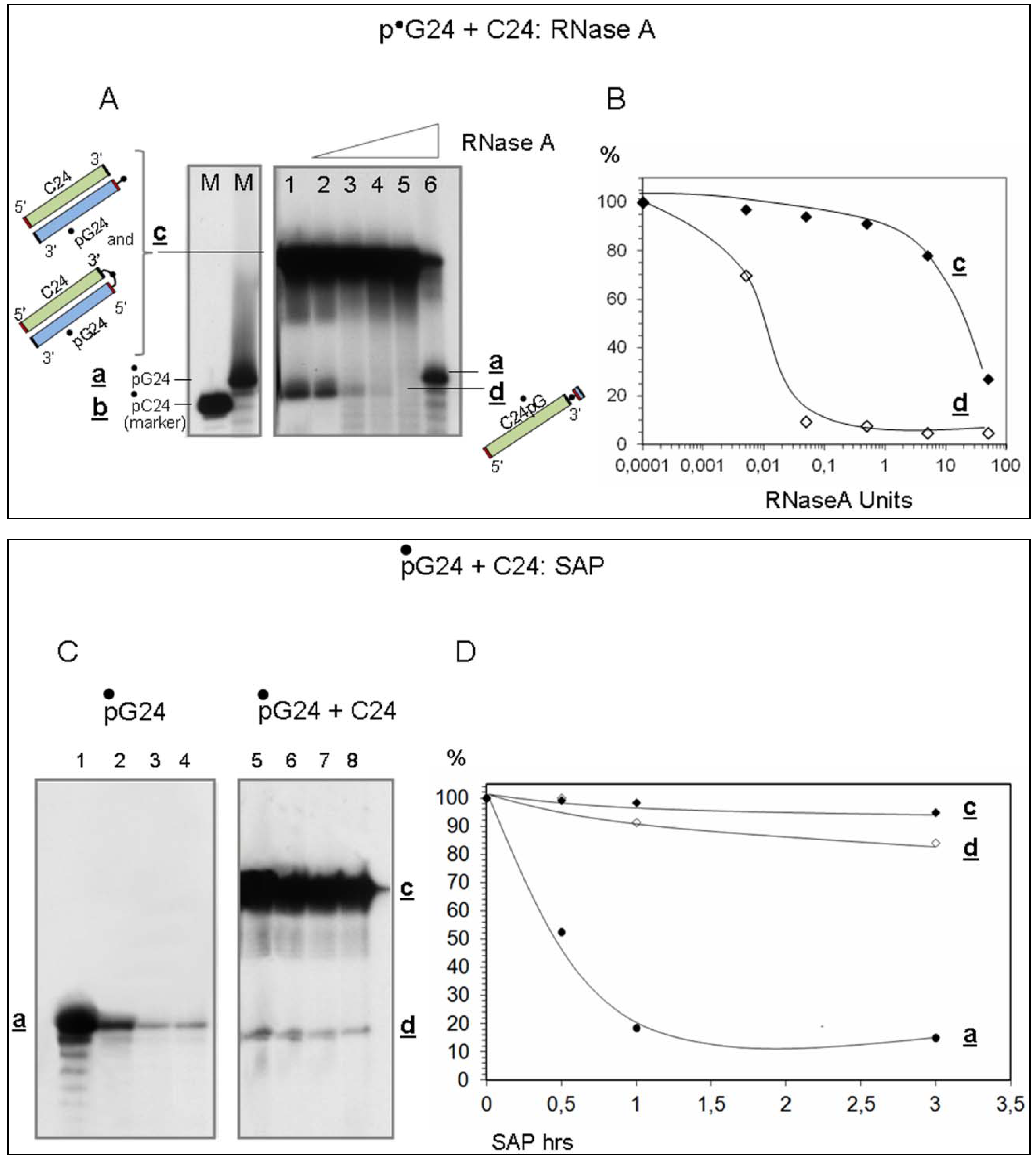
3.2.3. Determination of the Position of the 32P-labelled Phosphate Group
Ribonuclease A
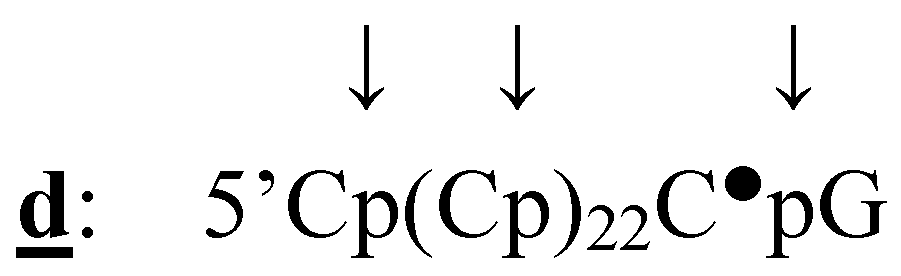
Shrimp Alkaline Phosphatase (SAP)
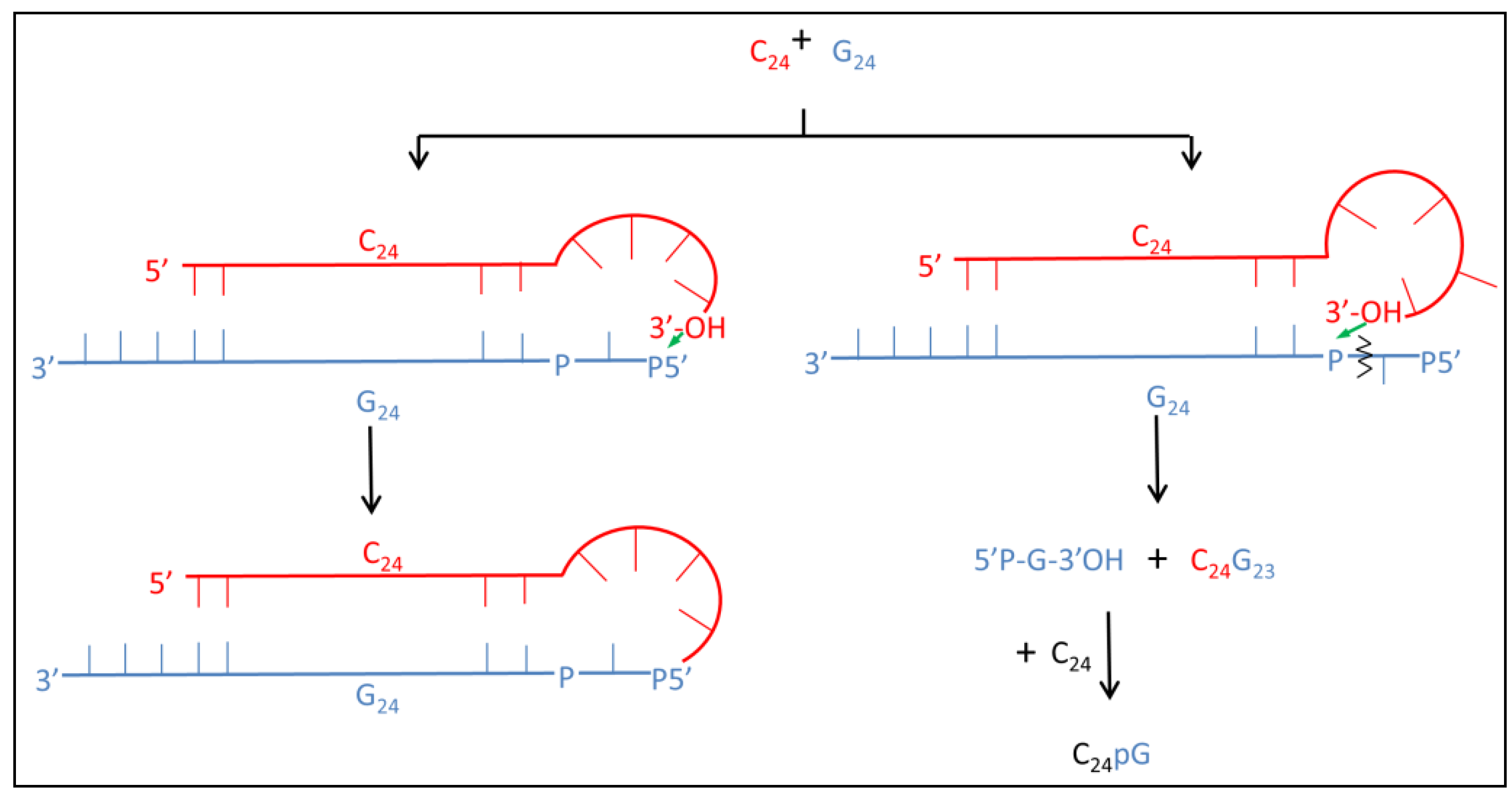
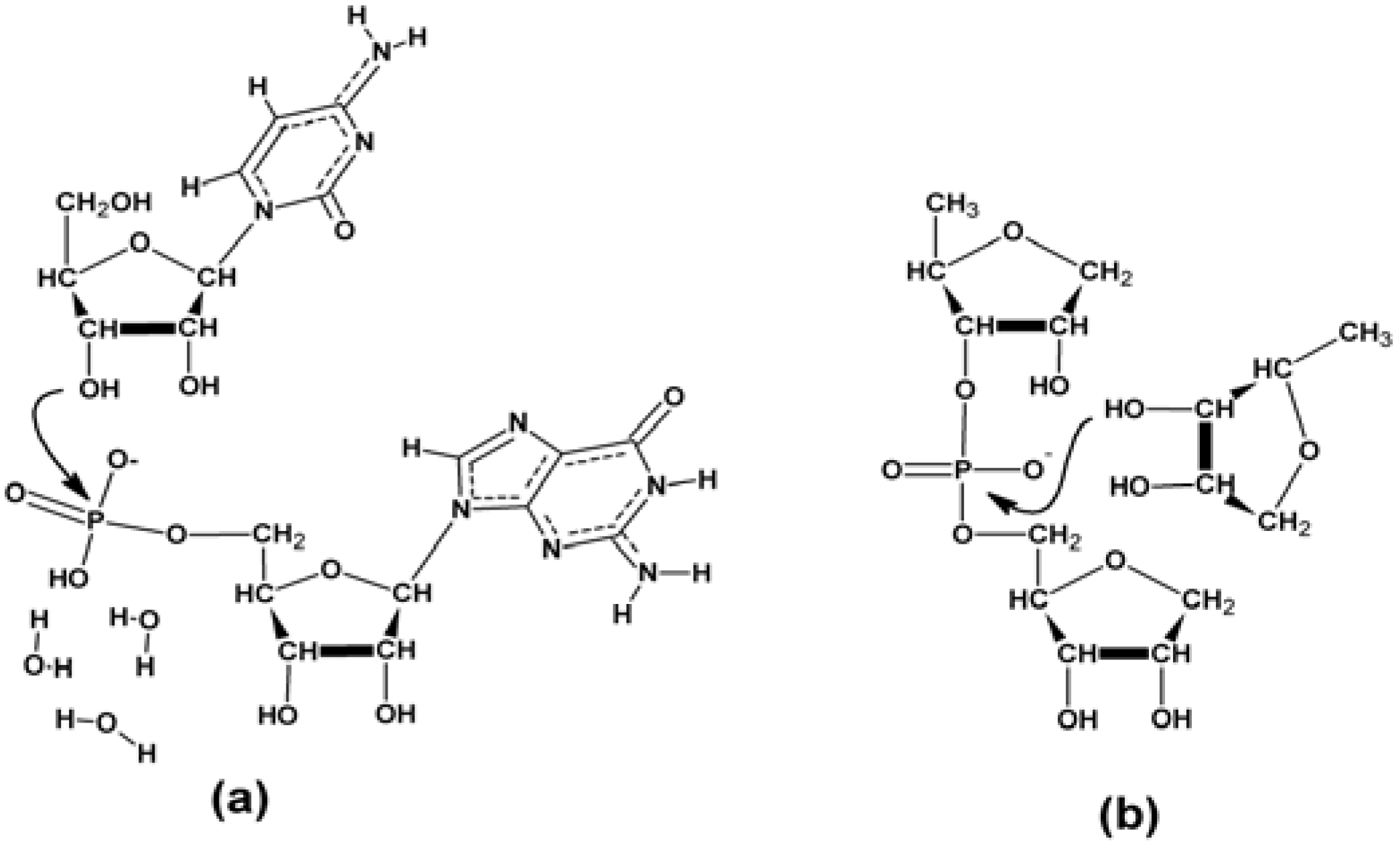
3.2.4. The Ligation Following Intermolecular Cleavage (LIC) Mechanism
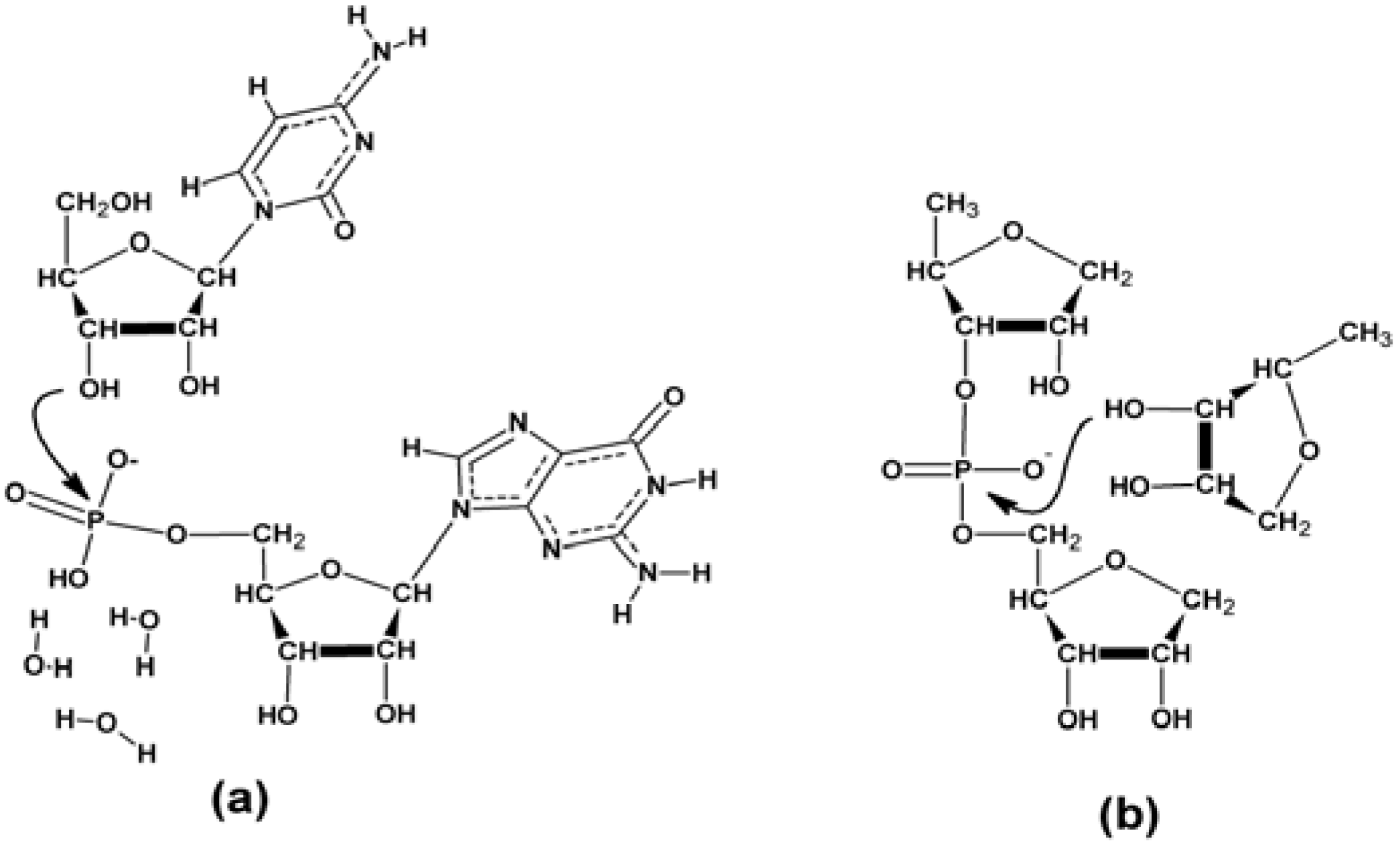
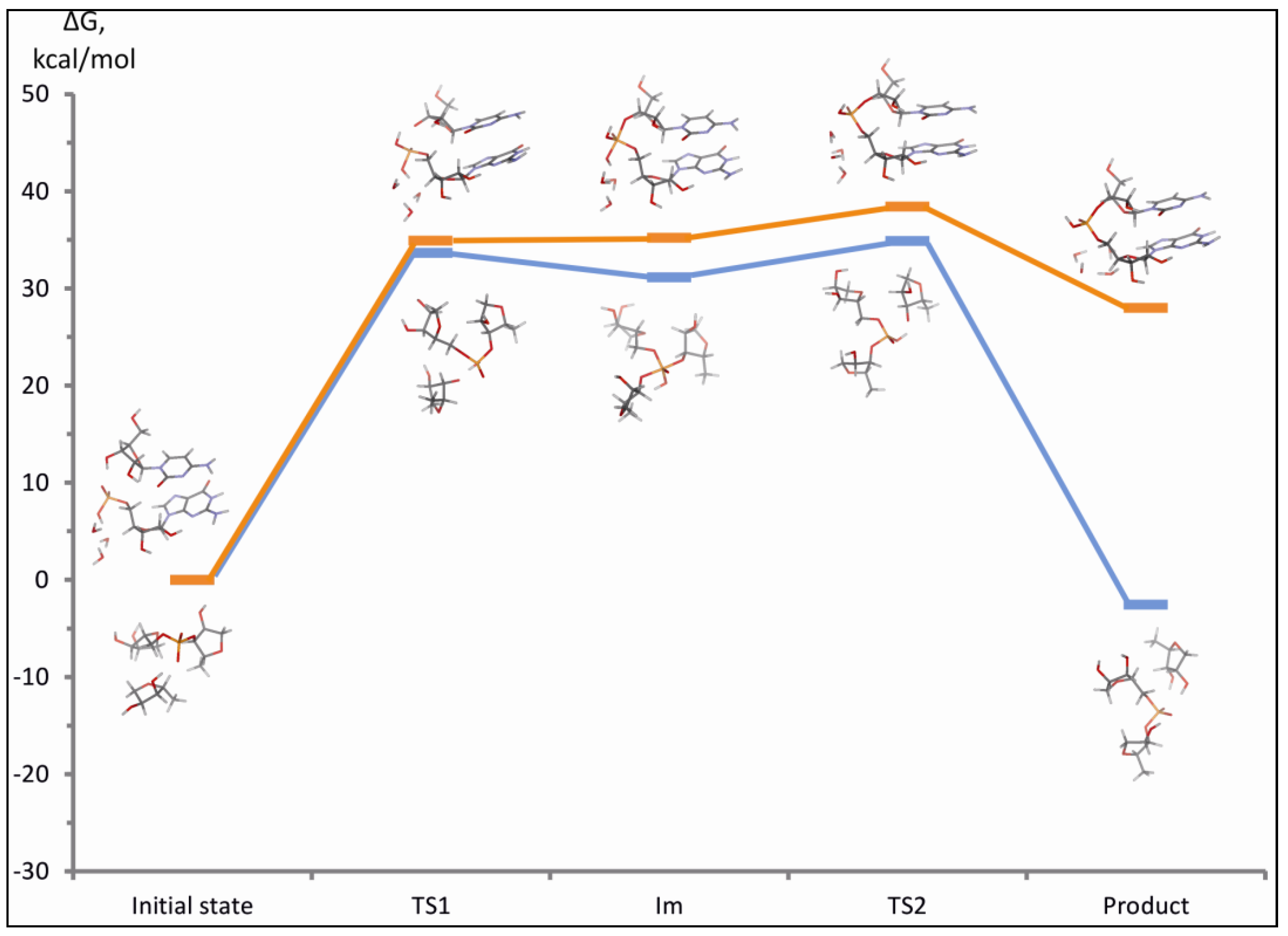
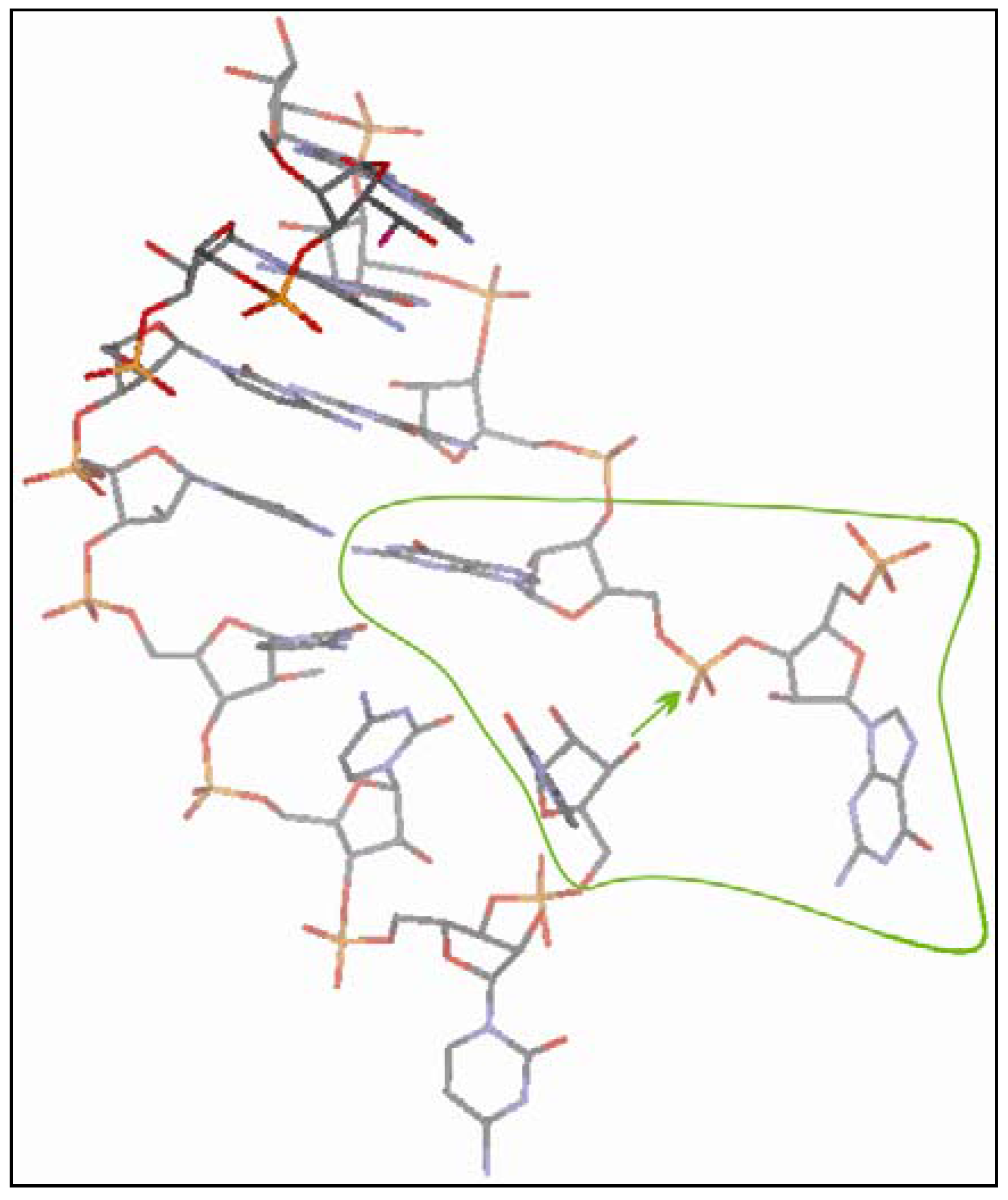
3.2.5. Sequence Requirements

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
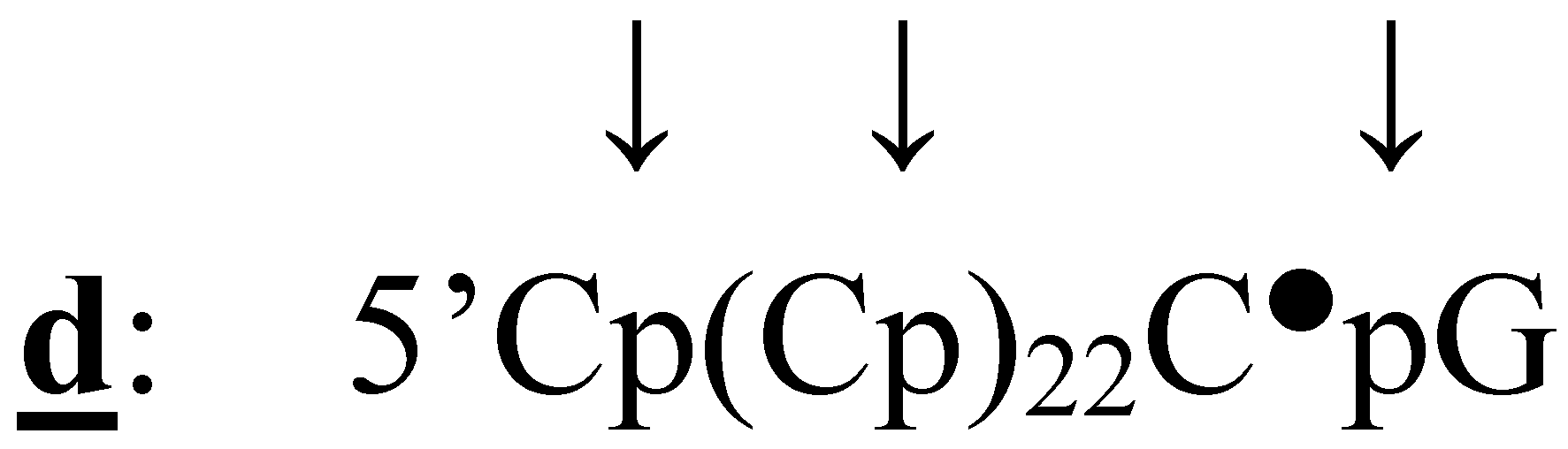{kind=link}
| Construct (A + D) | [A]/[D] | Yield (%) |
|---|---|---|
| C24 + •pG24 | 38.5 | 10 |
| C23U + •pG24 | 154 | 10 |
| C23G + •pG24 | 115 | 10 |
| C23A + •pG24 | 115 | 10 |
| A12C12 + •pG24 | 23 | 5 |
| A6C18 + •pG24 | 23 | 5 |
| C12A12 + •pG24 | 23 | 5 |
| A24 + •pG24 | 0 | 0 |
| U24 + •pG24 | 0 | 0 |
| A12U12 + •pG24 | 0 | 0 |
| G24 + •pC24 | 3.8 | 1 |
| G23U + •pC24 | 19.2 | 2 |
| G23A + •pC24 | 7.7 | 2 |
| G23C + •pC24 | 7.7 | 0.5 |
| C24 + •pG9 | 7.7 | 100 |
| C9 + •pG9 | 7.7 | 0 |
| C12A12 + •pG9 | 7.7 | 100 |
| A12C12 + •pG9 | 7.7 | 100 |
| C24 + •p NEO G | 4.7 | 15 |
3.2.6. Complexity
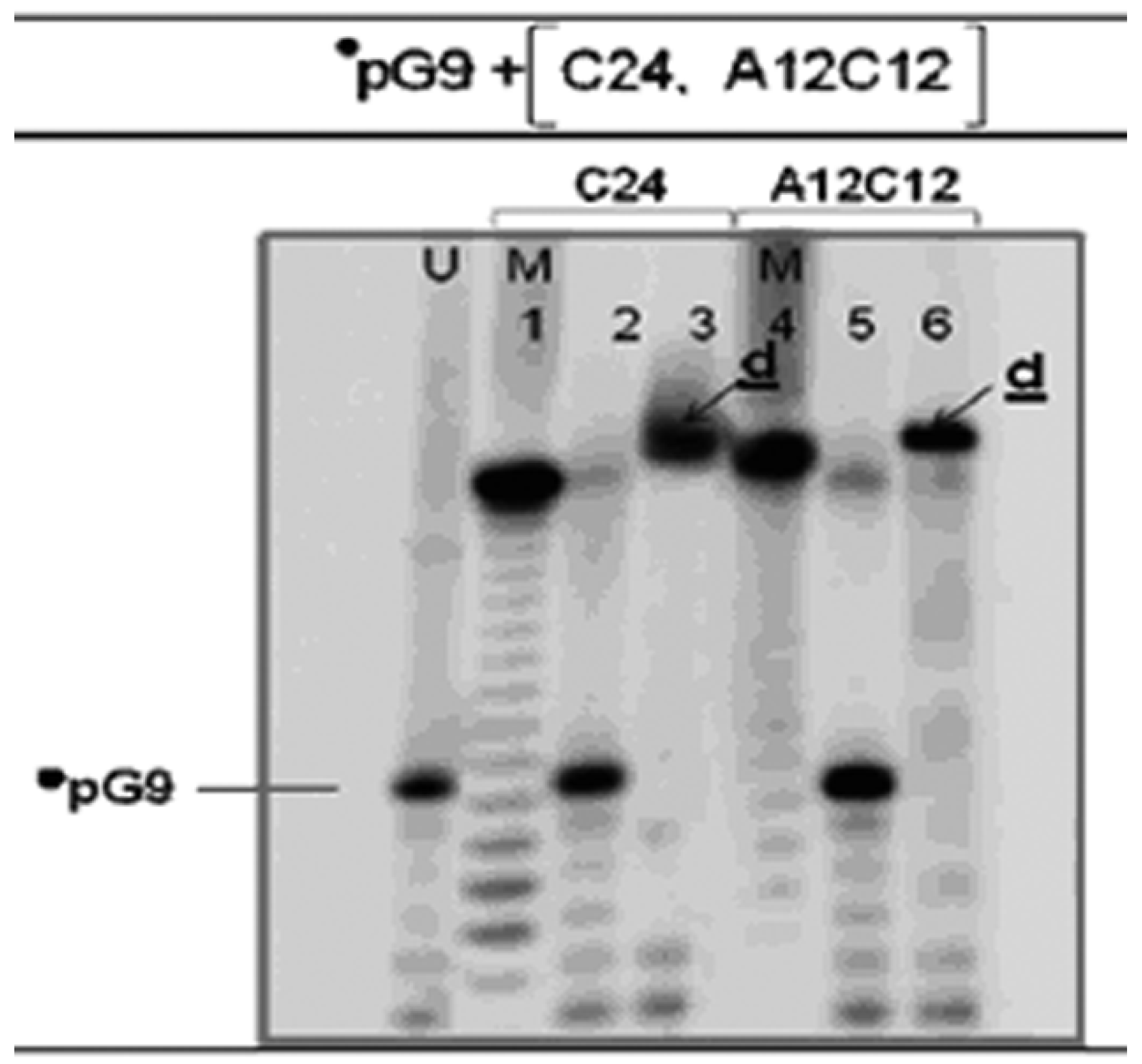
3.3. The •pG9 Oligonucleotides also Actively Perform the LIC Reaction
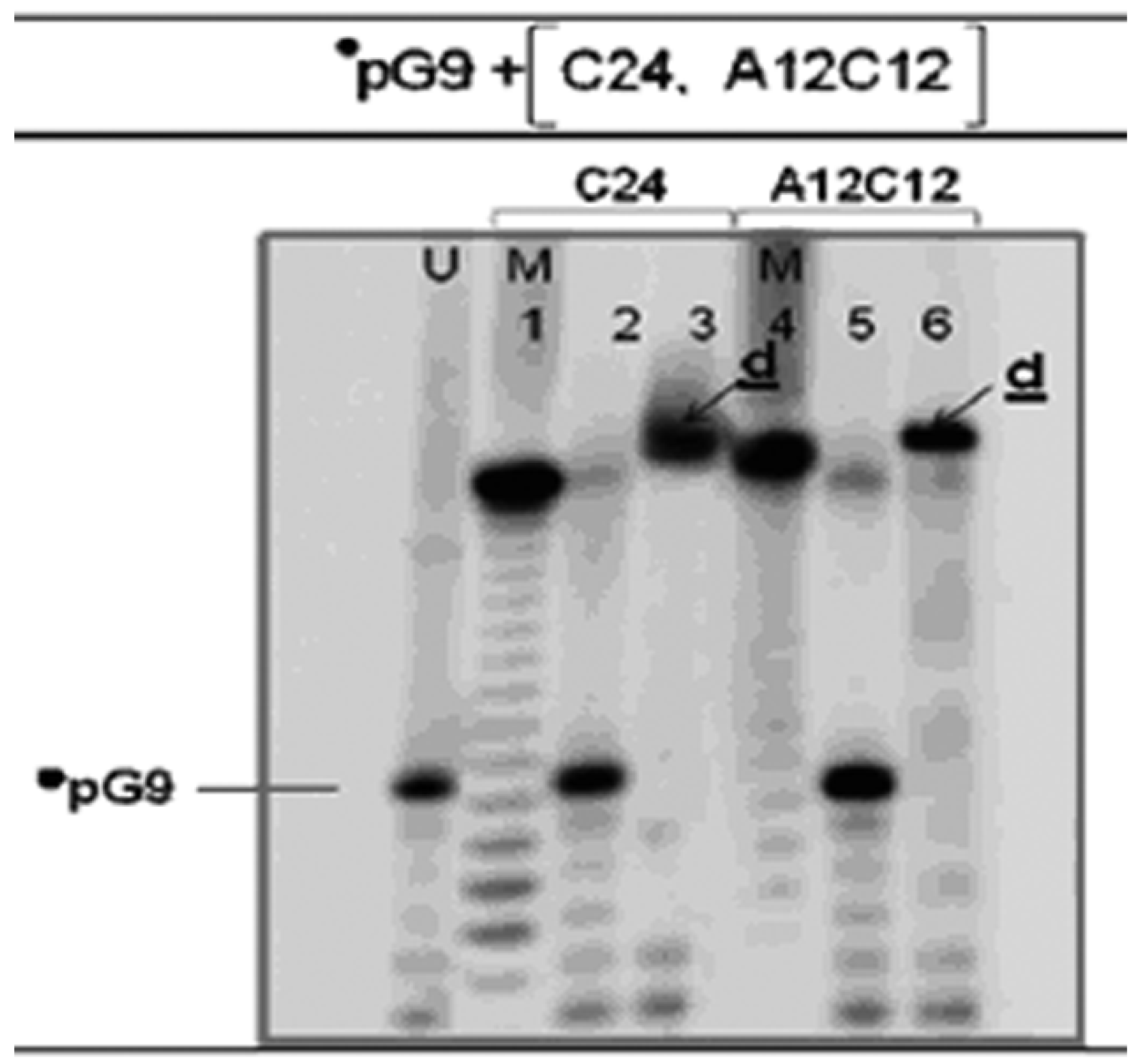
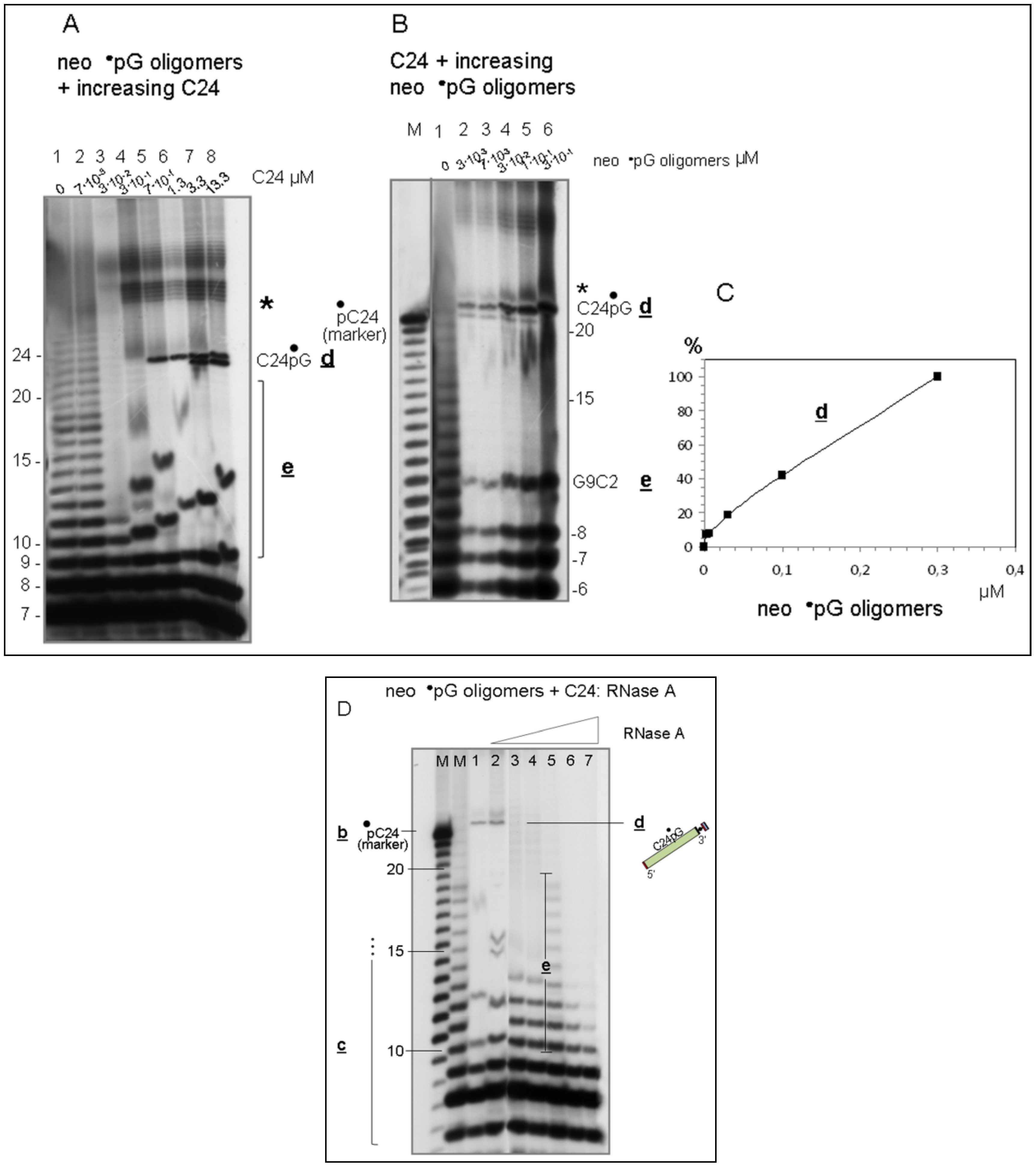
3.4. Spontaneously Polymerized Oligos Perform the LIC Reaction
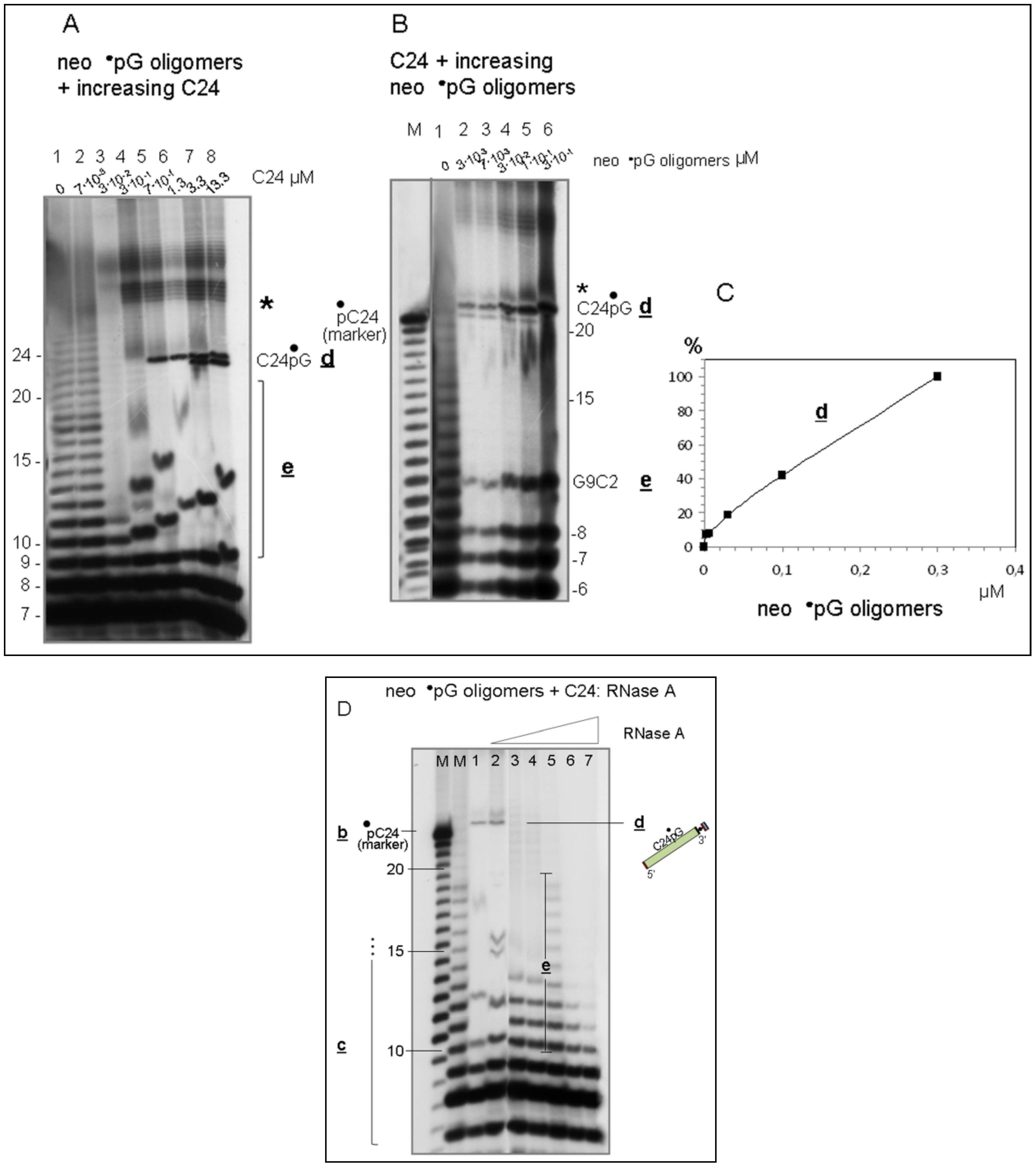
3.5. Discussion and Evolutionary Implications
3.6. Future Plans
Acknowledgments
Conflicts of Interest
References
- Guerrier-Takada, C.; Gardiner, K.; Marsh, T.; Pace, N.; Altman, S. The RNA moiety of ribonuclease P is the catalytic subunit of the enzyme. Cell 1983, 35, 849–857. [Google Scholar] [CrossRef]
- Zaug, A.J.; Grabowski, P.J.; Cech, T.R. Autocatalytic cyclization of an excised intervening sequence RNA is a cleavage-ligation reaction. Nature 1983, 301, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Cech, T.R. The chemistry of self-splicing RNA and RNA enzymes. Science 1987, 236, 1532–1539. [Google Scholar] [CrossRef] [PubMed]
- Zaug, A.J.; Cech, T.R. The intervening sequence RNA of Tetrahymena is an enzyme. Science 1996, 231, 470–475. [Google Scholar] [CrossRef]
- Doherty, E.A.; Doudna, J.A. Ribozyme structures and mechanisms. Annu. Rev. Biochem. 2000, 69, 597–615. [Google Scholar] [CrossRef] [PubMed]
- De Rose, V.J. Two decades of RNA catalysis. Chem. Biol. 2002, 9, 961–969. [Google Scholar]
- Chetverin, A.B.; Chetverina, H.V.; Demidenko, A.A.; Ugarov, V.I. Nonhomologous RNA recombination in a cell-free system: Evidence for a transesterification mechanism guided by secondary structure. Cell. 1997, 88, 503–513. [Google Scholar] [CrossRef]
- Chetverin, A.B. The puzzle of RNA recombination. FEBS Lett. 1999, 460, 1–5. [Google Scholar] [CrossRef]
- Chetverina, H.V.; Demidenko, A.A.; Ugarov, V.I.; Chetverin, A.B. Spontaneous rearrangements in RNA sequences. FEBS Lett. 1999, 450, 89–94. [Google Scholar] [CrossRef]
- Figlerowicz, M.; Bibiłło, A. RNA motifs mediating in vivo site-specific nonhomologous recombination in (+) RNA virus enforce in vitro nonhomologous crossovers with HIV-1 reverse transcriptase. RNA 2000, 6, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Chetverin, A.B. Replicable and recombinogenic RNAs. FEBS Lett. 2004, 567, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Chetverin, A.B.; Kopein, D.S.; Chetverina, H.V.; Demidenko, A.A.; Ugarov, V.I. Viral RNA-directed RNA polymerases use diverse mechanisms to promote recombination between RNA molecules. J. Biol. Chem. 2005, 280, 8748–8755. [Google Scholar] [CrossRef] [PubMed]
- Striggles, J.C.; Martin, M.B.; Schmidt, F.J. Frequency of RNA-RNA interaction in a model of the RNA World. RNA 2006, 12, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Lutay, A.V.; Zenkova, M.A.; Vlassov, V.V. Nonenzymatic recombination of RNA: Possible mechanism for the formation of novel sequences. Chem. Biodivers. 2007, 4, 762–767. [Google Scholar] [CrossRef] [PubMed]
- Obermayer, B.; Krammer, H.; Braun, D.; Gerland, U. Emergence of information transmission in a prebiotic RNA reactor. Phys. Rev. Lett. 2011, 107, 018101. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, G.; Pino, S.; Ciciriello, F.; Di Mauro, E. Generation of long RNA chains in water. J. Biol. Chem. 2009, 284, 33206–33216. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, G.; Saladino, R.; Botta, G.; Giorgi, A.; Scipioni, A.; Pino, S.; Di Mauro, E. Generation of RNA molecules by base catalyzed click-like reaction. ChemBioChem 2012, 13, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Crestini, C.; Neri, V.; Ciciriello, F.; Costanzo, G.; Di Mauro, E. Origin of informational polymers: The concurrent roles of formamide and phosphates. ChemBioChem 2006, 7, 1707–1714. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, G.; Saladino, R.; Crestini, C.; Ciciriello, F.; Di Mauro, E. Formamide as main building block in the origin of life. BMC Evol. Biol. 2007, 7. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, G.; Saladino, R.; Crestini, C.; Ciciriello, F.; Di Mauro, E. Nucleoside phosphorylation by phosphate minerals. J. Biol. Chem. 2007, 282, 16729–16735. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Botta, G.; Pino, S.; Costanzo, G.; Di Mauro, E. Genetics first or metabolism first? The formamide clue. Chem. Soc. Rev. 2012, 41, 5526–5565. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Crestini, C.; Pino, S.; Costanzo, G.; Di Mauro, E. Formamide and the origin of life. Phys. Life Rev. 2012, 9, 84–104. [Google Scholar] [CrossRef] [PubMed]
- Benner, S.A.; Kim, H.J.; Cardigan, M.A. Asphalt, water, and the prebiotic synthesis of ribose, ribonucleosides, and RNA. Accounts Chem. Res. 2012, 45, 2025–2034. [Google Scholar] [CrossRef] [PubMed]
- Deck, C.; Jauker, M.; Richert, C. Efficient enzyme-free. Copying of all four nucleobases templated by immobi-lized RNA. Nature Chem. 2011, 3, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Pino, S.; Ciciriello, F.; Costanzo, G.; Di Mauro, E. Nonenzymatic RNA ligation in water. J. Biol. Chem. 2008, 283, 36494–36503. [Google Scholar] [CrossRef] [PubMed]
- Pino, S.; Costanzo, G.; Giorgi, A.; Di Mauro, E. Sequence complementarity-driven nonenzymatic ligation of RNA. Biochemistry 2011, 50, 2994–3003. [Google Scholar] [CrossRef] [PubMed]
- Pino, S.; Biasiucci, M.; Scardamaglia, M.; Gigli, G.; Betti, M.G.; Mariani, C.; Di Mauro, E. Nonenzymatic ligation of an RNA oligonucleotide analyzed by atomic force microscopy. J. Phys. Chem. B 2011, 115, 6296–6303. [Google Scholar] [CrossRef] [PubMed]
- Levene, P.A.; Jorpes, E. The rate of hydrolysis of ribonucleotides. J. Biol. Chem. 1929, 81, 575–580. [Google Scholar]
- Saladino, R.; Crestini, C.; Ciciriello, F.; Di Mauro, E.; Costanzo, G. Origin of informational polymers: differential stability of phosphoester bonds in ribo monomers and oligomers. J. Biol. Chem. 2006, 281, 5790–5796. [Google Scholar] [CrossRef] [PubMed]
- Stephens, R.M.; Schneider, T.D. Features of spliceosome evolution and function inferred from an analysis of the information at human splice sites. J. Mol. Biol. 1992, 228, 1124–1136. [Google Scholar] [CrossRef]
- Yonath, A. Polar bears, antibiotics, and the evolving ribosome (Nobel Lectures). 2010, 49. [Google Scholar] [CrossRef]
- Turk, R.M.; Chumachenko, N.V.; Yarus, M. Multiple translational products from a five-nucleotide ribozyme. Proc. Natl. Acad. Sci. USA 2010, 107, 4585–4589. [Google Scholar] [CrossRef] [PubMed]
- Cuchillo, C.M.; Parés, X.; Guasch, A.; Barman, T.; Travers, F.; Nogués, M.V. The role of 2′,3′-cyclic phosphodiesters in the bovine pancreatic ribonuclease A catalysed cleavage of RNA: Intermediates or products? FEBS Lett. 1993, 333, 207–210. [Google Scholar] [CrossRef]
- Raines, R.T. Ribonuclease A. Chem. Rev. 1998, 98, 1045–1066. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.M.; Perdew, J.P.; Staroverov, V.N.; Scuseria, G.E. Climbing the density functional ladder: Nonempirical meta-generalized gradient approximation designed for molecules and solids. Phys. Rev. Lett. 2003, 91, 146401. [Google Scholar] [CrossRef] [PubMed]
- Grimme, S. Semiempirical GGA-type density functional constructed with a long-range dispersion correction. J. Comp. Chem. 2006, 27, 1787–1799. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, A.; Horn, H.; Ahlrichs, R. Fully optimized contracted Gaussian-basis sets for atoms Li to Kr. J. Chem. Phys. 1992, 97, 2571–2577. [Google Scholar] [CrossRef]
- Schaefer, A.; Huber, C.; Ahlrichs, R. Fully optimized contracted Gaussian-basis sets of triple zeta valence quality for atoms Li to Kr. J. Chem. Phys. 1994, 100, 5829–5835. [Google Scholar] [CrossRef]
- Barone, V.; Cossi, M. Quantum calculation of molecular energies and energy gradients in solution by a conductor solvent model. J. Phys. Chem. A 1998, 102, 1995–2001. [Google Scholar] [CrossRef]
- Cossi, M.; Rega, N.; Scalmani, G.; Barone, V. Energies, structures, and electronic properties of molecules in solution with the C-PCM solvation model. J. Comp. Chem. 2003, 24, 669–681. [Google Scholar] [CrossRef] [PubMed]
- Klamt, A.; Schüürmann, G. COSMO: A new approach to dielectric screening in solvents with explicit expressions for the screening energy and its gradient. J. Chem. Soc. Perkin Trans. 1993, 2, 799–805. [Google Scholar] [CrossRef]
- Gaussian 09; Revision, A.1; computer program for electronic structure modeling; Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G.A.; et al. Gaussian, Inc.: Wallingford, CT, USA, 2009. [Google Scholar]
- Soukup, G.A.; Breaker, R.R. Relationship between internucleotide linkage geometry and the stability of RNA. RNA 1999, 5, 1308–1325. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Breaker, R.R. Kinetics of RNA degradation by specific base catalysis of transesterification involving the 2′-hydroxyl group. J. Am. Chem. Soc. 1999, 121, 5364–5372. [Google Scholar] [CrossRef]
- Dawson, W.; Yamamoto, K.; Kawai, G. A new entropy model for RNA: Part I. A critique of the standard Jacobson-Stockmayer model applied to multiple cross links. J. Nucleic Acids Investig. 2012, 3, e3. [Google Scholar] [CrossRef]
- Dawson, W.; Yamamoto, K.; Shimizu, K.; Kawai, G. A new entropy model for RNA: Part II. Persistence-related entropic contributions to RNA secondary structure free energy calculations. J. Nucleic Acids Investig. 2013, 4, e2. [Google Scholar] [CrossRef]
- McConaughy, B.L.; Laird, C.D.; McCarthy, B.J. Nucleic acid reassociation in formamide. Biochemistry 1969, 8, 3289–3295. [Google Scholar] [CrossRef] [PubMed]
- Casey, J.; Davidson, N. Rates of formation and thermal stabilities of RNA:DNA and DNA:DNA duplexes at high concentrations of formamide. Nucleic Acids Res. 1977, 4, 1539–1552. [Google Scholar] [CrossRef] [PubMed]
- Trifonov, E.N. The triplet code from first principles. J. Biomol. Struc. Dyn. 2004, 22, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, A.E.; Trifonov, E.N. “Anticipated” Nucleosome positioning pattern in prokaryotes. Gene 2011, 488, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Hickey, D.A.; Benkel, B. Introns as relict retrotransposons: Implications for the evolutionary origin of eukaryotic mRNA splicing mechanisms. J. Theor. Biol. 1986, 121, 283–291. [Google Scholar] [CrossRef]
- Saladino, R.; Crestini, C.; Costanzo, G.; Di Mauro, E. On the Prebiotic Synthesis of Nucleobases, Nucleotides, Oligonucleotides, pre-RNA and pre-DNA Molecules. In Prebiotic Chemistry; Walde, P., Ed.; Topics in Current Chemistry; Springer: Berlin, Germany, 2005; Volume 259, pp. 29–68. [Google Scholar]
- Cleaves, H.J.; Michalkova Scott, A.; Hill, F.C.; Leszczynski, J.; Sahai, N.; Hazen, R. Mineral-organic interfacial processes: Potential roles in the origins of life. Chem. Soc. Rev. 2012, 41, 5502–5525. [Google Scholar] [CrossRef] [PubMed]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pino, S.; Costanzo, G.; Giorgi, A.; Šponer, J.; Šponer, J.E.; Mauro, E.D. Ribozyme Activity of RNA Nonenzymatically Polymerized from 3′,5′-Cyclic GMP. Entropy 2013, 15, 5362-5383. https://doi.org/10.3390/e15125362
Pino S, Costanzo G, Giorgi A, Šponer J, Šponer JE, Mauro ED. Ribozyme Activity of RNA Nonenzymatically Polymerized from 3′,5′-Cyclic GMP. Entropy. 2013; 15(12):5362-5383. https://doi.org/10.3390/e15125362
Chicago/Turabian StylePino, Samanta, Giovanna Costanzo, Alessandra Giorgi, Jiří Šponer, Judit E. Šponer, and Ernesto Di Mauro. 2013. "Ribozyme Activity of RNA Nonenzymatically Polymerized from 3′,5′-Cyclic GMP" Entropy 15, no. 12: 5362-5383. https://doi.org/10.3390/e15125362
APA StylePino, S., Costanzo, G., Giorgi, A., Šponer, J., Šponer, J. E., & Mauro, E. D. (2013). Ribozyme Activity of RNA Nonenzymatically Polymerized from 3′,5′-Cyclic GMP. Entropy, 15(12), 5362-5383. https://doi.org/10.3390/e15125362