
Fasting and Exercise Induce Changes in Serum Vitamin D Metabolites in Healthy Men

 , , ,
, , ,  ,
,
Abstract
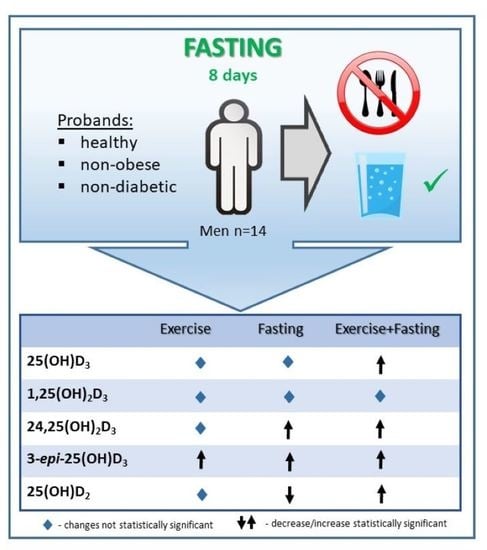
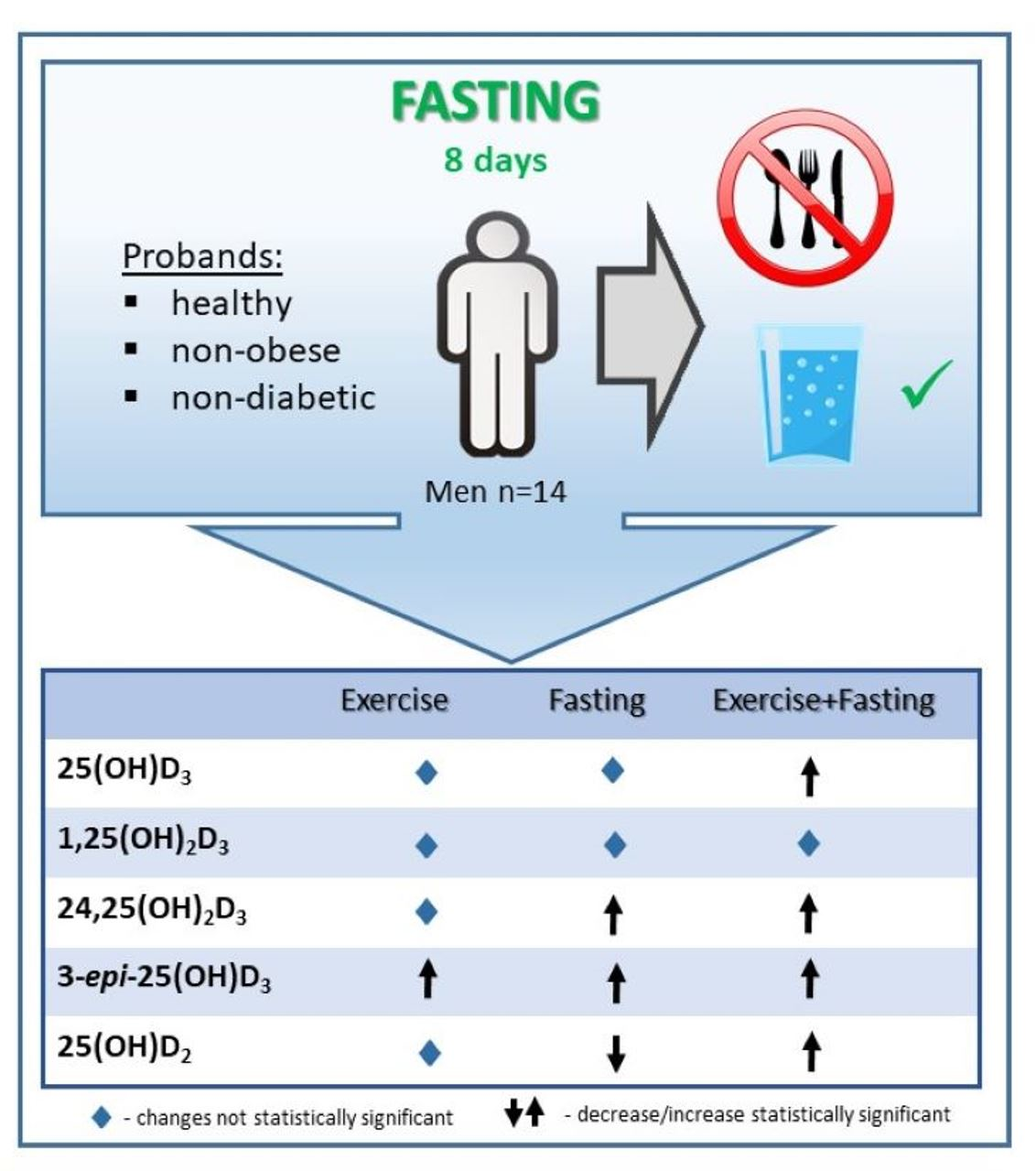
:
1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Study Group Characteristics
2.3. Anthropometric Measurements
2.4. Exercise Test
2.5. General Blood Parameter
2.6. Measurements of Serum Vitamin D Metabolites
2.7. Statistical Analysis
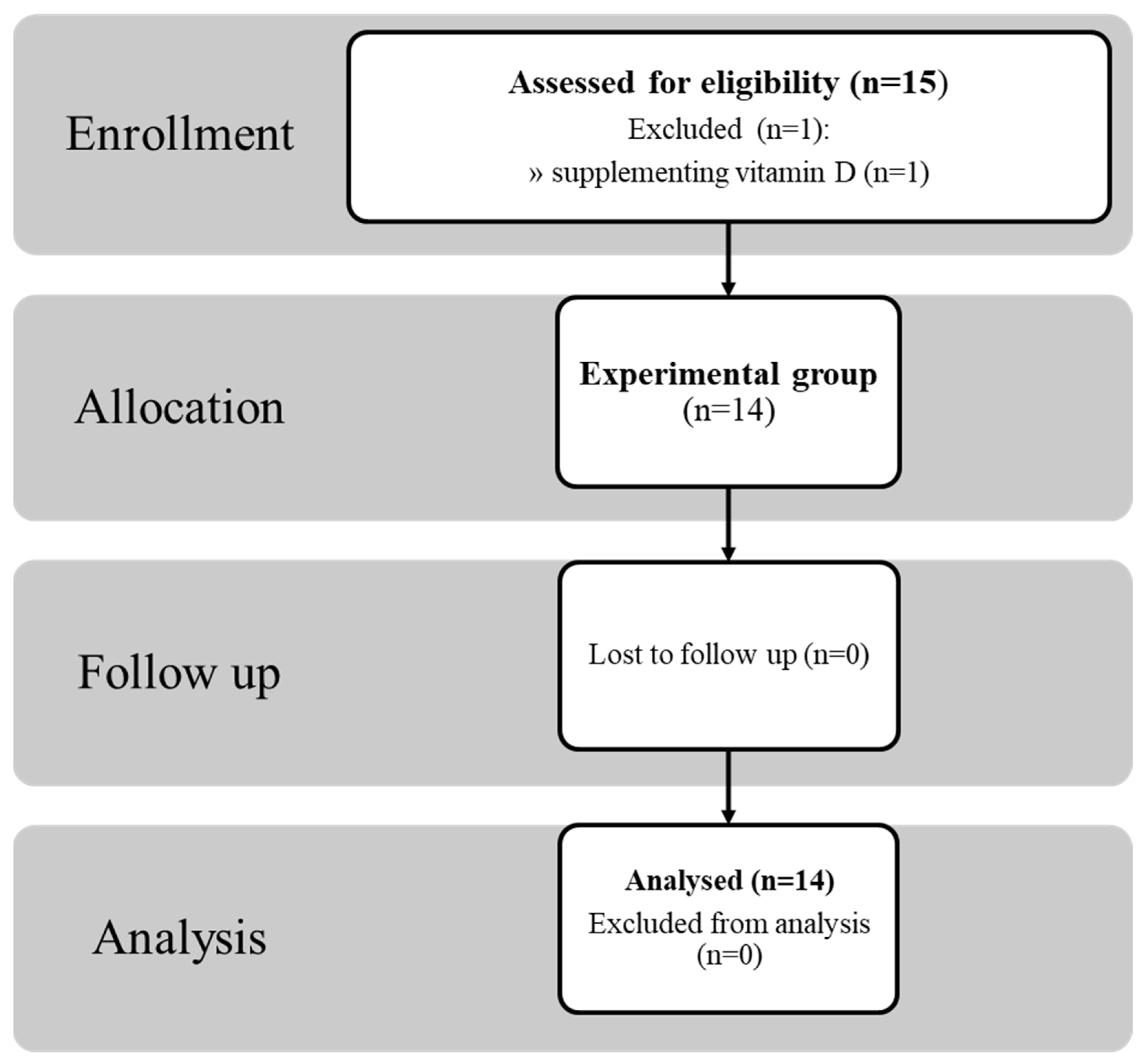
3. Results
3.1. Effect of 8 d of Fasting on Body Composition and Performance
3.2. General Outcomes
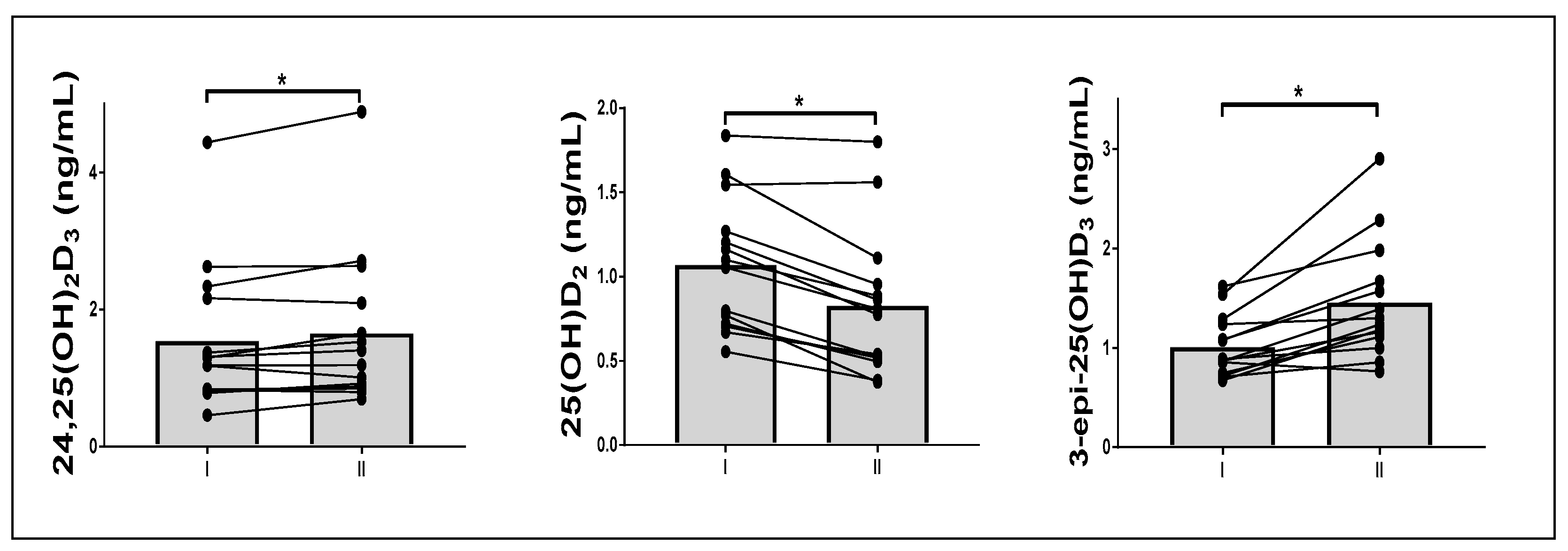
3.3. Effect of Fasting on Vitamin D Metabolite Response to a Single Exercise
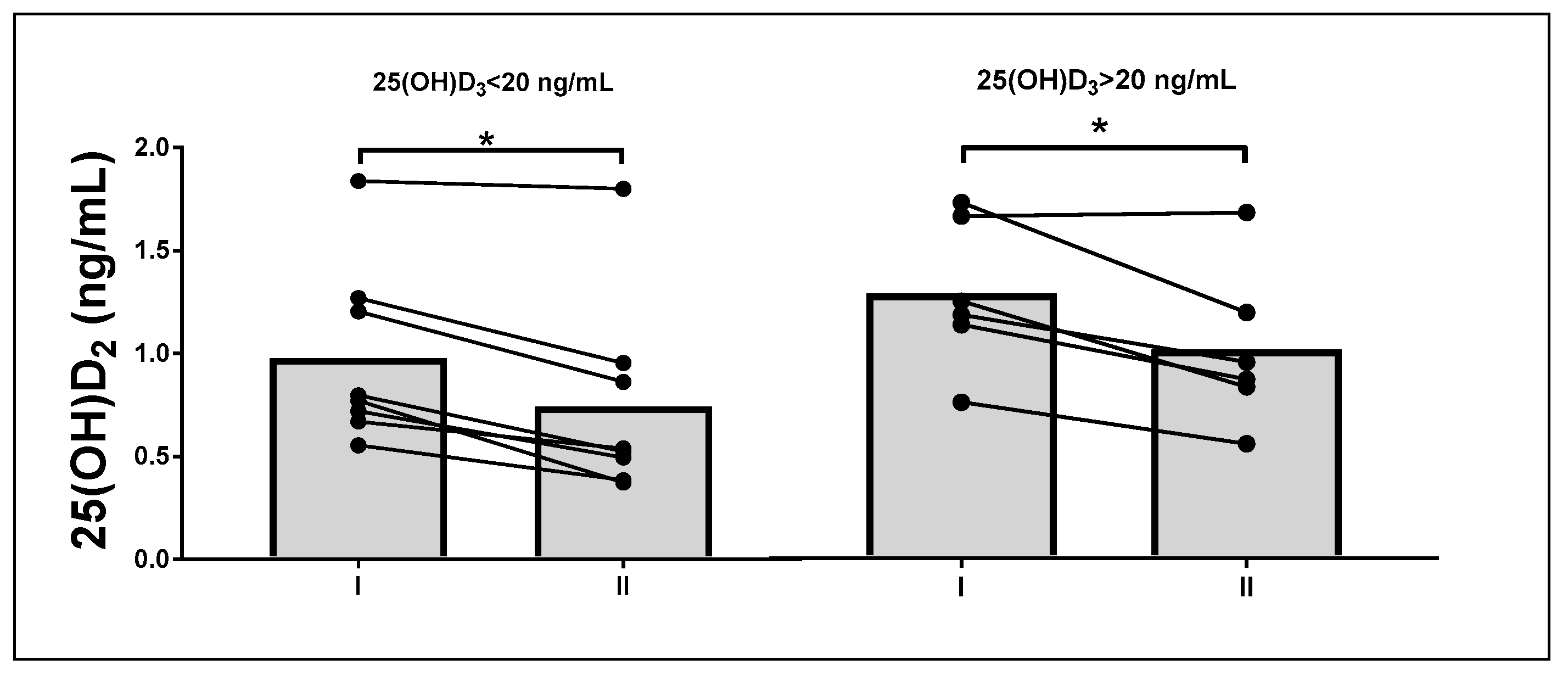
3.4. Subgroup Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Y.; Zhu, J.; DeLuca, H.F. Where is the vitamin D receptor? Arch Biochem. Biophys. 2012, 523, 123–133. [Google Scholar] [CrossRef]
- Wierzbicka, J.; Piotrowska, A.; Żmijewski, M.A. The renaissance of vitamin D. Acta Biochim. Pol. 2014, 61, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.T.; Tavera-Mendoza, L.E.; Laperriere, D.; Libby, E.; MacLeod, N.B.; Nagai, Y.; Bourdeau, V.; Konstorum, A.; Lallemant, B.; Zhang, R.; et al. Large-scale in silico and microarray-based identification of direct 1,25-dihydroxyvitamin D3 target genes. Mol. Endocrinol. 2005, 19, 2685–2695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutsey, P.L.; Eckfeldt, J.H.; Ogagarue, E.R.; Folsom, A.R.; Michos, E.D.; Gross, M. The 25-hydroxyvitamin D3 C-3 epimer: Distribution, correlates, and reclassification of 25-hydroxyvitamin D status in the population-based Atherosclerosis Risk in Communities Study (ARIC). Clin. Chim. Acta 2015, 442, 75–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, A.J.; Ritter, C.S.; Weiskopf, A.S.; Vouros, P.; Sasso, G.J.; Uskokovic, M.R.; Wang, G.; Reddy, G.S. Isolation and identification of 1alpha-hydroxy-3-epi-vitamin D3, a potent suppressor of parathyroid hormone secretion. J. Cell. Biochem. 2005, 96, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Yin, K.; Agrawal, D.K. Vitamin D and inflammatory diseases. J. Inflamm. Res. 2014, 7, 69–87. [Google Scholar] [CrossRef] [Green Version]
- Spach, K.M.; Nashold, F.E.; Dittel, B.N.; Hayes, C.E. IL-10 signaling is essential for 1,25-dihydroxyvitamin D-3-mediated inhibition of experimental autoimmune encephalomyelitis. J. Immunol. 2006, 177, 6030–6037. [Google Scholar] [CrossRef] [Green Version]
- Bosworth, C.R.; Levin, G.; Robinson-Cohen, C.; Hoofnagle, A.N.; Ruzinski, J.; Young, B.; Schwartz, S.M.; Himmelfarb, J.; Kestenbaum, B.; de Boer, I.H. The serum 24,25-dihydroxyvitamin D concentration, a marker of vitamin D catabolism, is reduced in chronic kidney disease. Kidney Int. 2012, 82, 693–700. [Google Scholar] [CrossRef] [Green Version]
- Mason, R.S.; Rybchyn, M.S.; Abboud, M.; Brennan-Speranza, T.C.; Fraser, D.R. The Role of Skeletal Muscle in Maintaining Vitamin D Status in Winter. Curr. Dev. Nutr. 2019, 3, nzz087. [Google Scholar] [CrossRef]
- Sun, X.; Cao, Z.B.; Taniguchi, H.; Tanisawa, K.; Higuchi, M. Effect of an Acute Bout of Endurance Exercise on Serum 25(OH)D Concentrations in Young Adults. J. Clin. Endocrinol. Metab. 2017, 102, 3937–3944. [Google Scholar] [CrossRef]
- Blum, M.; Dallal, G.E.; Dawson-Hughes, B. Body size and serum 25 hydroxy vitamin D response to oral supplements in healthy older adults. J. Am. Coll. Nutr. 2008, 27, 274–279. [Google Scholar] [CrossRef]
- Brouwer, D.A.J.; van Beek, J.; Ferwerda, H.; Brugman, A.M.; van der Klis, F.R.M.; van der Heiden, H.J.; Muskiet, F.A.J. Rat adipose tissue rapidly accumulates and slowly releases an orally-administered high vitamin D dose. Brit. J. Nutr. 1998, 79, 527–532. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Cao, Z.B.; Tanisawa, K.; Taniguchi, H.; Kubo, T.; Higuchi, M. Effects of chronic endurance exercise training on serum 25(OH)D concentrations in elderly Japanese men. Endocrine 2018, 59, 330–337. [Google Scholar] [CrossRef]
- Mieszkowski, J.; Stankiewicz, B.; Kochanowicz, A.; Niespodzinski, B.; Kowalik, T.; Zmijewski, M.A.; Kowalski, K.; Rola, R.; Bienkowski, T.; Antosiewicz, J. Ultra-Marathon-Induced Increase in Serum Levels of Vitamin D Metabolites: A Double-Blind Randomized Controlled Trial. Nutrients 2020, 12, 3629. [Google Scholar] [CrossRef]
- Abboud, M.; Puglisi, D.A.; Davies, B.N.; Rybchyn, M.; Whitehead, N.P.; Brock, K.E.; Cole, L.; Gordon-Thomson, C.; Fraser, D.R.; Mason, R.S. Evidence for a Specific Uptake and Retention Mechanism for 25-Hydroxyvitamin D (25OHD) in Skeletal Muscle Cells. Endocrinology 2013, 154, 3022–3030. [Google Scholar] [CrossRef] [Green Version]
- Aatsinki, S.M.; Elkhwanky, M.S.; Kummu, O.; Karpale, M.; Buler, M.; Viitala, P.; Rinne, V.; Mutikainen, M.; Tavi, P.; Franko, A.; et al. Fasting-Induced Transcription Factors Repress Vitamin D Bioactivation, a Mechanism for Vitamin D Deficiency in Diabetes. Diabetes 2019, 68, 918–931. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Kong, J.; Sun, T.; Li, G.; Szeto, F.L.; Liu, W.; Deb, D.K.; Wang, Y.; Zhao, Q.; Thadhani, R.; et al. 1,25-Dihydroxyvitamin D(3) suppresses inflammation-induced expression of plasminogen activator inhibitor-1 by blocking nuclear factor-kappaB activation. Arch. Biochem. Biophys. 2011, 507, 241–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, E.G.; Einhorn, T.A.; Norman, A.W. 24R,25-dihydroxyvitamin D3: An essential vitamin D3 metabolite for both normal bone integrity and healing of tibial fracture in chicks. Endocrinology 1997, 138, 3864–3872. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.J.; Taylor, R.L.; Reddy, G.S.; Grebe, S.K. C-3 epimers can account for a significant proportion of total circulating 25-hydroxyvitamin D in infants, complicating accurate measurement and interpretation of vitamin D status. J. Clin. Endocrinol. Metab. 2006, 91, 3055–3061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehan, V.K.; Torday, J.S.; Peleg, S.; Gennaro, L.; Vouros, P.; Padbury, J.; Rao, D.S.; Reddy, G.S. 1Alpha,25-dihydroxy-3-epi-vitamin D3, a natural metabolite of 1alpha,25-dihydroxy vitamin D3: Production and biological activity studies in pulmonary alveolar type II cells. Mol. Genet. Metab. 2002, 76, 46–56. [Google Scholar] [CrossRef]
- Binkley, N.; Ramamurthy, R.; Krueger, D. Low vitamin D status: Definition, prevalence, consequences, and correction. Endocrinol. Metab. Clin. N. Am. 2010, 39, 287–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karefylakis, C.; Pettersson-Pablo, P.; Sarnblad, S.; Rask, E.; Bitar, M.; Magnuson, A.; Eriksson, C.G. Vitamin D C3 epimer in a mid-Swedish region-Analytical measurement and epidemiology. Clin. Chim. Acta 2018, 478, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Maimoun, L.; Simar, D.; Caillaud, C.; Coste, O.; Barbotte, E.; Peruchon, E.; Rossi, M.; Mariano-Goulart, D. Response of calciotropic hormones and bone turnover to brisk walking according to age and fitness level. J. Sci. Med. Sport 2009, 12, 463–467. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| I | II | Change | p | |
|---|---|---|---|---|
| BW (kg) | 79.9 ± 10.98 | 74.8 ± 11.0 | −5.6 ± 1.4 | <0.01 |
| Fat (kg) | 15.8 ± 5.5 | 13.9 ± 5.7 | −2.0 ± 0.6 | <0.01 |
| FFM (kg) | 64.1 ± 5.8 | 60.7 ± 6.1 | −3.8 ± 1.3 | <0.01 |
| BMI (kg/m2) | 25.1 ± 3.3 | 23.4 ± 3.4 | −1.8 ± 0.5 | <0.01 |
| VO2 max (ml/min/kg) | 47.0 ± 12.0 | 40.0 ± 7.3 | −7.1 ± 7.8 | <0.01 |
| Load (W) | 212.1 ± 38.1 | 180.0 ± 36.7 | −32.3 ± 25.9 | <0.01 |
| I | I h | p (I vs. I h) | II | II h | p (II vs. II h) | |
|---|---|---|---|---|---|---|
| 25(OH)D3 (ng/mL) | 21.7 ± 6.9 | 22.3 ± 8.1 | 0.17 | 21.0 ± 7.9 | 23.2 ± 9.1 | 0.00 |
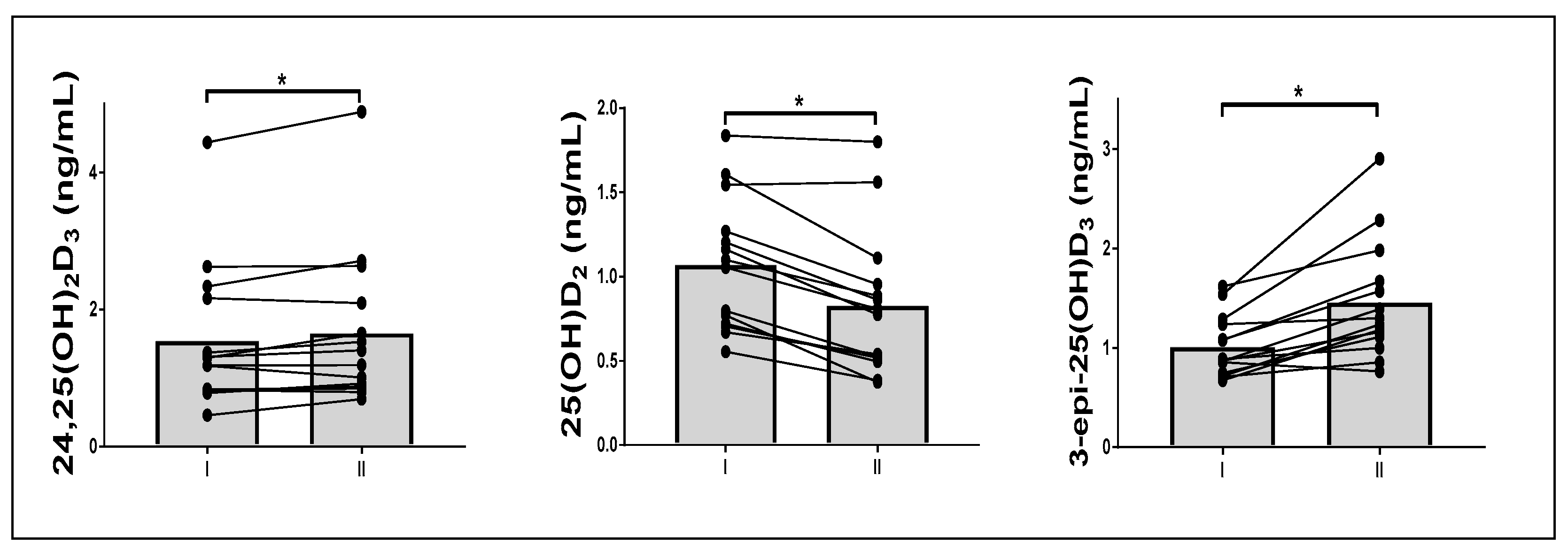
| 24,25(OH)2D3 (ng/mL) | 1.5 ± 1.1 | 1.6 ± 1.1 | 0.36 | 1.7 ± 1.2 | 1.9 ± 1.4 | 0.02 |
| 25(OH)D2 (ng/mL) | 1.1 ± 0.4 | 1.1 ± 0.5 | 0.19 | 0.8 ± 0.3 | 0.8 ± 0.3 | 0.01 |
| 3-epi-25(OH)D3 (ng/mL) | 1.0 ± 0.3 | 1.1 ± 0.3 | 0.00 | 1.5 ± 0.6 | 1.6 ± 0.7 | 0.00 |
| 1,25(OH)2D3 (pg/mL) | 64.1 ± 23.4 | 69.7 ± 27.9 | 0.34 | 72.6 ± 23.4 | 74.5 ± 28.7 | 0.76 |
| Baseline 25(OH)D3 | p | ||
|---|---|---|---|
| <20 ng/mL | >20 ng/mL | ||
| 25(OH)D3 (ng/mL) | 17.4 ± 2.5 | 27.5 ± 6.5 | <0.01 |
| 24,25(OH)2D3 (ng/mL) | 0.9 ± 0.3 | 2.4 ± 1.2 | 0.01 |
| 25(OH)D2 (ng/mL) | 1.0 ± 0.4 | 1.2 ± 0.3 | 0.33 |
| 3-epi-25(OH)D3 (ng/mL) | 0.8 ± 0.2 | 1.3 ± 0.3 | 0.01 |
| 1,25(OH)2D3 (pg/mL) | 48.8 ± 16.6 | 82.5 ± 15.8 | 0.01 |
| 25(OH)D3 < 20 ng/mL | 25(OH)D3 > 20 ng/mL | rANOVA | ||||
|---|---|---|---|---|---|---|
| II | II h | II | II h | p | ||
| 25(OH)D3 (ng/mL) | 15.57 ± 2.55 | 17.07 ± 2.86 | 27.27 ± 7.41 | 30.45 ± 8.42 * | 0.04 | 0.31 |
| 24,25(OH)2D3 (ng/mL) | 0.95 ± 0.27 | 1.02 ± 0.17 | 2.56 ± 1.25 | 2.93 ± 1.6 * | 0.05 | 0.24 |
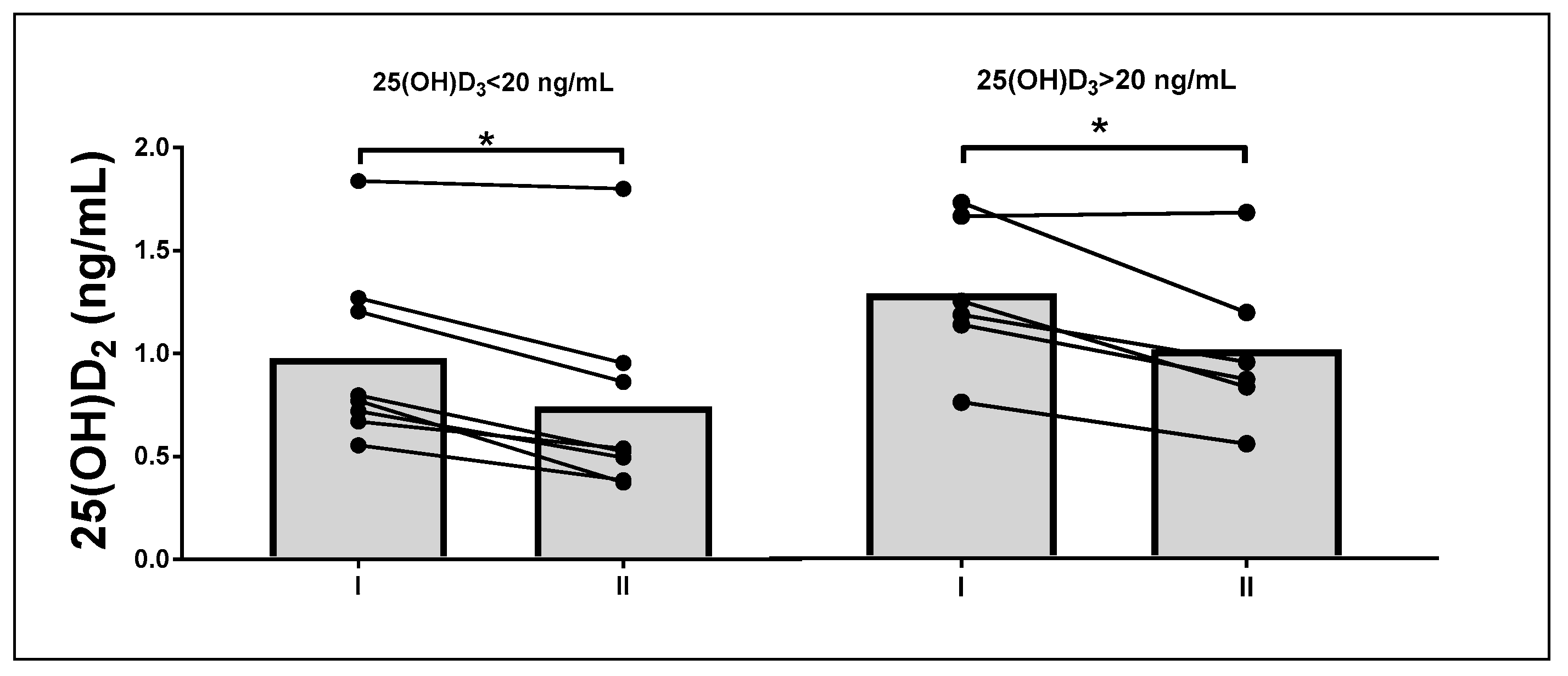
| 25(OH)D2 (ng/mL) | 0.59 ± 0.23 | 0.65 ± 0.23 | 0.94 ± 0.36 | 1.01 ± 0.34 | 0.78 | 0.01 |
| 3-epi-25(OH)D3 (ng/mL) | 1.08 ± 0.22 | 1.16 ± 0.21 | 1.93 ± 0.61 | 2.08 ± 0.68 | 0.10 | 0.07 |
| 1,25(OH)2D3 (pg/mL) | 61.87 ± 22.89 | 59.32 ± 28.83 | 85.42 ± 18.21 | 92.81 ± 16.12 | 0.45 | 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Żychowska, M.; Rola, R.; Borkowska, A.; Tomczyk, M.; Kortas, J.; Anczykowska, K.; Pilis, K.; Kowalski, K.; Pilch, W.; Antosiewicz, J. Fasting and Exercise Induce Changes in Serum Vitamin D Metabolites in Healthy Men. Nutrients 2021, 13, 1963. https://doi.org/10.3390/nu13061963
Żychowska M, Rola R, Borkowska A, Tomczyk M, Kortas J, Anczykowska K, Pilis K, Kowalski K, Pilch W, Antosiewicz J. Fasting and Exercise Induce Changes in Serum Vitamin D Metabolites in Healthy Men. Nutrients. 2021; 13(6):1963. https://doi.org/10.3390/nu13061963
Chicago/Turabian StyleŻychowska, Małgorzata, Rafał Rola, Andżelika Borkowska, Maja Tomczyk, Jakub Kortas, Katarzyna Anczykowska, Karol Pilis, Konrad Kowalski, Wanda Pilch, and Jędrzej Antosiewicz. 2021. "Fasting and Exercise Induce Changes in Serum Vitamin D Metabolites in Healthy Men" Nutrients 13, no. 6: 1963. https://doi.org/10.3390/nu13061963