
Effect of C-Type Natriuretic Peptide (CNP) on Spermatozoa Maturation in Adult Rat Epididymis

Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Establishment of Castrated Rat Model
2.3. Determination of the CNP/NPR-B Expression in Different Tissues of Rats by RT-PCR
2.4. Protein Concentration of CNP and cGMP in the Epididymal Fluid of Rats Using ELISA
2.5. Immunohistochemistry
2.6. Expression of NPR-B in the Spermatozoa by Immunofluorescence
2.7. Mobility and Maturation of Epididymal Sperm Treated with CNP In Vitro
2.8. Detection of the Signaling Pathway of CNP in Epididymal Sperm Motility
2.9. Statistical Analyses
3. Results
3.1. Expression of CNP/NPR-B in Different Rat Tissues and in Epididymis at Various Times after Birth
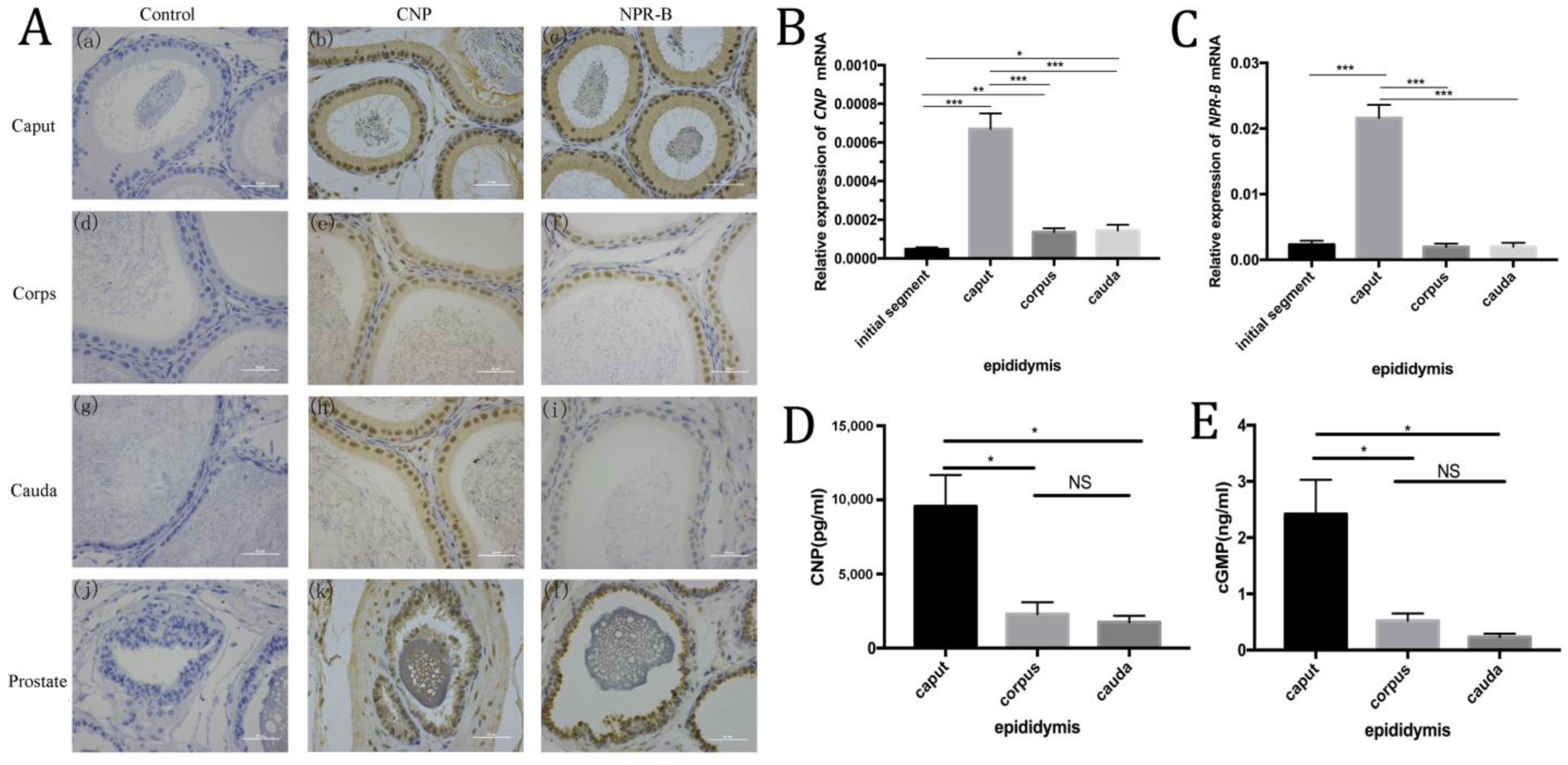
3.2. Expression of CNP/NPR-B in Different Segments of Rat Epididymis
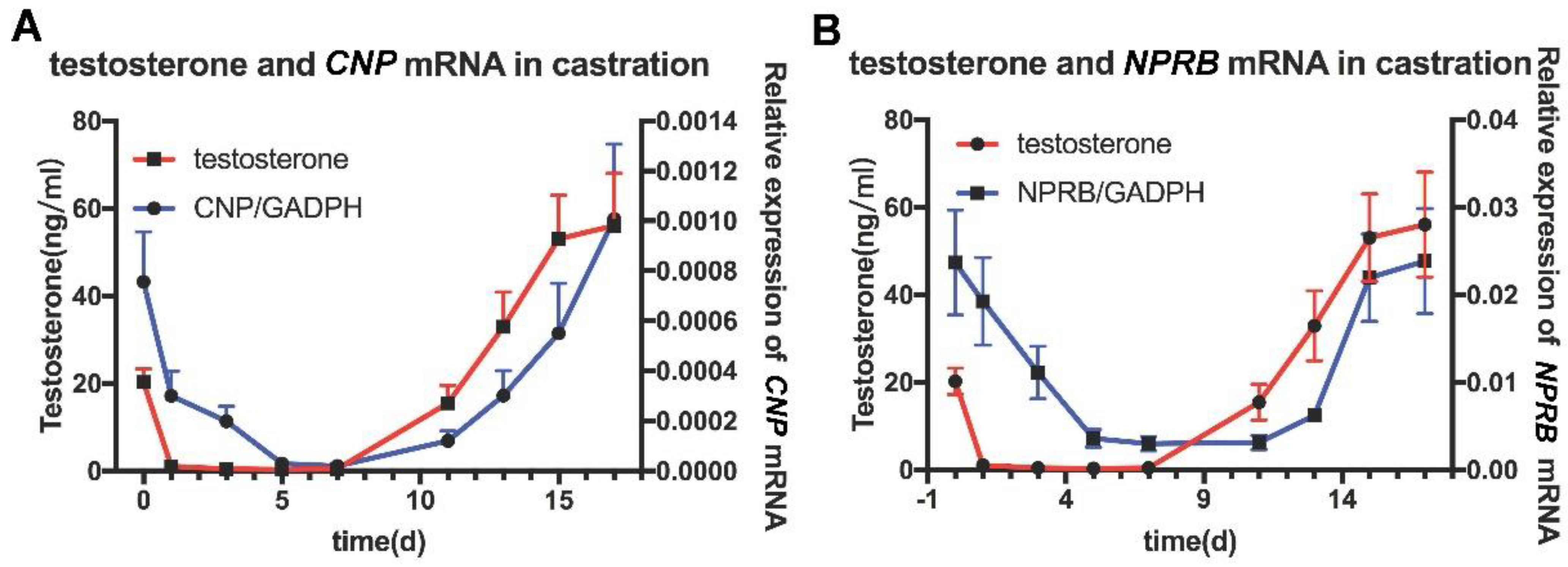
3.3. Expression of CNP, NPR-B in the Castrated Rat Model
3.4. Effect of CNP on Motility of the Epididymis Sperm In Vitro
3.5. Effect of CNP on Maturation-Related Factors of the Epididymis Sperm In Vitro
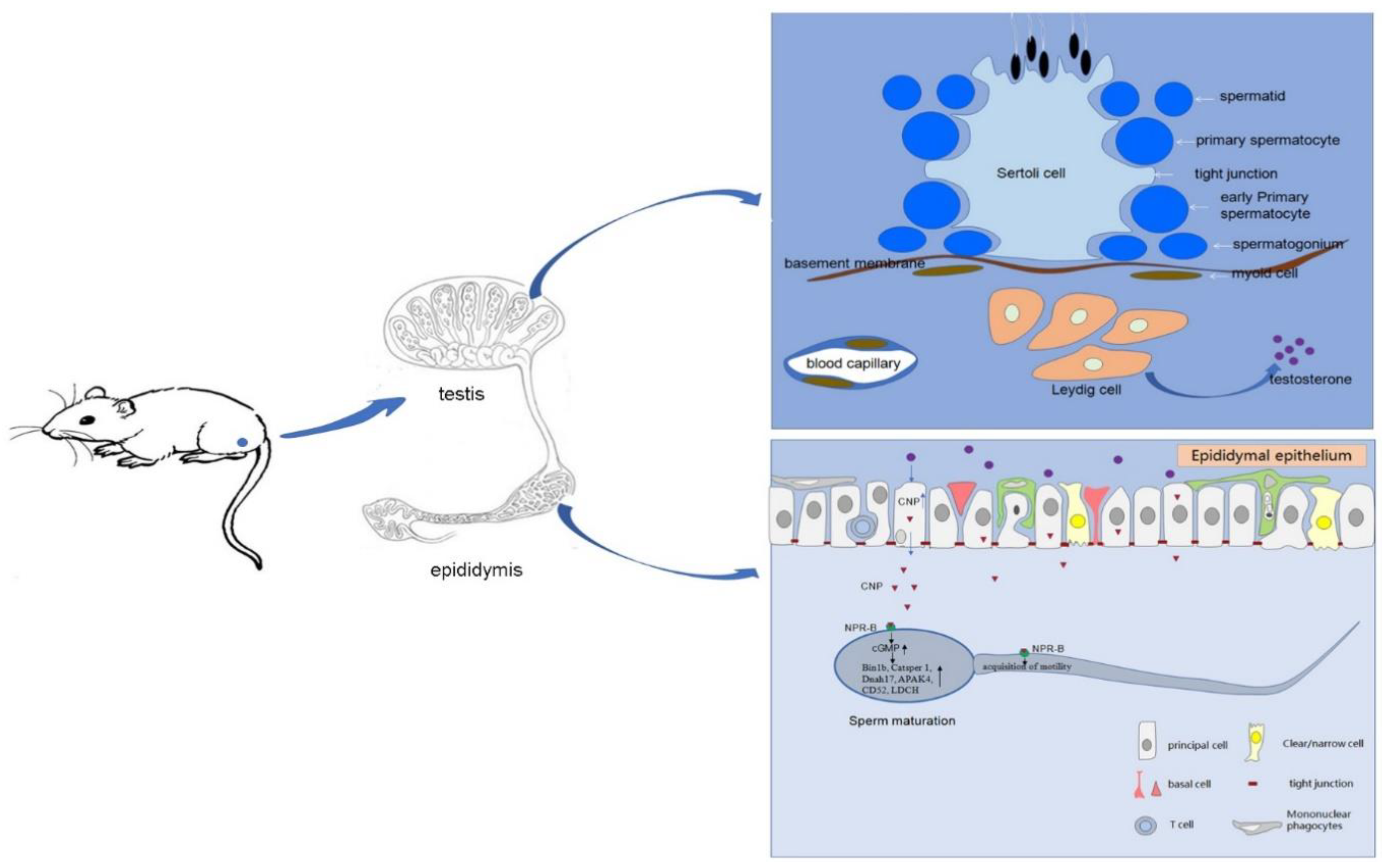
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Dun, M.D.; Aitken, R.J.; Nixon, B. The role of molecular chaperones in spermatogenesis and the post-testicular maturation of mammalian spermatozoa. Hum. Reprod. Update 2012, 18, 420–435. [Google Scholar] [CrossRef]
- Sullivan, R.; Mieusset, R. The human epididymis: Its function in sperm maturation. Hum. Reprod. Update 2016, 22, 574–587. [Google Scholar] [CrossRef] [PubMed]
- Björkgren, I.; Sipilä, P. The impact of epididymal proteins on sperm function. Reproduction 2019, 158, R155–R167. [Google Scholar] [CrossRef] [PubMed]
- Pal, P.; Biswas, S.; Mukhopadhyay, P.K. Molecular perspective concerning fluoride and arsenic mediated disorders on epididymal maturation of spermatozoa: A concise review. Hum. Exp. Toxicol. 2021, 40, 2025–2038. [Google Scholar] [CrossRef] [PubMed]
- Ozkocer, S.E.; Konac, E. The current perspective on genetic and epigenetic factors in sperm maturation in the epididymis. Andrologia 2021, 53, e13989. [Google Scholar] [CrossRef]
- Sudoh, T.; Minamino, N.; Kangawa, K.; Matsuo, H. C-type natriuretic peptide (cnp): A new member of natriuretic peptide family identified in porcine brain. Biochem. Biophys. Res. Commun. 1990, 168, 863–870. [Google Scholar] [CrossRef]
- Zhang, M.; Su, Y.Q.; Sugiura, K.; Xia, G.; Eppig, J.J. Granulosa cell ligand nppc and its receptor npr2 maintain meiotic arrest in mouse oocytes. Science 2010, 330, 366–369. [Google Scholar] [CrossRef]
- Chrisman, T.D.; Schulz, S.; Potter, L.R.; Garbers, D.L. Seminal plasma factors that cause large elevations in cellular cyclic GMP are C-type natriuretic peptides. J. Biol. Chem. 1993, 268, 3698–3703. [Google Scholar] [CrossRef]
- Nielsen, S.J.; Gøtze, J.P.; Jensen, H.L.; Rehfeld, J.F. ProCNP and CNP are expressed primarily in male genital organs. Regul. Pept. 2008, 146, 204–212. [Google Scholar] [CrossRef]
- Nielsen, S.J.; Rehfeld, J.F.; Pedersen, F.; Kastrup, J.; Videbaek, R.; Goetze, J.P. Measurement of Pro-C-Type Natriuretic Peptide in Plasma. Clin. Chem. 2005, 51, 2173–2176. [Google Scholar] [CrossRef][Green Version]
- Sogawa, C.; Fujiwara, Y.; Tsukamoto, S.; Ishida, Y.; Yoshii, Y.; Furukawa, T.; Kunieda, T.; Saga, T. Mutant phenotype analysis suggests potential roles for C-type natriuretic peptide receptor (NPR-B) in male mouse fertility. Reprod. Biol. Endocrinol. 2014, 12, 64. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Lei, L.; Zhao, Q.; Gong, Y.; Guan, G.; Huang, S. C-Type Natriuretic Peptide/Natriuretic Peptide Receptor 2 Is Involved in Cell Proliferation and Testosterone Production in Mouse Leydig Cells. World J. Mens. Health. 2019, 37, 186–198. [Google Scholar] [CrossRef]
- Yu, Y.; Chen, Y.; Mei, C.; Li, N.; Wu, K.; Huang, D. C-type natriuretic peptide stimulates function of the murine Sertoli cells via activation of the NPR-B/cGMP/PKG signaling pathway. Acta Biochim. Pol. 2021, 68, 603–609. [Google Scholar] [CrossRef]
- Kong, N.; Xu, X.; Zhang, Y.; Wang, Y.; Hao, X.; Zhao, Y.; Qiao, J.; Xia, G.; Zhang, M. Natriuretic peptide type C induces sperm attraction for fertilization in mouse. Sci. Rep. 2017, 7, 39711. [Google Scholar] [CrossRef]
- Xia, H.; Chen, Y.; Wu, K.J.; Zhao, H.; Xiong, C.L.; Huang, D.H. Role of C-type natriuretic peptide in the function of normal human sperm. Asian J. Androl. 2016, 18, 80–84. [Google Scholar]
- Wu, K.; Mei, C.; Chen, Y.; Guo, L.; Yu, Y.; Huang, D. C-type natriuretic peptide regulates sperm capacitation by the cGMP/PKG signalling pathway via Ca2+ influx and tyrosine phosphorylation. Reprod. Biomed. Online 2019, 38, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar]
- Mosli, H.H.; Esmat, A.; Atawia, R.T.; Shoieb, S.M.; Mosli, H.A.; Abdel-Naim, A.B. Metformin Attenuates Testosterone-Induced Prostatic Hyperplasia in Rats: A Pharmacological Perspective. Sci. Rep. 2015, 5, 15639. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Dong, X.; Fu, S.; Wang, X.; Li, H.; Song, G.; Huang, D. C-Type Natriuretic Peptide (CNP) Could Improve Sperm Motility and Reproductive Function of Asthenozoospermia. Int. J. Mol. Sci. 2022, 23, 10370. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Suzuki, K.; Yu, X.; Chaurand, P.; Araki, Y.; Lareyre, J.J.; Caprioli, R.M.; Orgebin-Crist, M.C.; Matusik, R.J. Epididymis-specific lipocalin promoters. Asian J. Androl. 2007, 9, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Robertson, M.J.; Kent, K.; Tharp, N.; Nozawa, K.; Dean, L.; Mathew, M.; Grimm, S.L.; Yu, Z.; Légaré, C.; Fujihara, Y.; et al. Large-scale discovery of male reproductive tract-specific genes through analysis of RNA-seq datasets. BMC Biol. 2020, 18, 103. [Google Scholar] [CrossRef]
- Oh, J.; Lee, J.; Woo, J.M.; Choi, E.; Park, I.; Han, C.; Baek, N.; Lee, H.; Kim, D.H.; Cho, C. Systematic identification and integrative analysis of novel genes expressed specifically or predominantly in mouse epididymis. BMC Genom. 2006, 7, 314. [Google Scholar] [CrossRef]
- Breton, S.; Ruan, Y.C.; Park, Y.J.; Kim, B. Regulation of epithelial function, differentiation, and remodeling in the epididymis. Asian J. Androl. 2016, 18, 3–9. [Google Scholar] [CrossRef]
- Xu, B.; Abdel-Fattah, R.; Yang, L.; Crenshaw, S.A.; Black, M.B.; Hinton, B.T. Testicular Lumicrine Factors Regulate ERK, STAT, and NFKB Pathways in the Initial Segment of the Rat Epididymis to Prevent Apoptosis1. Biol. Reprod. 2011, 84, 1282–1291. [Google Scholar] [CrossRef][Green Version]
- Jahnukainen, K.; Chrysis, D.; Hou, M.; Parvinen, M.; Eksborg, S.; Soder, O. Increased apoptosis occurring during the first wave of spermatogenesis is stage-specific and primarily affects midpachytene spermatocytes in the rat testis. Biol. Reprod. 2004, 70, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Barrachina, F.; Battistone, M.A.; Castillo, J.; Mallofré, C.; Jodar, M.; Breton, S.; Oliva, R. Sperm acquire epididymis-derived proteins through epididymosomes. Hum. Reprod. 2022, 37, 651–668. [Google Scholar] [CrossRef]
- Li, P.; Chan, H.C.; He, B.; So, S.C.; Chung, Y.W.; Shang, Q.; Zhang, Y.D.; Zhang, Y.L. An antimicrobial peptide gene found in the male reproductive system of rats. Science 2001, 291, 1783–1785. [Google Scholar] [CrossRef] [PubMed]
- Özbek, M.; Hitit, M.; Öztop, M.; Beyaz, F.; Ergün, E.; Ergün, L. Spatiotemporal expression patterns of natriuretic peptides in rat testis and epididymis during postnatal development. Andrologia 2019, 51, e13387. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Fok, K.L.; Dai, P.; Qiao, F.; Zhang, M.; Liu, H.; Sang, M.; Ye, M.; Liu, Y.; Zhou, Y.; et al. Spatio-temporal landscape of mouse epididymal cells and specific mitochondria-rich segments defined by large-scale single-cell RNA-seq. Cell Discov. 2021, 7, 34. [Google Scholar] [CrossRef] [PubMed]
- Browne, J.A.; Yang, R.; Leir, S.H.; Eggener, S.E.; Harris, A. Expression profiles of human epididymis epithelial cells reveal the functional diversity of caput, corpus and cauda regions. Mol. Hum. Reprod. 2016, 22, 69–82. [Google Scholar] [CrossRef]
- Zhao, W.; Quansah, E.; Yuan, M.; Gou, Q.; Mengal, K.; Li, P.; Wu, S.; Xu, C.; Yi, C.; Cai, X. Region-specific gene expression in the epididymis of Yak. Theriogenology 2019, 139, 132–146. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Z.; Kang, S.Y.; Lee, S.J.; Cho, K.W. Localization of receptors for natriuretic peptide and endothelin in the duct of the epididymis of the freshwater turtle. Gen. Comp. Endocrinol. 2000, 118, 26–38. [Google Scholar] [CrossRef]
- Mietens, A.; Tasch, S.; Feuerstacke, C.; Eichner, G.; Volkmann, J.; Schermuly, R.T.; Grimminger, F.; Müller, D.; Middendorff, R. Phosphodiesterase 5 (PDE5) inhibition, ANP and NO rapidly reduce epididymal duct contractions, but long-term PDE5 inhibition in vivo does not. Mol. Cell Endocrinol. 2012, 349, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Thong, A.; Müller, D.; Feuerstacke, C.; Mietens, A.; Stammler, A.; Middendorff, R. Neutral endopeptidase (CD10) is abundantly expressed in the epididymis and localized to a distinct population of epithelial cells—Its relevance for CNP degradation. Mol. Cell Endocrinol. 2014, 382, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.J.; Hardy, M.P.; Inigo, I.V.; Huhtaniemi, I.; Bardin, C.W.; Moo-Young, A.J. Effects of androgen on androgen receptor expression in rat testicular and epididymal cells: A quantitative immunohistochemical study. Biol. Reprod. 2000, 63, 368–376. [Google Scholar] [CrossRef]
- Vreeburg, J.T.; Holland, M.K.; Cornwall, G.A.; Orgebin-Crist, M.C. Secretion and transport of mouse epididymal proteins after injection of 35S-methionine. Biol. Reprod. 1990, 43, 113–120. [Google Scholar] [CrossRef]
- Pujianto, D.A.; Loanda, E.; Sari, P.; Midoen, Y.H.; Soeharso, P. Sperm-associated antigen 11A is expressed exclusively in the principal cells of the mouse caput epididymis in an androgen-dependent manner. Reprod. Biol. Endocrinol. 2013, 11, 59. [Google Scholar] [CrossRef]
- Zhu, C.F.; Liu, Q.; Zhang, L.; Yuan, H.X.; Zhen, W.; Zhang, J.S.; Chen, Z.J.; Hall, S.H.; French, F.S.; Zhang, Y.L. RNase9, an androgen-dependent member of the RNase A family, is specifically expressed in the rat epididymis. Biol. Reprod. 2007, 76, 63–73. [Google Scholar] [CrossRef]
- Fernandez, C.D.; Porto, E.M.; Arena, A.C.; Kempinas Wde, G. Effects of altered epididymal sperm transit time on sperm quality. Int. J. Androl. 2008, 31, 427–437. [Google Scholar] [CrossRef]
- Khurana, M.L.; Pandey, K.N. Receptor-mediated stimulatory effect of atrial natriuretic factor, brain natriuretic peptide, and C-type natriuretic peptide on testosterone production in purified mouse Leydig cells: Activation of cholesterol side-chain cleavage enzyme. Endocrinology 1993, 133, 2141–2149. [Google Scholar] [CrossRef]
- Reis, A.M.; Honorato-Sampaio, K. C-type natriuretic peptide: A link between hyperandrogenism and anovulation in a mouse model of polycystic ovary syndrome. Clin. Sci. 2018, 132, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Olney, R.C.; Prickett, T.C.; Yandle, T.G.; Espiner, E.A.; Han, J.C.; Mauras, N. Amino-terminal propeptide of C-type natriuretic peptide and linear growth in children: Effects of puberty, testosterone, and growth hormone. J. Clin. Endocrinol. Metab. 2007, 92, 4294–5298. [Google Scholar] [CrossRef] [PubMed][Green Version]
- van Der Horst, G.; Seier, J.V.; Spinks, A.C.; Hendricks, S. The maturation of sperm motility in the epididymis and vas deferens of the vervet monkey, Cercopithecus aethiops. Int. J. Androl. 1999, 22, 197–207. [Google Scholar] [CrossRef]
- Guyonnet, B.; Dacheux, F.; Dacheux, J.L.; Gatti, J.L. The epididymal transcriptome and proteome provide some insights into new epididymal regulations. J. Androl. 2011, 32, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Björkgren, I.; Alvarez, L.; Blank, N.; Balbach, M.; Turunen, H.; Laajala, T.D.; Toivanen, J.; Krutskikh, A.; Wahlberg, N.; Huhtaniemi, I.; et al. Targeted inactivation of the mouse epididymal beta-defensin 41 alters sperm flagellar beat pattern and zona pellucida binding. Mol. Cell Endocrinol. 2016, 427, 143–154. [Google Scholar] [CrossRef]
- Diao, R.; Fok, K.L.; Chen, H.; Yu, M.K.; Duan, Y.; Chung, C.M.; Li, Z.; Wu, H.; Li, Z.; Zhang, H.; et al. Deficient human β-defensin 1 underlies male infertility associated with poor sperm motility and genital tract infection. Sci. Transl. Med. 2014, 6, 249ra108. [Google Scholar] [CrossRef]
- Dey, S.; Brothag, C.; Vijayaraghavan, S. Signaling Enzymes Required for Sperm Maturation and Fertilization in Mammals. Front. Cell Dev. Biol. 2019, 7, 341. [Google Scholar] [CrossRef]
- Cohen, D.J.; Rochwerger, L.; Ellerman, D.A.; Morgenfeld, M.M.; Busso, D.; Cuasnicú, P.S. Relationship between the association of rat epididymal protein “DE” with spermatozoa and the behavior and function of the protein. Mol. Reprod. Dev. 2000, 56, 180–188. [Google Scholar] [CrossRef]
- Chen, H.; Alves, M.B.R.; Belleannée, C. Contribution of epididymal epithelial cell functions to sperm epigenetic changes and the health of progeny. Hum. Reprod. Update 2021, 28, 51–66. [Google Scholar] [CrossRef]
- Zhao, Y.; Diao, H.; Ni, Z.; Hu, S.; Yu, H.; Zhang, Y. The epididymis-specific antimicrobial peptide β-defensin 15 is required for sperm motility and male fertility in the rat (Rattus norvegicus). Cell Mol. Life Sci. 2011, 68, 697–708. [Google Scholar] [CrossRef]
- Gundogan, G.I.; Irez, T.; Bozkurt, H.H. Is there a relationship between infertility and fertilin β protein distribution? Rev. Int. Androl. 2022, 20, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Fàbrega, A.; Guyonnet, B.; Dacheux, J.L.; Gatti, J.L.; Puigmulé, M.; Bonet, S.; Pinart, E. Expression, immunolocalization and processing of fertilins ADAM-1 and ADAM-2 in the boar (Sus domesticus) spermatozoa during epididymal maturation. Reprod. Biol. Endocrinol. 2011, 9, 96. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Serial Number | Gene | Forward Primer | Reverse Primer | Length (bp) | Annealing Temperature |
|---|---|---|---|---|---|
| NM_017008.4 | GADPH | 5′ CAGTGCCAGCCTCGTCTCAT 3′ | 5′ AGGGGCCATCCACAGTCTTC 3′ | 202 | 60 °C |
| NM_053750.1 | CNP | 5′ CAACGCGCGCAAATACAAA 3′ | 5′ TAACATCCCAGACCGCTCATG 3′ | 163 | 60 °C |
| XM_006238010.4 | NPR-B | 5′ CCAAATTCTACCCCACTCTG 3′ | 5′ TAAGAAGTGTCACCACCTGC 3′ | 183 | 60 °C |
| XM_039101330.1 | Catsper 1 | 5′ TTTACCTGCCTCTTCCTCTTCT 3′ | 5′ ACCAGGTTGAGGAAGATGAAGT 3′ | 227 | 60 °C |
| NM_024402.1 | Akap4 | 5′ AGAGTCATCGCAGCATCCAA 3′ | 5′ TAATTCGCCGTCTGACCTGG 3′ | 258 | 60 °C |
| NM_053983.2 | CD52 | 5′ AGAAAAACCCCTGGGAAACC 3′ | 5′ GTTGGGGTGTCTCTTGCTAC 3′ | 184 | 60 °C |
| NM_017266.2 | LDHC | 5′ TCCACCTGTGAAGGCTCAAC 3′ | 5′ ATGCAGAAGATCCAGTGCCTC 3′ | 221 | 60 °C |
| XM_039087513.1 | Dnah17 | 5′ TGGAGGAGGTCCTCTACTCT 3′ | 5′ CCCATCACGAACATCTCGTT 3′ | 121 | 60 °C |
| NM_145087.2 | Bin1b | 5′ AGTCTCATCTGCTTTCCTGCAC 3′ | 5′ CACGGTGTTTCTGATTCCAGG 3′ | 128 | 60 °C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, H.; Yu, Y.; Mei, C.; Zhang, T.; Kang, Y.; Li, N.; Huang, D. Effect of C-Type Natriuretic Peptide (CNP) on Spermatozoa Maturation in Adult Rat Epididymis. Curr. Issues Mol. Biol. 2023, 45, 1681-1692. https://doi.org/10.3390/cimb45020108
Zhao H, Yu Y, Mei C, Zhang T, Kang Y, Li N, Huang D. Effect of C-Type Natriuretic Peptide (CNP) on Spermatozoa Maturation in Adult Rat Epididymis. Current Issues in Molecular Biology. 2023; 45(2):1681-1692. https://doi.org/10.3390/cimb45020108
Chicago/Turabian StyleZhao, Hu, Yuejin Yu, Chunlei Mei, Tianyu Zhang, Yafei Kang, Na Li, and Donghui Huang. 2023. "Effect of C-Type Natriuretic Peptide (CNP) on Spermatozoa Maturation in Adult Rat Epididymis" Current Issues in Molecular Biology 45, no. 2: 1681-1692. https://doi.org/10.3390/cimb45020108
APA StyleZhao, H., Yu, Y., Mei, C., Zhang, T., Kang, Y., Li, N., & Huang, D. (2023). Effect of C-Type Natriuretic Peptide (CNP) on Spermatozoa Maturation in Adult Rat Epididymis. Current Issues in Molecular Biology, 45(2), 1681-1692. https://doi.org/10.3390/cimb45020108