Practical Management of Iron Overload Disorder (IOD) in Black Rhinoceros (BR; Diceros bicornis)

 , ,
, ,
Abstract
:Simple Summary
Abstract
Table of Contents
- 1.
- Introduction
- 1.1.
- Why and how do we know BR have problems accumulating iron?
- 1.2.
- Why does iron overload matter to wellness?
- 1.3.
- How does IOD work in BR?
- 1.4.
- What health complications connected to iron are of concern?
- 2.
- Evidence-Based Veterinary Practice: Technical and Clinical Aspects
- 2.1.
- Clinical Signs
- 2.2.
- Diagnostic Testing
- 2.2.1.
- Serum iron
- 2.2.2.
- Total iron binding capacity (TIBC) and transferrin saturation
- 2.2.3.
- Ferritin
- 2.3.
- Perspectives on Integrative Monitoring
- 2.4.
- Inflammatory Biomarker Connections to IOD
- 2.5.
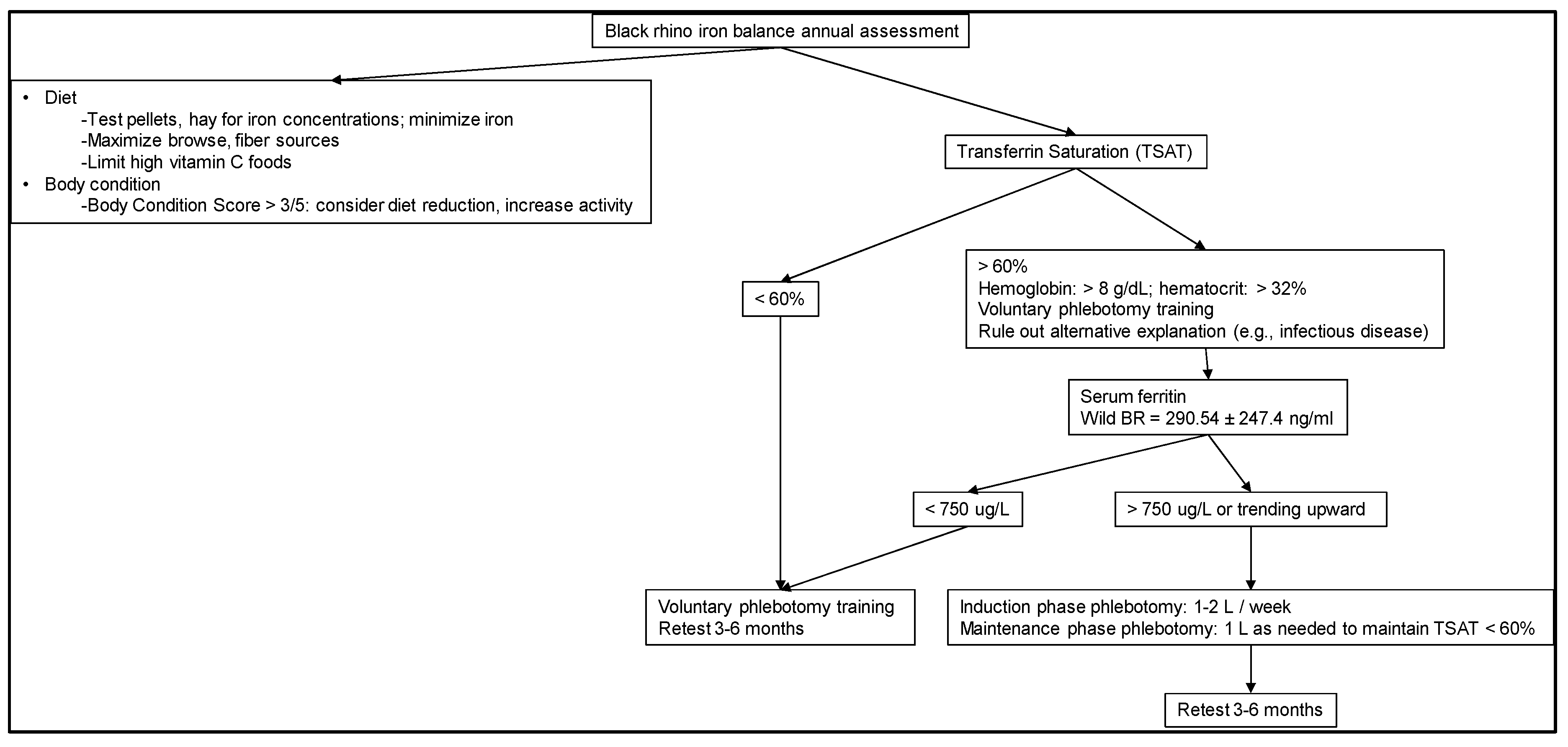
- Recommendations for Diagnosis
- 3.
- Evidence-Based Nutrition Practices
- 3.1.
- Could we make the diet more like the wild?
- 3.2.
- Has diet ever changed the impact of IOD?
- 3.3.
- What are the best practices for feeding black rhinos?
- 4.
- Treatment and Prevention
- 4.1.
- Hematologic Sampling Recommendations
- 4.2.
- Therapeutic Large Volume Phlebotomy
- 4.3.
- Techniques for VTLVP
- 4.4.
- Pharmacologic Chelation Therapy
- 4.5.
- History of Synthetic Chelation for BR
- 5.
- Animal Husbandry and Operant Conditioning Practices
- 6.
- Conclusions
1. Introduction
1.1. Why and How Do We Know BR Have Problems Accumulating Iron?
1.2. Why Does Iron Overload Matter to Wellness?
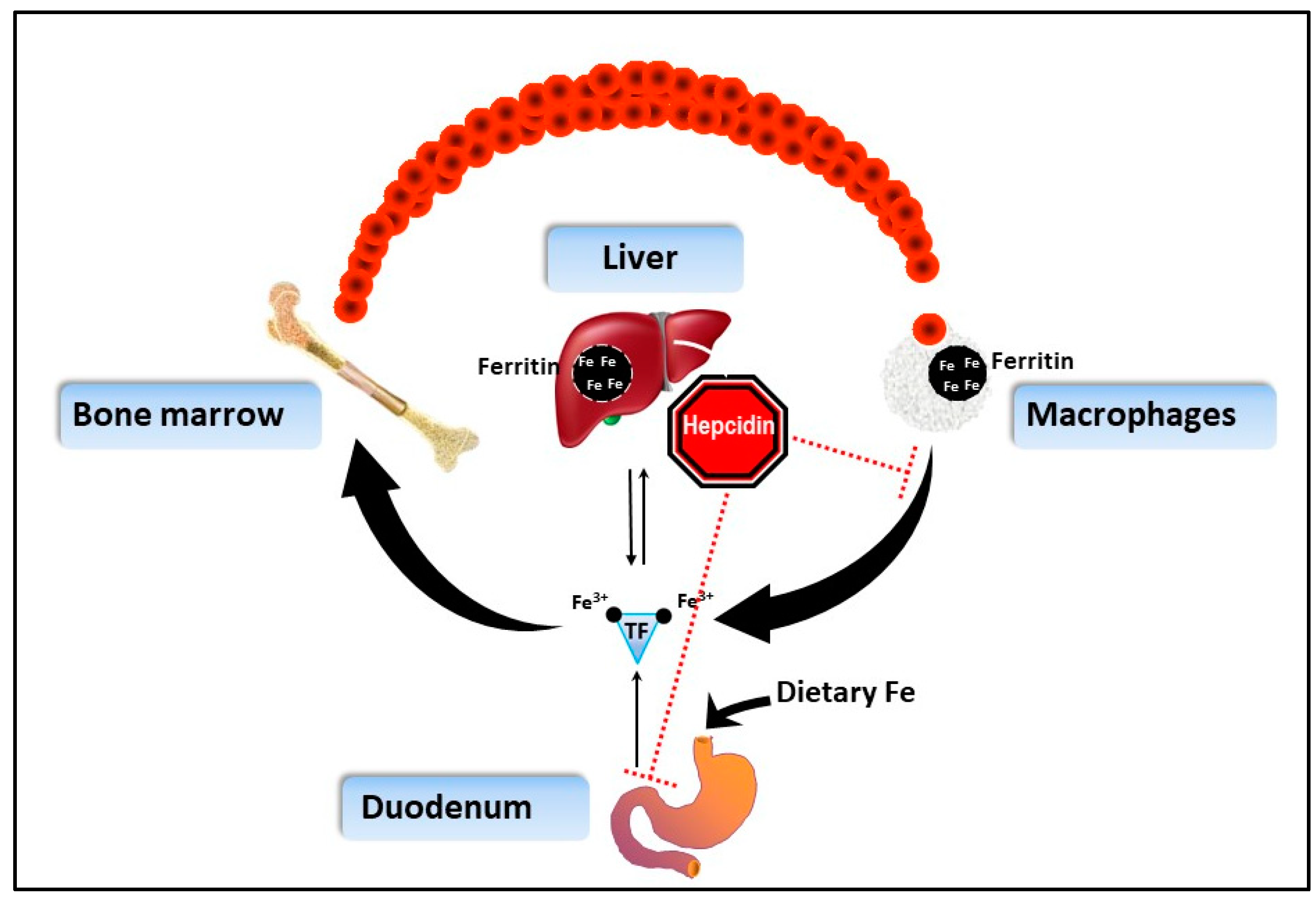
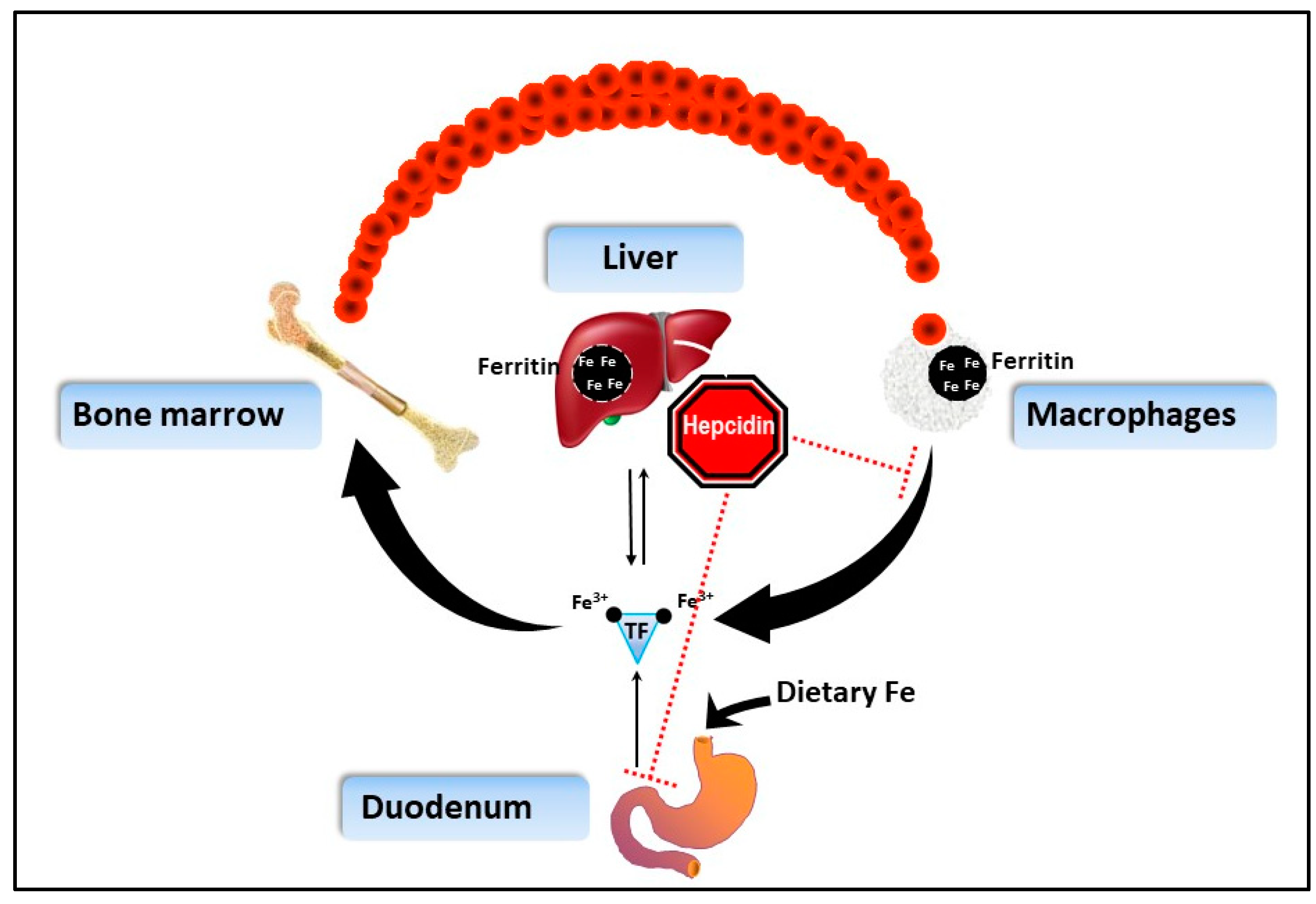
1.3. How Does IOD Work in BR?
1.4. What Health Complications Connected to Iron Are of Concern?
2. Evidence-Based Veterinary Practice: Technical and Clinical Aspects
2.1. Clinical Signs
2.2. Diagnostic Testing
- Serum iron (the amount of unbound iron in the blood)
- Total iron-binding capacity (TIBC; indirect transferrin level)
- Transferrin saturation (the percentage of TIBC occupied by iron) [46]
- Ferritin (iron carrier protein; used with the exclusion of other diseases that may elevate ferritin)
2.2.1. Serum Iron—Review and Comparative Evaluation
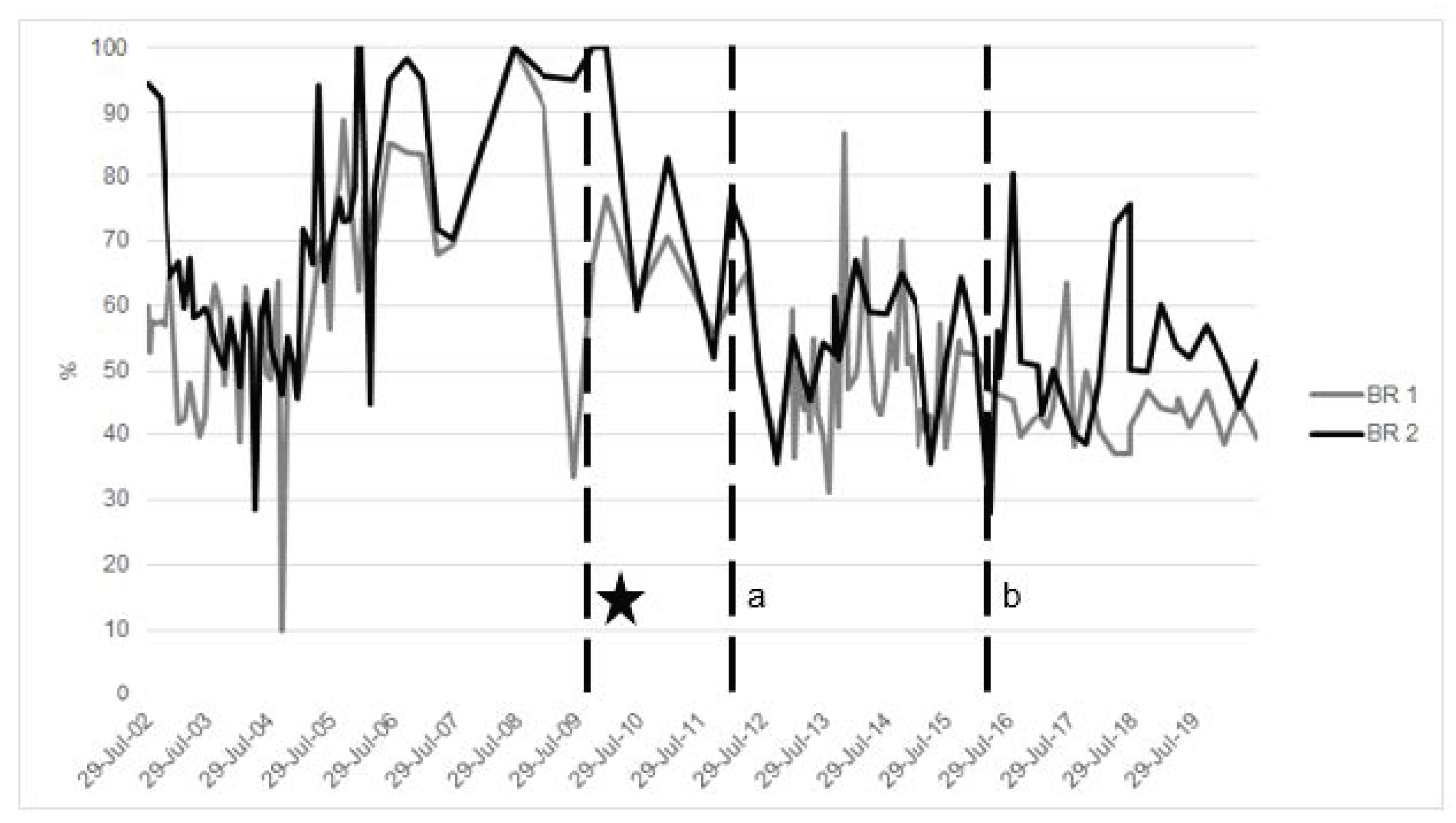
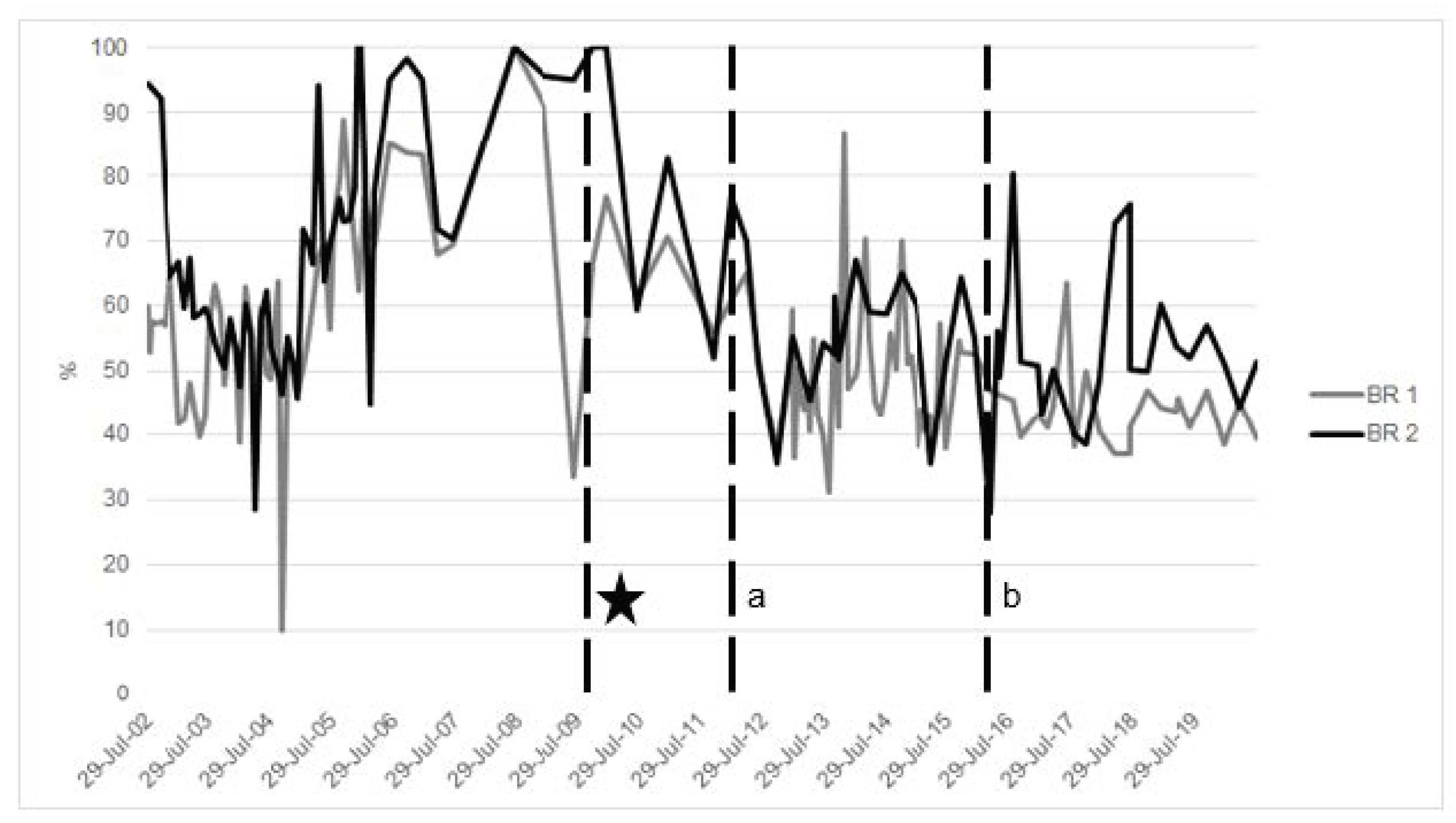
2.2.2. Total Iron-binding Capacity (TIBC) and Transferrin Saturation—Review and Comparative Species Evaluation
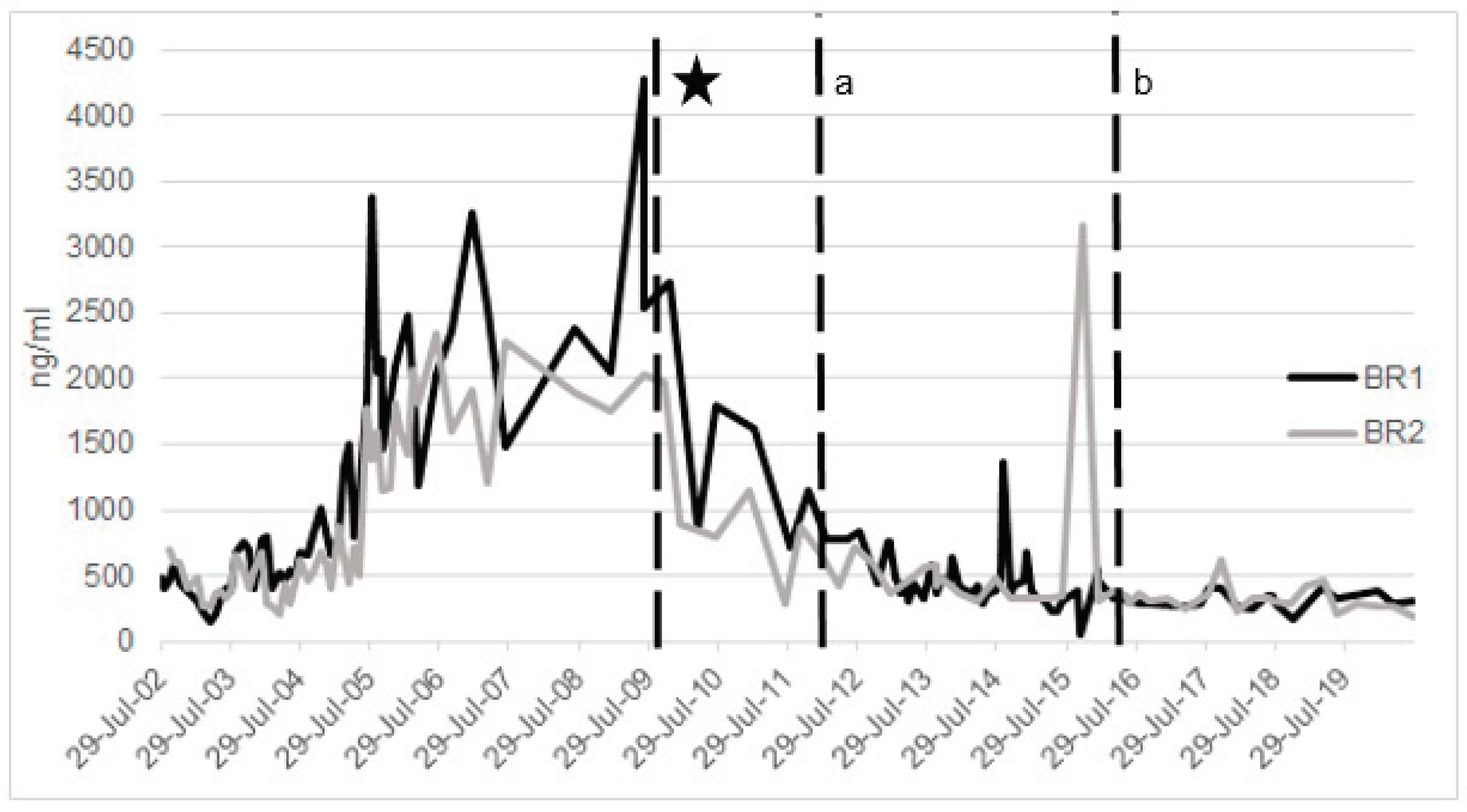
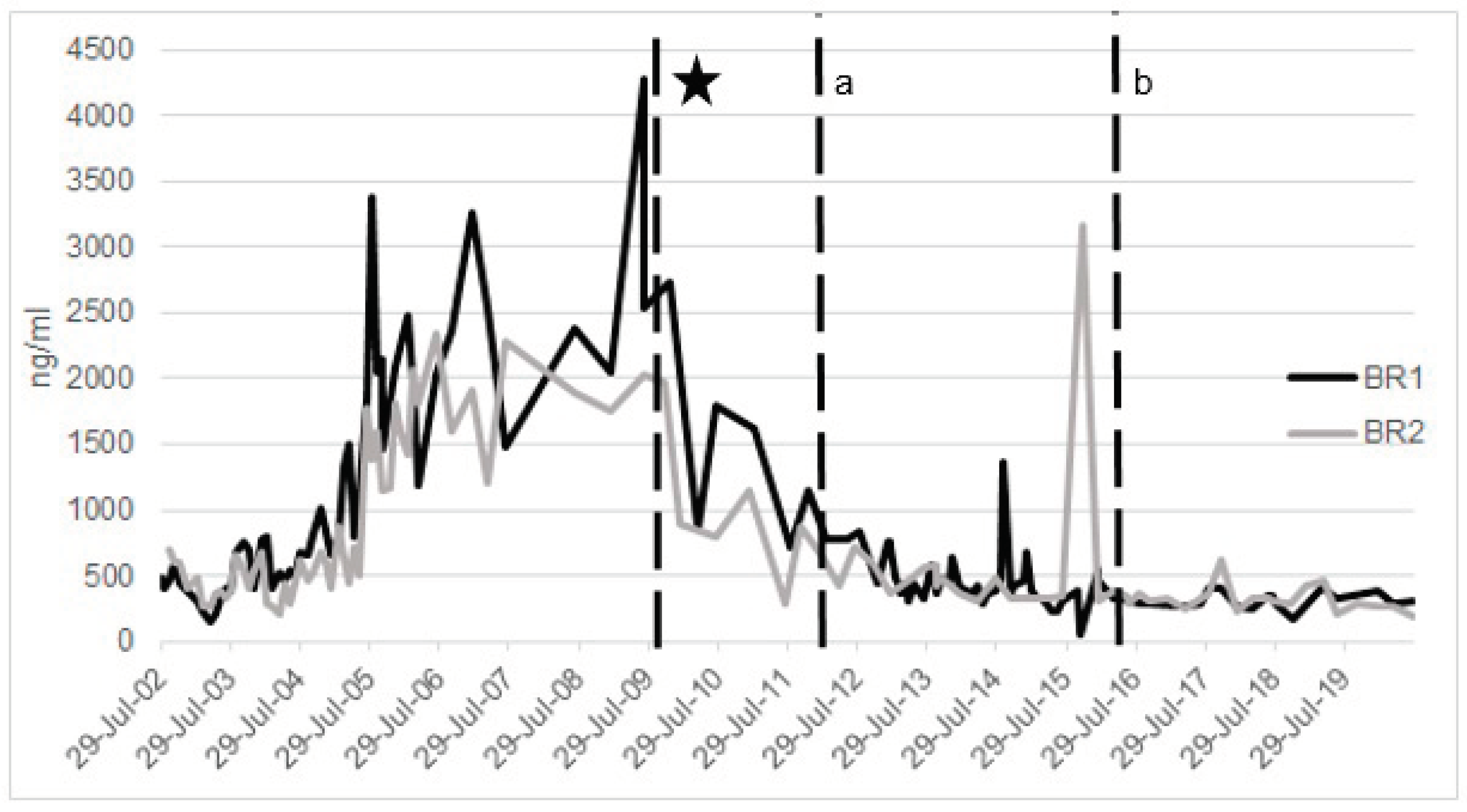
2.2.3. Ferritin
2.3. Perspectives on Integrative Monitoring
2.4. Inflammatory Biomarker Connections to IOD
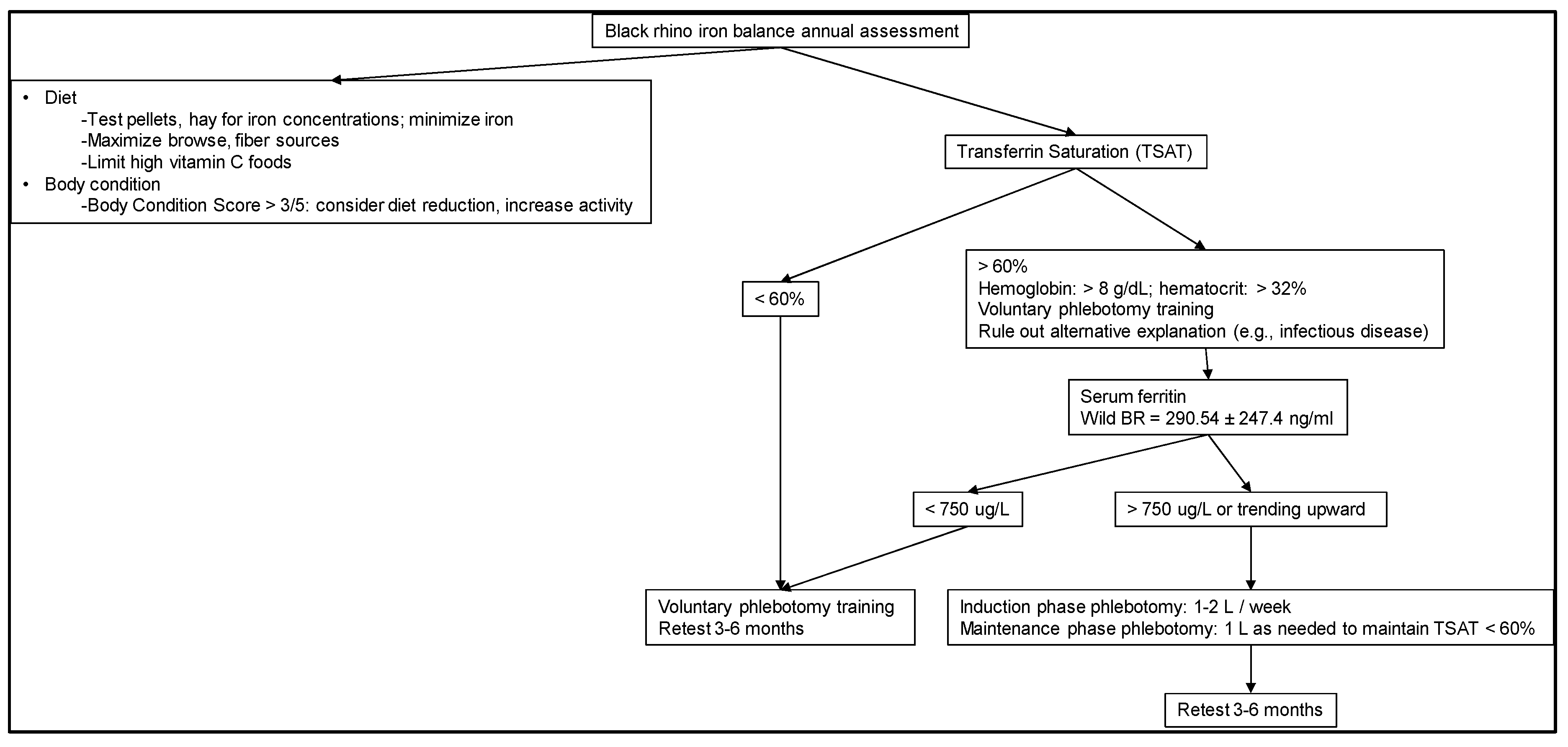
2.5. Recommendations for Diagnosis
3. Evidence-Based Nutrition Practices
3.1. Could We Make the Diet More Like the Wild?
3.2. Has Diet Ever Changed the Impact of IOD?
3.3. What Are the Best Practices for Feeding Black Rhinos?
- Iron should be limited. Iron concentration in the diet is recommended to not exceed 300 mg/kg dry matter (DM) or about 6 g of iron per day for a 1300 kg BR fed 1.5% BW in DM [5]. Based on the availability of feed items such as low-iron pelleted feed and browse (Table 1), a dietary iron concentration less than 300 mg/kg (dry matter basis) is a practical recommendation for BR [5,108]. Consider testing exhibit soil and vegetation to ensure they are not significant sources of iron in the diet of BRs [5,105].
- Monitor individual body weight and body condition. Feed 1–3% of body weight (BW) on an as fed basis, 1–2% on a dry matter basis. Maintaining appropriate body weight is critical as iron imbalance and obesity are presumed to be related; iron balance also is implicated with metabolic syndrome and associated negative health impacts [82]; however, the exact mechanism is not yet clear. Individual assessment of BR for bodyweight regularly across time (ideally weekly) and tracking with diet consumption is recommended [50,100]. Body condition scoring systems (e.g., a 1–5 scoring system based on ~seven body areas) are subjective, with varied recommendations on what is considered ideal depending on housing conditions along with medical and physiological considerations. Typically, a score of 3–4 out of a 5-point scale is considered ideal under human care. [109,110]. To optimize health, adjust diets as needed to ensure animals are within a target body weight range set for the individual animal.
- Feed at least twice daily. It is recommended to feed pellets in two feedings each day with forage to ensure maximum absorption of macronutrients [108]. Iron is not the only nutrient to consider in feeding complex diets to rhinos under human care. A single feeding would not be ideal for multiple reasons, including digestive efficiency, microbial community maintenance, satiety, and natural foraging behaviors.
- Feed appropriate pelleted feeds. The pellets milled for zoo animals vary widely in nutrient composition, and not all available pellets are appropriate for browsing species. Pellet formulations for BR are recommended to be high in fiber and low in starch and soluble sugars (Neutral Detergent Fiber (NDF) = 40–60%) [5]. Starch and sugar must be limited as these items can be associated with severe dental plaque, have metabolic impacts, and contribute to obesity [50,111,112]. A maximum of one-third of total calories is recommended to come from a pelleted concentrate. This limit avoids high pellet inclusion rates, which could be negative for dental health, body weight, and proper digestive health due to lack of long particle fibers [99].
- Alfalfa hay should be limited. This recommendation is due to its high protein, calcium, and iron, which can also create diarrhea and colic [5,50,108]. Conversely, low-quality hay (straw, wet/moldy, low nutrient content) is also not recommended due to the risk of intestinal impaction and/or colic. The iron in alfalfa is also held in a potentially highly bioavailable form (plant ferritin) [50].
- Maximize browse and provide access to hay. Preferably high quality roughage, ideally grass hay — not legume-based, as well as clean water and salt ad libitum [108]. Browse options may vary based on season and region, with options to freeze or ensile [110,113]. As browse best approximates the natural physical form of BR diets; it has the potential for iron-binding [114].
- Total dietary vitamin E concentrations should be 150–200 IU/kg diet. Extra supplementation may be necessary in addition to vitamin E in pelleted feed dependent on serum evaluation [17]. Vitamin E is a critical antioxidant that protects against ROS created by and including iron [50,115,116]. As BR lack some natural antioxidant production, ensuring dietary alpha-tocopherol (vitamin E) serves as a necessary preventive [50,115,116].
- Phosphorus levels in the serum should be monitored and supplemented where appropriate. BR have a predisposition to deficiency and continued concern for hemolytic issues; additionally, there is a link between phosphorus and iron metabolism [8,117]. Supplementation of monosodium phosphate and/or wheat bran in addition to phosphorus provided in a pelleted diet is recommended based on serum assessment. Naturally low phosphorus carriage in BR RBC (2–5% other mammals) [116] is thought to be connected to RBC fragility and potentially elevated RBC turnover [8,14,23]. In support of supplementation, higher levels of dietary phosphorus have been documented to combat anemic hemolytic crises in this species [8,23,117].
- The calcium to phosphorus (Ca:P) ratio of the diet should be 2:1 (no less than 1:1). A well-formulated pellet will provide appropriate calcium and phosphorus to meet the nutrient requirements of BR. An appropriate ratio eliminates the need for calcium supplementation, which can be contaminated with iron [102]. Grass hay typically is 1:1 and alfalfa 3:1, the latter of which can lead to hypercalcemia and hypophosphatemia. The amount of phosphorus added as a supplement should not unbalance the Ca:P ratio in the diet, in the amounts recommended based on body weight. The diet is balanced primarily with the pelleted portion, which is the majority of the dry matter of the diet and typically has the optimal 2:1 ratio (Table 1). Inverted serum Ca:P ratios are incredibly rare in rhinos; instead, hypercalcemia cases are far more common. As black rhinos physiologically appear to have an increased need for phosphorus, which is utilized for RBC turnover, they appear able to maintain serum Ca:P ratios of 2:1 despite a potential intake between 1:1 to 2:1.
- Avoid non-specific mineral supplements and mineral salt blocks. Plain salt blocks have minimal to no iron content and are appropriate [50].
- Training and enrichment diet items should be low in sugar, starch, and iron. Target less than 10% of the total diet comprising of training and enrichment foods. Take into consideration high-sugar, high-starch, and high-iron items (such as molasses-based foods), which often are included in balanced diets for BR (Table 1) [50].
4. Treatment and Prevention
4.1. Hematologic Sampling Recommendations
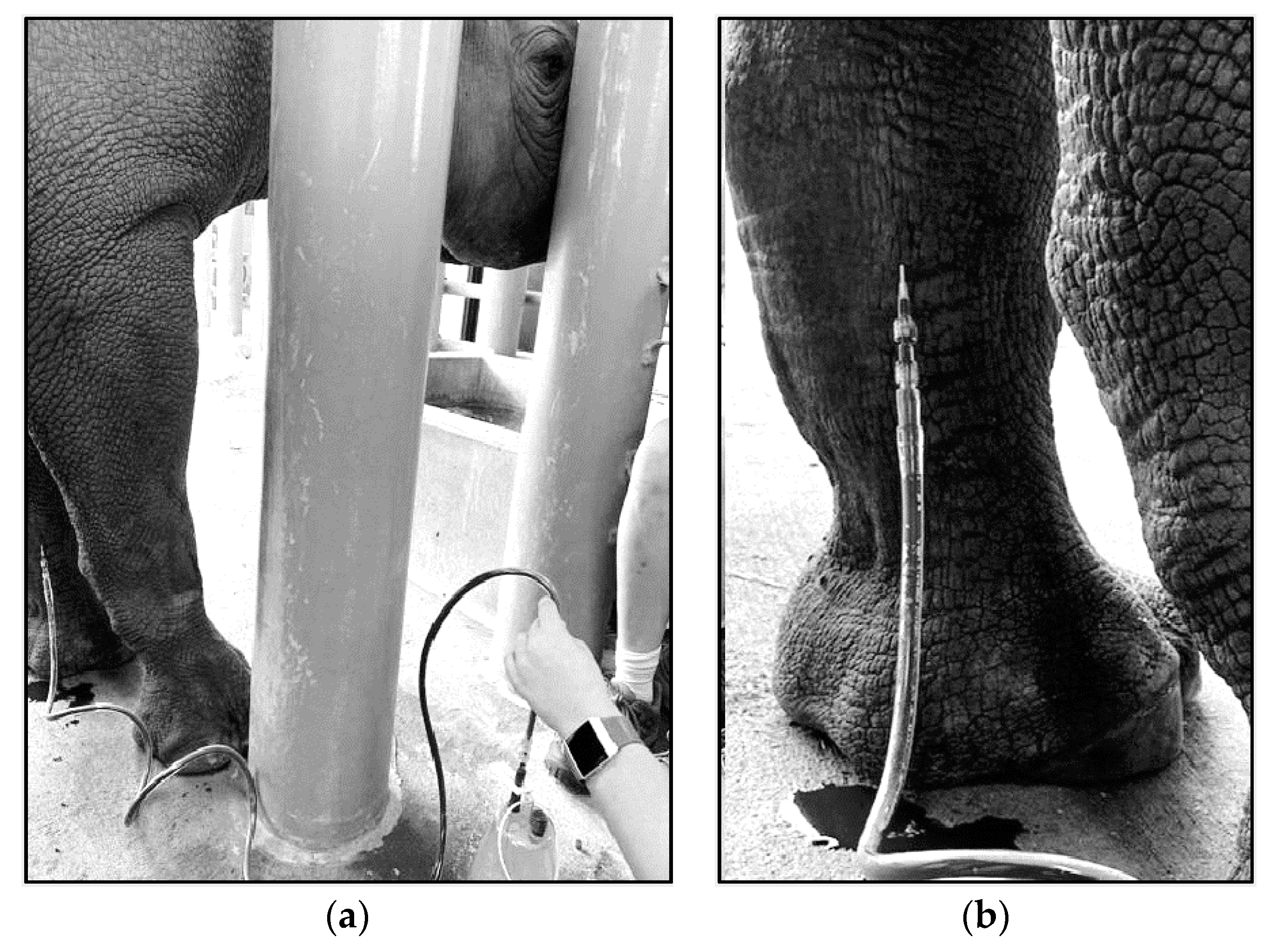
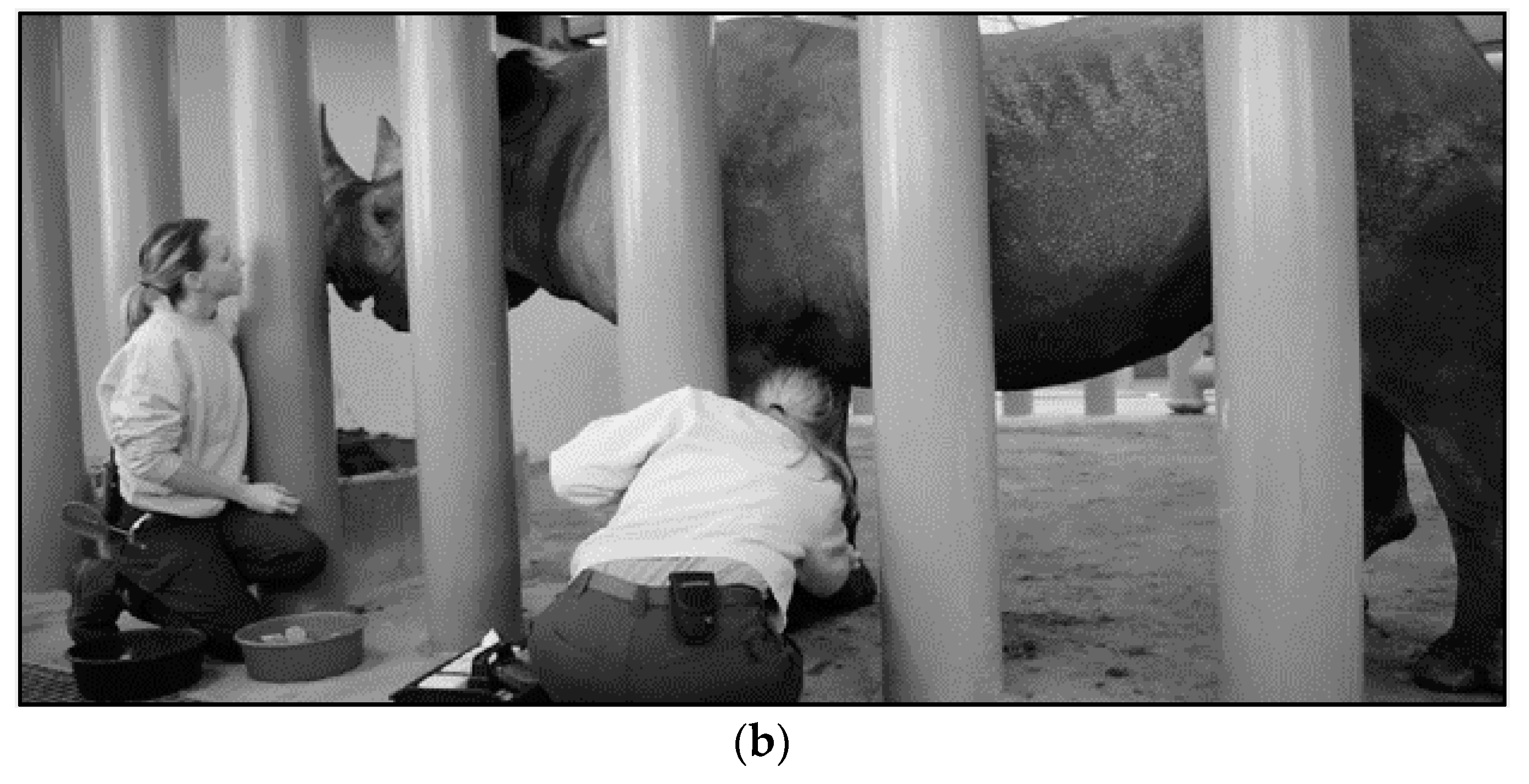
4.2. Therapeutic Large Volume Phlebotomy
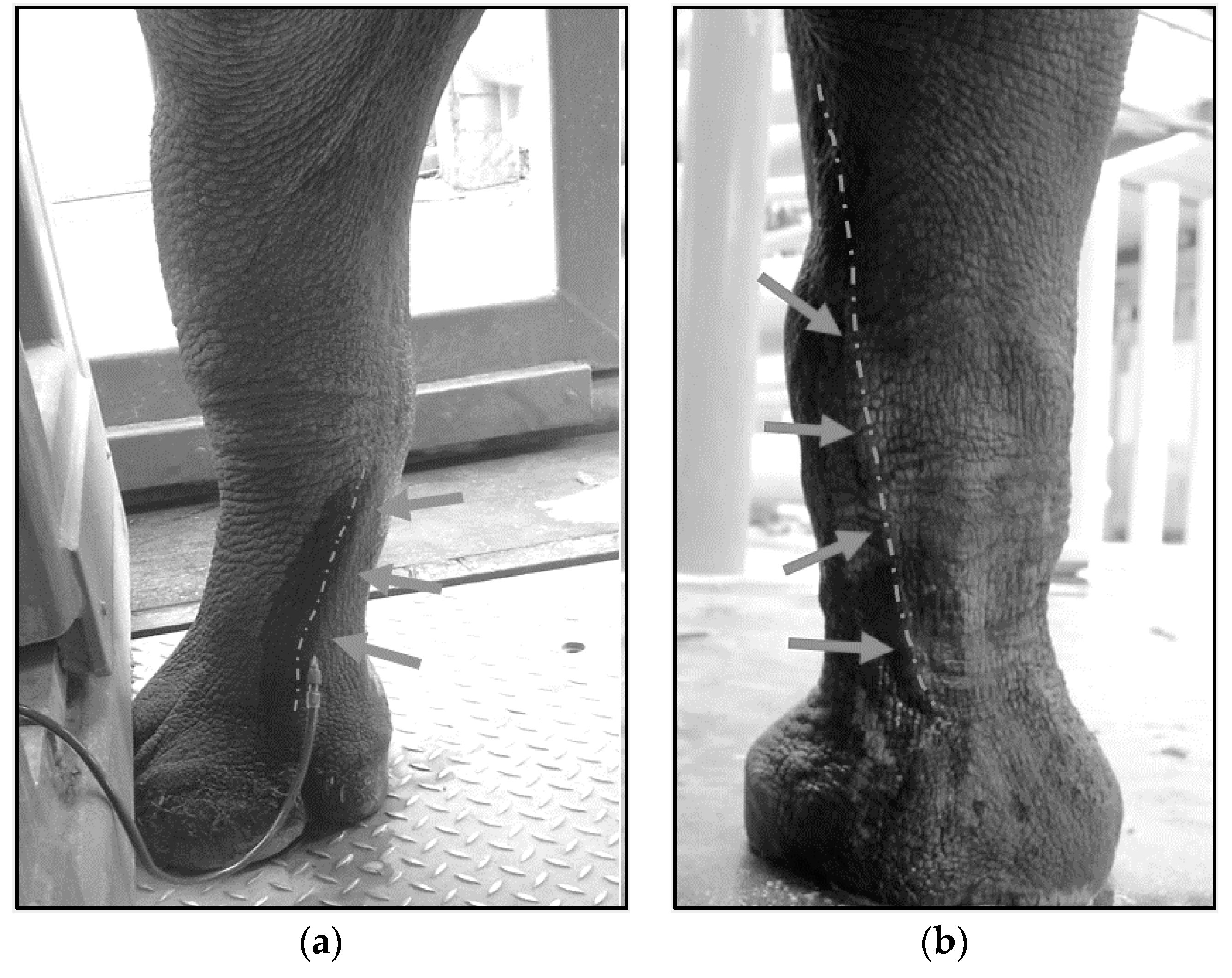
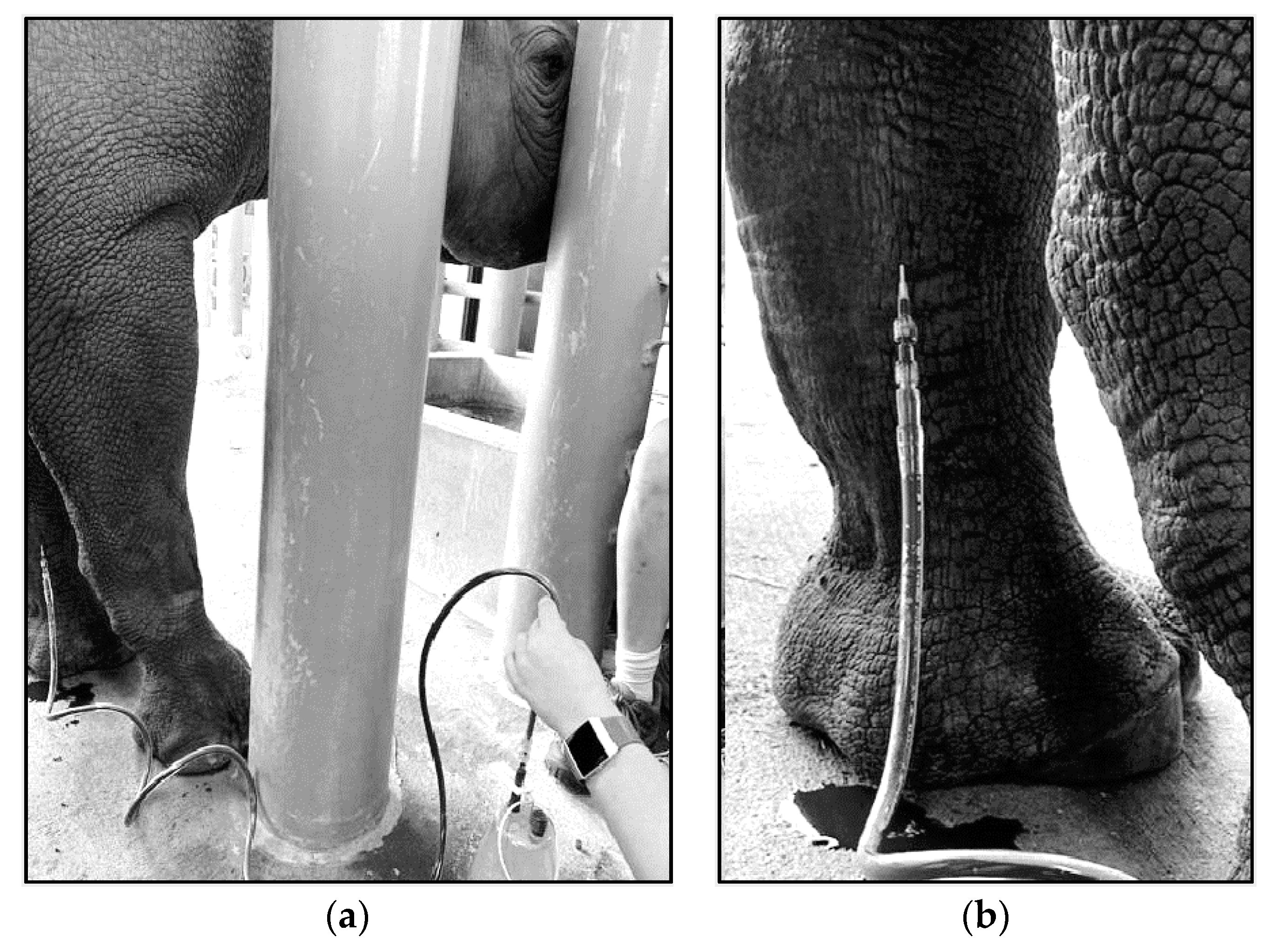
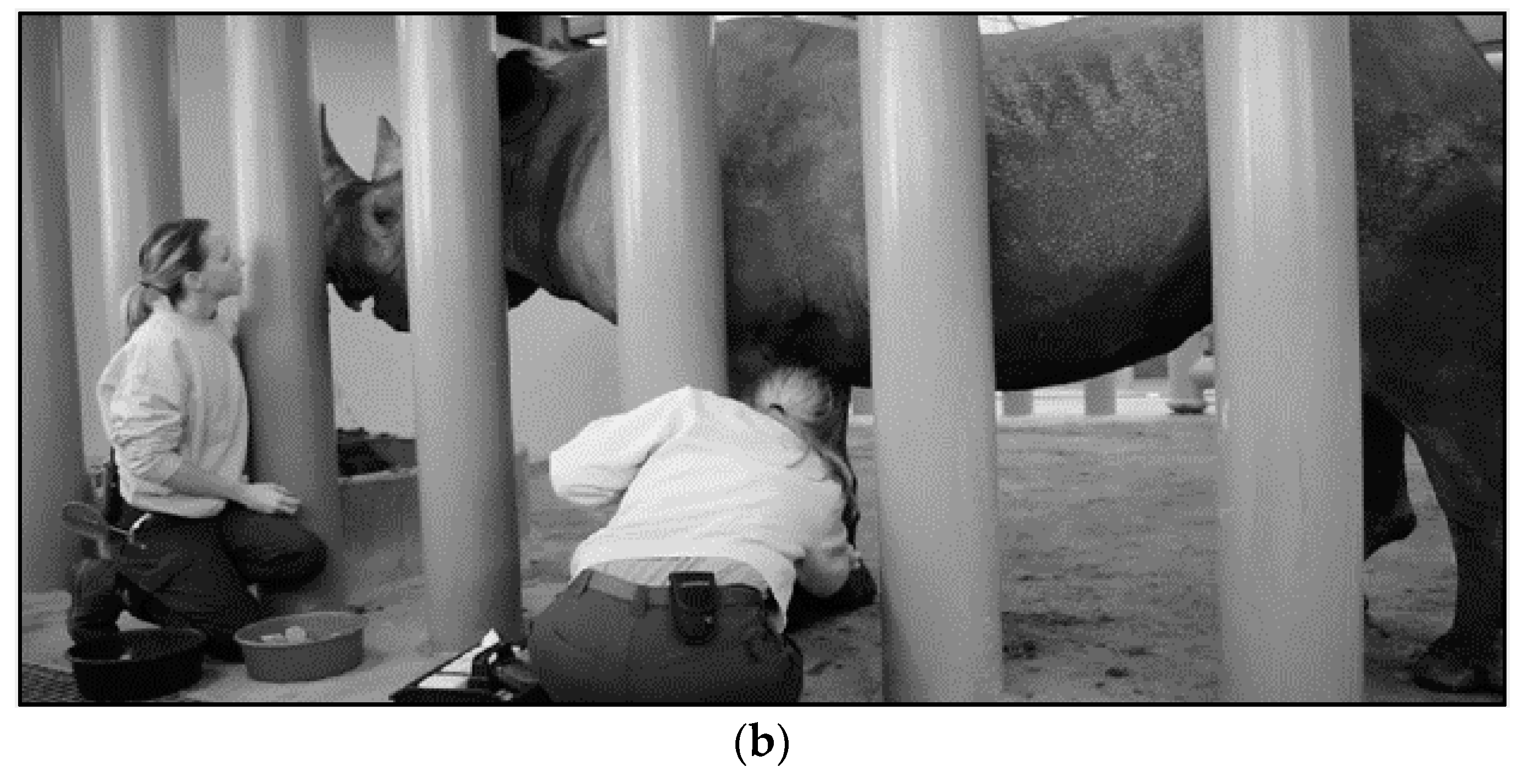
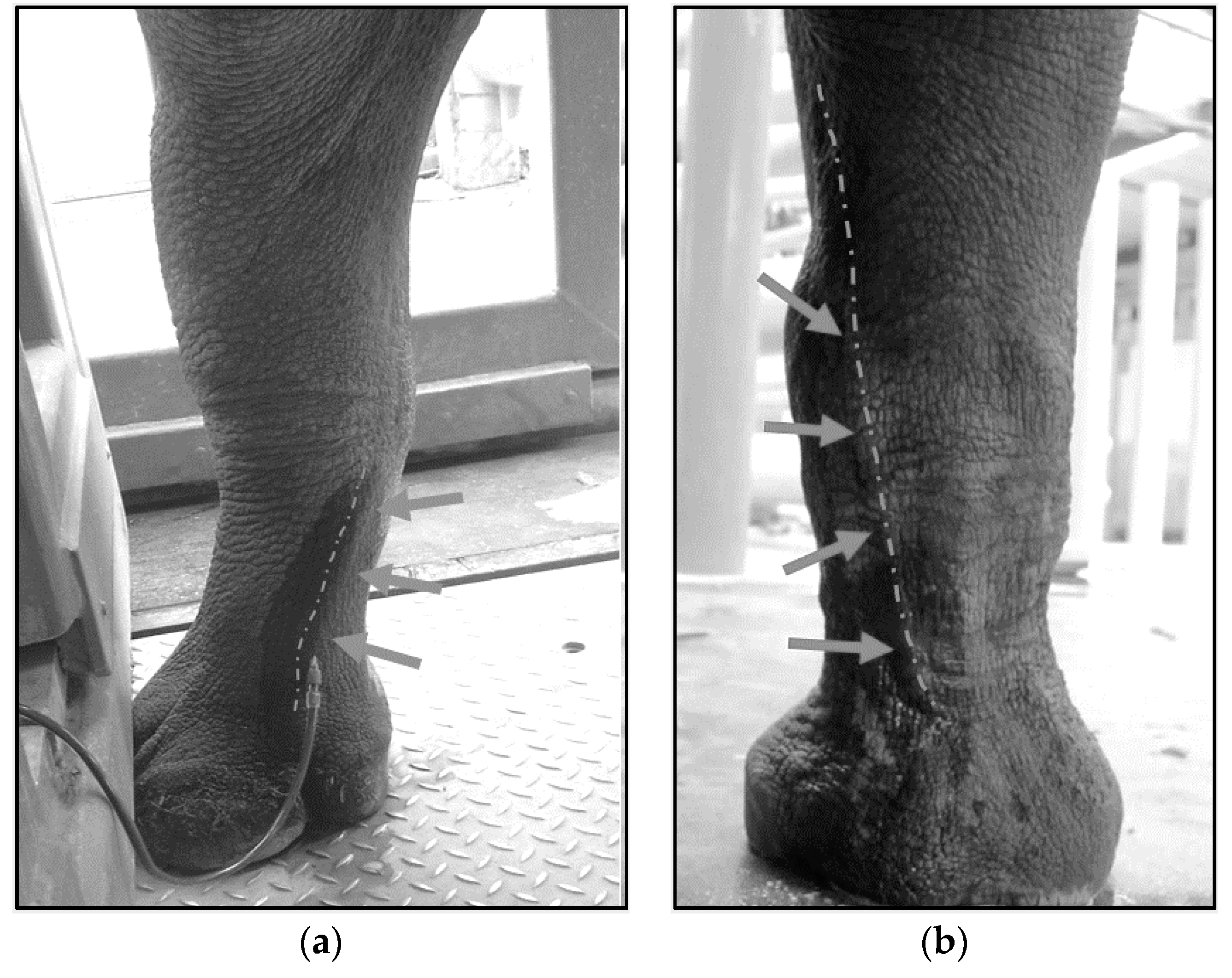
4.3. Techniques for VTLVP
4.4. Pharmacologic Chelation Therapy
4.5. History of Synthetic Chelation for BR
5. Animal Husbandry and Operant Conditioning Practices
6. Conclusions
- (1)
- Evaluate iron-related blood parameters to help understand baseline and then status over time and in relation to other health measures (Figure 2).
- (2)
- Focus attention on diet and nutrition, especially in terms of limiting iron.
- (3)
- Treat animals when indicated, preferably early in disease course; consider treatment options and guidance from experts on phlebotomy and potential chelation.
- (4)
- Invest time in training BR to allow for routine sampling as well as potential treatments. While it seems challenging, it is accomplishable without significant equipment or changes in stall design.
- (5)
- Balance the high cost of not assessing or managing IOD in BR with the low cost of using available tools to help guide treatments that can be very effective and prolong the life of the animal, improve reproductive success, and reduce disease issues, all of which have immeasurable costs.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Olias, P.; Mundhenk, L.; Bothe, M.; Ochs, A.; Gruber, A.; Klopfleisch, R. Iron Overload Syndrome in the Black Rhinoceros (Diceros bicornis): Microscopical Lesions and Comparison with Other Rhinoceros Species. J. Comp. Pathol. 2012, 147, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Paglia, D.E.; Tsu, I.-H. Review of laboratory and necropsy evidence for iron storage disease acquired by browser rhinoceroses. J. Zoo Wildl. Med. 2012, 43, S92–S104. [Google Scholar] [CrossRef] [PubMed]
- IUCN: International Union for Conservation of Nature. Available online: https://www.iucn.org/sites/dev/files/import/downloads/black_rhino_v3.pdf (accessed on 28 September 2020).
- Ferrie, G.M. AZA Regional Studbook Eastern Black Rhinoceros (Diceros bicornis Michaeli); AZA: Bay Lake, FL, USA, 2020. [Google Scholar]
- Clauss, M.; Dierenfeld, E.; Goff, J.; Klasing, K.; Koutsos, L.; Lavin, S.R.; Livingston, S.; Nielson, B.; Schlegel, M.; Sullivan, K.; et al. Iod in rhinos—Nutrition group report: Report from the nutrition working group of the international workshop on iron overload disorder in browsing rhinoceros (February 2011). J. Zoo Wildl. Med. 2012, 43. [Google Scholar] [CrossRef] [PubMed]
- Dennis, P.M.; Funk, J.A.; Rajala-Schultz, P.; Blumer, E.S.; Miller, R.E.; Wittum, T.E.; Saville, W.J.A. A review of some of the health issues of captive black rhinoceroses (Diceros bicornis). J. Zoo Wildl. Med. 2007, 38, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Andrews, N.C. Disorders of Iron Metabolism. N. Engl. J. Med. 1999, 341, 1986–1995. [Google Scholar] [CrossRef]
- Paglia, D.D.E. Human Medical Experience Provides Paradigms Relevant to Captive Breeding of Endangered Wildlife: Rationale for Prevention and Therapy of Hemolytic and Iron Overload Propensities in Browser Rhinoceroses, Tapirs and Other Susceptible Species; Aazv: Yule, FL, USA, 2017. [Google Scholar]
- Wood, J.C. Guidelines for quantifying iron overload. Hematology 2014, 2014, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Knutson, M.; Wessling-Resnick, M. Iron Metabolism in the Reticuloendothelial System. Crit. Rev. Biochem. Mol. Biol. 2003, 38, 61–88. [Google Scholar] [CrossRef]
- Helary, S.F.; Shaw, J.A.; Brown, D.; Clauss, M.; Owen-Smith, N. Black rhinoceros (Diceros bicornis) natural diets: Comparing iron levels across seasons and geographical locations. J. Zoo Wildl. Med. 2012, 43, S48–S54. [Google Scholar] [CrossRef]
- Beutler, E.; West, C.; Speir, J.A.; Wilson, I.A.; Worley, M. The HFE Gene of Browsing and Grazing Rhinoceroses: A Possible Site of Adaptation to a Low-Iron Diet. Blood Cells Mol. Dis. 2001, 27, 342–350. [Google Scholar] [CrossRef] [Green Version]
- Ganz, T.; Goff, J.; Klasing, K.; Nemeth, E.; Roth, T. Iod in rhinos—Immunity group report: Report from the immunity, genetics and toxicology working group of the international workshop on iron overload disorder in browsing rhinoceros (February 2011). J. Zoo Wildl. Med. 2012, 43, S117–S119. [Google Scholar] [CrossRef]
- Linzmeier, R.; Thompson, R.; Lamere, S.; Lee, P. Regulation of Iron Balance in Rhinoceroses. In Proceedings of the AAZV Annual Conference, Los Angeles, CA, USA, 11–17 October 2008; pp. 36–37. [Google Scholar]
- Kock, N.; Foggin, C.; Kock, M.D.; Kock, R. Hemosiderosis in the Black Rhinoceros (Diceros Bicornis): A Comparison of Free-Ranging and Recently Captured with Translocated and Captive Animals. J. Zoo Wildl. Med. 1992, 23, 230–234. [Google Scholar]
- Paglia, D.E.; Miller, R.E.; Renner, S.W. Is Impairment of Oxidant Neutralization the Common Denominator. In Proceedings of the Annual Conference-American Association of Zoo Veterinarians, Puerto Vallarta, Mexico, 3–8 November 1996; 1996; pp. 37–41. [Google Scholar]
- Dierenfeld, E.S.; Atkinson, S.; Craig, A.M.; Walker, K.C.; Clauss, M. Mineral concentrations in serum/plasma and liver tissue of captive and free-ranging Rhinoceros species. Zoo Biol. 2005, 24, 51–72. [Google Scholar] [CrossRef]
- Kontoghiorghe, K.C.; Kontoghiorghes, K.G. New developments and controversies in iron metabolism and iron chelation therapy. World J. Methodol. 2016, 6, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Kell, D.D.B. Iron behaving badly: Inappropriate iron chelation as a major contributor to the aetiology of vascular and other progressive inflammatory and degenerative diseases. BMC Med. Genom. 2009, 2, 2. [Google Scholar] [CrossRef]
- Kell, D.B.; Pretorius, E. No effects without causes: The Iron Dysregulation and Dormant Microbes hypothesis for chronic, inflammatory diseases. Biol. Rev. 2018, 93, 1518–1557. [Google Scholar] [CrossRef]
- Kontoghiorghes, G.J. New targeted therapies and diagnostic methods for iron overload diseases. Front. Biosci. 2018, 10, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Klopfleisch, R.; Olias, P. The Pathology of Comparative Animal Models of Human Haemochromatosis. J. Comp. Pathol. 2012, 147, 460–478. [Google Scholar] [CrossRef]
- Sullivan, K.E. Mitigating Iron Overload Disorder in Black Rhinoceros (Diceros bicornis); University of Florida: Gainesville, FL, USA, 2016. [Google Scholar]
- Ganz, T. Hepcidin and iron regulation, 10 years later. Blood 2011, 117, 4425–4433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paglia, D.E. Comparative Pathology of Iron-Storage Disorders in Captive Rhinoceroses: Potential Insights into Etiology and Pathogenesis. In Proceedings of the AAZV Annu. Conf., Knoxville, TN, USA, 20–26 October 2007; pp. 90–91. [Google Scholar]
- Saito, H. Metabolism of iron stores. Nagoya J. Med. Sci. 2014, 76, 235–254. [Google Scholar] [PubMed]
- Theil, E.C. Ferritin: The Protein Nanocage and Iron Biomineral in Health and in Disease. Inorg. Chem. 2013, 52, 12223–12233. [Google Scholar] [CrossRef] [PubMed]
- Theil, E.C. Ferritin and Its Role in Iron Homeostasis. Gas Sens. Cells 2014, 2, 358–380. [Google Scholar] [CrossRef]
- Doherty, C.P. Host-pathogen interactions: The role of iron. J. Nutr. 2007, 137, 1341–1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer-Fodor, E.; Miklášová, N.; Berindan-Neagoe, I.; Saha, B. Iron, inflammation and invasion of cancer cells. Med. Pharm. Rep. 2015, 88, 272–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer, E.; Le Gall-David, S.; Martin, B.; Fong, S.B.; Loréal, O.; Deugnier, Y.; Bonnaure-Mallet, M.; Meuric, V. Increased transferrin saturation is associated with subgingival microbiota dysbiosis and severe periodontitis in genetic haemochromatosis. Sci. Rep. 2018, 8, 15532. [Google Scholar] [CrossRef]
- Meuric, V.; Lainé, F.; Boyer, E.; Le Gall-David, S.; Oger, E.; Ebourgeois, D.; Bouchard, P.; Bardou-Jacquet, E.; Turmel, V.; Bonnaure-Mallet, M.; et al. Periodontal status and serum biomarker levels in HFE haemochromatosis patients. A case-series study. J. Clin. Periodontol. 2017, 44, 892–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, J.; Yamada, S.; Kajikawa, T.; Ozaki, N.; Awata, T.; Yamaba, S.; Fujihara, C.; Murakami, S. Iron plays a key role in the cytodifferentiation of human periodontal ligament cells. J. Period. Res. 2013, 49, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Facchini, F.S. Effect of Phlebotomy on Plasma Glucose and Insulin Concentrations. Diabetes Care 1998, 21, 2190. [Google Scholar] [CrossRef]
- Kordonouri, O. Diabetes-Perspektiven 2011: Dialog—Erfahrung—Gewinn. DMW Dtsch. Med. Wochenschr. 2011, 136, 1105. [Google Scholar] [CrossRef]
- Lee, D.W. Iron Dysregulation and Neurodegeneration: The Molecular Connection. Mol. Interv. 2006, 6, 89–97. [Google Scholar] [CrossRef]
- Paglia, D.E.; Kenny, D.E.; Dierenfeld, E.S.; Tsu, I.-H. Role of excessive maternal iron in the pathogenesis of congenital leukoencephalomalacia in captive black rhinoceroses (Diceros bicornis). Am. J. Veter. Res. 2001, 62, 343–349. [Google Scholar] [CrossRef]
- Jones, R.L.; Peterson, C.M.; Grady, R.W.; Kumbaraci, T.; Cerami, A.; Graziano, J.H. Effects of iron chelators and iron overload on Salmonella infection. Nat. Cell Biol. 1977, 267, 63–65. [Google Scholar] [CrossRef] [PubMed]
- Dimier, I.H.; Bout, D.T. Bout Interferon-γ-activated primary enterocytes inhibit Toxoplasma gondii replication: A role for intracellular iron. Immunology 1998, 94, 488–495. [Google Scholar] [CrossRef]
- Khalili, H.; Chan, S.S.M.; Lochhead, P.; Ananthakrishnan, A.N.; Hart, A.R.; Chan, A.T. The role of diet in the aetiopathogenesis of inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 525–535. [Google Scholar] [CrossRef] [Green Version]
- Wright, J.A.; Richards, T.; Srai, S.K.S. The role of iron in the skin and cutaneous wound healing. Front. Pharmacol. 2014, 5, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowenstine, L.J.; Stasiak, I.M. Update on Iron Overload in Zoologic Species. In Fowler’s Zoo and Wild Animal Medicine, Volume 8; Elsevier BV: Amsterdam, The Netherlands, 2015; Volume 8, pp. 674–681. [Google Scholar]
- Lowenstine, L.J.; Munson, L. Iron Overload in the Animal Kingdom. In Zoo and Wild Animal Medicine. Current Therapy, 4th ed.; Fowler, M.E., Miller, R.E., Eds.; W.B. Saunders Co.: Philadelphia, PA, USA, 1999; pp. 260–268. [Google Scholar]
- Molenaar, F.M.; Sainsbury, A.W.; Waters, M.; Amin, R.; Fall, T.; Holm, B.; Karlsson, Å.; Ahlgren, K.M.; Kämpe, O.; Von Euler, H. High serum concentrations of iron, transferrin saturation and gamma glutamyl transferase in captive black rhinoceroses (Diceros bicornis). Veter. Rec. 2008, 162, 716–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mylniczenko, N.D.; Sullivan, K.E.; Corcoran, M.E.; Fleming, G.J.; Valdes, E.V. Management strategies of iron accumulation in a captive population of black rhinoceroses (Diceros bicornis minor). J. Zoo Wildl. Med. 2012, 43, S83–S91. [Google Scholar] [CrossRef] [PubMed]
- Bohn, A.A. Diagnosis of Disorders of Iron Metabolism in Dogs and Cats. Veter. Clin. N. Am. Small Anim. Pr. 2013, 43, 1319–1330. [Google Scholar] [CrossRef]
- Miller, M.; Chavey, P.S.; Hofmeyr, J.; Mathebula, N.; Doering, A.; Buss, P.; Olea-Popelka, F. Evaluation of serum ferritin and serum iron in free-ranging black rhinoceros (Diceros bicornis) as a tool to understand factors affecting iron-overload disorder. J. Zoo Wildl. Med. 2016, 47, 820–826. [Google Scholar] [CrossRef] [PubMed]
- Kernan, K.F.; Carcillo, J.A. Hyperferritinemia and inflammation. Int. Immunol. 2017, 29, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Knovich, M.A.; Storey, J.A.; Coffman, L.G.; Torti, S.V.; Torti, F.M. Ferritin for the clinician. Blood Rev. 2009, 23, 95–104. [Google Scholar] [CrossRef]
- Sullivan, K.E.; Valdes, E.V. Update on Rhinoceros Nutrition. In Fowler’s Zoo and Wild Animal Medicine Current Therapy, Volume 9; Elsevier BV: Amsterdam, The Netherlands, 2019; Volume 9, pp. 699–706. [Google Scholar]
- Orino, K.; Lehman, L.; Tsuji, Y.; Ayaki, H.; Torti, S.V.; Torti, F.M. Ferritin and the response to oxidative stress. Biochem. J. 2001, 357, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J. Free Radicals and Antioxidant Protection: Mechanisms and Significance in Toxicology and Disease. Hum. Toxicol. 1988, 7, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Cloonan, S.M.; Mumby, S.; Adcock, I.M.; Choi, A.M.K.; Chung, K.F.; Quinlan, G.J. The “Iron”-y of Iron Overload and Iron Deficiency in Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2017, 196, 1103–1112. [Google Scholar] [CrossRef] [Green Version]
- Kushner, J.P.; Porter, J.P.; Olivieri, N.F. Secondary Iron Overload. Hematol. Ed. Prog. 2001, 2001, 47–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brasch, R.C.; Wesbey, G.E.; Gooding, C.A.; Koerper, M.A. Magnetic resonance imaging of transfusional hemosiderosis complicating thalassemia major. Radiololgy 1984, 150, 767–771. [Google Scholar] [CrossRef]
- Relia, N.; Kaushik, C. Renal hemosiderosis: A case of black kidneys causing renal failure. J. Postgrad. Med. 2010, 56, 216. [Google Scholar] [CrossRef]
- Roberts, W.C. Renal hemosiderosis (blue kidney) in patients with valvular heart disease. Am. J. Pathol. 1966, 48, 409–419. [Google Scholar]
- Bothwell, T.H.; MacPhail, A.P. Hereditary hemochromatosis: Etiologic, pathologic, and clinical aspects. Semin. Hematol. 1998, 35, 55–71. [Google Scholar]
- Levi, S.; Tiranti, V. Neurodegeneration with Brain Iron Accumulation Disorders: Valuable Models Aimed at Understanding the Pathogenesis of Iron Deposition. Pharmaceuticals 2019, 12, 27. [Google Scholar] [CrossRef] [Green Version]
- Spelman, L.H.; Osborn, K.G.; Anderson, M.P. Pathogenesis of hemosiderosis in lemurs: Role of dietary iron, tannin, and ascorbic acid. Zoo Biol. 1989, 8, 239–251. [Google Scholar] [CrossRef]
- Smith, K.M.; McAloose, D.; Raphael, B.L.; Calle, P.P.; Moore, R.P.; James, S.B.; Torregrossa, A.-M. Hematologic iron analyte values as an indicator of hepatic hemosiderosis in Callitrichidae. Am. J. Primatol. 2008, 70, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Klasing, K.C.; Dierenfeld, E.S.; Koutsos, E.A. Avian iron storage disease: Variations on a common theme? J. Zoo Wildl. Med. 2012, 43, S27–S34. [Google Scholar] [CrossRef] [PubMed]
- Farina, L.L.; Heard, D.J.; Leblanc, D.M.; Hall, J.O.; Stevens, G.; Wellehan, J.F.X.; Detrisac, C.J. Iron storage disease in captive Egyptian fruit bats (Rousettus aegyptiacus): Relationship of blood iron parameters to hepatic iron concentrations and hepatic histopathology. J. Zoo Wildl. Med. 2005, 36, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Mazzaro, L.M.; Johnson, S.P.; A Fair, P.; Bossart, G.; Carlin, K.P.; Jensen, E.D.; Smith, C.R.; Andrews, G.; Chavey, P.S.; Venn-Watson, S. Iron Indices in Bottlenose Dolphins (Tursiops truncatus). Comp. Med. 2012, 62, 508–515. [Google Scholar]
- Johnson, S.P.; Venn-Watson, S.; Cassle, S.E.; Smith, C.R.; Jensen, E.D.; Ridgway, S.H. Use of phlebotomy treatment in Atlantic bottlenose dolphins with iron overload. J. Am. Veter. Med. Assoc. 2009, 235, 194–200. [Google Scholar] [CrossRef] [Green Version]
- Cork, S.C. Iron storage diseases in birds. Avian Pathol. 2000, 29, 7–12. [Google Scholar] [CrossRef]
- DeHeer, H.L.; McManus, P. Frequency and severity of tracheal wash hemosiderosis and association with underlying disease in 96 cats: 2002–2003. Veter. Clin. Pathol. 2005, 34, 17–22. [Google Scholar] [CrossRef]
- Gambino, R.; Desvarieux, E.; Orth, M.; Matan, H.; Ackattupathil, T.; Lijoi, E.; Wimmer, C.; Bower, J.; Gunter, E. The relation between chemically measured total iron-binding capacity concentrations and immunologically measured transferrin concentrations in human serum. Clin. Chem. 1997, 43, 2408–2412. [Google Scholar] [CrossRef]
- Harvey, J.W. Evaluation of Erythrocytes. Vet. Hematol. 2012, 49–121. [Google Scholar] [CrossRef]
- Pouillevet, H.; Soetart, N.; Boucher, D.; Wedlarski, R.; Jaillardon, L. Inflammatory and oxidative status in European captive black rhinoceroses: A link with Iron Overload Disorder? PLoS ONE 2020, 15, e0231514. [Google Scholar] [CrossRef]
- Elsayed, M.; Sharif, M.; Stack, A. Transferrin Saturation. Internat. Rev. Cytol. 2016, 75, 71–97. [Google Scholar] [CrossRef]
- Yamanishi, H.; Iyama, S.; Yamaguchi, Y.; Kanakura, Y.; Iwatani, Y. Total Iron-binding Capacity Calculated from Serum Transferrin Concentration or Serum Iron Concentration and Unsaturated Iron-binding Capacity. Clin. Chem. 2003, 49, 175–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huebers, H.A.; Eng, M.J.; Josephson, B.M.; Ekpoom, N.; Rettmer, R.L.; Labbé, R.F.; Pootrakul, P.; Finch, C.A. Plasma iron and transferrin iron-binding capacity evaluated by colorimetric and immunoprecipitation methods. Clin. Chem. 1987, 33, 273–277. [Google Scholar] [CrossRef]
- Anderson, G.J. Non-transferrin-bound iron and cellular toxicity. J. Gastroenterol. Hepatol. 2002, 14, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Bacon, B.R.; Adams, P.C.; Kowdley, K.V.; Powell, L.W.; Tavill, A.S. Diagnosis and management of hemochromatosis: 2011 Practice Guideline by the American Association for the Study of Liver Diseases. Hepatology 2011, 54, 328–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piperno, A. Classification and diagnosis of iron overload. Haematologica 1998, 83, 447–455. [Google Scholar]
- Kell, D.B.; Pretorius, E. Serum ferritin is an important inflammatory disease marker, as it is mainly a leakage product from damaged cells. Metallomics 2014, 6, 748–773. [Google Scholar] [CrossRef] [Green Version]
- Hintze, K.J.; Theil, E.C. DNA and mRNA elements with complementary responses to hemin, antioxidant inducers, and iron control ferritin-L expression. Proc. Natl. Acad. Sci. USA 2005, 102, 15048–15052. [Google Scholar] [CrossRef] [Green Version]
- Muckenthaler, M.U.; Galy, B.; Hentze, M.W. Systemic Iron Homeostasis and the Iron-Responsive Element/Iron-Regulatory Protein (IRE/IRP) Regulatory Network. Annu. Rev. Nutr. 2008, 28, 197–213. [Google Scholar] [CrossRef]
- Serraj, K.; Alaoui, H.; Hamaz, S.; Mourot-Cottet, R.; Keller, O.; Andrès, E. Démarche Diagnostique Devant Une Hyperferritinémie. Méd. Thérapeut. 2015, 21, 298–304. [Google Scholar]
- Smith, J.E.; Chavey, P.S.; Miller, R.E. Iron Metabolism in Captive Black (Diceros bicornis) and White (Ceratotherium simum) Rhinoceroses. J. Zoo Wildl. Med. 2009, 26, 525–531. [Google Scholar]
- Schook, M.W.; Wildt, D.E.; Raghanti, M.A.; Wolfe, B.A.; Dennis, P.M. Increased inflammation and decreased insulin sensitivity indicate metabolic disturbances in zoo-managed compared to free-ranging black rhinoceros (Diceros bicornis). Gen. Comp. Endocrinol. 2015, 217–218, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Orino, K.; Miura, T.; Muto, S.; Watanabe, K. Sequence analysis of canine and equine ferritin H and L subunit cDNAs. DNA Seq. 2005, 16, 58–64. [Google Scholar] [CrossRef]
- Wojtusik, J.; Roth, T.L. Investigation of factors potentially associated with serum ferritin concentrations in the black rhinoceros (Diceros bicornis) using a validated rhinoceros-specific assay. J. Zoo Wildl. Med. 2018, 49, 297–306. [Google Scholar] [CrossRef]
- Roth, T.L.; Reinhart, P.R.; Kroll, J.L. Serum ferritin concentration is not a reliable biomarker of iron overload disorder progression or hemochromatosis in the sumatran rhinoceros (Dicerorhinus sumatrensis). J. Zoo Wildl. Med. 2017, 48, 645–658. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, E.; Ganz, T. The role of hepcidin in iron metabolism. Acta Haematol. 2009, 122, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Roth, T.L.; Wojtusik, J.; (Center for Conservation and Research of Endangered Wildlife (CREW), Cincinnati, OH, USA). Summary of Recent Iron Studies, Cincinnati Zoo and Botanical Garden. Personal Communication, 2017. [Google Scholar]
- Fredholm, D.V.; (Disney’s Animal Kingdom, Bay Lake, FL, USA). Personal Communication, 2016.
- Roth, T.L.; Switzer, A.; Watanabe-Chailland, M.; Bik, E.M.; Relman, D.A.; Romick-Rosendale, L.E.; Ollberding, N.J. Reduced Gut Microbiome Diversity and Metabolome Differences in Rhinoceros Species at Risk for Iron Overload Disorder. Front. Microbiol. 2019, 10, 2291. [Google Scholar] [CrossRef]
- Roth, T.L. The Quest for a Reliable Biomarker of Iron Overload Disorder in the Rhinoceros. In Proceedings of the AAZV Annual Conference, St. Louis, MO, USA, 28 September–4 October 2019; pp. 163–164. [Google Scholar]
- D’Assignies, G.; Paisant, A.; Bardou-Jacquet, E.; Boulic, A.; Bannier, E.; Lainé, F.; Ropert, M.; Morcet, J.; Saint-Jalmes, H.; Gandon, Y. Non-invasive measurement of liver iron concentration using 3-Tesla magnetic resonance imaging: Validation against biopsy. Eur. Radiol. 2017, 28, 2022–2030. [Google Scholar] [CrossRef]
- Fredholm, D.V.; Shmalberg, J.; Scott, K.C.; Mylniczenko, N.D. A Comparison of Oxidative Stress Markers and Antioxidant Status in Two Species of Rhinoceros, Diceros bicornis and Ceratotherium simum. In Proceedings of the AAZV Annual Conference, Atlanta, GA, USA, 16–22 July 2016; pp. 118–119. [Google Scholar]
- Cray, C. Reference Intervals in Avian and Exotic Hematology. Veter. Clin. N. Am. Exot. Anim. Pr. 2015, 18, 105–116. [Google Scholar] [CrossRef]
- Genova, J.H.M. Serum Proteins Electrophoresis by Agarose Gel- M-Spike Screening and Beyond—Review. Int. J. Sci. Res. 2017, 6, 1463–1466. [Google Scholar] [CrossRef]
- Gevanthor, J.F.; Tatum, L.M.; Deem, S.L.; Citino, S.B. Preliminary Evaluation of Serum Proteinelectrophoresis as a Diagnostic Tool in the Black Rhinoceros (Diceros bicornis). In Proceedings of the AAZV AAWV Joint Conference, Ohama, NE, USA, 17–22 October 1998; pp. 124–127. [Google Scholar]
- Camiolo, G.; Tibullo, D.; Giallongo, C.; Romano, A.; Parrinello, N.L.; Musumeci, G.; Tibullo, D.; Vicario, N.; Brundo, M.V.; Amenta, F.; et al. α-Lipoic Acid Reduces Iron-induced Toxicity and Oxidative Stress in a Model of Iron Overload. Int. J. Mol. Sci. 2019, 20, 609. [Google Scholar] [CrossRef] [Green Version]
- Ali, Y.F.; Desouky, O.S.; Selim, N.S.; Ereiba, K.M. Assessment of the role of α-lipoic acid against the oxidative stress of induced iron overload. J. Radiat. Res. Appl. Sci. 2015, 8, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Dierenfeld, E.S.; Wildman, R.E.C.; Steve, R.; Dierenfeld, E.S.; Wildman, R.E.C.; Romo, S. Feed Intake, Diet Utilization, and Composition of Browses Consumed by the Sumatran Rhino (Dicerorhinus sumatrensis) in a North American Zoo. Zoo Biol. 2000, 19, 169–180. [Google Scholar] [CrossRef]
- Van Soest, P.J. Nutritional Ecology of the Ruminant; Cornell University Press: Ithaca, NY, USA, 1994; pp. 140–155. [Google Scholar] [CrossRef]
- Clauss, M.; Hatt, J. The feeding of rhinoceros in captivity. Int. Zoo Yearb. 2006, 40, 197–209. [Google Scholar] [CrossRef]
- Dairy One. Interactive Feed Composition Libraries. Available online: https://dairyone.com/services/forage-laboratory-services/feed-composition-library/interactive-feed-composition-libraries/ (accessed on 28 October 2020).
- Koutsos, L.; Clauss, M.; Valdes, E. Designing Iron Controlled Diets for Exotic Hoofstock—Variability in Raw Materials and Manufacturing Contributions to Total Dietary Iron. In Proceedings of the Eleventh Symposia of the Comparative Nutrition Society, Rio del Mar, Puerto Rico, 1–4 August 2016; pp. 77–80. [Google Scholar]
- Adams, R. Variability in Mineral and Trace Element Content of Dairy Cattle Feeds. J. Dairy Sci. 1975, 58, 1538–1548. [Google Scholar] [CrossRef]
- Berger, L.L. Variation in the Trace Mineral Content of Feedstuffs1. Prof. Anim. Sci. 1996, 12, 1–5. [Google Scholar] [CrossRef]
- Mimiko, J.; Stringer, E.P.J. Case Study: Iron in Black Rhinoceros Diets: The Impact of Pasture. In Proceedings of the Twelfth Conference on Zoo and Wildlife Nutrition, Zoo and Wildlife Nutrition Foundation and AZA Nutrition Advisory Group, Frisco, TX, USA, 23–29 September 2017. [Google Scholar]
- Ward, A.M.; Hunt, A.S. Summary of Mineral and Iron Binding Polyphenolic Plant Compound Levels in Diets Offered Captive Black Rhinoceros (Diceros bicornis) in 3 Zoos and 1 Ranch in Texas. In Proceedings of the 4th Nutrition Advisory Group Conference on Zoo and Wildlife Nutrition, Bay Lake, FL, USA, 18–23 September 2001; pp. 173–186. [Google Scholar]
- Clauss, M.; Castell, J.C.; Kienzle, E.; Dierenfeld, E.S.; Flach, E.J.; Behlert, O.; Ortmann, S.; Streich, W.J.; Hummel, J.; Hatt, J.-M. The influence of dietary tannin supplementation on digestive performance in captive black rhinoceros (Diceros bicornis). J. Anim. Physiol. Anim. Nutr. 2007, 91, 449–458. [Google Scholar] [CrossRef]
- Metrione, L.; Eyres, A. Rhino Husbandry Manual International Rhino Foundation; International Rhino Foundation: Fort Worth, TX, USA, 2014. [Google Scholar]
- Reuter, H.; Adcock, K. Standardised Body Condition Scoring System for Black Rhinoceros (Diceros bicornis). Pachyderm 1998, 26, 116–121. [Google Scholar]
- Pilgrim, M.; Biddle, R. Best Practice Guidelines Black Rhinoceros (Diceros bicornis), 2nd ed.; The European Association of Zoos and Aquaria (EAZA): Amsterdam, The Netherlands, 2020; pp. 35–41. [Google Scholar]
- Vervuert, I.; Klein, S.; Coenen, M. Effects of feeding state on glycaemic and insulinaemic responses to a starchy meal in horses: A methodological approach. Animal 2009, 3, 1246–1253. [Google Scholar] [CrossRef]
- Vervuert, I.; Voigt, K.; Hollands, T.; Cuddeford, D.; Coenen, M. Effect of feeding increasing quantities of starch on glycaemic and insulinaemic responses in healthy horses. Veter. J. 2009, 182, 67–72. [Google Scholar] [CrossRef]
- Sullivan, K.E.S.; Lavin, S.; Livingston, E.V.V. Palatability of Bunker Ensiled Willow as a Winter Diet Item for Browsing Herbivores at Disney’s Animal Kingdom. In Proceedings of the Nutrition Advisory Group to the AZA 9th bi- Annual Conference, Kansas City, MO, USA, 22–28 October 2011. [Google Scholar]
- Lavin, S.R. Plant phenolics and their potential role in mitigating iron overload disorder in wild animals. J. Zoo Wild. Med. 2012, 43, S74–S82. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, J.; Packer, L. Vitamin E in health and disease. Free. Radic. Biol. Med. 1993, 15, 109. [Google Scholar] [CrossRef]
- Harley, E.; Paglia, D.; Weber, B. Oxidative damage and purine metabolism: Investigation of haemolytic anaemia in the black rhinoceros. Clin. Biochem. 1997, 30, 259. [Google Scholar] [CrossRef]
- Paglia, D.E. Acute episodic hemolysis in the African black rhinoceros as an analogue of human glucose-6-phosphate dehydrogenase deficiency. Am. J. Hematol. 1993, 42, 36–45. [Google Scholar] [CrossRef]
- Cook, J.D.; Reddy, M.B. Effect of ascorbic acid intake on nonheme-iron absorption from a complete diet. Am. J. Clin. Nutr. 2001, 73, 93–98. [Google Scholar] [CrossRef]
- Paglia, D. Recommended Phlebotomy Guidelines for Prevention and Therapy of Captivity-Induced Iron-Storage Disease. In Proceedings of the AAZV Annual Conference, San Diego, CA, USA, 28 August–3 September 2004; pp. 122–127. [Google Scholar]
- Oh, K.Y.; Kim, K.-H. Clinical applications of therapeutic phlebotomy. J. Blood Med. 2016, 7, 139–144. [Google Scholar] [CrossRef] [Green Version]
- Abdulmalek, A.A.; Abdulrahman, I.A.E.; Tariek, A.A.; Ali, S.A.; Khalid, M.M.; Sultan, M.A.; Mansour, T.A.; Kawther, A.A.; Thamer, H.M.A.; Fatheya, A.H. Evaluation of Hemochromatosis, Diagnosis and Management: Simple Literature Review. Arch. Pharm. Pract. 2020, 11, 101–103. [Google Scholar]
- Ginzburg, Y.Z.; Vinchi, F. Therapeutic Phlebotomy. In Transfusion Medicine and Hemostasis; Elsevier BV: Amsterdam, The Netherlands, 2019; pp. 501–503. [Google Scholar]
- Chikazawa, S.; Hori, Y.; Hoshi, F.; Kanai, K.; Ito, N.; Sato, J.; Orino, K.; Watanabe, K.; Higuchi, S.-I. Development of a sandwich enzyme-linked immunosorbent assay to detect and measure serum levels of canine ferritin. J. Veter. Med Sci. 2012, 75, 515–517. [Google Scholar] [CrossRef] [Green Version]
- Stringer, B.; Bapodra, P.; (Columbus Zoo and Aquarium, Powell, OH, USA). Personal Communication, 2020.
- Jacokes, C. Adapting Large Volume Phlebotomy to Denver Zoo’s Black Rhinoceros. In Proceedings of the 45th Annual American Association of Zookeepers National Conference, Denver, CO, USA, 4–8 October 2018. [Google Scholar]
- Rhino Resource Center 2007 Newsletter. Available online: http://www.rhinoresourcecenter.com/pdf_files/118/1187007151.pdf (accessed on 28 September 2020).
- Nelson, S.M.N. Voluntary Therapeutic Large Volume Phlebotomy of Ex-Situ Black Rhinoceroses (Diceros bicornis) For the Treatment and Prevention of Iron Overload Disorder. In Proceedings of the Association of Zoo Veterinary Technicians Annual Conference, Colorado Springs, CO, USA, 23–26 September 2019. [Google Scholar]
- Liu, Z.D.; Hider, R.C. Design of iron chelators with therapeutic application. Co-ord. Chem. Rev. 2002, 232, 151–171. [Google Scholar] [CrossRef]
- Bergeron, R.J.; Wiegand, J.; Brittenham, G.M. HBED: The Continuing Development of a Potential Alternative to Deferoxamine for Iron-Chelating Therapy. Blood 1999, 93, 370–375. [Google Scholar] [CrossRef]
- Dean, R.T.; Nicholson, P. The Action of Nine Chelators on Iron-Dependent Radical Damage. Free. Radic. Res. 1994, 20, 83–101. [Google Scholar] [CrossRef]
- Singh, S.; Choudhury, R.; Hider, R.C.; Porter, J.B. Design and Development of Orally Active Chelating Agents: Selective Targeting of Chelatable Iron Pools. In Proceedings of the International Conference “New Trends in Therapy for Hemoglobinopathies and Thalassaemias”, Paris, France, 19–22 September 1994. [Google Scholar]
- Crisponi, G.; Remelli, M. Iron chelating agents for the treatment of iron overload. Co-ord. Chem. Rev. 2008, 252, 1225–1240. [Google Scholar] [CrossRef]
- Hershko, C.; Grady, R.W.; Cerami, A. Mechanism of iron chelation in the hypertransfused rat: Definition of two alternative pathways of iron mobilization. J. Lab. Clin. Med. 1978, 92, 144–151. [Google Scholar] [PubMed]
- Hershko, C.; Link, G.; Cabantchik, I. Pathophysiology of Iron Overload. Ann. N. Y. Acad. Sci. 1998, 850, 191–201. [Google Scholar] [CrossRef]
- Hershko, C.; Grady, R.W.; Link, G. Development and evaluation of the improved iron chelating agents EHPG, HBED and their dimethyl esters. Haematology 1984, 17, 25–33. [Google Scholar]
- Hider, R.C.; Epemolu, O.; Singh, S.; Porter, J.B. Iron Chelator Design. Adv. Experim. Med. Biol. 1994, 356, 343–349. [Google Scholar] [CrossRef]
- Brooks, M.; Lee, B.P.J. Case Study: Winos for Rhinos: Feeding Grape Pomace to Black Rhinoceros (Diceros bicornis) as a Method for Mitigating Iron Storage Disease. In Proceedings of the Twelfth Conference on Zoo and Wildlife Nutrition, Zoo and Wildlife Nutrition Foundation and AZA Nutrition Advisory Group, Frisco, TX, USA, 23–29 September 2017. [Google Scholar]
- Huntley, N.F.; Naumann, H.D.; Kenny, A.L.; Kerley, M.S. Black rhinoceros (Diceros bicornis) and domestic horse (Equus caballus) hindgut microflora demonstrate similar fermentation responses to grape seed extract supplementationin vitro. J. Anim. Physiol. Anim. Nutr. 2016, 101, e195–e209. [Google Scholar] [CrossRef] [PubMed]
- Lavin, S.R.; Sullivan, K.E.; Wooley, S.C.; Robinson, R.; Singh, S.; Stone, K.; Russell, S.; Valdes, E.V. Nutrient and plant secondary compound composition and iron-binding capacity in leaves and green stems of commonly used plant browse (Carolina willow; Salix caroliniana) fed to zoo-managed browsing herbivores. Zoo Biol. 2015, 34, 565–575. [Google Scholar] [CrossRef]
- Hurrell, R.F.; Reddy, M.; Cook, J.D. Inhibition of non-haem iron absorption in man by polyphenolic-containing beverages. Br. J. Nutr. 1999, 81, 289–295. [Google Scholar] [CrossRef]
- International Rhino Foundation. 2019 Rhino Research Grantees. Available online: https://rhinos.org/research-publications/irf-funded-research/ (accessed on 25 September 2020).
- Muscher-Banse, A.S.; Marholt, L.; Eigendorf, N.; Wilkens, M.R.; Schröder, B.; Breves, G.; Cehak, A. Segmental diversity of phosphate transport along the intestinal axis in horses. J. Anim. Sci. 2017, 95, 165. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Murakami, Y.; Yano, H.; Fujikawa, H.; Osawa, T.; Asai, Y. Phytate and phosphorus movements in the digestive tract of horses. Equin. Veter. J. 2010, 31, 505–507. [Google Scholar] [CrossRef] [PubMed]
- Muya, S.M.; Oguge, N.O. Effects of browse availability and quality on black rhino (Diceros bicornis michaeli Groves 1967) diet in Nairobi National Park, Kenya. Afr. J. Ecol. 2000, 38, 62–71. [Google Scholar] [CrossRef] [Green Version]
- Helary, S.; Shaw, J.; Brown, D.; Owen-Smith, N. Natural Diets: Comparing Iron Levels through Seasons and Geographical Locations. In Proceedings of the International Workshop on Iron Storage Disease in Black Rhinoceros, Orlando, FL, USA, 23–26 February 2011. [Google Scholar]
- Morris, P.J.; Avgeris, S.E.; Baumgartner, R.E. Hemochromatosis in a Greater Indian Hill Mynah (Gracula religiosa): Case Report and Review of the Literature. J. Assoc. Avian Veter. 1989, 3, 87. [Google Scholar] [CrossRef]
- Davis, W.D.; Arrowsmith, W.R. The treatment of hemochromatosis by massive venesection. Ann. Intern. Med. 1953, 39, 723–734. [Google Scholar] [CrossRef]
- Solomon, M.; Hammell, M. Charging Ahead in Rhinoceros Welfare: Iron Overload Disorder Management in Black Rhinoceros (Diceros bicornis). Proceedings of Animal Behavior Management Alliance (ABMA), Tampa, FL, USA, 18–22 April 2016. [Google Scholar]
- Buowari, O.Y. Complications of venepuncture. Adv. Biosci. Biotechnol. 2013, 4, 126–128. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, K.; Livingston, S.; Williams, S.; Mylniczenko, N.; Rodriguez, C.; Pye, G.V.E. Iron Overload Disorder in Browsing Rhinos 2016 Workshop: A Review of Current Goals and Practical Action Planning. In Proceedings of the Comparative Nutrition Society 11th Biennial Symposium, Rio del Mar, Puerto Rico, 1–4 August 2016. [Google Scholar]
- Paglia, D.E.; Dennis, P. Role of Chronic Iron Overload in Multiple Disorders of Captive Black Rhinoceroses (Diceros bicornis). In Proceedings of the American Association of Zoo Veterinarians Annual Meeting, Columbus, OH, USA, 9–14 October 1999. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Moisture | Dry Matter | Iron | Vitamin C | Calcium | Phosphorus | |
|---|---|---|---|---|---|---|
| Food Item | % | % | ppm (DM) * | mg/100 g (AF) ** | (%DM) | (%DM) |
| Pelleted feed examples: | ||||||
| Mazuri ADF 25 Herbivore diet | 10.5 | 89.5 | 652 | nd | 1.51 | 0.95 |
| Mazuri ADF 16 Herbivore diet | 8.2 | 91.8 | 490 | nd | 1.16 | 0.84 |
| Mazuri Browser Rhino Cube 5Z1P | 10.4 | 89.6 | 222 | nd | 1.22 | 0.68 |
| Produce examples: | ||||||
| Cucumber (raw, whole) | 97.6 | 2.4 | 86 | 3.2 | 0.89 | 1.35 |
| Carrot (raw, whole) | 88.9 | 11.1 | 35 | 5.9 | 0.31 | 0.25 |
| Celery (raw, whole) | 95.1 | 4.9 | 29 | 3.1 | 0.98 | 0.51 |
| Sweet potato (raw, whole) | 79.8 | 20.2 | 22 | 2.4 | 0.36 | 0.29 |
| Apple (raw, whole) | 88.0 | 12.0 | 11 | 4.6 | 0.05 | 0.08 |
| Produce with higher vitamin C or iron level examples: | ||||||
| Green Leaf lettuce (fresh, raw) | 94.9 | 5.1 | 278 | 9.2 | 0.74 | 0.66 |
| Spinach (fresh, raw) | 91.4 | 8.6 | 264 | 28.1 | 1.02 | 0.73 |
| Romaine (fresh, raw) | 95.6 | 4.4 | 152 | 11.5 | 0.73 | 0.67 |
| Green beans (fresh, raw) | 92.2 | 7.8 | 101 | 12.2 | 0.55 | 0.53 |
| Cantaloupe melon (fresh, whole) | 93.1 | 6.9 | 74 | 36.7 | 0.11 | 0.17 |
| Cauliflower (raw, whole) | 94.0 | 6.0 | 60 | 48.2 | 0.55 | 0.75 |
| Tomatoes (raw, whole) | 95.7 | 4.3 | 53 | 13.7 | 0.18 | 0.47 |
| Honeydew melon (fresh, whole) | 91.6 | 8.4 | 37 | 18.0 | 0.11 | 0.33 |
| Watermelon (fresh, whole) | 92.3 | 7.7 | 33 | 8.1 | 0.14 | 0.33 |
| Banana (raw, whole with peel) | 82.4 | 17.6 | 15 | 8.7 | 0.07 | 0.12 |
| Orange (raw, whole with peel) | 82.6 | 17.4 | 14 | 71.0 | 0.57 | 0.13 |
| Hay/Fresh Browse examples: | ||||||
| Alfalfa Hay | 12.0 | 88.0 | 386 | nd | 1.84 | 0.31 |
| Coastal Bermudagrass Hay | 10.0 | 90.0 | 52 | nd | 0.57 | 0.19 |
| Timothy Hay | 11.0 | 89.0 | 48 | nd | 0.54 | 0.09 |
| Mulberry (whole branch fresh) | 67.6 | 32.4 | 84 | nd | 1.7 | 0.38 |
| Willow (whole branch fresh) | 63.2 | 36.8 | 63 | nd | 0.77 | 0.13 |
| Spineless cactus pads (Opuntia) | 91.5 | 8.5 | 22 | nd | 3.67 | 0.17 |
| Supplement examples: | ||||||
| Dicalcium Phosphate | 4.6 | 95.4 | 12,200 | nd | 23.4 | 19.7 |
| Trace mineral block | 0.1 | 99.9 | 1790 | nd | 0.3 | 0.05 |
| Dried Beet Pulp with molasses | 7 | 93 | 731 | nd | 1.15 | 0.1 |
| Molasses | 8.0 | 92.0 | 577 | 0.0 | 0.18 | 0.02 |
| Wheat bran | 8.4 | 91.6 | 186 | nd | 0.15 | 1.48 |
| Steamed Rolled Oats | 8.5 | 91.5 | 41 | 0.0 | 0.05 | 0.48 |
| VTLVP at DAK: Average Quarterly Volumes | ||||||||
|---|---|---|---|---|---|---|---|---|
| BR 1 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 |
| Q1 (January–March) | 10.0 L | 5.5 L | 7.0 L | 4.0 L | 3.7 L | 4.0 L | 1.0 L | 4.0 L |
| Q2 (April–June) | 4.0 L | 3.0 L | 6.0 L | 3.5 L | 4.0 L | 4.0 L | 0.5 L | NA |
| Q3 (July–September) | 5.5 L | 0.5 L | 0.0 L | 4.0 L | 0.0 L | 4.0 L | 4.0 L | 4.0 L |
| Q4 (October–December) | 7.5 L | 3.0 L | 2.5 L | 0.0 L | 0.0 L | 4.0 L | 3.0 L | - |
| BR 2 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 |
| Q1 (January–March) | NA | NA | NA | NA | 6.0 L | 2.0 L | 2.0 L | 2.0 L |
| Q2 (April–June) | NA | NA | NA | NA | 3.0 L | 3.0 L | 3.0 L | NA |
| Q3 (July–September) | NA | NA | NA | NA | 2.0 L | 4.0 L | 4.0 L | 0.0 L |
| Q4 (October–December) | NA | NA | NA | NA | 1.0 L | 2.0 L | 4.0 L | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sullivan, K.E.; Mylniczenko, N.D.; Nelson, S.E., Jr.; Coffin, B.; Lavin, S.R. Practical Management of Iron Overload Disorder (IOD) in Black Rhinoceros (BR; Diceros bicornis). Animals 2020, 10, 1991. https://doi.org/10.3390/ani10111991
Sullivan KE, Mylniczenko ND, Nelson SE Jr., Coffin B, Lavin SR. Practical Management of Iron Overload Disorder (IOD) in Black Rhinoceros (BR; Diceros bicornis). Animals. 2020; 10(11):1991. https://doi.org/10.3390/ani10111991
Chicago/Turabian StyleSullivan, Kathleen E., Natalie D. Mylniczenko, Steven E. Nelson, Jr., Brandy Coffin, and Shana R. Lavin. 2020. "Practical Management of Iron Overload Disorder (IOD) in Black Rhinoceros (BR; Diceros bicornis)" Animals 10, no. 11: 1991. https://doi.org/10.3390/ani10111991
APA StyleSullivan, K. E., Mylniczenko, N. D., Nelson, S. E., Jr., Coffin, B., & Lavin, S. R. (2020). Practical Management of Iron Overload Disorder (IOD) in Black Rhinoceros (BR; Diceros bicornis). Animals, 10(11), 1991. https://doi.org/10.3390/ani10111991