Integration of Physiological and Molecular Traits Would Help to Improve the Insights of Drought Resistance in Highbush Blueberry Cultivars


Abstract
1. Introduction
2. Results
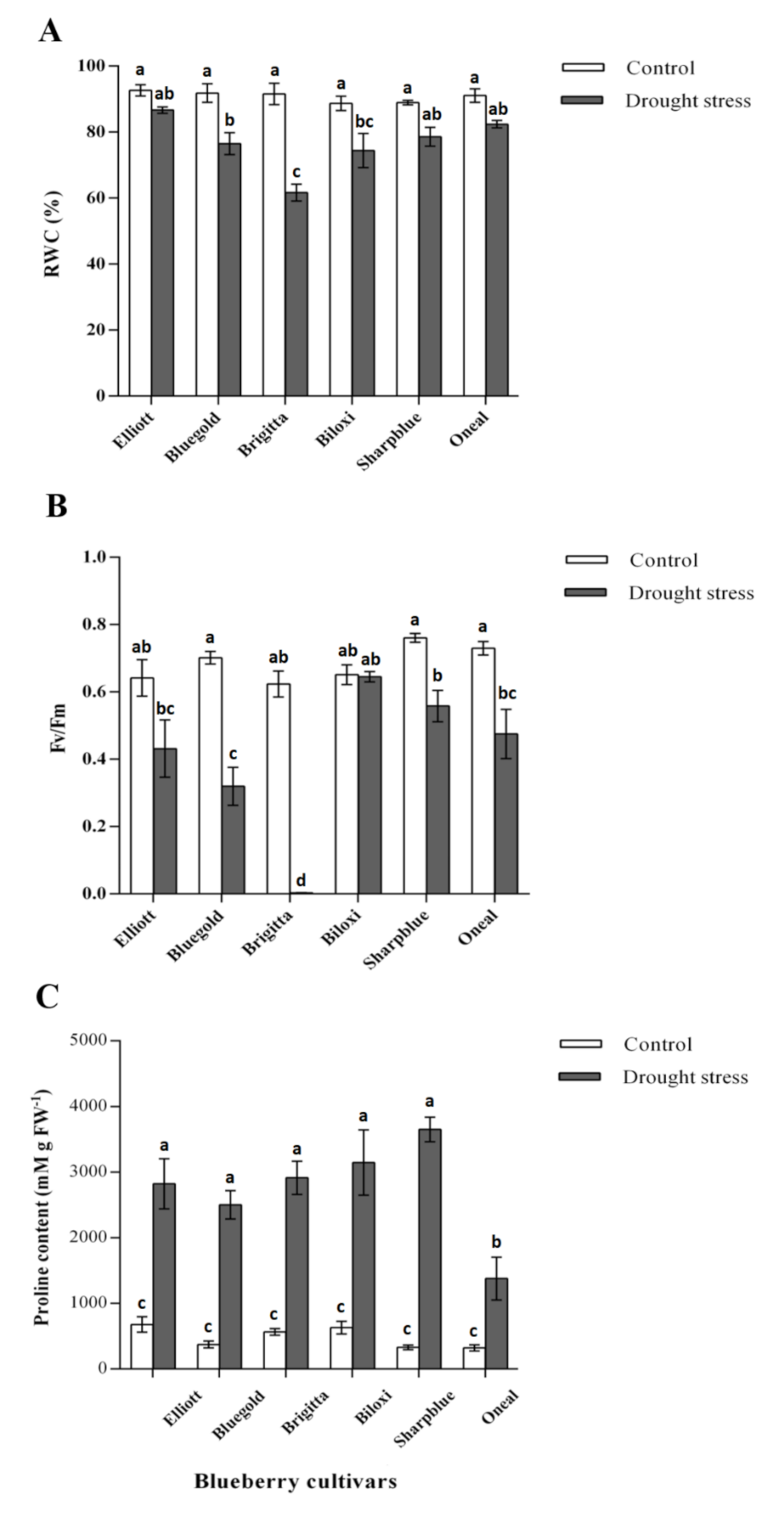
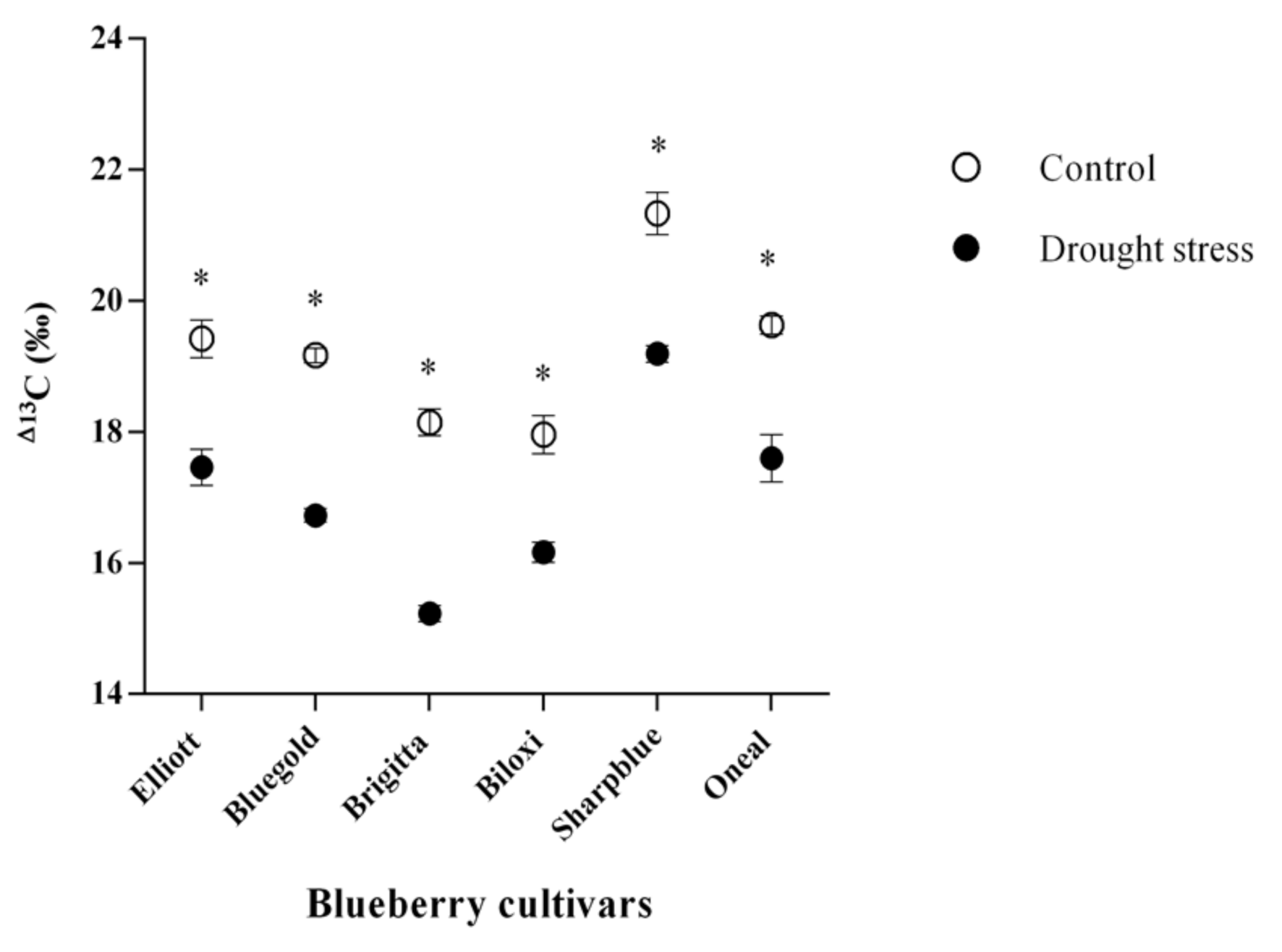
2.1. Physiological Measurements of Blueberry Plants upon Water Deficit Stress
2.2. Integrated Estimation of Plant Blueberry Cultivar Water Deficit Resistance
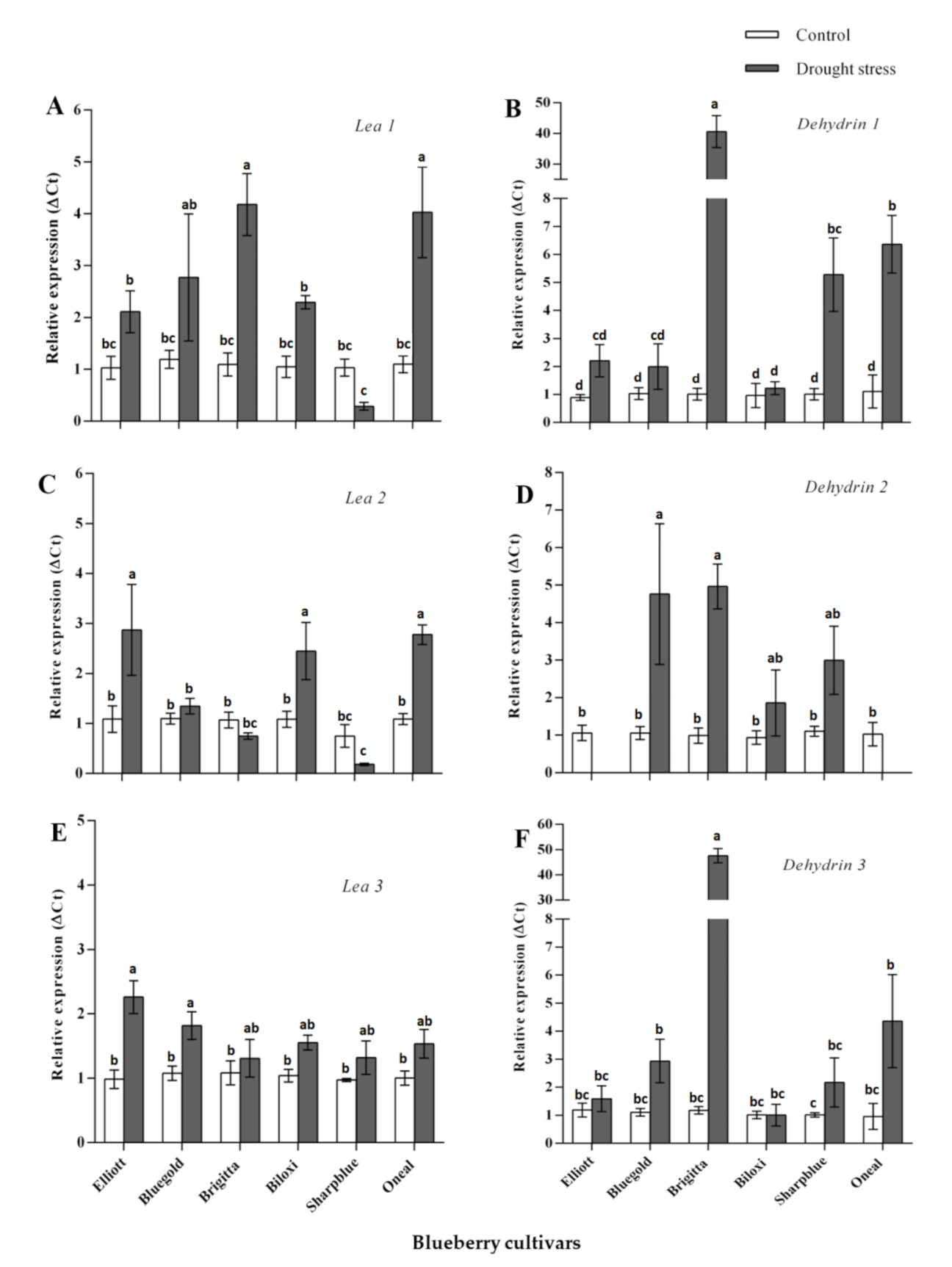
2.3. Analysis of Transcriptional Profile of Candidate Gene of the Late Embryogenesis Abundant (LEA) Family upon Water Deficit Stress
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plants, Growth Conditions, and Drought Treatment
5.2. Physiological Measurements of Blueberry Cultivars upon Water Deficit Stress
5.2.1. Relative Water Content
5.2.2. Chlorophyll Fluorescence
5.2.3. Leaf Proline Content
5.2.4. Carbon Isotope Discrimination (Δ13C)
5.3. Integrated Estimation of Plant Blueberry Cultivar Physiological Activity
5.4. Analysis of Transcriptional Profiles of the LEA Family Candidate Genes upon Water Deficit Stress
5.4.1. In Silico Analysis and Primer Design
5.4.2. RNA Isolation from Blueberry Leaves, cDNA Synthesis, and Gene Expression Analysis
5.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sankar, B.; Jaleel, C.A.; Manivannan, P.; Kishorekumar, A.; Somasundaram, R.; Panneerselvam, R. Drought-induced biochemical modifications and proline metabolism in Abelmoschus esculentus (L.) Moench. Acta Bot. Croat. 2007, 66, 43–56. [Google Scholar]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Stocker, T.F.; Qin, D.; Plattner, G.K.; Tignor, M.M.B.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. Climate Change 2013 the Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC—Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2013; Volume 9781107057, ISBN 9781107415324. [Google Scholar]
- Shinozaki, K.; Urano, K.; Maruyama, K.; Takahashi, F. Drought Stress, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2016; Volume 1, ISBN 9780123948083. [Google Scholar]
- Larkunthod, P.; Nounjan, N.; Siangliw, J.L.; Toojinda, T.; Sanitchon, J.; Jongdee, B.; Theerakulpisut, P. Physiological responses under drought stress of improved drought-tolerant rice lines and their parents. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 46, 679–687. [Google Scholar] [CrossRef]
- Monneveux, P.; Ramírez, D.A.; Pino, M.T. Drought tolerance in potato (S. tuberosum L.). Can we learn from drought tolerance research in cereals? Plant Sci. 2013, 205–206, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Salehi-Lisar, S.Y.; Bakhshayeshan-Agdam, H. Drought stress in plants: Causes, consequences, and tolerance. In Drought Stress Tolerance in Plants; Hossain, M.A., Wani, S.H., Bhattacharjee, S., Burritt, D.J., Tran, L.-S.P., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 1, pp. 1–16. ISBN 978-3-319-28899-4. [Google Scholar]
- Ganji Arjenaki, F.; Jabbari, R.; Morshedi, A. Evaluation of drought stress on relative water content, chlorophyll content and mineral elements of wheat (Triticum aestivum L.) varieties. Int. J. Agric. Crop Sci. 2012, 4, 726–729. [Google Scholar]
- De Silva, M.A.; Jifon, J.L.; da Silva, J.A.G.; Sharma, V. Use of physiological parameters as fast tools to screen for drought tolerance in sugarcane. Braz. J. Plant Physiol. 2007, 19, 193–201. [Google Scholar] [CrossRef]
- Guo, R.; Shi, L.; Jiao, Y.; Li, M.; Zhong, X.; Gu, F.; Liu, Q.; Xia, X.; Li, H. Metabolic responses to drought stress in the tissues of drought-tolerant and drought-sensitive wheat genotype seedlings. AoB Plants 2018, 10, ply016. [Google Scholar] [CrossRef]
- Suprasanna, P.; Nikalje, G.C.; Rai, A.N. Osmolyte accumulation and implications in plant abiotic stress tolerance. In Osmolytes and Plants Acclimation to Changing Environment: Emerging Omics Technologies; Springer: New Delhi, India, 2016; pp. 1–12. ISBN 9788132226161. [Google Scholar]
- Zegaoui, Z.; Planchais, S.; Cabassa, C.; Djebbar, R.; Belbachir, O.A.; Carol, P. Variation in relative water content, proline accumulation and stress gene expression in two cowpea landraces under drought. J. Plant Physiol. 2017, 218, 26–34. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef]
- Zargar, S.M.; Gupta, N.; Nazir, M.; Mahajan, R.; Malik, F.A.; Sofi, N.R.; Shikari, A.B.; Salgotra, R.K. Impact of drought on photosynthesis: Molecular perspective. Plant Gene 2017. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Osmolovskaya, N.; Shumilina, J.; Kim, A.; Didio, A.; Grishina, T.; Bilova, T.; Keltsieva, O.A.; Zhukov, V.; Tikhonovich, I.; Tarakhovskaya, E.; et al. Methodology of drought stress research: Experimental setup and physiological characterization. Int. J. Mol. Sci. 2018, 19, 4089. [Google Scholar] [CrossRef] [PubMed]
- Marino, G.; Haworth, M.; Scartazza, A.; Tognetti, R.; Centritto, M. A comparison of the variable j and carbon-isotopic composition of sugars methods to assess mesophyll conductance from the leaf to the canopy scale in drought-stressed cherry. Int. J. Mol. Sci. 2020, 21, 1222. [Google Scholar] [CrossRef] [PubMed]
- Brito, G.G.; Suassuna, N.D.; Silva, V.N.; Sofiatti, V.; Diola, V.; Morello, C.L. Leaf-level carbon isotope discrimination and its relationship with yield components as a tool for cotton phenotyping in unfavorable conditions. Acta Sci. Agron. 2014, 36, 335. [Google Scholar] [CrossRef][Green Version]
- Nagaraju, M.; Kumar, S.A.; Reddy, P.S.; Kumar, A.; Rao, D.M.; Kavi Kishor, P.B. Genome-scale identification, classification, and tissue specific expression analysis of late embryogenesis abundant (LEA) genes under abiotic stress conditions in Sorghum bicolor L. PLoS ONE 2019, 14, e0209980. [Google Scholar] [CrossRef] [PubMed]
- Pedrosa, A.M.; Martins, C.D.P.S.; Gonçalves, L.P.; Costa, M.G.C. Late Embryogenesis Abundant (LEA) constitutes a large and diverse family of proteins involved in development and abiotic stress responses in sweet orange (Citrus sinensis L. Osb.). PLoS ONE 2015, 10, e0145785. [Google Scholar] [CrossRef]
- Jin, X.; Cao, D.; Wang, Z.; Ma, L.; Tian, K.; Liu, Y.; Gong, Z.; Zhu, X.; Jiang, C.; Li, Y. Genome-wide identification and expression analyses of the LEA protein gene family in tea plant reveal their involvement in seed development and abiotic stress responses. Sci. Rep. 2019, 9, 14123. [Google Scholar] [CrossRef]
- Magwanga, R.O.; Lu, P.; Kirungu, J.N.; Lu, H.; Wang, X.; Cai, X.; Zhou, Z.; Zhang, Z.; Salih, H.; Wang, K.; et al. Characterization of the late embryogenesis abundant (LEA) proteins family and their role in drought stress tolerance in upland cotton. BMC Genet. 2018, 19, 6. [Google Scholar] [CrossRef] [PubMed]
- Adak, N.; Gubbuk, H.; Tetik, N. Yield, quality and biochemical properties of various strawberry cultivars under water stress. J. Sci. Food Agric. 2018, 98, 304–311. [Google Scholar] [CrossRef]
- Morales, C.G.; Pino, M.T.; del Pozo, A. Phenological and physiological responses to drought stress and subsequent rehydration cycles in two raspberry cultivars. Sci. Hortic. (Amst.) 2013, 162, 234–241. [Google Scholar] [CrossRef]
- Retamales, J.B.; Hancock, J.F. Blueberries; CABI—Centre for Agriculture and Bioscience International: Oxfordshire, UK, 2012; Volume 21, ISBN 9781845938260. [Google Scholar]
- Estrada, F.; Escobar, A.; Romero-Bravo, S.; González-Talice, J.; Poblete-Echeverría, C.; Caligari, P.D.S.; Lobos, G.A. Fluorescence phenotyping in blueberry breeding for genotype selection under drought conditions, with or without heat stress. Sci. Hortic. (Amst.) 2015, 181, 147–161. [Google Scholar] [CrossRef]
- Lobos, G.A.; Hancock, J.F. Breeding blueberries for a changing global environment: A review. Front. Plant Sci. 2015, 6, 782. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Bryla, D.R.; Strik, B.C. Effects of cultivar and plant spacing on the seasonal water requirements of highbush blueberry. J. Am. Soc. Hortic. Sci. 2007, 132, 270–277. [Google Scholar] [CrossRef]
- Kumari, A.; Khan, I.; Singh, A.K.; Singh, S.K. Morphological, physiological and biochemical responses of poplar plants to drought stress. J. AgriSearch 2018, 5, 2026–2032. [Google Scholar] [CrossRef]
- De Almeida Silva, M.; Jifon, J.L.; Sharma, V.; da Silva, J.A.G.; Caputo, M.M.; Damaj, M.B.; Guimarães, E.R.; Ferro, M.I.T. Use of physiological parameters in screening drought tolerance in sugarcane genotypes. Sugar Tech 2011, 13, 191–197. [Google Scholar] [CrossRef]
- Pinheiro, C.; Chaves, M.M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 2011, 62, 869–882. [Google Scholar] [CrossRef]
- Dixon, L.S.; Godoy, J.V.; Carter, A.H. Evaluating the utility of carbon isotope discrimination for wheat breeding in the Pacific Northwest. Plant Phenom. 2019, 2019, 1–11. [Google Scholar] [CrossRef]
- Li, R.; Guo, P.-g.; Michael, B.; Stefania, G.; Salvatore, C. Evaluation of chlorophyll content and fluorescence parameters as indicators of drought tolerance in barley. Agric. Sci. China 2006, 5, 751–757. [Google Scholar] [CrossRef]
- Monneveux, P.; Sheshshayee, M.S.; Akhter, J.; Ribaut, J.M. Using carbon isotope discrimination to select maize (Zea mays L.) inbred lines and hybrids for drought tolerance. Plant Sci. 2007, 173, 390–396. [Google Scholar] [CrossRef]
- Leprince, O.; Buitink, J. Desiccation tolerance: From genomics to the field. Plant Sci. 2010, 179, 554–564. [Google Scholar] [CrossRef]
- Banerjee, A.; Roychoudhury, A. Group II late embryogenesis abundant (LEA) proteins: Structural and functional aspects in plant abiotic stress. Plant Growth Regul. 2016, 79, 1–17. [Google Scholar] [CrossRef]
- Fu, Y.; Ma, H.; Chen, S.; Gu, T.; Gong, J. Control of proline accumulation under drought via a novel pathway comprising the histone methylase CAU1 and the transcription factor ANAC055. J. Exp. Bot. 2018, 69, 579–588. [Google Scholar] [CrossRef]
- Man, D.; Bao, Y.-X.; Han, L.-B.; Zhang, X. Drought tolerance associated with proline and hormone metabolism in two tall fescue cultivars. HortScience 2011, 46, 1027–1032. [Google Scholar] [CrossRef]
- Battaglia, M.; Olvera-Carrillo, Y.; Garciarrubio, A.; Campos, F.; Covarrubias, A.A. The enigmatic LEA Ppoteins and other hydrophilins. Plant Physiol. 2008, 148, 6–24. [Google Scholar] [CrossRef]
- Kavi Kishor, P.B.; Sreenivasulu, N. Is proline accumulation per se correlated with stress tolerance or is proline homeostasis a more critical issue? Plant Cell Environ. 2014, 37, 300–311. [Google Scholar] [CrossRef]
- Zhao, P.; Liu, F.; Zheng, G.; Liu, H. Group 3 late embryogenesis abundant protein in Arabidopsis: Structure, regulation, and function. Acta Physiol. Plant. 2011, 33, 1063–1073. [Google Scholar] [CrossRef]
- Hand, S.C.; Menze, M.A.; Toner, M.; Boswell, L.; Moore, D. LEA proteins during water stress: Not just for plants anymore. Annu. Rev. Physiol. 2011, 73, 115–134. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Richards, C.L.; Bossdorf, O.; Muth, N.Z.; Gurevitch, J.; Pigliucci, M. Jack of all trades, master of some? On the role of phenotypic plasticity in plant invasions. Ecol. Lett. 2006, 9, 981–993. [Google Scholar] [CrossRef]
- Lloyd, G.; McCown, B. Commercially feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot tip culture. Combined Proceedings of the International Plant Propagators Society. Int. Plant Propagators’ Soc. Proc. 1980, 30, 421–427. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Freeland, R.S. Review of soil moisture sensing using soil electrical conductivity. Trans. ASAE 1989, 32, 2190–2194. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology; Springer: New York, NY, USA, 2008; ISBN 978-0-387-78340-6. [Google Scholar]
- Montecinos, S.; Salinas, P.; Oses, R.; González-Silvestre, L.; Molina-Montenegro, M. Linking climatic variability with spatial performance in two varieties of quinoa distributed in a semi-arid zone. Am. J. Plant Sci. 2012, 3, 1682–1687. [Google Scholar] [CrossRef]
- Gupta, V.; Estrada, A.D.; Blakley, I.; Reid, R.; Patel, K.; Meyer, M.D.; Andersen, S.U.; Brown, A.F.; Lila, M.A.; Loraine, A.E. RNA-Seq analysis and annotation of a draft blueberry genome assembly identifies candidate genes involved in fruit ripening, biosynthesis of bioactive compounds, and stage-specific alternative splicing. Gigascience 2015, 4, 5. [Google Scholar] [CrossRef]
- Rowland, L.J.; Alkharouf, N.; Darwish, O.; Ogden, E.L.; Polashock, J.J.; Bassil, N.V.; Main, D. Generation and analysis of blueberry transcriptome sequences from leaves, developing fruit, and flower buds from cold acclimation through deacclimation. BMC Plant Biol. 2012, 12, 46. [Google Scholar] [CrossRef]
- Dhanaraj, A.L.; Slovin, J.P.; Rowland, L.J. Analysis of gene expression associated with cold acclimation in blueberry floral buds using expressed sequence tags. Plant Sci. 2004, 166, 863–872. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. The plant hormone abscisic acid mediates the drought-induced expression but not the seed-specific expression of rd22, a gene responsive to dehydration stress in Arabidopsis thaliana. MGG Mol. Gen. Genet. 1993. [Google Scholar] [CrossRef]
- Davies, C.; Robinson, S.P. Sugar accumulation in grape berries (cloning of two putative vacuolar invertase cDNAs and their expression in grapevine tissues). Plant Physiol. 1996, 111, 275–283. [Google Scholar] [CrossRef]
- Vashisth, T.; Johnson, L.K.; Malladi, A. An efficient RNA isolation procedure and identification of reference genes for normalization of gene expression in blueberry. Plant Cell Rep. 2011, 30, 2167–2176. [Google Scholar] [CrossRef]
- Zifkin, M.; Jin, A.; Ozga, J.A.; Zaharia, L.I.; Schernthaner, J.P.; Gesell, A.; Abrams, S.R.; Kennedy, J.A.; Constabel, C.P. Gene expression and metabolite profiling of developing highbush blueberry fruit indicates transcriptional regulation of flavonoid metabolism and activation of abscisic acid metabolism. Plant Physiol. 2012, 158, 200–224. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Source of Variation | d.f | MS | F | p |
|---|---|---|---|---|
| Relative water content (RWC) | ||||
| Intercept | 1 | 403,822.9 | 13,284.36 | p < 0.001 |
| Cultivar | 5 | 184.7 | 6.08 | p < 0.001 |
| Irrigation | 1 | 2855.2 | 93.93 | p < 0.001 |
| Cultivar*Irrigation | 5 | 173.3 | 5.70 | p < 0.001 |
| Error | 46 | 30.4 | ||
| Photochemical efficiency of PSII (Fv/Fm) | ||||
| Intercept | 1 | 20.89461 | 1607.872 | p < 0.001 |
| Cultivar | 5 | 0.17368 | 13.365 | p < 0.001 |
| Irrigation | 1 | 1.20068 | 92.394 | p < 0.001 |
| Cultivar*Irrigation | 5 | 0.11042 | 8.497 | p < 0.001 |
| Error | 60 | 0.01300 | ||
| Proline content | ||||
| Intercept | 1 | 155,304,002 | 946.2628 | p < 0.001 |
| Cultivar | 5 | 1,726,505 | 10.5195 | p < 0.001 |
| Irrigation | 1 | 76,087,947 | 463.6017 | p < 0.001 |
| Cultivar*Irrigation | 5 | 1,340,468 | 8.1674 | p < 0.001 |
| Error | 48 | 164,124 | ||
| Carbon Isotopic Discrimination of 13C | ||||
| Intercept | 1 | 38,045.46 | 181,554.1 | p < 0.001 |
| Cultivar | 5 | 14.85 | 70.9 | p < 0.001 |
| Irrigation | 1 | 64.67 | 308.6 | p < 0.001 |
| Cultivar*Irrigation | 5 | 0.38 | 1.8 | 0.1302 |
| Error | 46 | 0.21 | ||
| Cultivar | PPA Control (%) | PPA Water Deficit (%) | WDRI (PPAWD/PPAC) |
|---|---|---|---|
| Sharpblue | 56.76 | 45.63 | 0.804 |
| O’Neal | 55.77 | 44.10 | 0.791 |
| Bluegold | 54.56 | 36.91 | 0.677 |
| Elliott | 53.11 | 44.05 | 0.830 |
| Biloxi * | 52.14 | 47.05 | 0.903 |
| Brigitta | 51.18 | 21.40 | 0.418 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balboa, K.; Ballesteros, G.I.; Molina-Montenegro, M.A. Integration of Physiological and Molecular Traits Would Help to Improve the Insights of Drought Resistance in Highbush Blueberry Cultivars. Plants 2020, 9, 1457. https://doi.org/10.3390/plants9111457
Balboa K, Ballesteros GI, Molina-Montenegro MA. Integration of Physiological and Molecular Traits Would Help to Improve the Insights of Drought Resistance in Highbush Blueberry Cultivars. Plants. 2020; 9(11):1457. https://doi.org/10.3390/plants9111457
Chicago/Turabian StyleBalboa, Karen, Gabriel I. Ballesteros, and Marco A. Molina-Montenegro. 2020. "Integration of Physiological and Molecular Traits Would Help to Improve the Insights of Drought Resistance in Highbush Blueberry Cultivars" Plants 9, no. 11: 1457. https://doi.org/10.3390/plants9111457
APA StyleBalboa, K., Ballesteros, G. I., & Molina-Montenegro, M. A. (2020). Integration of Physiological and Molecular Traits Would Help to Improve the Insights of Drought Resistance in Highbush Blueberry Cultivars. Plants, 9(11), 1457. https://doi.org/10.3390/plants9111457