Efficient Agrobacterium-Mediated Transformation of the Commercial Hybrid Poplar Populus Alba × Populus glandulosa Uyeki

Abstract
1. Introduction
2. Results
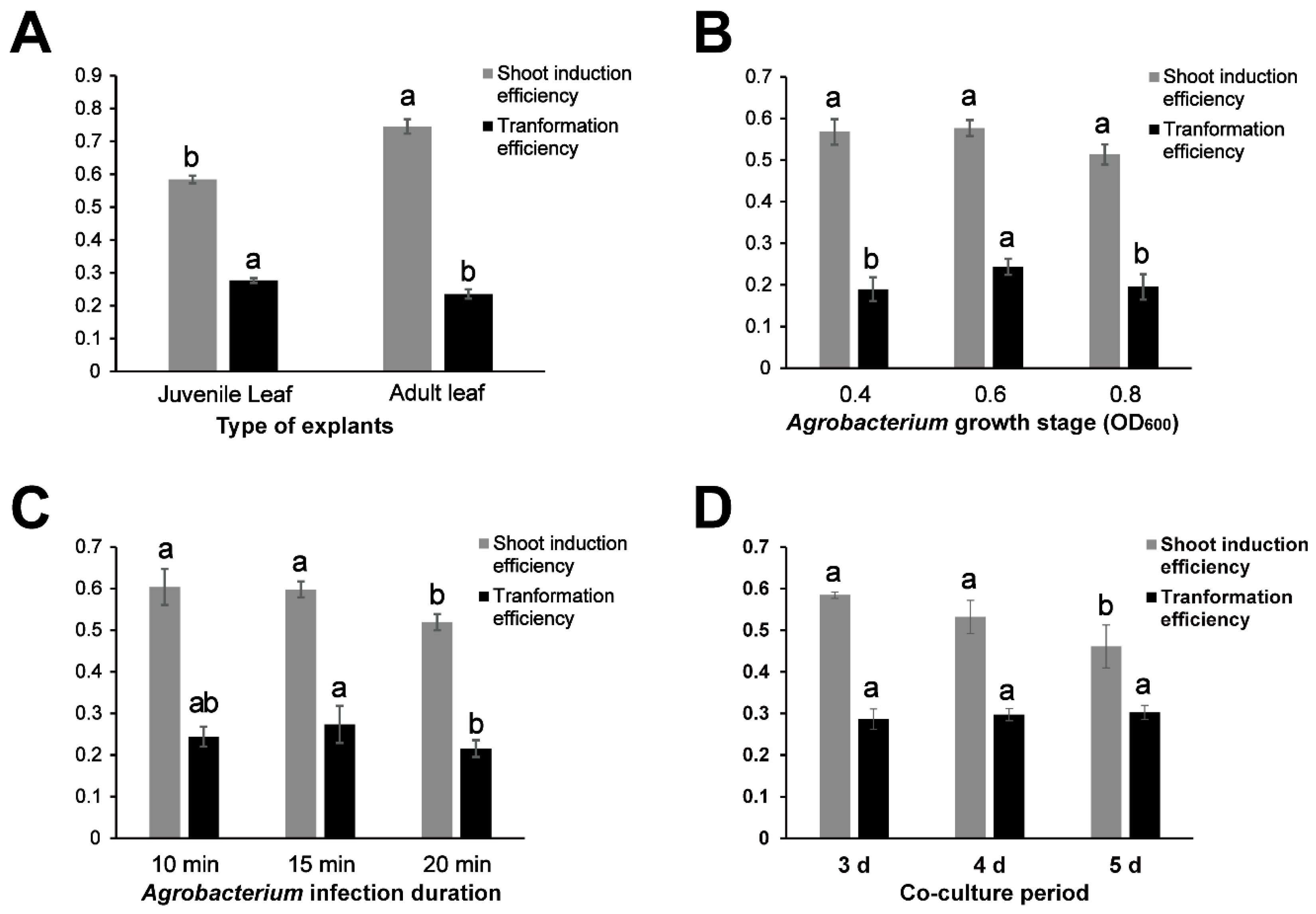
2.1. Optimization of the Transformation Procedure
2.2. Simple and Fast Transformation System
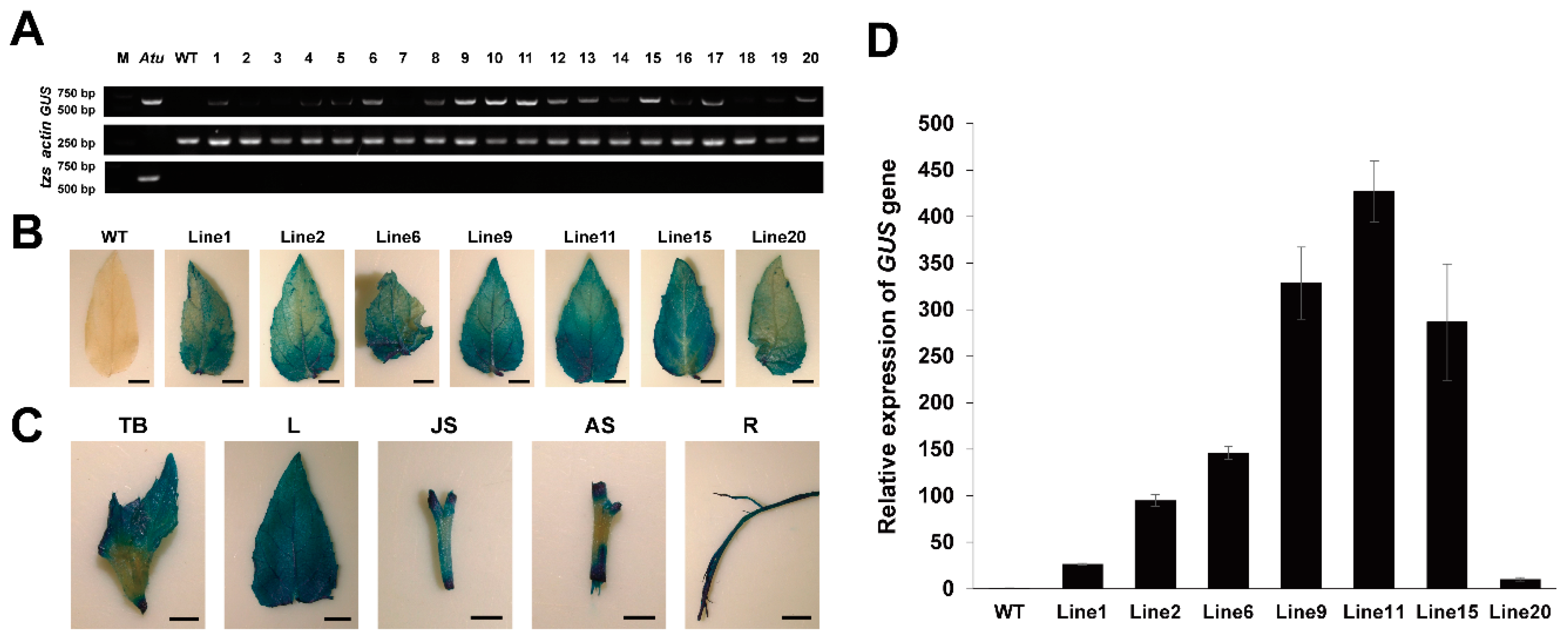
2.3. Positive Plant Identification by PCR Analysis
2.4. Quantitative Assay of GUS Expression by qRT-PCR
2.5. β-Glucuronidase Histochemical Assays
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Agrobacterium Culture and Transformation Procedure
4.3. Plant Genomic DNA Isolation and PCR Analysis
4.4. Quantitative Analysis of GUS Expression by qRT-PCR
4.5. β-Glucuronidase Histochemical Assay
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Balatinecz, J.J.; Kretschmann, D.E.; Leclercq, A. Achievements in the utilization of poplar wood-guideposts for the future. For. Chron. 2001, 77, 265–269. [Google Scholar] [CrossRef]
- Confalonieri, M.; Belenghi, B.; Balestrazzi, A.; Negri, S.; Facciotto, G.; Schenone, G.; Delledonne, M. Transformation of elite white poplar (Populus alba L.) cv. ‘Villafranca’ and evaluation of herbicide resistance. Plant Cell Rep. 2000, 19, 978–982. [Google Scholar] [CrossRef] [PubMed]
- Parsons, T.J.; Sinkar, V.P.; Stettler, R.F.; Nester, E.W.; Gordon, M.P. Transformation of poplar by Agrobacterium tumefaciens. Biotechnology 1986, 4, 533–536. [Google Scholar] [CrossRef]
- Vainstein, A.; Marton, I.; Zuker, A.; Danziger, M.; Tzfira, T. Permanent genome modifications in plant cells by transient viral vectors. Trends Biotechnol. 2011, 29, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Jiang, L. Transient expression of fluorescent fusion proteins in protoplasts of suspension cultured cells. Nat. Protoc. 2007, 2, 2348–2353. [Google Scholar] [CrossRef]
- Taylor, N.J.; Fauquet, C.M. Microparticle bombardment as a tool in plant science and agricultural biotechnology. DNA Cell Biol. 2002, 21, 963–977. [Google Scholar] [CrossRef]
- Victor, B.B.; Amy, M.B.; Richard, M.; Serge, F.; Lisa, G.; Sonali, G.; Steven, H.S. Genetic transformation: A powerful tool for dissection of adaptive traits in trees. New Phytol. 2005, 167, 9–18. [Google Scholar]
- Tzfira, T.; Jensen, C.S.; Wang, W.; Zuker, A.; Vincour, B.; Altman, A.; Vainstein, A. Transgenic Populus tremula: A step-by-step protocol for its Agrobacterium-mediated transformation. Plant Mol. Biol. 1997, 15, 219–235. [Google Scholar] [CrossRef]
- Cseke, L.J.; Cseke, S.B.; Podila, G.K. High efficiency poplar transformation. Plant Cell Rep. 2007, 26, 1529–1538. [Google Scholar] [CrossRef]
- Delledonne, M.; Allegro, G.; Belenghi, B.; Balestrazzi, A.; Picco, F.; Levine, A.; Zelasco, S.; Calligari, P.; Confalonieri, M. Transformation of white poplar (Populus alba L.) with a novel Arabidopsis thaliana cysteine proteinase inhibitor and analysis of insect pest resistance. Mol. Breed. 2001, 7, 35–42. [Google Scholar] [CrossRef]
- Biswas, K.K.; Mohri, T.; Kogawara, S.; Hase, Y.; Oono, Y. An improved system for shoot regeneration from stem explants of Lombardy poplar (Populus nigra L. var. italica Koehne). Am. J. Plant Sci. 2012, 3, 1181–1186. [Google Scholar] [CrossRef]
- Eriksson, M.E.; Israelsson, M.; Olsson, O.; Moritz, T. Increased gibberellin biosynthesis in transgenic trees promotes growth, biomass production and xylem fiber length. Nat. Biotechnol. 2000, 18, 784–788. [Google Scholar] [CrossRef]
- Song, J.; Lu, S.; Chen, Z.Z.; Lourenco, R.; Chiang, V.L. Genetic transformation of Populus trichocarpa genotype Nisqually-1: A functional genomic tool for woody plants. Plant Cell Physiol. 2006, 47, 1582–1589. [Google Scholar] [CrossRef]
- Han, K.H.; Meilan, R.; Ma, C.; Strauss, S.H. An Agrobacterium tumefaciens transformation protocol effective on a variety of cottonwood hybrids (genus Populus). Plant Cell Rep. 2000, 19, 315–320. [Google Scholar] [CrossRef]
- Movahedi, A.; Zhang, J.; Amirian, R.; Zhuge, Q. An efficient Agrobacterium-mediated transformation system for poplar. Int. J. Mol. Sci. 2014, 15, 10780–10793. [Google Scholar] [CrossRef] [PubMed]
- Peña, L.; Séguin, A. Recent advances in the genetic transformation of trees. Trends Biotechnol. 2001, 19, 500–506. [Google Scholar] [CrossRef]
- Han, X.; Ma, S.; Kong, X.; Takano, T.; Liu, S. Efficient Agrobacterium-mediated transformation of hybrid poplar Populus davidiana Dode × Populus bollena Lauche. Int. J. Mol. Sci. 2013, 14, 2515–2528. [Google Scholar] [CrossRef]
- Wang, H.H.; Wang, C.T.; Liu, H.; Tang, R.J.; Zhang, H.X. An efficient Agrobacterium-mediated transformation and regeneration system for leaf explants of two elite aspen hybrid clones Populus alba × P. berolinensis and Populus davidiana × P. bolleana. Plant Cell Rep. 2011, 30, 2037–2044. [Google Scholar] [CrossRef] [PubMed]
- Thakur, A.K.; Sarita, S.; Srivastava, D.K. Plant regeneration and genetic transformation studies in petiole tissue of Himalayan poplar (Populus ciliata Wall). Curr. Sci. 2005, 89, 664–668. [Google Scholar]
- Yadav, R.; Arora, P.; Kumar, D.; Katyal, D.; Dilbaghi, N.; Chaudhury, A. High frequency direct plant regeneration from leaf, internode, and root segments of Eastern Cottonwood (Populus deltoides). Plant Biotechnol. Rep. 2009, 3, 175–182. [Google Scholar] [CrossRef]
- Mohri, T.; Yamamoto, N.; Shinohara, K. Agrobacterium-mediated transformation of lombardy poplar (Populus nigra L. var. italica Koehne) using stem segments. J. For. Res. 1996, 1, 13–16. [Google Scholar] [CrossRef]
- Yevtushenko, D.P.; Misra, S. Efficient Agrobacterium-mediated transformation of commercial hybrid poplar Populus nigra L. × P. maximowiczii A. Henry. Plant Cell Rep. 2010, 29, 211–221. [Google Scholar] [CrossRef]
- Kang, B.; Osburn, L.; Kopsell, D.; Tuskan, G.A.; Cheng, Z.M. Micropropagation of Populus trichocarpa ‘Nisqually-1’: The genotype deriving the Populus reference genome. Plant Cell Tiss. Org. Cult. 2009, 99, 251–257. [Google Scholar] [CrossRef]
- Klocko, A.L.; Meilan, R.; James, R.R.; Viswanath, V.; Ma, C.; Payne, P.; Miller, L.; Skinner, J.S.; Oppert, B.; Cardineau, G.A.; et al. Bt-Cry3Aa transgene expression reduces insect damage and improves growth in field-grown hybrid poplar. Can. J. For. Res. 2013, 44, 28–35. [Google Scholar] [CrossRef]
- Harfouche, A.; Meilan, R.; Altman, A. Molecular and physiological responses to abiotic stress in forest trees and their relevance to tree improvement. Tree Physiol. 2014, 34, 1181–1198. [Google Scholar] [CrossRef]
- Coleman, H.D.; Yan, J.; Mansfield, S.D. Sucrose synthase affects carbon partitioning to increase cellulose production and altered cell wall ultrastructure. Proc. Natl. Acad. Sci. USA 2009, 106, 13118–13123. [Google Scholar] [CrossRef]
- Hu, J.; Wang, L.; Yan, D.; Lu, M.Z. Research and application of transgenic poplar in China, in challenges and opportunities for the world’s forests in the 21st century. Springer Dordr. 2014, 81, 567–584. [Google Scholar]
- Ramanathan, T.; Gurudeeban, S.; Satyavani, K. In vitro plant regeneration from leaf primordia of gum-bearing tree Aegle marmelos. J. For. Res. 2010, 4, 208–212. [Google Scholar] [CrossRef][Green Version]
- Cavusoglu, A.; Ipekci-Altas, Z.; Bajrovic, K.; Gozukirmizi, N.; Zehir, A. Direct and indirect plant regeneration from various explants of eastern cottonwood clones (Populus deltoides Bartram ex Marsh.) with tissue culture. Afr. J. Biotechnol. 2011, 10, 3216–3221. [Google Scholar]
- Husaini, A.M. Pre-and post-agroinfection strategies for efficient leaf disk transformation and regeneration of transgenic strawberry plants. Plant Cell Rep. 2010, 29, 97–110. [Google Scholar] [CrossRef]
- Li, B.; Xie, C.; Qiu, H. Production of selectable marker-free transgenic tobacco plants using a non-selection approach: Chimerism or escape, transgene inheritance, and efficiency. Plant Cell Rep. 2009, 28, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Cervera, M.; Pina, J.A.; Juarez, J.; Navarro, L.; Pena, L. A broad exploration of a transgenic population of citrus: Stability of gene expression and phenotype. Theor. Appl. Genet. 2000, 11, 1683–1685. [Google Scholar] [CrossRef]
- Tang, J.; Scarth, R.; Fristensky, B. Effects of genomic position and copy number of acyl-ACP thioesterase transgenes on the level of the target fatty acids in Brassica napus L. Mol. Breed. 2003, 12, 71–81. [Google Scholar] [CrossRef]
- Zheng, L.; Liu, G.; Meng, X.; Li, Y.; Wang, Y. A versatile Agrobacterium-mediated transient gene expression system for herbaceous plants and trees. Biochem. Genet. 2012, 50, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Gururaj, H.B.; Padma, M.N.; Giridhar, P.; Ravishankar, G.A. Functional validation of Capsicum frutescens aminotransferase gene involved in vanillylamine biosynthesis using Agrobacterium mediated genetic transformation studies in Nicotiana tabacum and Capsicum frutescens calli cultures. Plant Sci. 2012, 195, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Porebski, S.; Bailey, L.G.; Baum, B.R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Rep. 1997, 15, 8–15. [Google Scholar] [CrossRef]
- Zhang, D.; Du, Q.; Xu, B.; Zhang, Z.; Li, B. The actin multigene family in Populus: Organization, expression and phylogenetic analysis. Mol. Genet. Genom. 2010, 284, 105–119. [Google Scholar] [CrossRef]
- Han, Z.F.; Hunter, D.M.; Sibbald, S.; Zhang, J.S.; Tian, L. Biological activity of the tzs gene of nopaline Agrobacterium tumefaciens GV3101 in plant regeneration and genetic transformation. Mol. Plant Microbe Interact. 2013, 26, 1359–1365. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Components | M1 (Plant Propagation and Pre-Culture) | M2 (Co-Culture) | M3 (Shoot Induction) | M4 (Root Induction) |
|---|---|---|---|---|
| MS | 4.43 g/L | 4.43 g/L | 4.43 g/L | 2.215 g/L |
| Sucrose | 30 g/L | 30 g/L | 30 g/L | 30 g/L |
| Agar | 5.8 g/L | 5.8 g/L | 5.8 g/L | 5.8 g/L |
| NAA | — | 0.05 mg/L | 0.05 mg/L | 0.02mg/L |
| 6-BA | — | 0.5 mg/L | 0.5 mg/L | — |
| IBA | — | — | — | 0.05 mg/L |
| Kanamycin | — | — | 30 mg/L | 30 mg/L |
| Cefotaxime | — | — | 200 mg/L | 200 mg/L |
| Timentin | — | — | 200 mg/L | 200 mg/L |
| pH | 5.8 | 5.8 | 5.8 | 5.8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, C.; Lu, L.; Guo, Y.; Xu, H.; Li, R. Efficient Agrobacterium-Mediated Transformation of the Commercial Hybrid Poplar Populus Alba × Populus glandulosa Uyeki. Int. J. Mol. Sci. 2019, 20, 2594. https://doi.org/10.3390/ijms20102594
Song C, Lu L, Guo Y, Xu H, Li R. Efficient Agrobacterium-Mediated Transformation of the Commercial Hybrid Poplar Populus Alba × Populus glandulosa Uyeki. International Journal of Molecular Sciences. 2019; 20(10):2594. https://doi.org/10.3390/ijms20102594
Chicago/Turabian StyleSong, Chengwei, Liang Lu, Yayu Guo, Huimin Xu, and Ruili Li. 2019. "Efficient Agrobacterium-Mediated Transformation of the Commercial Hybrid Poplar Populus Alba × Populus glandulosa Uyeki" International Journal of Molecular Sciences 20, no. 10: 2594. https://doi.org/10.3390/ijms20102594
APA StyleSong, C., Lu, L., Guo, Y., Xu, H., & Li, R. (2019). Efficient Agrobacterium-Mediated Transformation of the Commercial Hybrid Poplar Populus Alba × Populus glandulosa Uyeki. International Journal of Molecular Sciences, 20(10), 2594. https://doi.org/10.3390/ijms20102594