Purification, Characterization and Degradation Performance of a Novel Dextranase from Penicillium cyclopium CICC-4022


 ,
,
Abstract
1. Introduction
2. Results and Discussions
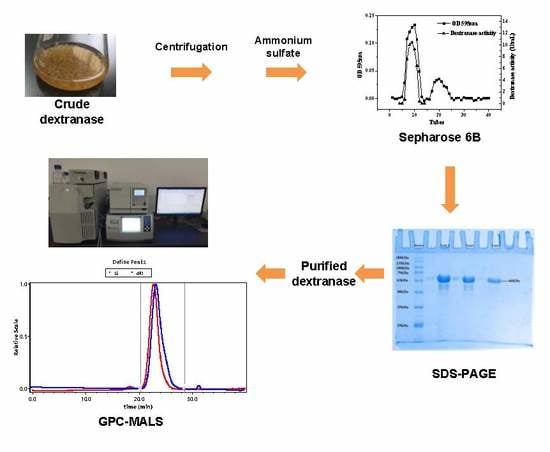
2.1. Purification of Dextranase
2.2. Dextranase Enzymatic Properties
2.2.1. Effect of Temperature on Dextranase Activity and Thermal Stability
2.2.2. Effect of pH on Dextranase Activity and pH Stability
2.2.3. Substrate Specificity and Final Products Analysis of Enzyme
2.2.4. Effects of Metal Ions and Compounds on Enzyme Activity
2.2.5. Kinetic Parameters of Dextranase
2.3. Determination of Mw and Mw/Mn of Dextran
2.3.1. Standard Curve of Dextran of GPC
2.3.2. Comparison of GPC-MALS with GPC and Error Analysis
2.3.3. Degradation of Dextran Polymers by Dextranase
3. Materials and Methods
3.1. Materials
3.2. Preparation of Crude Dextranase
3.3. Dextranase Activity Assay
multiple of enzyme solution]/[Volume of enzyme solution (mL) × Time (min)]
3.4. Determination of Protein Concentration
3.5. Purification of Dextranase
3.6. Dextranase Enzymatic Properties
3.6.1. Effect of Temperature and pH on Dextranase Activity and Stability
3.6.2. Substrate Specificity and Analysis of Hydrolysis Products
3.6.3. Effects of Metal ions and other Compounds on Dextranase Activity
3.6.4. Enzyme Kinetics
3.7. Determination of Mw and Mw/Mn
3.7.1. Gel permeation chromatography (GPC)
3.7.2. Degradation of dextrans by dextranase
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bashari, M.; Eibaid, A.; Wang, J.P.; Tian, Y.Q.; Xu, X.M.; Jin, Z.Y. Influence of low ultrasound intensity on the degradation of dextran catalyzed by dextranase. Ultrason. Sonochem. 2013, 20, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Cai, R.H.; Yan, W.L.; Lyu, M.S.; Fang, Y.W.; Wang, S.J. Purification and characterization of a biofilm-degradable dextranase from a marine bacterium. Mar. Drugs 2018, 16, 51. [Google Scholar] [CrossRef] [PubMed]
- Shahid, F.; Aman, A.; Nawaz, M.A.; Karim, A.; Qader, S.A. Chitosan hydrogel microspheres: An effective covalent matrix for crosslinking of soluble dextranase to increase stability and recycling efficiency. Bioprocess Biosyst. Eng. 2017, 40, 451–461. [Google Scholar] [CrossRef]
- Wu, D.T.; Zhang, H.B.; Huang, L.J.; Hu, X.Q. Purification and characterization of extracellular dextranase from a novel producer, Hypocrea lixii F1002, and its use in oligodextran production. Process Biochem. 2011, 46, 1942–1950. [Google Scholar] [CrossRef]
- Walker, G.V.; Heng, N.C.K.; Carne, A.; Tagg, J.R.; Wescombe, P.A. Salivaricin E and abundant dextranase activity may contribute to the anti-cariogenic potential of the probiotic candidate Streptococcus salivarius JH. Microbiology 2016, 162, 476–486. [Google Scholar] [CrossRef]
- Qiu, Y.X.; Mao, M.Y.; Jang, D.; Hong, X.; Yang, Y.M.; Hu, T. Co-operative effect of exogenous dextranase and sodium fluoride on multispecies biofilms. J. Dent. Sci. 2016, 11, 41–47. [Google Scholar] [CrossRef]
- Marotta, M.; Martino, A.; Rosa, A.D.; Farina, E.; Carteni, M.; Rosa, M.D. Degradation of dental plaque glucans and prevention of glucan formation using commercial enzymes. Process Biochem. 2002, 38, 101–108. [Google Scholar] [CrossRef]
- Ren, W.; Wang, S.J.; Lü, M.S.; Wang, X.B.; Fang, Y.W.; Jiao, Y.L.; Hu, J.E. Optimization of four types of antimicrobial agents to increase the inhibitory ability of marine Arthrobacter oxydans KQ11 dextranase mouthwash. Chin. J. Oceanol. Limn. 2016, 34, 354–366. [Google Scholar] [CrossRef]
- Wang, X.B.; Lu, M.S.; Wang, S.J.; Fang, Y.W.; Wang, D.L.; Ren, W.; Zhao, G.M. The atmospheric and room-temperature plasma (ARTP) method on the dextranase activity and structure. Int. J. Biol. Macromol. 2014, 70, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Hild, E.; Brumbley, S.M.; O’Shea, M.G.; Nevalainen, H.; Bergquist, P.L. A Paenibacillus sp. dextranase mutant pool with improved thermostability and activity. Appl. Microbiol. Biotechnol. 2007, 75, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Zohra, R.R.; Aman, A.; Ansari, A.; Haider, M.S.; Qader, S.A.U. Purification, characterization and end product analysis of dextran degrading endodextranase from Bacillus licheniformis, KIBGE-IB25. Int. J. Biol. Macromol. 2015, 78, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Netsopa, S.; Niamsanit, S.; Araki, T.; Kongkeitkajorn, M.B.; Milintawisamai, N. Purification and Characterization Including Dextran Hydrolysis of Dextranase from Aspergillus allahabadii X26. Sugar Tech 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Li, R.H.; Zhang, H.B.; Wu, M.; Hu, X.Q. Purification, characterization, and application of a thermostable dextranase from Talaromyces pinophilus. J. Ind. Microbiol. Biotechnol. 2017, 44, 317–327. [Google Scholar] [CrossRef]
- Cai, R.H.; Lu, M.S.; Fang, Y.W.; Jiao, Y.L.; Zhu, Q.; Liu, Z.P.; Wang, S.J. Screening, production, and characterization of dextranase from Catenovulum sp. Ann. Microbiol. 2014, 64, 147–155. [Google Scholar] [CrossRef]
- Klahan, P.; Okuyama, M.; Jinnai, K.; Ma, M.; Kikuchi, A.; Kumagai, Y.; Tagami, T.; Kimura, A. Engineered dextranase from Streptococcus mutans enhances the production of longer isomaltooligosaccharides. Biosci. Biotechnol. Biochem. 2018, 82, 1–8. [Google Scholar] [CrossRef]
- Kim, Y.M.; Kim, D. Characterization of novel thermostable dextranase from Thermotoga lettingae TMO. Appl. Microbiol. Biotechnol. 2010, 85, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Lu, H.Q.; Hang, F.X.; Li, S.B.; Liu, J.D. Improved dextranase production by Chaetomium gracile through optimization of carbon source and fermentation parameters. Sugar Tech 2017, 19, 432–437. [Google Scholar] [CrossRef]
- Zohra, R.R.; Aman, A.; Zohra, R.R.; Ansari, A.; Ghani, M.; Qader, S.A.U. Dextranase: Hyper production of dextran degrading enzyme from newly isolated strain of Bacillus licheniformis. Carbohydr. Polym. 2013, 92, 2149–2153. [Google Scholar] [CrossRef]
- Gozu, Y.; Ishizaki, Y.; Hosoyama, Y.; Miyazaki, T.; Nishikawa, A.; Tonozuka, T. A glycoside hydrolase family 31 dextranase with high transglucosylation activity from Flavobacterium johnsoniae. Biosci. Biotechnol. Biochem. 2016, 80, 1562–1567. [Google Scholar] [CrossRef]
- Sufiate, B.L.; Soares, F.E.F.; Moreira, S.S.; Gouveia, A.S.; Cardoso, E.F.; Braga, F.R.; Araújo, J.V.; Queiroz, J.H. In vitro and in silico characterization of a novel dextranase from Pochonia chlamydosporia. Biotech 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Bhatia, S.; Bhakri, G.; Arora, M.; Batta, S.K.; Uppal, S.K. Kinetic and thermodynamic properties of partially purified dextranase from Paecilomyces lilacinus and its application in dextran removal from cane juice. Sugar Tech 2016, 18, 1–7. [Google Scholar] [CrossRef]
- Khalikova, E.; Susi, P.; Usanov, N.; Korpela, T. Purification and properties of extracellular dextranase from a Bacillus sp. J. Chromatogr. B 2003, 796, 315–326. [Google Scholar] [CrossRef]
- Kim, Y.M.; Ko, E.A.; Kang, H.K.; Kim, D. Construction, expression and characterization of fusion enzyme from Arthrobacter oxydans dextranase and Klebsiella pneumoniaeamylase. Biotechnol Lett. 2009, 31, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zhou, N.D.; Tian, Y.P. Purification, characterization, and biocatalytic potential of a novel dextranase from Chaetomium globosum. Biotechnol Lett. 2018, 40, 1407–1418. [Google Scholar] [CrossRef]
- Abdelnaby, M.A.; Ismail, A.M.S.; Abdelfattah, A.M. Preparation and some properties of immobilized Penicillium funiculosum 258 dextranase. Process Biochem. 1999, 34, 391–398. [Google Scholar] [CrossRef]
- Zhang, L.X.; Duan, R.; Chen, L.J.; Ling, X.P.; Lu, Y.H. Expression of α-1,6-dextranase from Penicillium minioluteum in P.pastoris. Sci. Technol. Food Ind. 2011, 11, 194–197. [Google Scholar]
- Wynter, C.V.A.; Chang, M.; Jersey, J.D.; Patel, B.; Inkerman, P.A.; Hamiton, S. Isolation and characterization of a thermostable dextranase. Enzym. Microb. Technol. 1997, 20, 242–247. [Google Scholar] [CrossRef]
- Lee, J.H.; Nam, S.H.; Park, H.J.; Kim, Y.M.; Kim, N.; Kim, G.; Seo, E.S.; Kang, S.S.; Kim, D. Biochemical characterization of dextranase from Arthrobacter oxydans and its cloning and expression in Escherichia coli. Food Sci. Biotechnol. 2010, 19, 757–762. [Google Scholar] [CrossRef]
- Purushe, S.; Prakash, D.; Nawani, N.N.; Dhakephalkar, P.; Kapadnis, B. Biocatalytic potential of an alkalophilic and thermophilic dextranase as a remedial measure for dextran removal during sugar manufacture. Bioresour. Technol. 2012, 115, 2–7. [Google Scholar] [CrossRef]
- Galvez, M.A.; Lopez, M.A. Production and characterization of a dextranase from an isolated Paecilomyces lilacinus strain. Appl. Microbiol. Biotechnol. 1991, 36, 327–331. [Google Scholar]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Biochem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Abdelwahed, N.A.M.; Ahmed, E.F.; El-Gammal, E.W.; Hawas, U.W. Application of statistical design for the optimization of dextranase production by a novel fungus isolated from Red Sea sponge. Biotech 2014, 4, 533–544. [Google Scholar] [CrossRef]
- Carlsson, N.; Borde, A.; Wölfel, S.; Kerman, B.; Larsson, A. Quantification of protein concentration by the Bradford method in the presence of pharmaceutical polymers. Anal. Biochem. 2011, 411, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Lineweaver, H.; Burk, D. The determination of enzyme dissociation constant. J. Am. Chem. Soc. 1934, 56, 658–666. [Google Scholar] [CrossRef]
- Pawcenis, D.; Thomas, J.L.; Łojewski, T.; Milczarek, J.M.; Łojewska, J. Towards determination of absolute molar mass of cellulose polymer by size exclusion chromatography with multiple angle laser light scattering detection. J. Chromatogr A 2015, 1409, 53–59. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Step | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg) | Purification (Fold) | Yield (%) |
|---|---|---|---|---|---|
| Culture dextranase | 7.08 | 533.15 | 75.32 | 1.00 | 100.00 |
| (NH4)2SO4 precipitation | 4.19 | 465.71 | 111.06 | 1.71 | 87.35 |
| Sepharose 6B | 0.44 | 155.54 | 350.29 | 16.09 | 29.17 |
| Substrate | Main Linkage | Relative Activity (%) |
|---|---|---|
| DextranT3 | α-1,6 | 65.17 ± 0.7 |
| DextranT5 | α-1,6 | 65.54 ± 0.6 |
| DextranT20 | α-1,6 | 90.64 ± 0.4 |
| DextranT40 | α-1,6 | 98.50 ± 0.2 |
| DextranT70 | α-1,6 | 100.00 ± 0.5 |
| DextranT500 | α-1,6 | 99.62 ± 0.1 |
| DextranT2000 | α-1,6 | 97.00 ± 0.4 |
| Soluble starch | α-1,4; α-1,6 | 20.83 ± 0.8 |
| Cellulose | β-1,4 | 0 |
| Sucrose | β-1,2 | 0 |
| Chitin | β-1,4 | 0 |
| Chitosan | β-1,4 | 0 |
| β-Cyclodextrin | β-1,4 | 0 |
| Compound | Relative Activity (%) | Relative Activity (%) | Relative Activity (%) |
|---|---|---|---|
| (0.001 M) | (0.005 M) | (0.01 M) | |
| Control | 100 | 100 | 100 |
| Cu2+ | 24.02 ± 0.15 | 22.53 ± 0.05 | 16.41 ± 0.2 |
| Fe3+ | 99.55 ± 0.10 | 64.61 ± 0.07 | 67.27 ± 0.07 |
| Zn2+ | 90.95 ± 0.02 | 59.35 ± 0.13 | 16.85 ± 0.05 |
| Ca2+ | 101.81 ± 0.03 | 93.30 ± 0.04 | 40.73 ± 0.06 |
| Cr2+ | 90.95 ± 0.08 | 72.74 ± 0.12 | 48.25 ± 0.06 |
| K+ | 97.74 ± 0.06 | 114.83 ± 0.10 | 105.75 ± 0.05 |
| Li+ | 116.28 ± 0.05 | 115.78 ± 0.09 | 103.98 ± 0.12 |
| Na+ | 104.98 ± 0.07 | 111.48 ± 0.02 | 105.75 ± 0.05 |
| Pb2+ | 59.29 ± 0.05 | 22.53 ± 0.03 | 20.39 ± 0.15 |
| Ni2+ | 95.48 ± 0.15 | 97.61 ± 0.05 | 67.71 ± 0.16 |
| NH4+ | 107.58 ± 0.04 | 105.21 ± 0.07 | 99.53 ± 0.15 |
| Fe2+ | 104.74 ± 0.06 | 100.95 ± 0.02 | 109.95 ± 0.12 |
| Tris | 103.79 ± 0.08 | 100.00 ± 0.07 | 100.47 ± 0.10 |
| SDS | 106.16 ± 0.03 | 46.94 ± 0.05 | 34.15 ± 0.13 |
| Urea | 112.79 ± 0.04 | 109.00 ± 0.11 | 99.05 ± 0.04 |
| EDTA | 102.37 ± 0.07 | 80.58 ± 0.14 | 81.05 ± 0.05 |
| Dextran T5 b | Dextran T70 b | Dextran T2000 b | |
|---|---|---|---|
| Km (mM) | 2.72 ± 0.04 | 2.61 ± 0.02 | 2.45 ± 0.01 |
| Vmax (µmol Glucose min−1 mg−1 Protein) | 189.50 ± 0.03 | 192.70 ± 0.04 | 203.21 ± 0.01 |
| Kacat (s−1) | 208.45 ± 0.01 | 211.97 ± 0.02 | 223.53 ± 0.03 |
| Kcat/Km (M−1 s−1) | 76636.0 ± 0.06 | 81214.6 ± 0.03 | 91236.7 ± 0.05 |
| Standard | Mw (kDa) | Mw δ (%) | Mw/Mn | Mw/Mn δ (%) | ||||
|---|---|---|---|---|---|---|---|---|
| GPC | GPC-MALS | GPC | GPC-MALS | GPC | GPC-MALS | GPC | GPC-MALS | |
| Dextran (Mw ≈ 1.27 kDa) | 0.84 | 1.32 | −33.86 | 3.94 | 1.72 | 1.20 | 36.51 | −4.76 |
| Dextran (Mw ≈ 5.52 kDa) | 3.09 | 5.4 | −44.02 | −2.17 | 3.03 | 1.57 | 89.38 | −1.88 |
| Dextran (Mw ≈ 23.8 kDa) | 21.4 | 23.99 | −10.08 | 0.80 | 1.28 | 1.229 | 24.62 | 7.69 |
| Dextran (Mw ≈ 48.6 kDa) | 31.57 | 49.98 | −35.04 | 2.84 | 1.28 | 1.24 | −5.88 | −5.15 |
| Dextran (Mw ≈ 80.9 kDa) | 59.26 | 82.26 | −26.74 | 1.68 | 1.64 | 1.28 | 12.33 | −5.48 |
| Dextran (Mw ≈ 667.8 kDa) | 827.80 | 621.5 | 23.96 | −6.93 | 5.24 | 2.049 | 160.70 | 1.94 |
| Reaction Time (min) | 0.2 U/mL | 0.4 U/mL | 0.6 U/mL | |||
|---|---|---|---|---|---|---|
| Mw (kDa) | Mn/Mw | Mw (kDa) | Mn/Mw | Mw (kDa) | Mn/Mw | |
| 0 | 1869 | 3.236 | 1859 | 1.523 | 1859 | 3.358 |
| 4 | 1553 | 2.322 | 865.9 | 3.048 | 251 | 5.891 |
| 8 | 720.6 | 4.524 | 242.3 | 4.272 | 83.69 | 3.991 |
| 12 | 426.5 | 5.329 | 69.25 | 3.12 | 48.34 | 2.198 |
| 16 | 216 | 4.837 | 40.74 | 2.504 | 24.13 | 2.133 |
| 20 | 119.5 | 4.373 | 29.54 | 2.071 | 20.78 | 1.917 |
| 30 | 51.16 | 1.966 | 16.77 | 1.82 | 11.6 | 1.731 |
| 40 | 32.41 | 1.618 | 11.35 | 1.672 | 9.247 | 1.507 |
| 50 | 22.26 | 2.187 | 9.685 | 1.542 | 7.156 | 1.502 |
| 60 | 13.79 | 1.627 | 8.052 | 1.433 | 6.262 | 1.36 |
| 90 | 11.39 | 1.494 | 6.362 | 1.306 | ||
| 120 | 8.239 | 1.375 | 5.954 | 1.238 | ||
| 150 | 7.358 | 1.396 | ||||
| Reaction Time (min) | Dextran (1%) | Dextran (3%) | Dextran (5%) | |||
|---|---|---|---|---|---|---|
| Mw (kDa) | Mn/Mw | Mw (kDa) | Mn/Mw | Mw (kDa) | Mn/Mw | |
| 0 | 1726 | 2.995 | 1759 | 3.358 | 1750 | 3.829 |
| 4 | 49.94 | 2.534 | 251 | 5.891 | 487.8 | 11.383 |
| 8 | 16.06 | 1.702 | 83.69 | 3.991 | 194.4 | 6.03 |
| 12 | 10.65 | 1.39 | 48.34 | 2.198 | 107.1 | 5.032 |
| 16 | 7.196 | 1.388 | 24.13 | 2.133 | 63.9 | 3.519 |
| 20 | 5.893 | 1335 | 20.78 | 1.917 | 42.4 | 2.964 |
| 30 | 4.993 | 1.234 | 11.6 | 1.731 | 23.88 | 2.237 |
| 40 | 4.434 | 1.53 | 9.247 | 1.507 | 16.54 | 1.979 |
| 50 | 7.156 | 1.502 | 12.8 | 1.693 | ||
| 60 | 6.262 | 1.36 | 10.05 | 1.651 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, R.; Zhong, L.; Xie, F.; Wei, L.; Gan, L.; Wang, X.; Liao, A. Purification, Characterization and Degradation Performance of a Novel Dextranase from Penicillium cyclopium CICC-4022. Int. J. Mol. Sci. 2019, 20, 1360. https://doi.org/10.3390/ijms20061360
Huang R, Zhong L, Xie F, Wei L, Gan L, Wang X, Liao A. Purification, Characterization and Degradation Performance of a Novel Dextranase from Penicillium cyclopium CICC-4022. International Journal of Molecular Sciences. 2019; 20(6):1360. https://doi.org/10.3390/ijms20061360
Chicago/Turabian StyleHuang, Ruijie, Lei Zhong, Fengwei Xie, Liming Wei, Lanfang Gan, Xuejiao Wang, and Anping Liao. 2019. "Purification, Characterization and Degradation Performance of a Novel Dextranase from Penicillium cyclopium CICC-4022" International Journal of Molecular Sciences 20, no. 6: 1360. https://doi.org/10.3390/ijms20061360
APA StyleHuang, R., Zhong, L., Xie, F., Wei, L., Gan, L., Wang, X., & Liao, A. (2019). Purification, Characterization and Degradation Performance of a Novel Dextranase from Penicillium cyclopium CICC-4022. International Journal of Molecular Sciences, 20(6), 1360. https://doi.org/10.3390/ijms20061360