
Microbial Pathogens Linked to Vaginal Microbiome Dysbiosis and Therapeutic Tools for Their Treatment


Abstract
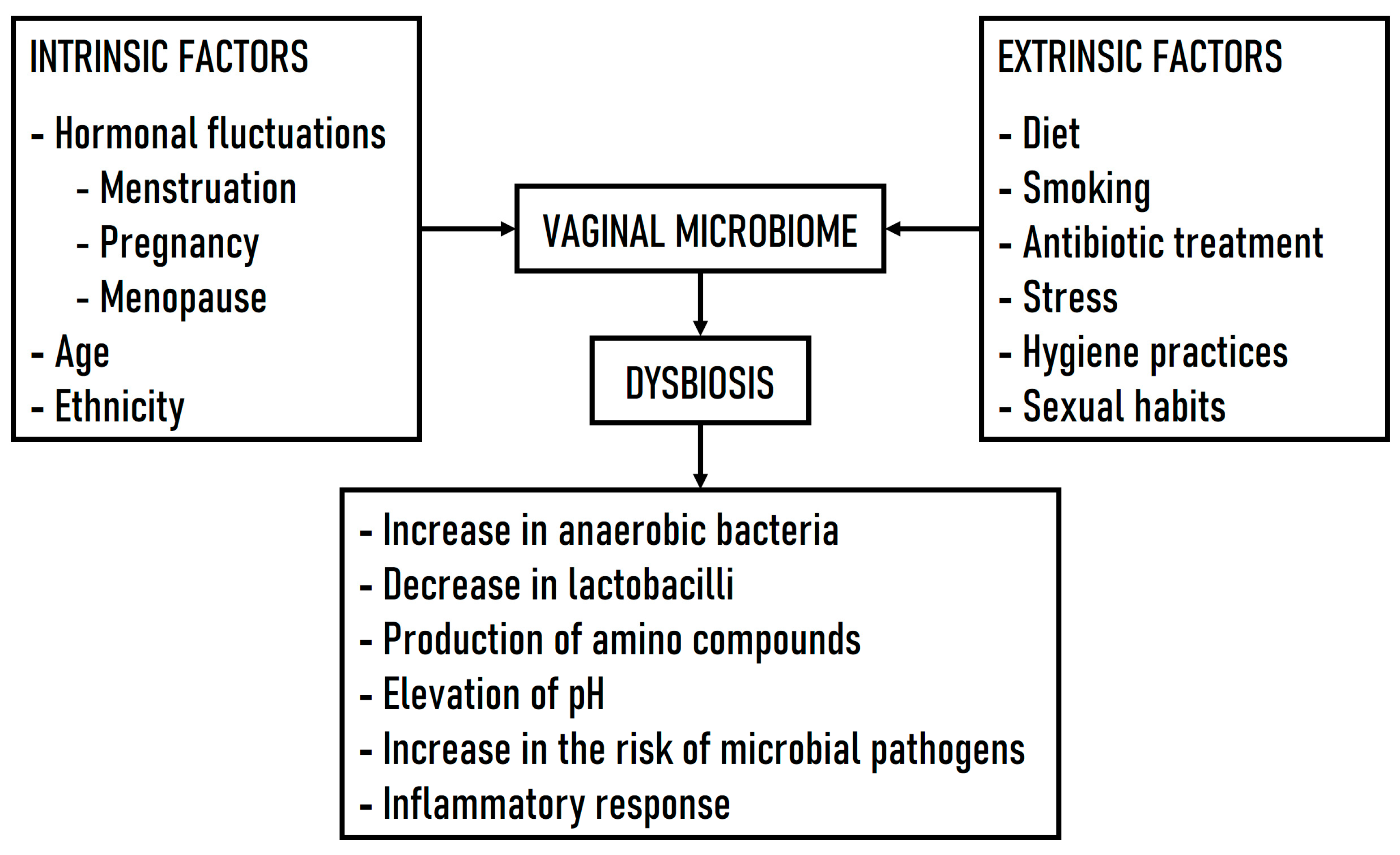
1. Introduction
2. Methods
3. Bacterial Vaginosis
3.1. Diagnostic Methods for Bacterial Vaginosis

{kind=link}
{kind=link}
| Test Method | Criteria | Advantages | Disadvantages | Performance Data |
|---|---|---|---|---|
| Reference Methods | ||||
| Amsel criteria [35] | Clinical/microscopy-based (wet mount microscopy/phase contrast)
| Rapid point-of-care test |
| Comparison with Nugent score:
|
| Nugent score [36] | Microscopy-based method (Gram stain) 0–3: No indication of BV 4–6: No conclusive indication of BV 7–10: Indication of BV | Objective criteria |
| Comparison with Amsel criteria:
|
| Hay–Ison criteria [37] | Microscopy-based method (Gram-stain) 0: No bacteria1: No indication of BV 2: No conclusive indication of BV 3: Indication of BV 4: Gram-positive cocci | Objective criteria |
| Comparison with Amsel criteria:
|
| Molecular methods | ||||
| FISH [44] | Fluorescence microscopy-based analysis of polymicrobial structures using 16S rRNA probes | Biofilm visualization in situ/dysbiosis visualization |
|
Comparison with reference methods:
|
| Sequencing [45] | Gene sequencing (NGS) | Quantitative determination of the VM |
|
Comparison with reference methods:
|
| Multiplex qPCR [46,47] | Multiplex quantitative PCR | Commercially available automated test, indirect biofilm detection possible |
|
Comparison with reference methods:
|
3.2. Bacterial Vaginosis Biofilm Formation
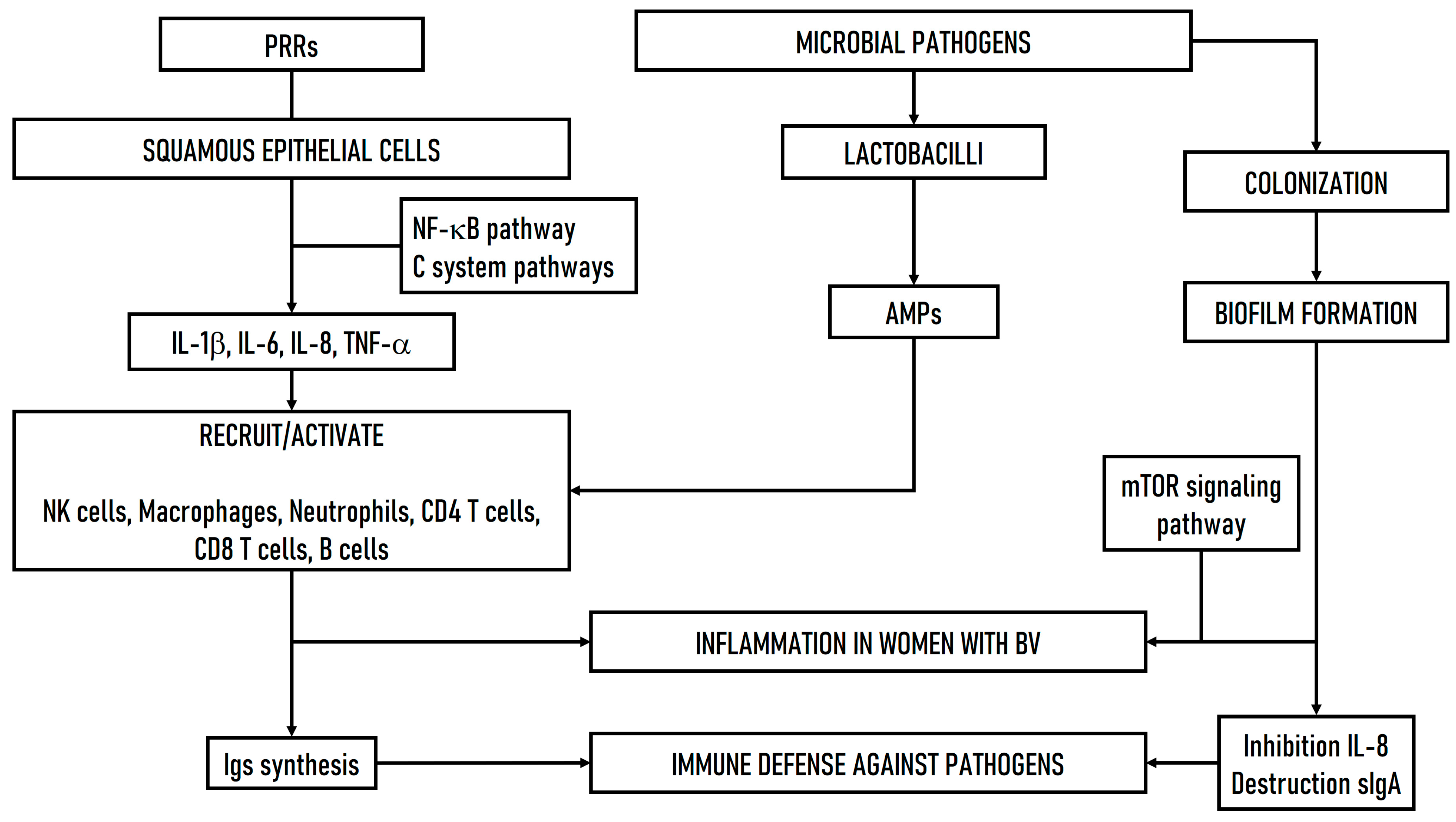
3.3. Immune Response
4. Other Microbial Pathologies Associated with VM Dysbiosis
4.1. Aerobic Vaginitis
4.2. Pelvic Inflammatory Disease
4.3. Vulvovaginal Candidiasis
4.4. Susceptibility to Sexually Transmitted Infections
4.4.1. Human Immunodeficiency Virus (HIV)
4.4.2. Human Papillomavirus (HPV)
4.4.3. Trichomonas Vaginalis Infection
5. Therapeutic Tools
5.1. Antibiotic Therapies
5.2. Live Biotherapeutic Products
| Reference | Type of Study | Treatment | Results |
|---|---|---|---|
| Laue et al. [159] | RCT. N = 36 female volunteers aged ≥18 years at reproductive age with stable menstrual cycles or postmenopausal women presenting BV, initially treated with metronidazole. | Fermented dairy drink: Contained pasteurized whole milk and the yogurt starter cultures Streptococcus thermophilus (95%) and Lactobacillus delbrueckii subsp. bulgaricus (5%). The dairy drink was supplemented with L. crispatus strain LbV 88, L. paragasseri strain LbV 150N, L. mulieris strain LbV 116, and L. rhamnosus strain LbV96 at a concentration of each strain of 1 × 107 CFU/mL for 4 weeks. | After 4 weeks, Amsel scores, discharge and odor, and Nugent scores decreased in the verum group compared to the control group. Supplemental consumption of yogurt containing these probiotic strains improved the recovery rate and symptoms of BV and tended to improve the vaginal microbial pattern. |
| Heczko et al. [160] | RCT. N = 241 (18–50 years) females with diagnoses of R-BV and AV with standard metronidazole treatment. | prOVag® oral probiotic capsules: L. paragasseri strain 57C, Limosilactobacillus fermentum strain 57A, and Lactiplantibacillus plantarum strain 57B, at a concentration of 108 CFU/capsule for 3 months. | prOVag extended the time to clinical relapse of BV/AV symptoms by up to 51% compared to placebo. AV recurrence was delayed by up to 76%. Probiotic use also reduced and maintained a low vaginal pH and Nugent score, and increased vaginal Lactobacillus counts compared to standard treatment. |
| Pendharkar et al. [161] | Observational. N = 50 (18–55 years) females with diagnoses of BV or R-VVC and treated with clindamycin/metronidazole (C/M) and fluconazole. | EcoVag® oral probiotic capsules: L. paragasseri strain DSM 14,869 and L. rhamnosus strain DSM 14870, at a concentration of 108 CFU/strain/capsule for 6–12 months. | In trial I, BV was treated with C/M followed by 5 days of EcoVag® capsules. Trial II included three groups: BV with prolonged C/M and EcoVag®, R-VVC with extended fluconazole and EcoVag®, and fluconazole treatment alone. In trial I, the 6-month cure rate for BV was 50%, and in trial II, both the 6- and 12-month cure rates were 67%. For VVC, women receiving fluconazole and EcoVag® had cure rates of 100% and 89% at 6 and 12 months, respectively, while those receiving fluconazole only had cure rates of 100% and 70%. Lactobacilli were associated with BV cure in both trials, while EcoVag® strains were significant only in trial II. |
| Davar et al. [162] | RCT. N = 59 (mean age 32.3 years) females with diagnosis of R-VVC and treated, initially treated with fluconazole. | Pro-Digest probiotic oral capsules: Lactobacillus acidophilus (7.5 × 109 CFU/capsule, Bifidobacterium bifidum (6 × 109 CFU/capsule), and B. longum (1:5 × 109 CFU/capsule) for 6 months. | The results showed that taking probiotics without antifungal drugs could be highly effective in treating VVC, resulting in a lower recurrence rate as well. |
| Cohen et al. [163] | RCT. N = 228 (18–45 years) females with untreated BV (asymptomatic or symptomatic). | Lactin-V probiotic capsules: L. crispatus strain CTV-05, at 2 × 109 CFU/dose for 10 weeks. | Recurrence of BV at week 12 was lower in the Lactin-V group compared to the placebo group. L. crispatus CTV-05 was detected in 79% of Lactin-V participants at the 12-week visit, and adverse events were similar between the Lactin-V and placebo groups. |
| Reznichenko et al. [164] | Randomized prospective. N = 186 (18–45 years) females with symptomatic BV, cured with metronidazole. | Oral probiotic cocktail: L. crispatus strain LMG S-29995, Levilactobacillus brevis, and L. acidophilus in proportions of 60%, 20%, and 20%, respectively. Treatment for 16 weeks. | Oral intake of probiotics can significantly reduce the percentage of recurrences of BV in recently treated women and prolong the time to recurrence of the disease. |
| Zhang et al. [165] | Parallel controlled trial. N = 70 (18–65 years) females with BV and treated with metronidazole. | Oral probiotics: L. rhamnosus strain GR-1 and Limosilactobacillus reuteri strain RC-14, treated orally for 30–90 days as an adjunct to metronidazole. | There was no significant difference in the overall 30-day total cure rate between the adjunctive probiotic group and the metronidazole group. There was also no significant difference in vaginal microbial diversity and structure between the two groups at 0, 30, or 90 days. The probiotic species were rarely detected in either the vaginal microbiota or the fecal microbiota after administration, which may reveal the cause of the non-effectiveness of oral probiotics. |
| Park et al. [166] | RCT. N = 101 (19–50 years) females with a Nugent score of 4–6. | MED-01 oral probiotic capsules: Ligilactobacillus salivarius strain MG242, L. fermentum strain MG901, L. plantarum strain MG989, Lacticaseibacillus paracasei strain MG4272, and L. rhamnosus strain MG4288 at 1.0 × 109 CFU/capsule each. Treatment for 12 weeks. | Quantitative PCR analysis confirmed that L. plantarum was significantly increased in the vagina, whereas harmful bacteria such as Mobiluncus spp., B. vaginale, and F. vaginae were suppressed after 12 weeks of MED-01 supplementation. These results confirmed that MED-01 can be used as a probiotic for the treatment of BV by improving VM. |
| Tomusiak et al. [167] | RCT. N = 376 (18–40 years) females with a Nugent score of 4–6. | inVag(®) vaginal gelatin capsules: contained >109 CFU/capsule with 25% L. fermentum strain 57A, 25% L. plantarum strain 57B, and 50% L. paragasseri strain 57C. Treatment for 3 months. | The probiotic inVag was safely administered to sustainably restore the healthy VM with a significant increase in the abundance of Lactobacillus. |
| Recine et al. [168] | Prospective case–control. N = 250 (mean age 29.4 years) non-pregnant sexually active women with a diagnosis of BV treated with metronidazole. | NORMOGIN® vaginal tablets: L. rhamnosus strain BMX 54 (containing > 104 CFU/tablet) for 7 months. | Probiotic supplementation with vaginal L. rhamnosus appears to be useful in inhibiting bacterial growth, especially after antibiotic therapy. Patients who underwent prophylactic therapy with NORMOGIN had a significantly lower rate of recurrence compared to patients treated with antibiotics alone. |
| Palma et al. [169] | RCT. N = 117 (>18 years) females with a diagnosis of BV or yeast vaginitis associated with HPV-infection. | Probiotic tablets: L. rhamnosus strain BMX 54, at 104 CFU/tablet for 9 months. | Women in the long-term probiotic-use group were twice as likely as those in the short-term probiotic-use group to resolve HPV-related cytological abnormalities. Complete clearance of HPV was observed in 11.6% of short-term probiotic users compared to 31.2% of long-term users. |
| Marcotte et al. [170] | Prospective–exploratory. N = 39 (18–50 years) females with a diagnosis of BV and treated with triple oral antibiotic (cefixime, doxycycline, and metronidazole). | Vaginal probiotic capsules: L. rhamnosus strain DSM 14,870 and L. paragasseri strain DSM 14869, for 190 days. | Supplementation with vaginal probiotic capsules resulted in colonization of the vagina by the Lactobacillus strains contained in the capsules. Low initial cure rates of BV after a single dose of metronidazole, and the probiotic did not improve BV cure rates or alleviate recurrence, which could be due to treatment failure or very limited power of the study. |
| Mändar et al. [171] | RCT. N = 182 (18–50 years) females with a diagnosis of R-BV and R-VVC. | Vaginal and oral probiotic capsules: L. crispatus strains DSM32717, DSM32720, DSM32718, and DSM32716, at 3 × 1010 CFU/strain/capsule for 3 months. | Both oral and vaginal capsules effectively improved symptoms in patients with R-BV, including reducing discharge volume, odor, and itching/irritation. In patients with R-VVC, both types of capsules successfully relieved the main symptoms of discharge volume and itching/irritation. |
| Russo et al. [172] | Clinical trial. N = 48 females positive for C. albicans and symptoms of VVC after clotrimazole therapy. | Oral synbiotic formulation: L. acidophilus strain GLA-14 and L. rhamnosus strain HN001 plus bovine lactoferrin, for 3–6 months. | After clotrimazole treatment, a significant improvement in symptoms was shown in both groups (control and intervention). However, only women treated with the synbiotic formulation showed a significant improvement in itching and discharge at 3 and 6 months. During the 6-month follow-up, there were significantly fewer relapses in the intervention group compared to placebo. |
| Lyra et al. [173] | RCT. N = 50 (18–50 years) women devoid of vaginal complaints (Nugent score 0–3 and vaginal pH < 4.5). | V-Caps HMPC synbiotic capsules: Capsules: contained 1010 CFU of L. acidophilus strain La-14 and L. rhamnosus strain HN001 in a 4:1 ratio, potato maltodextrin as a carrier, and stearate and silicon dioxide as flow agents. Treatment for 3 weeks. | VM remained stable and dominated by lactobacilli throughout the intervention, and vaginal pH remained optimal (pH of 4.0–4.5). Immune markers elafin and human β-defensin 3 were significantly decreased in the verum group but did not correlate with changes in the microbiota. |
| Vicariotto et al. [174] | Prospective–observational. N = 50 (45–65 years) postmenopausal healthy women. | Synbiotic capsules: L. plantarum strain PBS067, Bifidobacterium animalis subsp. lactis strain BL050, and L. rhamnosus strain LRH020, at 3 × 109 CFU/g (1 × 109 CFU/g of each probiotic strain), plus vitamin B3 and maltodextrin during 28 days. | Significant improvements in the VHI score and substantial reductions in inflammatory cytokine levels (IL-6, IL-1β, and TNF-α) were achieved. In addition, the probiotic intervention facilitated the restoration of the VM, evidenced by an increase in the abundance of lactobacilli. |
| Baldacci et al. [175] | Clinical trial. N ≥ 3000 females with R-BV after metronidazole treatment. | NORMOGIN™ synbiotic vaginal tablets: L. rhamnosus strain BMX 54 plus lactose, for 6 months. | NORMOGIN has been shown in a large sample of women enrolled in clinical trials not only to be able to significantly reduce the BV recurrences after the standard of care administration but also to control the vaginal pathobiosis pathway, restoring the physiological eubiosis from dysbiosis. |
| Vivekanandan et al. [176] | RCT. N = 66 (≥40 and ≤65 years) females with BV symptoms. | VagiBIOM synbiotic suppositories: 1–2% L. crispatus strain Bi16, L. paragasseri strain Bi19, Heyndrickxia coagulans (formerly Bacillus coagulans) strain Bi34, L. acidophilus strain Bi14, coconut oil fatty acids, prebiotic complex (0.05–2%), hyaluronic acid (0.1–0.3%), silica gel (0.1–0.3%), and lactic acid (0.01–0.025%). Treatment for 4 weeks. | After 4 weeks of intervention with VagiBIOM or a placebo, the mean scores for vaginal pH, VAS itch, and Nugent total score were significantly reduced from the baseline. Compared to the baseline scores, the VHI scores improved significantly after 28 days of intervention. In addition, VagiBIOM improved vaginal lactobacilli diversity. |
| Lev-Sagie et al. [177] | Exploratory. Recipients N = 5 (18–50 years) females with a diagnosis of R-BV. | VMT: Donors N = 2, one was non-sexually active for 8 years and other was in a 25-year monogamous relationship. Donors had a negative history of vaginal symptoms and underwent an examination to verify the absence of BV. Follow-up at 5–21 months after VMT. | Of the five patients treated, VMT was associated with complete long-term remission, defined as significant improvements in symptoms and reconstitution of a Lactobacillus-dominated VM in four patients to the end of follow-up. No adverse effects were observed in any of the five women. Notably, remission in three patients required repeated VMT, including a donor change in one patient, to achieve a durable clinical response. |
| Wrønding et al. [178] | Case study. Recipient N = 1 (30 years) females with vaginal microbiome dysbiosis. | VMT: Transfer of cervicovaginal secretions from a healthy donor with a Lactobacillus-dominant vaginal microbiome. | After one VMT, there was a complete shift in microbiome composition to 81.2% L. crispatus and 9% L. mulieris with concomitant resolution of vaginal symptoms. |
| Oerlemans et al. [179] | Interventional. N = 20 (18–50 years) females with a diagnosis of VVC and treatment with fluconazole. | Probiotic gel: L. plantarum strain WCFS1, Lactiplantibacillus pentosus strain KCA1, and L. rhamnosus strain GG, at 109–1010 CFU/g for 4 weeks. | Forty-five percent of women using the probiotic gel experienced symptom relief without the need for additional fluconazole medication, and their fungal levels were similar to those of women treated with fluconazole. Fluconazole alone reduced the abundance of vaginal lactobacilli. |
| Yoshikata et al. [180] | RCT. N = 70 (20–40 years) females with atrophic vaginitis, BV, and genitourinary symptoms. | Foaming wash (Delicate Softwash®), feminine cream (Delicate Softgelcream®) and vaginal gel (Inner gel®): L. rhamnosus strain vitaP1 and L. plantarum strain KCTC3108. Treatment for 4 weeks. | Vaginal pH and pathogenic microorganisms were reduced in both treatment groups (post- and premenopausal women) compared to the control group, which was more significant in the postmenopausal group. Genitourinary symptoms improved significantly in 60% of premenopausal women and 81.3% of postmenopausal women compared to the control group. |
| Shen et al. [181] | Human clinical trial. N = 50 (18–55 years) females with diagnosis of BV. | Postbiotic gel: L. paracasei strain ProSci-92 and L. rhamnosus strain ProSci-109, at 3 × 109 CFU/g of each probiotic strain, plus 5% of skim milk powder and 3% full-fat soybean. Treatment for 1 week. | The results showed that the application of the synbiotic gel improved the symptoms of BV, as indicated by an improvement in the abnormalities of the patients’ vaginal secretions. After application of the gel, the relative abundance of vaginal lactobacilli increased compared to baseline. Significant negative correlations were found between lactobacilli and potential vaginal pathogens (including Gardnerella, Prevotella, and Fannyhessea), as well as vaginal secretion abnormalities. |
6. Discussion
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| VM | Vaginal microbiome |
| AMPs | Antimicrobial peptides |
| BV | Bacterial vaginosis |
| STIs | Sexually transmitted infections |
| AV | Aerobic vaginitis |
| PID | Pelvic inflammatory disease |
| VVC | Vulvovaginal candidiasis |
| LBPs | Live biotherapeutic products |
| qPCR | Quantitative real-time PCR |
| HIV | Human immunodeficiency virus |
| HPV | Human papillomavirus |
| DCs | Dendritic cells |
| DAMPs | Damage-associated molecular patterns |
| C system | Complement system |
| PRRs | Pattern recognition receptors |
| TLRs | Toll-like receptors |
| NOD | Nucleotide-binding oligomerization domain |
| TNF-α | Tumor necrosis factor-α |
| PAMPs | Pathogen-associated molecular patterns |
| NF-κB | Nuclear factor-κB |
| GBS | Group B Streptococcus agalactiae |
| SCFAs | Short-chain fatty acids |
| RCT | Randomized controlled trial |
| VMT | Vaginal microbiota transplantation |
| CFUs | Colony-forming units |
| R-BV | Recurrent bacterial vaginosis |
| R-VVC | Recurrent vulvovaginal candidiasis |
| VAS | Visual analog scale |
| VHI | Vaginal health index |
References
- Smith, S.B.; Ravel, J. The vaginal microbiota, host defense, and reproductive physiology. J. Physiol. 2017, 595, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Greenbaum, S.; Greenbaum, G.; Moran-Gilad, J.; Weintraub, A.Y. Ecological dynamics of the vaginal microbiome in relation to health and disease. Am. J. Obstet. Gynecol. 2019, 220, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108, 4680–4687. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Forney, L.J.; Ravel, J. Vaginal microbiome: Rethinking health and disease. Annu. Rev. Microbiol. 2012, 66, 371–389. [Google Scholar] [CrossRef]
- Pekmezovic, M.; Mogavero, S.; Naglik, J.R.; Hube, B. Host-Pathogen Interactions during Female Genital Tract Infections. Trends Microbiol. 2019, 27, 982–996. [Google Scholar] [CrossRef]
- Chee, W.J.Y.; Chew, S.Y.; Than, L.T.L. Vaginal microbiota and the potential of Lactobacillus derivatives in maintaining vaginal health. Microb. Cell Fact. 2020, 19, 203. [Google Scholar] [CrossRef]
- Christensen, A.O.; Li, G.; Young, C.H.; Snow, B.; Khan, S.A.; DeVore, S.B.; Edwards, S.; Bouma, G.J.; Navratil, A.M.; Cherrington, B.D.; et al. Peptidylarginine deiminase enzymes and citrullinated proteins in female reproductive physiology and associated diseases. Biol. Reprod. 2022, 107, 1395–1410. [Google Scholar] [CrossRef]
- Fuochi, V.; Cardile, V.; Petronio Petronio, G.; Furneri, P.M. Biological properties and production of bacteriocins-like-inhibitory substances by Lactobacillus sp. strains from human vagina. J. Appl. Microbiol. 2019, 126, 1541–1550. [Google Scholar] [CrossRef]
- Miko, E.; Barakonyi, A. The Role of Hydrogen-Peroxide (H2O2) Produced by Vaginal Microbiota in Female Reproductive Health. Antioxidants 2023, 12, 1055. [Google Scholar] [CrossRef]
- Vagios, S.; Mitchell, C.M. Mutual Preservation: A Review of Interactions Between Cervicovaginal Mucus and Microbiota. Front. Cell. Infect. Microbiol. 2021, 11, 676114. [Google Scholar] [CrossRef]
- Vornhagen, J.; Quach, P.; Santana-Ufret, V.; Alishetti, V.; Brokaw, A.; Armistead, B.; Qing Tang, H.; MacDonald, J.W.; Bammler, T.K.; Adams Waldorf, K.M.; et al. Human Cervical Mucus Plugs Exhibit Insufficiencies in Antimicrobial Activity Towards Group B Streptococcus. J. Infect. Dis. 2018, 217, 1626–1636. [Google Scholar] [CrossRef] [PubMed]
- Yarbrough, V.L.; Winkle, S.; Herbst-Kralovetz, M.M. Antimicrobial peptides in the female reproductive tract: A critical component of the mucosal immune barrier with physiological and clinical implications. Hum. Reprod. Update 2015, 21, 353–377. [Google Scholar] [CrossRef] [PubMed]
- Gutzeit, O.; Gulati, A.; Izadifar, Z.; Stejskalova, A.; Rhbiny, H.; Cotton, J.; Budnik, B.; Shahriar, S.; Goyal, G.; Junaid, A.; et al. Cervical mucus in linked human Cervix and Vagina Chips modulates vaginal dysbiosis. NPJ Womens Health 2025, 3, 5. [Google Scholar] [CrossRef] [PubMed]
- Holdcroft, A.M.; Ireland, D.J.; Payne, M.S. The Vaginal Microbiome in Health and Disease—What Role Do Common Intimate Hygiene Practices Play? Microorganisms 2023, 11, 298. [Google Scholar] [CrossRef]
- Kalia, N.; Singh, J.; Kaur, M. Microbiota in vaginal health and pathogenesis of recurrent vulvovaginal infections: A critical review. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 5. [Google Scholar] [CrossRef]
- Shen, L.; Zhang, W.; Yuan, Y.; Zhu, W.; Shang, A. Vaginal microecological characteristics of women in different physiological and pathological period. Front. Cell. Infect. Microbiol. 2022, 12, 959793. [Google Scholar] [CrossRef]
- Gajer, P.; Brotman, R.M.; Bai, G.; Sakamoto, J.; Schütte, U.M.; Zhong, X.; Koenig, S.S.; Fu, L.; Ma, Z.S.; Zhou, X.; et al. Temporal dynamics of the human vaginal microbiota. Sci. Transl. Med. 2012, 4, 132ra52. [Google Scholar] [CrossRef]
- Sukhera, J. Narrative Reviews: Flexible, Rigorous, and Practical. J. Grad. Med. Educ. 2022, 14, 414–417. [Google Scholar] [CrossRef]
- Zheng, N.; Guo, R.; Wang, J.; Zhou, W.; Ling, Z. Contribution of Lactobacillus iners to Vaginal Health and Diseases: A Systematic Review. Front. Cell. Infect. Microbiol. 2021, 11, 792787. [Google Scholar] [CrossRef]
- Abou Chacra, L.; Fenollar, F.; Diop, K. Bacterial Vaginosis: What Do We Currently Know? Front. Cell. Infect. Microbiol. 2022, 11, 672429. [Google Scholar] [CrossRef]
- Carter, K.A.; Fischer, M.D.; Petrova, M.I.; Balkus, J.E. Epidemiologic Evidence on the Role of Lactobacillus iners in Sexually Transmitted Infections and Bacterial Vaginosis: A Series of Systematic Reviews and Meta-Analyses. Sex. Transm. Dis. 2023, 50, 224–235. [Google Scholar] [CrossRef]
- Lev-Sagie, A.; De Seta, F.; Verstraelen, H.; Ventolini, G.; Lonnee-Hoffmann, R.; Vieira-Baptista, P. The Vaginal Microbiome: II. Vaginal Dysbiotic Conditions. J. Low. Genit. Tract Dis. 2022, 26, 79–84. [Google Scholar] [CrossRef]
- Muzny, C.A.; Taylor, C.M.; Swords, W.E.; Tamhane, A.; Chattopadhyay, D.; Cerca, N.; Schwebke, J.R. An Updated Conceptual Model on the Pathogenesis of Bacterial Vaginosis. J. Infect. Dis. 2019, 220, 1399–1405. [Google Scholar] [CrossRef]
- Osei Sekyere, J.; Oyenihi, A.B.; Trama, J.; Adelson, M.E. Species-Specific Analysis of Bacterial Vaginosis-Associated Bacteria. Microbiol. Spectr. 2023, 11, e0467622. [Google Scholar] [CrossRef]
- Diop, K.; Bretelle, F.; Raoult, D.; Fenollar, F. ‘Corynebacterium fournierii’, a new bacterial species isolated from the vaginal sample of a patient with bacterial vaginosis. New Microbes New Infect. 2017, 18, 6–7. [Google Scholar] [CrossRef]
- Diop, K.; Diop, A.; Bretelle, F.; Cadoret, F.; Michelle, C.; Richez, M.; Cocallemen, J.F.; Raoult, D.; Fournier, P.E.; Fenollar, F. Olegusella massiliensis gen. nov., sp. nov., strain KHD7T, a new bacterial genus isolated from the female genital tract of a patient with bacterial vaginosis. Anaerobe 2017, 44, 87–95. [Google Scholar] [CrossRef]
- Diop, K.; Raoult, D.; Bretelle, F.; Fenollar, F. “Ezakiella massiliensis” sp. nov., a new bacterial species isolated from human female genital tract. New Microbes New Infect. 2017, 15, 16–17. [Google Scholar] [CrossRef]
- Swidsinski, S.; Moll, W.M.; Swidsinski, A. Bacterial Vaginosis—Vaginal Polymicrobial Biofilms and Dysbiosis. Dtsch. Arztebl. Int. 2023, 120, 347–354. [Google Scholar] [CrossRef]
- Segui-Perez, C.; de Jongh, R.; Jonkergouw, R.L.W.; Pelayo, P.; Balskus, E.P.; Zomer, A.; Strijbis, K. Prevotella timonensis degrades the vaginal epithelial glycocalyx through high fucosidase and sialidase activities. mBio 2024, 15, e0069124. [Google Scholar] [CrossRef]
- Mondal, A.S.; Sharma, R.; Trivedi, N. Bacterial vaginosis: A state of microbial dysbiosis. Med. Microecol. 2023, 16, 100082. [Google Scholar] [CrossRef]
- Nelson, T.M.; Borgogna, J.L.; Brotman, R.M.; Ravel, J.; Walk, S.T.; Yeoman, C.J. Vaginal biogenic amines: Biomarkers of bacterial vaginosis or precursors to vaginal dysbiosis? Front. Physiol. 2015, 6, 253. [Google Scholar] [CrossRef] [PubMed]
- Amegashie, C.P.; Gilbert, N.M.; Peipert, J.F.; Allsworth, J.E.; Lewis, W.G.; Lewis, A.L. Relationship between Nugent score and vaginal epithelial exfoliation. PLoS ONE 2017, 12, e0177797. [Google Scholar] [CrossRef] [PubMed]
- Machado, A.; Cerca, N. Influence of Biofilm Formation by Gardnerella vaginalis and Other Anaerobes on Bacterial Vaginosis. J. Infect. Dis. 2015, 212, 1856–1861. [Google Scholar] [CrossRef] [PubMed]
- Swidsinski, A.; Loening-Baucke, V.; Swidsinski, S.; Sobel, J.D.; Dörffel, Y.; Guschin, A. Clue Cells and Pseudo Clue Cells in Different Morphotypes of Bacterial Vaginosis. Front. Cell. Infect. Microbiol. 2022, 12, 905739. [Google Scholar] [CrossRef]
- Amsel, R.; Totten, P.A.; Spiegel, C.A.; Chen, K.C.S.; Eschenbach, D.; Holmes, K.K. Nonspecific vaginitis. Diagnostic criteria and microbial and epidemiologic associations. Am. J. Med. 1983, 74, 14–22. [Google Scholar] [CrossRef]
- Nugent, R.P.; Krohn, M.A.; Hillier, S.L. Reliability of diagnosing bacterial vaginosis is improved by a standardized method of gram stain interpretation. J. Clin. Microbiol. 1991, 29, 297–301. [Google Scholar] [CrossRef]
- Ison, C.A.; Hay, P.E. Validation of a simplified grading of Gram stained vaginal smears for use in genitourinary medicine clinics. Sex. Transm. Infect. 2002, 78, 413–415. [Google Scholar] [CrossRef]
- Sherrard, J.; Wilson, J.; Donders, G.; Mendling, W.; Jensen, J.S. 2018 European (IUSTI/WHO) International Union against sexually transmitted infections (IUSTI) World Health Organisation (WHO) guideline on the management of vaginal discharge. Int. J. STD AIDS. 2018, 29, 1258–1725. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, L.; Zhao, M.; Wang, Y.; Bai, H.; Wang, Y.; Rui, C.; Fan, C.; Li, J.; Li, N.; et al. Deep Neural Networks Offer Morphologic Classification and Diagnosis of Bacterial Vaginosis. J. Clin. Microbiol. 2021, 59, e02236-20. [Google Scholar] [CrossRef]
- Cartwright, C.P.; Pherson, A.J.; Harris, A.B.; Clancey, M.S.; Nye, M.B. Multicenter study establishing the clinical validity of a nucleic-acid amplification–based assay for the diagnosis of bacterial vaginosis. Diagn. Microbiol. Infect. Dis. 2018, 92, 173–178. [Google Scholar] [CrossRef]
- Gaydos, C.A.; Beqaj, S.; Schwebke, J.R.; Lebed, J.; Smith, B.; Davis, T.E.; Fife, K.H.; Nyirjesy, P.; Spurrell, T.; Furgerson, D.; et al. Clinical Validation of a Test for the Diagnosis of Vaginitis. Obstet. Gynecol. 2017, 130, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Hilbert, D.W.; Smith, W.L.; Chadwick, S.G.; Toner, G.; Mordechai, E.; Adelson, M.E.; Aguin, T.J.; Sobel, J.D.; Gygax, S.E. Development and Validation of a Highly Accurate Quantitative Real-Time PCR Assay for Diagnosis of Bacterial Vaginosis. J. Clin. Microbiol. 2016, 54, 1017–1024. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.S.; Gaydos, C.A. Molecular Diagnosis of Bacterial Vaginosis: An Update. J. Clin. Microbiol. 2018, 56, e00342-18. [Google Scholar] [CrossRef]
- Azeredo, J.; Azevedo, N.F.; Briandet, R.; Cerca, N.; Coenye, T.; Costa, A.R.; Desvaux, M.; Di Bonaventura, G.; Hébraud, M.; Jaglic, Z.; et al. Critical review on biofilm methods. Crit. Rev. Microbiol. 2017, 43, 313–351. [Google Scholar] [CrossRef]
- Shipitsyna, E.; Roos, A.; Datcu, R.; Hallén, A.; Fredlund, H.; Jensen, J.S.; Engstrand, L.; Unemo, M. Composition of the vaginal microbiota in women of reproductive age—Sensitive and specific molecular diagnosis of bacterial vaginosis is possible? PLoS ONE 2013, 8, e60670. [Google Scholar] [CrossRef]
- Breding, K.; Selbing, A.; Farnebäck, M. Diagnosis of Bacterial Vaginosis Using a Novel Molecular Real-Time PCR Test. J. Womens Health Gynecol. 2020, 7, 1–7. [Google Scholar]
- van den Munckhof, E.H.A.; van Sitter, R.L.; Boers, K.E.; Lamont, R.F.; Te Witt, R.; le Cessie, S.; Knetsch, C.W.; van Doorn, L.J.; Quint, W.G.V.; Molijn, A.; et al. Comparison of Amsel criteria, Nugent score, culture and two CE-IVD marked quantitative real-time PCRs with microbiota analysis for the diagnosis of bacterial vaginosis. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 959–966. [Google Scholar] [CrossRef]
- Madhivanan, P.; Krupp, K.; Li, T.; Ravi, K.; Selezneva, J.; Srinivas, V.; Arun, A.; Klausner, J.D. Performance of BVBlue rapid test in detecting bacterial vaginosis among women in Mysore, India. Infect. Dis. Obstet. Gynecol. 2014, 2014, 908313. [Google Scholar] [CrossRef]
- Liu, G.J.; Wang, B.; Zhang, Y.; Xing, G.W.; Yang, X.; Wang, S. A tetravalent sialic acid-coated tetraphenylethene luminogen with aggregation-induced emission characteristics: Design, synthesis and application for sialidase activity assay, high-throughput screening of sialidase inhibitors and diagnosis of bacterial vaginosis. Chem. Commun. 2018, 54, 10691–10694. [Google Scholar]
- Rodríguez-Nava, C.; Cortés-Sarabia, K.; Avila-Huerta, M.D.; Ortiz-Riaño, E.J.; Estrada-Moreno, A.K.; Alarcón-Romero, L.D.C.; Mata-Ruíz, O.; Medina-Flores, Y.; Vences-Velázquez, A.; Morales-Narváez, E. Nanophotonic Sialidase Immunoassay for Bacterial Vaginosis Diagnosis. ACS Pharmacol. Transl. Sci. 2021, 4, 365–371. [Google Scholar] [CrossRef]
- Swidsinski, A.; Mendling, W.; Loening-Baucke, V.; Ladhoff, A.; Swidsinski, S.; Hale, L.P.; Lochs, H. Adherent biofilms in bacterial vaginosis. Obstet. Gynecol. 2005, 106, 1013–1023. [Google Scholar] [CrossRef] [PubMed]
- Hickey, R.J.; Forney, L.J. Gardnerella vaginalis does not always cause bacterial vaginosis. J. Infect. Dis. 2014, 210, 1682–1683. [Google Scholar] [CrossRef]
- Kumar, N.; Behera, B.; Sagiri, S.S.; Pal, K.; Ray, S.S.; Roy, S. Bacterial vaginosis: Etiology and modalities of treatment—A brief note. J. Pharm. Bioallied Sci. 2011, 3, 496. [Google Scholar]
- Muzny, C.A.; Blanchard, E.; Taylor, C.M.; Aaron, K.J.; Talluri, R.; Griswold, M.E.; Redden, D.T.; Luo, M.; Welsh, D.A.; Van Der Pol, W.J.; et al. Identification of Key Bacteria Involved in the Induction of Incident Bacterial Vaginosis: A Prospective Study. J. Infect. Dis. 2018, 218, 966–978. [Google Scholar] [CrossRef] [PubMed]
- Swidsinski, A.; Doerffel, Y.; Loening-Baucke, V.; Swidsinski, S.; Verstraelen, H.; Vaneechoutte, M.; Lemm, V.; Schilling, J.; Mendling, W. Gardnerella biofilm involves females and males and is transmitted sexually. Gynecol. Obstet. Investigat. 2010, 70, 256–263. [Google Scholar] [CrossRef]
- Castro, J.; Machado, D.; Cerca, N. Unveiling the role of Gardnerella vaginalis in polymicrobial Bacterial Vaginosis biofilms: The impact of other vaginal pathogens living as neighbors. ISME J. 2019, 13, 1306–1317. [Google Scholar] [CrossRef]
- Hardy, L.; Jespers, V.; Abdellati, S.; De Baetselier, I.; Mwambarangwe, L.; Musengamana, V.; van de Wijgert, J.; Vaneechoutte, M.; Crucitti, T. A fruitful alliance: The synergy between Atopobium vaginae and Gardnerella vaginalis in bacterial vaginosis-associated biofilm. Sex. Transm. Infect. 2016, 92, 487–491. [Google Scholar] [CrossRef]
- Kalia, N.; Singh, J.; Kaur, M. Immunopathology of Recurrent Vulvovaginal Infections: New Aspects and Research Directions. Front. Immunol. 2019, 10, 2034. [Google Scholar] [CrossRef]
- Gholiof, M.; Adamson-De Luca, E.; Wessels, J.M. The female reproductive tract microbiotas, inflammation, and gynecological conditions. Front. Reprod. Health. 2022, 4, 963752. [Google Scholar] [CrossRef]
- Villa, P.; Cipolla, C.; D’Ippolito, S.; Amar, I.D.; Shachor, M.; Ingravalle, F.; Scaldaferri, F.; Puca, P.; Di Simone, N.; Scambia, G. The interplay between immune system and microbiota in gynecological diseases: A narrative review. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 5676–5690. [Google Scholar]
- Anahtar, M.N.; Byrne, E.H.; Doherty, K.E.; Bowman, B.A.; Yamamoto, H.S.; Soumillon, M.; Padavattan, N.; Ismail, N.; Moodley, A.; Sabatini, M.E.; et al. Cervicovaginal bacteria are a major modulator of host inflammatory responses in the female genital tract. Immunity 2015, 42, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Pellis, V.; De Seta, F.; Crovella, S.; Bossi, F.; Bulla, R.; Guaschino, S.; Radillo, O.; Garred, P.; Tedesco, F. Mannose binding lectin and C3 act as recognition molecules for infectious agents in the vagina. Clin. Exp. Immunol. 2005, 139, 120–126. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, R.; Li, M.; Zeng, Z.; Zhang, L.; Liao, Q. The interplay between microbiota, metabolites, immunity during BV. Med. Microecol. 2022, 11, 100049. [Google Scholar] [CrossRef]
- Giraldo, P.C.; de Carvalho, J.B.; do Amaral, R.L.; da Silveira Gonçalves, A.K.; Eleutério, J., Jr.; Guimarães, F. Identification of immune cells by flow cytometry in vaginal lavages from women with vulvovaginitis and normal microflora. Am. J. Reprod. Immunol. 2012, 67, 198–205. [Google Scholar] [CrossRef]
- Aldunate, M.; Srbinovski, D.; Hearps, A.C.; Latham, C.F.; Ramsland, P.A.; Gugasyan, R.; Cone, R.A.; Tachedjian, G. Antimicrobial and immune modulatory effects of lactic acid and short chain fatty acids produced by vaginal microbiota associated with eubiosis and bacterial vaginosis. Front. Physiol. 2015, 6, 164. [Google Scholar] [CrossRef]
- Delgado-Diaz, D.J.; Tyssen, D.; Hayward, J.A.; Gugasyan, R.; Hearps, A.C.; Tachedjian, G. Distinct Immune Responses Elicited From Cervicovaginal Epithelial Cells by Lactic Acid and Short Chain Fatty Acids Associated with Optimal and Non-optimal Vaginal Microbiota. Front. Cell. Infect. Microbiol. 2020, 9, 446. [Google Scholar] [CrossRef]
- Lewis, W.G.; Robinson, L.S.; Perry, J.; Bick, J.L.; Peipert, J.F.; Allsworth, J.E.; Lewis, A.L. Hydrolysis of secreted sialoglycoprotein immunoglobulin A (IgA) in ex vivo and biochemical models of bacterial vaginosis. J. Biol. Chem. 2012, 287, 2079–2089. [Google Scholar] [CrossRef]
- Berard, A.R.; Brubaker, D.K.; Birse, K.; Lamont, A.; Mackelprang, R.D.; Noël-Romas, L.; Perner, M.; Hou, X.; Irungu, E.; Mugo, N.; et al. Vaginal epithelial dysfunction is mediated by the microbiome, metabolome, and mTOR signaling. Cell Rep. 2023, 42, 112474. [Google Scholar] [CrossRef]
- Donders, G.G.G.; Bellen, G.; Grinceviciene, S.; Ruban, K.; Vieira-Baptista, P. Aerobic vaginitis: No longer a stranger. Res. Microbiol. 2017, 168, 845–858. [Google Scholar] [CrossRef]
- Turpin, R.; Tuddenham, S.; He, X.; Klebanoff, M.A.; Ghanem, K.G.; Brotman, R.M. Bacterial Vaginosis and Behavioral Factors Associated with Incident Pelvic Inflammatory Disease in the Longitudinal Study of Vaginal Flora. J. Infect. Dis. 2021, 224, S137–S144. [Google Scholar] [CrossRef]
- Willems, H.M.E.; Ahmed, S.S.; Liu, J.; Xu, Z.; Peters, B.M. Vulvovaginal Candidiasis: A Current Understanding and Burning Questions. J. Fungi. 2020, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Norenhag, J.; Hu, Y.O.O.; Brusselaers, N.; Fransson, E.; Ährlund-Richter, A.; Guðnadóttir, U.; Angelidou, P.; Zha, Y.; Hamsten, M.; et al. Vaginal microbiota and human papillomavirus infection among young Swedish women. NPJ Biofilms Microbiomes 2020, 6, 39. [Google Scholar] [CrossRef] [PubMed]
- Cherenack, E.M.; Broedlow, C.A.; Klatt, N.R. The vaginal microbiome and HIV transmission dynamics. Curr. Opin. HIV AIDS. 2024, 19, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Lokken, E.M.; Balkus, J.E.; Kiarie, J.; Hughes, J.P.; Jaoko, W.; Totten, P.A.; McClelland, R.S.; Manhart, L.E. Association of Recent Bacterial Vaginosis with Acquisition of Mycoplasma genitalium. Am. J. Epidemiol. 2017, 186, 194–201. [Google Scholar] [CrossRef]
- Santella, B.; Schettino, M.T.; Franci, G.; De Franciscis, P.; Colacurci, N.; Schiattarella, A.; Galdiero, M. Microbiota and HPV: The role of viral infection on vaginal microbiota. J. Med. Virol. 2022, 94, 4478–4484. [Google Scholar] [CrossRef]
- Gudnadottir, U.; Debelius, J.W.; Du, J.; Hugerth, L.W.; Danielsson, H.; Schuppe-Koistinen, I.; Fransson, E.; Brusselaers, N. The vaginal microbiome and the risk of preterm birth: A systematic review and network meta-analysis. Sci. Rep. 2022, 12, 7926. [Google Scholar] [CrossRef]
- Kaambo, E.; Africa, C.; Chambuso, R.; Passmore, J.S. Vaginal Microbiomes Associated with Aerobic Vaginitis and Bacterial Vaginosis. Front. Public Health 2018, 6, 78. [Google Scholar] [CrossRef]
- Tao, Z.; Zhang, L.; Zhang, Q.; Lv, T.; Chen, R.; Wang, L.; Huang, Z.; Hu, L.; Liao, Q. The Pathogenesis of Streptococcus anginosus in Aerobic Vaginitis. Infect. Drug Resist. 2019, 12, 3745–3754. [Google Scholar] [CrossRef]
- Wang, C.; Fan, A.; Li, H.; Yan, Y.; Qi, W.; Wang, Y.; Han, C.; Xue, F. Vaginal bacterial profiles of aerobic vaginitis: A case-control study. Diagn. Microbiol. Infect. Dis. 2020, 96, 114981. [Google Scholar] [CrossRef]
- Fan, A.; Yue, Y.; Geng, N.; Zhang, H.; Wang, Y.; Xue, F. Aerobic vaginitis and mixed infections: Comparison of clinical and laboratory findings. Arch. Gynecol. Obstet. 2013, 287, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Oerlemans, E.F.M.; Wuyts, S.; Bellen, G.; Wittouck, S.; De Boeck, I.; Ruban, K.; Allonsius, C.N.; van den Broek, M.F.L.; Donders, G.G.G.; Lebeer, S. The Dwindling Microbiota of Aerobic Vaginitis, an Inflammatory State Enriched in Pathobionts with Limited TLR Stimulation. Diagnostics. 2020, 10, 879. [Google Scholar] [CrossRef] [PubMed]
- Stika, C.S. Atrophic vaginitis. Dermatol. Ther. 2010, 23, 514–522. [Google Scholar] [CrossRef]
- Krauss-Silva, L.; Almada-Horta, A.; Alves, M.B.; Camacho, K.G.; Moreira, M.E.; Braga, A. Basic vaginal pH, bacterial vaginosis and aerobic vaginitis: Prevalence in early pregnancy and risk of spontaneous preterm delivery, a prospective study in a low socioeconomic and multiethnic South American population. BMC Pregnancy Childbirth 2014, 4, 107. [Google Scholar] [CrossRef]
- Valeriano, V.D.; Lahtinen, E.; Hwang, I.C.; Zhang, Y.; Du, J.; Schuppe-Koistinen, I. Vaginal dysbiosis and the potential of vaginal microbiome-directed therapeutics. Front. Microbiomes 2024, 3, 1363089. [Google Scholar] [CrossRef]
- den Heijer, C.D.J.; Hoebe, C.J.P.A.; Driessen, J.H.M.; Wolffs, P.; van den Broek, I.V.F.; Hoenderboom, B.M.; Williams, R.; de Vries, F.; Dukers-Muijrers, N.H.T.M. Chlamydia trachomatis and the Risk of Pelvic Inflammatory Disease, Ectopic Pregnancy, and Female Infertility: A Retrospective Cohort Study Among Primary Care Patients. Clin. Infect. Dis. 2019, 69, 1517–1525. [Google Scholar] [CrossRef]
- Hoenderboom, B.M.; van Benthem, B.H.B.; van Bergen, J.E.A.M.; Dukers-Muijrers, N.H.T.M.; Götz, H.M.; Hoebe, C.J.P.A.; Hogewoning, A.A.; Land, J.A.; van der Sande, M.A.B.; Morré, S.A.; et al. Relation between Chlamydia trachomatis infection and pelvic inflammatory disease, ectopic pregnancy and tubal factor infertility in a Dutch cohort of women previously tested for chlamydia in a chlamydia screening trial. Sex. Transm. Infect. 2019, 95, 300–306. [Google Scholar]
- Price, M.J.; Ades, A.E.; Welton, N.J.; Simms, I.; Macleod, J.; Horner, P.J. Proportion of Pelvic Inflammatory Disease Cases Caused by Chlamydia trachomatis: Consistent Picture from Different Methods. J. Infect. Dis. 2016, 214, 617–624. [Google Scholar] [CrossRef]
- Haggerty, C.L.; Taylor, B.D. Mycoplasma genitalium: An emerging cause of pelvic inflammatory disease. Infect. Dis. Obstet. Gynecol. 2011, 2011, 959816. [Google Scholar] [CrossRef]
- Xu, S.X.; Gray-Owen, S.D. Gonococcal Pelvic Inflammatory Disease: Placing Mechanistic Insights Into the Context of Clinical and Epidemiological Observations. J. Infect. Dis. 2021, 224, S56–S63. [Google Scholar] [CrossRef]
- Ravel, J.; Moreno, I.; Simón, C. Bacterial vaginosis and its association with infertility, endometritis, and pelvic inflammatory disease. Am. J. Obstet. Gynecol. 2021, 224, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Tamarelle, J.; Thiébaut, A.C.M.; de Barbeyrac, B.; Bébéar, C.; Ravel, J.; Delarocque-Astagneau, E. The vaginal microbiota and its association with human papillomavirus, Chlamydia trachomatis, Neisseria gonorrhoeae and Mycoplasma genitalium infections: A systematic review and meta-analysis. Clin. Microbiol. Infect. 2019, 25, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Kerry-Barnard, S.; Zhou, L.; Phillips, L.; Furegato, M.; Witney, A.A.; Sadiq, S.T.; Oakeshott, P. Vaginal microbiota in ethnically diverse young women who did or did not develop pelvic inflammatory disease: Community-based prospective study. Sex. Transm. Infect. 2022, 98, 503–509. [Google Scholar] [CrossRef]
- Taylor, B.D.; Darville, T.; Haggerty, C.L. Does bacterial vaginosis cause pelvic inflammatory disease? Sex. Transm. Dis. 2013, 40, 117–122. [Google Scholar] [CrossRef]
- Nyirjesy, P.; Brookhart, C.; Lazenby, G.; Schwebke, J.; Sobel, J.D. Vulvovaginal Candidiasis: A Review of the Evidence for the 2021 Centers for Disease Control and Prevention of Sexually Transmitted Infections Treatment Guidelines. Clin. Infect. Dis. 2022, 74, S162–S168. [Google Scholar] [CrossRef]
- Denning, D.W.; Kneale, M.; Sobel, J.D.; Rautemaa-Richardson, R. Global burden of recurrent vulvovaginal candidiasis: A systematic review. Lancet Infect. Dis. 2018, 18, e339–e347. [Google Scholar] [CrossRef]
- Fukazawa, E.I.; Witkin, S.S.; Robial, R.; Vinagre, J.G.; Baracat, E.C.; Linhares, I.M. Influence of recurrent vulvovaginal candidiasis on quality of life issues. Arch. Gynecol. Obstet. 2019, 300, 647–650. [Google Scholar] [CrossRef]
- Neal, C.M.; Martens, M.G. Clinical challenges in diagnosis and treatment of recurrent vulvovaginal candidiasis. SAGE Open Med. 2022, 10, 20503121221115201. [Google Scholar] [CrossRef]
- Sobel, J.D. Recurrent vulvovaginal candidiasis. Am. J. Obstet. Gynecol. 2016, 214, 15–21. [Google Scholar] [CrossRef]
- Rosati, D.; Bruno, M.; Jaeger, M.; Ten Oever, J.; Netea, M.G. Recurrent Vulvovaginal Candidiasis: An Immunological Perspective. Microorganisms 2020, 8, 144. [Google Scholar] [CrossRef]
- Hong, E.; Dixit, S.; Fidel, P.L.; Bradford, J.; Fischer, G. Vulvovaginal candidiasis as a chronic disease: Diagnostic criteria and definition. J. Low. Genit. Tract Dis. 2014, 18, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Yeater, K.M.; Hoyer, L.L. Cellular and molecular biology of Candida albicans estrogen response. Eukaryot. Cell 2006, 5, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Velloza, J.; Heffron, R. The Vaginal Microbiome and its Potential to Impact Efficacy of HIV Pre-exposure Prophylaxis for Women. Curr. HIV/AIDS Rep. 2017, 14, 153–160. [Google Scholar] [CrossRef]
- France, M.; Alizadeh, M.; Brown, S.; Ma, B.; Ravel, J. Towards a deeper understanding of the vaginal microbiota. Nat. Microbiol. 2022, 7, 367–378. [Google Scholar] [CrossRef]
- Shen, R.; Richter, H.E.; Smith, P.D. Interactions between HIV-1 and mucosal cells in the female reproductive tract. Am. J. Reprod. Immunol. 2014, 71, 608–617. [Google Scholar] [CrossRef]
- Hladik, F.; Sakchalathorn, P.; Ballweber, L.; Lentz, G.; Fialkow, M.; Eschenbach, D.; McElrath, M.J. Initial events in establishing vaginal entry and infection by human immunodeficiency virus type-1. Immunity 2007, 26, 257–270. [Google Scholar] [CrossRef]
- Wang, Y.; Noël-Romas, L.; Perner, M.; Knodel, S.; Molatlhegi, R.; Hoger, S.; Birse, K.; Zuend, C.F.; McKinnon, L.R.; Burgener, A.D. Non-Lactobacillus-Dominant and Polymicrobial Vaginal Microbiomes Are More Common in Younger South African Women and Predictive of Increased Risk of Human Immunodeficiency Virus Acquisition. Clin. Infect. Dis. 2023, 76, 1372–1381. [Google Scholar] [CrossRef]
- Schellenberg, J.J.; Card, C.M.; Ball, T.B.; Mungai, J.N.; Irungu, E.; Kimani, J.; Jaoko, W.; Wachihi, C.; Fowke, K.R.; Plummer, F.A. Bacterial vaginosis, HIV serostatus and T-cell subset distribution in a cohort of East African commercial sex workers: Retrospective analysis. AIDS 2012, 26, 387–393. [Google Scholar] [CrossRef]
- Armstrong, E.; Kaul, R. Beyond bacterial vaginosis: Vaginal lactobacilli and HIV risk. Microbiome 2021, 9, 239. [Google Scholar] [CrossRef]
- Mitra, A.; MacIntyre, D.A.; Marchesi, J.R.; Lee, Y.S.; Bennett, P.R.; Kyrgiou, M. The vaginal microbiota, human papillomavirus infection and cervical intraepithelial neoplasia: What do we know and where are we going next? Microbiome 2016, 4, 58. [Google Scholar] [CrossRef]
- Wang, Y.; Thakur, R.; Shen, Q.; He, Y.; Chen, C. Influences of vaginal microbiota on human papillomavirus infection and host immune regulation: What we have learned? Decoding Infect. Transm. 2023, 1, 100002. [Google Scholar] [CrossRef]
- Di Paola, M.; Sani, C.; Clemente, A.M.; Iossa, A.; Perissi, E.; Castronovo, G.; Tanturli, M.; Rivero, D.; Cozzolino, F.; Cavalieri, D.; et al. Characterization of cervico-vaginal microbiota in women developing persistent high-risk Human Papillomavirus infection. Sci. Rep. 2017, 7, 10200. [Google Scholar] [CrossRef]
- Lee, J.E.; Lee, S.; Lee, H.; Song, Y.M.; Lee, K.; Han, M.J.; Sung, J.; Ko, G. Association of the vaginal microbiota with human papillomavirus infection in a Korean twin cohort. PLoS ONE 2013, 8, e63514. [Google Scholar] [CrossRef]
- Mei, L.; Wang, T.; Chen, Y.; Wei, D.; Zhang, Y.; Cui, T.; Meng, J.; Zhang, X.; Liu, Y.; Ding, L.; et al. Dysbiosis of vaginal microbiota associated with persistent high-risk human papilloma virus infection. J. Transl. Med. 2022, 20, 12. [Google Scholar] [CrossRef]
- Alimena, S.; Davis, J.; Fichorova, R.N.; Feldman, S. The vaginal microbiome: A complex milieu affecting risk of human papillomavirus persistence and cervical cancer. Curr. Probl. Cancer. 2022, 46, 100877. [Google Scholar] [CrossRef]
- Balkus, J.E.; Richardson, B.A.; Rabe, L.K.; Taha, T.E.; Mgodi, N.; Kasaro, M.P.; Ramjee, G.; Hoffman, I.F.; Abdool Karim, S.S. Bacterial vaginosis and the risk of Trichomonas vaginalis acquisition among HIV-1-negative women. Sex. Transm. Dis. 2014, 41, 123–128. [Google Scholar] [CrossRef]
- Chiu, S.F.; Huang, P.J.; Cheng, W.H.; Huang, C.Y.; Chu, L.J.; Lee, C.C.; Lin, H.C.; Chen, L.C.; Lin, W.N.; Tsao, C.H.; et al. Vaginal Microbiota of the Sexually Transmitted Infections Caused by Chlamydia trachomatis and Trichomonas vaginalis in Women with Vaginitis in Taiwan. Microorganisms 2021, 9, 1864. [Google Scholar] [CrossRef]
- Margarita, V.; Fiori, P.L.; Rappelli, P. Impact of Symbiosis Between Trichomonas vaginalis and Mycoplasma hominis on Vaginal Dysbiosis: A Mini Review. Front. Cell. Infect. Microbiol. 2020, 10, 179. [Google Scholar] [CrossRef]
- Jarrett, O.D.; Srinivasan, S.; Richardson, B.A.; Fiedler, T.; Wallis, J.M.; Kinuthia, J.; Jaoko, W.; Mandaliya, K.; Fredricks, D.N.; McClelland, R.S. Specific Vaginal Bacteria Are Associated with an Increased Risk of Trichomonas vaginalis Acquisition in Women. J. Infect. Dis. 2019, 220, 1503–1510. [Google Scholar] [CrossRef]
- Marrazzo, J.M.; David, M.; Heather, W.D.; Schulte, J.D.O.; Sobel, J.D.; Hillier, S.L.; Deal, C.; Fredricks, D.N. Bacterial vaginosis: Identifying research gaps proceedings of a workshop sponsored by DHHS/NIH/NIAID. Sex. Transm. Dis. 2010, 37, 732–744. [Google Scholar] [CrossRef]
- Abbe, C.; Mitchell, C.M. Bacterial vaginosis: A review of approaches to treatment and prevention. Front. Reprod. Health 2023, 5, 1100029. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, C.S.; Sobel, J.D. Current Treatment of Bacterial Vaginosis—Limitations and Need for Innovation. J. Infect. Dis. 2016, 214, S14–S20. [Google Scholar] [CrossRef] [PubMed]
- WHO. Guidelines for the Management of Symptomatic Sexually Transmitted Infections; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Machado, D.; Castro, J.; Palmeira-de-Oliveira, A.; Martinez-de-Oliveira, J.; Cerca, N. Bacterial Vaginosis Biofilms: Challenges to Current Therapies and Emerging Solutions. Front. Microbiol. 2016, 6, 1528. [Google Scholar] [CrossRef] [PubMed]
- Schwebke, J.R.; Desmond, R.A. Tinidazole vs metronidazole for the treatment of bacterial vaginosis. Am. J. Obstet. Gynecol. 2011, 204, 211.e1–211.e2116. [Google Scholar] [CrossRef]
- Donders, G.G.G.; Zodzika, J.; Rezeberga, D. Treatment of bacterial vaginosis: What we have and what we miss. Expert. Opin. Pharmacother. 2014, 15, 645–657. [Google Scholar] [CrossRef]
- Bilardi, J.; Walker, S.; McNair, R.; Mooney-Somers, J.; Temple-Smith, M.; Bellhouse, C.; Fairley, C.; Chen, M.; Bradshaw, C. Women’s Management of Recurrent Bacterial Vaginosis and Experiences of Clinical Care: A Qualitative Study. PLoS ONE 2016, 11, e0151794. [Google Scholar] [CrossRef]
- Bradshaw, C.S.; Brotman, R.M. Making inroads into improving treatment of bacterial vaginosis—Striving for long-term cure. BMC Infect. Dis. 2015, 15, 292. [Google Scholar] [CrossRef]
- Marrazzo, J.M.; Fiedler, T.L.; Srinivasan, S.; Thomas, K.K.; Liu, C.; Ko, D.; Xie, H.; Saracino, M.; Fredricks, D.N. Extravaginal reservoirs of vaginal bacteria as risk factors for incident bacterial vaginosis. J. Infect. Dis. 2012, 205, 1580–1588. [Google Scholar] [CrossRef]
- Muzny, C.A.; Sobel, J.D. The Role of Antimicrobial Resistance in Refractory and Recurrent Bacterial Vaginosis and Current Recommendations for Treatment. Antibiotics 2022, 11, 500. [Google Scholar] [CrossRef]
- DuPont, H.L. The potential for development of clinically relevant microbial resistance to rifaximin-α: A narrative review. Clin. Microbiol. Rev. 2023, 36, e0003923. [Google Scholar] [CrossRef]
- Shaskolskiy, B.; Dementieva, E.; Leinsoo, A.; Runina, A.; Vorobyev, D.; Plakhova, X.; Kubanov, A.; Deryabin, D.; Gryadunov, D. Drug Resistance Mechanisms in Bacteria Causing Sexually Transmitted Diseases and Associated with Vaginosis. Front. Microbiol. 2016, 7, 747. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.W.; Song, J.J.; Zeng, S.H.; Huang, Y.L.; Luo, J.J.; Guo, W.L.; Li, X.Y. Plasmid-mediated azithromycin resistance in non-typhoidal Salmonella recovered from human infections. J. Antimicrob. Chemother. 2024, 79, 2688–2697. [Google Scholar] [CrossRef] [PubMed]
- Javed, A.; Parvaiz, F.; Manzoor, S. Bacterial vaginosis: An insight into the prevalence, alternative treatments regimen and it’s associated resistance patterns. Microb. Pathog. 2019, 127, 21–30. [Google Scholar] [CrossRef]
- Nourbakhsh, F.; Nasrollahzadeh, M.S.; Tajani, A.S.; Soheili, V.; Hadizadeh, F. Bacterial biofilms and their resistance mechanisms: A brief look at treatment with natural agents. Folia Microbiol. 2022, 67, 535–554. [Google Scholar] [CrossRef]
- Tempera, G.; Furneri, P.M. Management of aerobic vaginitis. Gynecol. Obstet. Investig. 2010, 70, 244–249. [Google Scholar] [CrossRef]
- Sousa, L.G.V.; Pereira, S.A.; Cerca, N. Fighting polymicrobial biofilms in bacterial vaginosis. Microb. Biotechnol. 2023, 16, 1423–1437. [Google Scholar] [CrossRef]
- Donders, G.G.; Ruban, K.; Bellen, G. Selecting anti-microbial treatment of aerobic vaginitis. Curr. Infect. Dis. Rep. 2015, 17, 477. [Google Scholar] [CrossRef]
- FDA. Early Clinical Trials with Live Biotherapeutic Products: Chemistry, Manufacturing, and Control Information; Food Drug and Administration: College Park, MD, USA, 2016.
- Cordaillat-Simmons, M.; Rouanet, A.; Pot, B. Live biotherapeutic products: The importance of a defined regulatory framework. Exp. Mol. Med. 2020, 52, 1397–1406. [Google Scholar] [CrossRef]
- Amabebe, E.; Bhatnagar, N.; Kamble, N.; Reynolds, S.; Anumba, D.O. Exploring the antimicrobial properties of vaginal Lactobacillus crispatus against preterm birth-associated bacteria. Reprod. Fertil. 2022, 3, L6–L8. [Google Scholar] [CrossRef]
- Lagenaur, L.A.; Hemmerling, A.; Chiu, C.; Miller, S.; Lee, P.P.; Cohen, C.R.; Parks, T.P. Connecting the Dots: Translating the Vaginal Microbiome into a Drug. J. Infect. Dis. 2021, 223, S296–S306. [Google Scholar] [CrossRef]
- López-Moreno, A.; Aguilera, M. Vaginal Probiotics for Reproductive Health and Related Dysbiosis: Systematic Review and Meta-Analysis. J. Clin. Med. 2021, 10, 1461. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Hugerth, L.W.; Schuppe-Koistinen, I.; Du, J. The right bug in the right place: Opportunities for bacterial vaginosis treatment. NPJ Biofilms Microbiomes 2022, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Lehtoranta, L.; Ala-Jaakkola, R.; Laitila, A.; Maukonen, J. Healthy Vaginal Microbiota and Influence of Probiotics Across the Female Life Span. Front. Microbiol. 2022, 13, 819958. [Google Scholar] [CrossRef] [PubMed]
- Muzny, C.A.; Łaniewski, P.; Schwebke, J.R.; Herbst-Kralovetz, M.M. Host-vaginal microbiota interactions in the pathogenesis of bacterial vaginosis. Curr. Opin. Infect. Dis. 2020, 33, 59–65. [Google Scholar] [CrossRef]
- van der Veer, C.; Hertzberger, R.Y.; Bruisten, S.M.; Tytgat, H.L.P.; Swanenburg, J.; de Kat Angelino-Bart, A.; Schuren, F.; Molenaar, D.; Reid, G.; de Vries, H.; et al. Comparative genomics of human Lactobacillus crispatus isolates reveals genes for glycosylation and glycogen degradation: Implications for in vivo dominance of the vaginal microbiota. Microbiome 2019, 7, 49. [Google Scholar] [CrossRef]
- Tachedjian, G.; Aldunate, M.; Bradshaw, C.S.; Cone, R.A. The role of lactic acid production by probiotic Lactobacillus species in vaginal health. Res. Microbiol. 2017, 168, 782–792. [Google Scholar] [CrossRef]
- Wang, S.; Wang, Q.; Yang, E.; Yan, L.; Li, T.; Zhuang, H. Antimicrobial Compounds Produced by Vaginal Lactobacillus crispatus Are Able to Strongly Inhibit Candida albicans Growth, Hyphal Formation and Regulate Virulence-Related Gene Expressions. Front. Microbiol. 2017, 8, 564. [Google Scholar] [CrossRef]
- Turovskiy, Y.; Ludescher, R.D.; Aroutcheva, A.A.; Faro, S.; Chikindas, M.L. Lactocin 160, a Bacteriocin Produced by Vaginal Lactobacillus rhamnosus, Targets Cytoplasmic Membranes of the Vaginal Pathogen, Gardnerella vaginalis. Probiotics Antimicrob. Proteins 2009, 1, 67–74. [Google Scholar] [CrossRef]
- Maldonado-Barragán, A.; Caballero-Guerrero, B.; Martín, V.; Ruiz-Barba, J.L.; Rodríguez, J.M. Purification and genetic characterization of gassericin E, a novel co-culture inducible bacteriocin from Lactobacillus gasseri EV1461 isolated from the vagina of a healthy woman. BMC Microbiol. 2016, 16, 37. [Google Scholar] [CrossRef]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C. Host interactions of probiotic bacterial surface molecules: Comparison with commensals and pathogens. Nat. Rev. Microbiol. 2010, 8, 171–184. [Google Scholar] [CrossRef]
- Pradines, B.; Domenichini, S.; Lievin-Le Moal, V. Adherent Bacteria and Parasiticidal Secretion Products of Human Cervicovaginal Microbiota-Associated Lactobacillus gasseri Confer Non-Identical Cell Protection against Trichomonas vaginalis-Induced Cell Detachment. Pharmaceuticals 2022, 15, 1350. [Google Scholar] [CrossRef] [PubMed]
- Edelman, S.M.; Lehti, T.A.; Kainulainen, V.; Antikainen, J.; Kylväjä, R.; Baumann, M.; Westerlund-Wikström, B.; Korhonen, T.K. Identification of a high-molecular-mass Lactobacillus epithelium adhesin (LEA) of Lactobacillus crispatus ST1 that binds to stratified squamous epithelium. Microbiology 2012, 158, 1713–1722. [Google Scholar] [CrossRef]
- Ojala, T.; Kankainen, M.; Castro, J.; Cerca, N.; Edelman, S.; Westerlund-Wikström, B.; Paulin, L.; Holm, L.; Auvinen, P. Comparative genomics of Lactobacillus crispatus suggests novel mechanisms for the competitive exclusion of Gardnerella vaginalis. BMC Genom. 2014, 15, 1070. [Google Scholar] [CrossRef] [PubMed]
- Donnarumma, G.; Molinaro, A.; Cimini, D.; De Castro, C.; Valli, V.; De Gregorio, V.; De Rosa, M.; Schiraldi, C. Lactobacillus crispatus L1: High cell density cultivation and exopolysaccharide structure characterization to highlight potentially beneficial effects against vaginal pathogens. BMC Microbiol. 2014, 14, 137. [Google Scholar] [CrossRef]
- Song, J.; Lang, F.; Zhao, N.; Guo, Y.; Zhang, H. Vaginal Lactobacilli Induce Differentiation of Monocytic Precursors Toward Langerhans-like Cells: In Vitro Evidence. Front. Immunol. 2018, 9, 2437. [Google Scholar] [CrossRef]
- Hummelen, R.; Changalucha, J.; Butamanya, N.L.; Cook, A.; Habbema, J.D.; Reid, G. Lactobacillus rhamnosus GR-1 and L. reuteri RC-14 to prevent or cure bacterial vaginosis among women with HIV. Int. J. Gynaecol. Obstet. 2010, 111, 245–248. [Google Scholar] [CrossRef]
- Laue, C.; Papazova, E.; Liesegang, A.; Pannenbeckers, A.; Arendarski, P.; Linnerth, B.; Domig, K.J.; Kneifel, W.; Petricevic, L.; Schrezenmeir, J. Effect of a yoghurt drink containing Lactobacillus strains on bacterial vaginosis in women—A double-blind, randomised, controlled clinical pilot trial. Benef. Microbes 2018, 9, 35–50. [Google Scholar] [CrossRef]
- Heczko, P.B.; Tomusiak, A.; Adamski, P.; Jakimiuk, A.J.; Stefański, G.; Mikołajczyk-Cichońska, A.; Suda-Szczurek, M.; Strus, M. Supplementation of standard antibiotic therapy with oral probiotics for bacterial vaginosis and aerobic vaginitis: A randomised, double-blind, placebo-controlled trial. BMC Womens Health 2015, 15, 115. [Google Scholar] [CrossRef]
- Pendharkar, S.; Brandsborg, E.; Hammarström, L.; Marcotte, H.; Larsson, P.G. Vaginal colonisation by probiotic lactobacilli and clinical outcome in women conventionally treated for bacterial vaginosis and yeast infection. BMC Infect. Dis. 2015, 15, 255. [Google Scholar] [CrossRef]
- Davar, R.; Nokhostin, F.; Eftekhar, M.; Sekhavat, L.; Bashiri Zadeh, M.; Shamsi, F. Comparing the Recurrence of Vulvovaginal Candidiasis in Patients Undergoing Prophylactic Treatment with Probiotic and Placebo During the 6 Months. Probiotics Antimicrob. Proteins 2016, 8, 130–133. [Google Scholar] [CrossRef]
- Cohen, C.R.; Wierzbicki, M.R.; French, A.L.; Morris, S.; Newmann, S.; Reno, H.; Green, L.; Miller, S.; Powell, J.; Parks, T.; et al. Randomized Trial of Lactin-V to Prevent Recurrence of Bacterial Vaginosis. N. Engl. J. Med. 2020, 382, 1906–1915. [Google Scholar] [CrossRef] [PubMed]
- Reznichenko, H.; Henyk, N.; Maliuk, V.; Khyzhnyak, T.; Tynna, Y.; Filipiuk, I.; Veresniuk, N.; Zubrytska, L.; Quintens, J.; Richir, K.; et al. Oral Intake of Lactobacilli Can Be Helpful in Symptomatic Bacterial Vaginosis: A Randomized Clinical Study. J. Low. Genit. Tract Dis. 2020, 24, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lyu, J.; Ge, L.; Huang, L.; Peng, Z.; Liang, Y.; Zhang, X.; Fan, S. Probiotic Lacticaseibacillus rhamnosus GR-1 and Limosilactobacillus reuteri RC-14 as an Adjunctive Treatment for Bacterial Vaginosis Do Not Increase the Cure Rate in a Chinese Cohort: A Prospective, Parallel-Group, Randomized, Controlled Study. Front. Cell. Infect. Microbiol. 2021, 11, 669901. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Lee, E.S.; Park, S.T.; Jeong, S.Y.; Yun, Y.; Kim, Y.; Jeong, Y.; Kang, C.H.; Choi, H.J. Efficacy and Safety of MED-01 Probiotics on Vaginal Health: A 12-Week, Multicenter, Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Nutrients 2023, 15, 331. [Google Scholar] [CrossRef]
- Tomusiak, A.; Strus, M.; Heczko, P.B.; Adamski, P.; Stefański, G.; Mikołajczyk-Cichońska, A.; Suda-Szczurek, M. Efficacy and safety of a vaginal medicinal product containing three strains of probiotic bacteria: A multicenter, randomized, double-blind, and placebo-controlled trial. Drug Des. Devel. Ther. 2015, 9, 5345–5354. [Google Scholar] [CrossRef]
- Recine, N.; Palma, E.; Domenici, L.; Giorgini, M.; Imperiale, L.; Sassu, C.; Musella, A.; Marchetti, C.; Muzii, L.; Panici, P.B. Restoring vaginal microbiota: Biological control of bacterial vaginosis. A prospective case-control study using Lactobacillus rhamnosus BMX 54 as adjuvant treatment against bacterial vaginosis. Arch. Gynecol. Obstet. 2016, 293, 101–107. [Google Scholar] [CrossRef]
- Palma, E.; Recine, N.; Domenici, L.; Giorgini, M.; Pierangeli, A.; Panici, P.B. Long-term Lactobacillus rhamnosus BMX 54 application to restore a balanced vaginal ecosystem: A promising solution against HPV-infection. BMC Infect. Dis. 2018, 18, 13. [Google Scholar] [CrossRef]
- Marcotte, H.; Larsson, P.G.; Andersen, K.K.; Zuo, F.; Mikkelsen, L.S.; Brandsborg, E.; Gray, G.; Laher, F.; Otwombe, K. An exploratory pilot study evaluating the supplementation of standard antibiotic therapy with probiotic lactobacilli in south African women with bacterial vaginosis. BMC Infect. Dis. 2019, 19, 824. [Google Scholar] [CrossRef]
- Mändar, R.; Sõerunurk, G.; Štšepetova, J.; Smidt, I.; Rööp, T.; Kõljalg, S.; Saare, M.; Ausmees, K.; Le, D.D.; Jaagura, M.; et al. Impact of Lactobacillus crispatus-containing oral and vaginal probiotics on vaginal health: A randomised double-blind placebo controlled clinical trial. Benef. Microbes 2023, 14, 143–152. [Google Scholar] [CrossRef]
- Russo, R.; Superti, F.; Karadja, E.; De Seta, F. Randomised clinical trial in women with Recurrent Vulvovaginal Candidiasis: Efficacy of probiotics and lactoferrin as maintenance treatment. Mycoses 2019, 62, 328–335. [Google Scholar] [CrossRef]
- Lyra, A.; Ala-Jaakkola, R.; Yeung, N.; Datta, N.; Evans, K.; Hibberd, A.; Lehtinen, M.J.; Forssten, S.D.; Ibarra, A.; Pesonen, T.; et al. A Healthy Vaginal Microbiota Remains Stable during Oral Probiotic Supplementation: A Randomised Controlled Trial. Microorganisms 2023, 11, 499. [Google Scholar] [CrossRef] [PubMed]
- Vicariotto, F.; Malfa, P.; Viciani, E.; Dell’Atti, F.; Squarzanti, D.F.; Marcante, A.; Castagnetti, A.; Ponchia, R.; Governini, L.; De Leo, V. Efficacy of Lactiplantibacillus plantarum PBS067, Bifidobacterium animalis subsp. lactis BL050, and Lacticaseibacillus rhamnosus LRH020 in the Amelioration of Vaginal Microbiota in Post-Menopausal Women: A Prospective Observational Clinical Trial. Nutrients 2024, 16, 402. [Google Scholar]
- Baldacci, F.; Baldacci, M.; Bertini, M. Lactobacillus rhamnosus BMX 54 + Lactose, A Symbiotic Long-Lasting Vaginal Approach to Improve Women’s Health. Int. J. Womens Health 2020, 12, 1099–1104. [Google Scholar] [CrossRef]
- Vivekanandan, V.; Khan, Z.H.; Venugopal, G.; Musunuru, B.; Mishra, P.; Srivastava, S.; Ramadass, B.; Subhadra, B. VagiBIOM Lactobacillus suppository improves vaginal health index in perimenopausal women with bacterial vaginosis: A randomized control trial. Sci. Rep. 2024, 14, 3317. [Google Scholar] [CrossRef]
- Lev-Sagie, A.; Goldman-Wohl, D.; Cohen, Y.; Dori-Bachash, M.; Leshem, A.; Mor, U.; Strahilevitz, J.; Moses, A.E.; Shapiro, H.; Yagel, S.; et al. Vaginal microbiome transplantation in women with intractable bacterial vaginosis. Nat. Med. 2019, 25, 1500–1504. [Google Scholar] [CrossRef]
- Wrønding, T.; Vomstein, K.; Bosma, E.F.; Mortensen, B.; Westh, H.; Heintz, J.E.; Mollerup, S.; Petersen, A.M.; Ensign, L.M.; DeLong, K.; et al. Antibiotic-free vaginal microbiota transplant with donor engraftment, dysbiosis resolution and live birth after recurrent pregnancy loss: A proof of concept case study. EClinicalMedicine 2023, 61, 102070. [Google Scholar] [CrossRef]
- Oerlemans, E.F.M.; Bellen, G.; Claes, I.; Henkens, T.; Allonsius, C.N.; Wittouck, S.; van den Broek, M.F.L.; Wuyts, S.; Kiekens, F.; Donders, G.G.G.; et al. Impact of a lactobacilli-containing gel on vulvovaginal candidosis and the vaginal microbiome. Sci. Rep. 2020, 10, 7976. [Google Scholar] [CrossRef]
- Yoshikata, R.; Yamaguchi, M.; Mase, Y.; Tatsuyuki, A.; Myint, K.Z.Y.; Ohta, H. Evaluation of the efficacy of Lactobacillus-containing feminine hygiene products on vaginal microbiome and genitourinary symptoms in pre- and postmenopausal women: A pilot randomized controlled trial. PLoS ONE 2022, 17, e0270242. [Google Scholar] [CrossRef]
- Shen, X.; Xu, L.; Zhang, Z.; Yang, Y.; Li, P.; Ma, T.; Guo, S.; Kwok, L.Y.; Sun, Z. Postbiotic gel relieves clinical symptoms of bacterial vaginitis by regulating the vaginal microbiota. Front. Cell. Infect. Microbiol. 2023, 13, 1114364. [Google Scholar] [CrossRef]
- Al-Nasiry, S.; Ambrosino, E.; Schlaepfer, M.; Morré, S.A.; Wieten, L.; Voncken, J.W.; Spinelli, M.; Mueller, M.; Kramer, B.W. The Interplay Between Reproductive Tract Microbiota and Immunological System in Human Reproduction. Front. Immunol. 2020, 11, 378. [Google Scholar] [CrossRef]
- Borrego-Ruiz, A.; Borrego, J.J. Neurodevelopmental Disorders Associated with Gut Microbiome Dysbiosis in Children. Children 2024, 11, 796. [Google Scholar] [CrossRef] [PubMed]
- Borrego-Ruiz, A.; Borrego, J.J. Microbial Dysbiosis in the Skin Microbiome and Its Psychological Consequences. Microorganisms 2024, 12, 1908. [Google Scholar] [CrossRef]
- Barrientos-Durán, A.; Fuentes-López, A.; de Salazar, A.; Plaza-Díaz, J.; García, F. Reviewing the Composition of Vaginal Microbiota: Inclusion of Nutrition and Probiotic Factors in the Maintenance of Eubiosis. Nutrients 2020, 12, 419. [Google Scholar] [CrossRef]
- Morsli, M.; Gimenez, E.; Magnan, C.; Salipante, F.; Huberlant, S.; Letouzey, V.; Lavigne, J.P. The association between lifestyle factors and the composition of the vaginal microbiota: A review. Eur. J. Clin. Microbiol. Infect. Dis. 2024, 43, 1869–1881. [Google Scholar] [CrossRef]
- Allsworth, J.E.; Peipert, J.F. Severity of bacterial vaginosis and the risk of sexually transmitted infection. Am. J. Obstet. Gynecol. 2011, 205, 113.e1–113.e6. [Google Scholar] [CrossRef]
- Lamont, R.F.; Taylor-Robinson, D. The role of bacterial vaginosis, aerobic vaginitis, abnormal vaginal flora and the risk of preterm birth. BJOG 2010, 117, 119–120. [Google Scholar] [CrossRef]
- Srinivasan, S.; Hoffman, N.G.; Morgan, M.T.; Matsen, F.A.; Fiedler, T.L.; Hall, R.W.; Ross, F.J.; McCoy, C.O.; Bumgarner, R.; Marrazzo, J.M.; et al. Bacterial communities in women with bacterial vaginosis: High resolution phylogenetic analyses reveal relationships of microbiota to clinical criteria. PLoS ONE 2012, 7, e37818. [Google Scholar] [CrossRef]
- DeLong, K.; Bensouda, S.; Zulfiqar, F.; Zierden, H.C.; Hoang, T.M.; Abraham, A.G.; Coleman, J.S.; Cone, R.A.; Gravitt, P.E.; Hendrix, C.W.; et al. Conceptual Design of a Universal Donor Screening Approach for Vaginal Microbiota Transplant. Front. Cell. Infect. Microbiol. 2019, 9, 306. [Google Scholar] [CrossRef]
- Meng, Y.; Sun, J.; Zhang, G. Vaginal microbiota transplantation is a truly opulent and promising edge: Fully grasp its potential. Front. Cell. Infect. Microbiol. 2024, 14, 1280636. [Google Scholar] [CrossRef]
- Borrego-Ruiz, A.; Borrego, J.J. Fecal Microbiota Transplantation as a Tool for Therapeutic Modulation of Neurological and Mental Disorders. SciBase Neurol. 2024, 2, 1018. [Google Scholar] [CrossRef]
- Rouanet, A.; Bolca, S.; Bru, A.; Claes, I.; Cvejic, H.; Girgis, H.; Harper, A.; Lavergne, S.N.; Mathys, S.; Pane, M.; et al. Live Biotherapeutic Products, A Road Map for Safety Assessment. Front. Med. 2020, 7, 237. [Google Scholar] [CrossRef] [PubMed]
- Heavey, M.K.; Durmusoglu, D.; Crook, N.; Anselmo, A.C. Discovery and delivery strategies for engineered live biotherapeutic products. Trends Biotechnol. 2022, 40, 354–369. [Google Scholar] [CrossRef] [PubMed]
- Ducarmon, Q.R.; Kuijper, E.J.; Olle, B. Opportunities and Challenges in Development of Live Biotherapeutic Products to Fight Infections. J. Infect. Dis. 2021, 223, S283–S289. [Google Scholar] [CrossRef]
- Pot, B.; Vandenplas, Y. Factors that influence clinical efficacy of live biotherapeutic products. Eur. J. Med. Res. 2021, 26, 40. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.H.; Wong, S.; Yu, J.; Lee, Y.Y.; Terauchi, J.; Lai, H.C.; Luo, J.C.; Kao, C.Y.; Yu, S.L.; Liou, J.M.; et al. Development of live biotherapeutic products: A position statement of Asia-Pacific Microbiota Consortium. Gut 2025, 74, 706–713. [Google Scholar] [CrossRef]
- Dreher-Lesnick, S.M.; Stibitz, S.; Carlson, P.E., Jr. U.S. Regulatory Considerations for Development of Live Biotherapeutic Products as Drugs. Microbiol. Spectr. 2017, 5, 10.1128. [Google Scholar] [CrossRef]
- Nakra, N.A.; Madan, R.P.; Buckley, N.; Huber, A.M.; Freiermuth, J.L.; Espinoza, L.; Walsh, J.; Parikh, U.M.; Penrose, K.J.; Keller, M.J.; et al. Loss of innate host defense following unprotected vaginal sex. J. Infect. Dis. 2016, 213, 840–847. [Google Scholar] [CrossRef]
- Sabo, M.C.; Balkus, J.E.; Richardson, B.A.; Srinivasan, S.; Kimani, J.; Anzala, O.; Schwebke, J.; Feidler, T.L.; Fredricks, D.N.; McClelland, R.S. Association between vaginal washing and vaginal bacterial concentrations. PLoS ONE 2019, 14, e0210825. [Google Scholar] [CrossRef]
- Michala, L. The Adolescent and Genital Dissatisfaction. Clin. Obstet. Gynecol. 2020, 63, 528–535. [Google Scholar] [CrossRef]
- Moulton, L.J.; Jernigan, A.M. Management of Retained Genital Piercings: A Case Report and Review. Case Rep. Obstet. Gynecol. 2017, 2017, 2402145. [Google Scholar] [CrossRef]
- Borrego-Ruiz, A. A current overview on adolescent alcohol misuse and its potential negative impacts. Alcohol. Drug Addict./Alkohol. Narkomania 2025, 37, 205–231. [Google Scholar] [CrossRef]
- Aslan, E.; Bechelaghem, N. To ‘douche’ or not to ‘douche’: Hygiene habits may have detrimental effects on vaginal microbiota. J. Obstet. Gynaecol. 2018, 38, 678–681. [Google Scholar] [CrossRef]
- Łaniewski, P.; Owen, K.A.; Khnanisho, M.; Brotman, R.M.; Herbst-Kralovetz, M.M. Clinical and Personal Lubricants Impact the Growth of Vaginal Lactobacillus Species and Colonization of Vaginal Epithelial Cells: An in Vitro Study. Sex. Transm. Dis. 2021, 48, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, E.M.; Łaniewski, P.; Herbst-Kralovetz, M.M.; Brotman, R.M. Personal and Clinical Vaginal Lubricants: Impact on Local Vaginal Microenvironment and Implications for Epithelial Cell Host Response and Barrier Function. J. Infect. Dis. 2019, 220, 2009–2018. [Google Scholar] [CrossRef]
- Jovandaric, M.Z.; Jovanović, K.; Raus, M.; Babic, S.; Igic, T.; Kotlica, B.; Milicevic, S. The Significance of Plant Nutrition in the Creation of the Intestinal Microbiota—Prevention of Chronic Diseases: A Narrative Review. Medicina 2024, 60, 1969. [Google Scholar] [CrossRef]
- Yang, H.J.; Kim, D.S.; Lee, K.W.; Kim, Y.H. The Urinary Microbiome; Axis Crosstalk and Short-Chain Fatty Acid. Diagnostics 2022, 12, 3119. [Google Scholar] [CrossRef]
- Borrego-Ruiz, A.; González-Domenech, C.M.; Borrego, J.J. The Role of Fermented Vegetables as a Sustainable and Health-Promoting Nutritional Resource. Appl. Sci. 2024, 14, 10853. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Published by MDPI on behalf of the Hellenic Society for Microbiology. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borrego-Ruiz, A.; Borrego, J.J. Microbial Pathogens Linked to Vaginal Microbiome Dysbiosis and Therapeutic Tools for Their Treatment. Acta Microbiol. Hell. 2025, 70, 19. https://doi.org/10.3390/amh70020019
Borrego-Ruiz A, Borrego JJ. Microbial Pathogens Linked to Vaginal Microbiome Dysbiosis and Therapeutic Tools for Their Treatment. Acta Microbiologica Hellenica. 2025; 70(2):19. https://doi.org/10.3390/amh70020019
Chicago/Turabian StyleBorrego-Ruiz, Alejandro, and Juan J. Borrego. 2025. "Microbial Pathogens Linked to Vaginal Microbiome Dysbiosis and Therapeutic Tools for Their Treatment" Acta Microbiologica Hellenica 70, no. 2: 19. https://doi.org/10.3390/amh70020019
APA StyleBorrego-Ruiz, A., & Borrego, J. J. (2025). Microbial Pathogens Linked to Vaginal Microbiome Dysbiosis and Therapeutic Tools for Their Treatment. Acta Microbiologica Hellenica, 70(2), 19. https://doi.org/10.3390/amh70020019