Abstract
New ornithopod remains have recently been discovered at the Vadillos-1 paleontological site, in Cuenca Province, Central Spain. These remains were collected in sandy and clayey deposits within “Wealden” alluvial facies, which correspond to an upper Barremian (Lower Cretaceous) alluvial-palustrine muddy floodplain paleoenvironment. The remains correspond to a part of a sacrum, including three fused sacral vertebral centra with sacral ribs and part of the sacral yoke, together with an isolated unfused sacral vertebra. An incomplete ischium has also been identified, likely belonging to the same individual. The sacral block exhibits a broad sulcus that begins at the most anterior sacral vertebra, continues through the middle vertebra, and ends in the area where the middle and the most posterior vertebrae are fused. The non fused sacral vertebra also displays a ventral sulcus. The incomplete ischium has a curved shaft with a triangular cross section towards the distal end. The combination of the morphological features of the sacrum and ischium allow its attribution to Iguanodon cf. bernissartensis. This species is identified in the area for the first time, adding new valuable data about the dinosaur assemblages from the Lower Cretaceous of Spain.
1. Introduction
Iguanodontia remains, especially styracosternans [1], are well known from the Lower Cretaceous of Western Europe, mainly in the United Kingdom, France, Belgium, Portugal, and Spain (e.g., [2,3,4,5,6,7,8,9,10,11]). Iguanodon bernissartensis Boulenger in Van Beneden, 1881 [12], is the large styracosternan dinosaur best represented in the Lower Cretaceous (upper Barremian-lower Aptian) European “Wealden” alluvial facies, and its remains have been frequently found on the Iberian Peninsula. Most of the known material comes from the Maestrazgo Basin, Northeast Spain (e.g., [13,14,15]), and particularly from the Morella sub-basin, being less known, but also recorded in the Cuenca Province (e.g., [16]). Other upper Barremian ornithopods described in the Iberian Peninsula are Mantellisaurus atherfieldensis Hooley, 1925 [17], from the fossil site of Las Hoyas (Cuenca), in the La Huérguina Formation [18], and from Mas de La Parreta, Morella [13,14,15]; and Morelladon beltrani Gasulla et al., 2015 [14], from the upper Barremian of Morella, in the Morella Formation [19]. Verdú et al. [19] also studied the remains from the lower Barremian of Galve (Teruel), in the Camarillas Formation, and proposed the species Iguanodon galvensis, supporting the evolutionary relationship with I. bernissartensis [20,21]. Other Spanish Barremian ornithopods are Magnamanus soriaensis Fuentes Vidarte et al. [22] from the lower Barremian of Golmayo (Soria) and Portellsaurus sosbaynati Santos-Cubedo et al. [23] from the lower Barremian of Portell (Castellón).
In the Barremian of Hoces de Beteta (Cuenca Province, Central Spain), indeterminate ornithopod remains were previously reported [24,25], but without detailed description or identification. The presence of styracosternan ornithopods and other reptile fossils, such as crocodiles and ankylosaurs, is not new for the Vadillos-1 paleontological site, as larger-sized fossils had previously been documented [26]. In this paper, several newly discovered ornithopod remains from the upper Barremian vertebrate site of Vadillos-1 (Hoces de Beteta) are examined, and an initial approach to their taxonomic assignment is presented. The studied specimen comprises a partial skeleton consisting of part of the sacrum and the ischium. Despite its fragmentary nature, this specimen can be referred to I. cf. bernissartensis based on a unique combination of shared characters. The present work provides a description and a taxonomic assignment of the sacral remains and a fragmentary ischium of an ornithopod, as well as assess the significance of these new findings. This discovery is particularly noteworthy because it originates from a region (Cuenca province) where this type of dinosaur has been scarcely studied. Therefore, it fills gaps in our knowledge on dinosaur diversity in the Iberian Peninsula and of the degree of extension of Iguanodon-like ornithopods in Western Europe.
2. Geological and Geographical Setting
The upper Barremian vertebrate site of Vadillos-1 (European Datum 1950: Lat. 40° 320 20.2700, Long. 2° 80 31.2100; UTM 30 X 572663, Y 4488011 N) is located in the Beteta Municipality, at the northern part of Cuenca Province, Central Spain. It is situated in the area known as the Beteta Gorges, close to the village of Puente de Vadillos that gives its name to the site (Figure 1A). The Lower Cretaceous in the region is represented mainly by continental strata without direct marine influence [27]. In the area of Vadillos-1, the upper Barremian stratigraphic succession is widely exposed as an anticline fold, consisting mainly of mudstones rich in vertebrate remains [26] and [28]. The site corresponds to materials in “Weald” facies historically assigned to the El Collado Sandstone and Clay Formation. However, the materials of this lithostratigraphic unit have been recently assigned in the region to the Tragacete Formation [29].
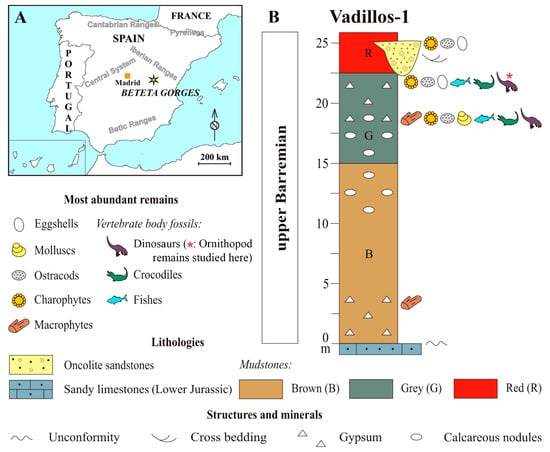
Figure 1.
General geographic location of the Beteta Gorges, Spain (A), and simplified biostratigraphic section of the upper Barremian of Vadillos-1 (B) with the main lithological characteristics and the most abundant fossil taxonomic groups, including the ornithopod remains studied herein. Modified from [26,28,30].
Vadillos-1 is specifically characterized by a succession, from base to top, of 15 m of brownish mudstones (B level), 8 m of greyish mudstones (G level) and >4 m of reddish clays (R level) separated by gradual boundaries, with carbonate nodules (in B and G levels) and interbedded sandstones (in R level) (Figure 1B). The depositional environment of Vadillos-1 is interpreted as an alluvial-palustrine muddy floodplain, showing intervals of more carbonate sedimentation, being temporary crossed by braided distributary channels with sandy infill [26,28,30].
So far, the fossil-rich layers, especially the G level of Vadillos-1, have yielded a substantial variety of micro and macro remains of plants, invertebrates and vertebrates. Plants have been categorized as unidentified macrophytes, spores of cryptogams, pollen grains from gymnosperms and angiosperms, as well as fructifications of charophytes (e.g., Atopochara, Globator, Clavator). Invertebrates are represented by ostracods (e.g., Cypridea) and molluscs (Unionoida, Viviparus sp.); and vertebrates include fishes, amphibians (Albanerpetontidae), turtles (cf. Eucryptodira), crocodylomorphs (Neosuchia), dinosaurs (ankylosaurs, ornithopods, theropods) and mammals [26,28,30,31,32]. In detail, Vadillos-1 shows an abundant record of small dental remains (Crocodylomorphs, [33]: Bernissartiidae, Atoposauridae, Goniopholididae, Unasuchus; and Theropods [30]: Dromaeosauridae, possibly non-Tyrannosaurid Tyrannosauroidea), suggesting, together with the existence of eggshell remains [28], the possible existence of a nearby nesting zone.
3. Materials and Methods
Vertebrate macrofossils appear in Vadillos-1 mostly as scattered and disarticulated remains, exposed on the surface of the sites. They mainly correspond to appendicular, vertebral and costal elements assignable to Dinosauria, usually fragmented and preserved in bioapatite, sometimes partially covered by a resistant carbonate matrix and with their interior partially replaced by calcite crystals with geode growth [26,34].
Among the remains selected for this work, in those that were adequately consolidated and showed a relatively clean surface without matrix, it was only necessary to apply a cleaning treatment by washing with demineralized water and a small brush to remove the scarce remains of adhering sediment. Differently, in those remains that were surrounded by a carbonate concretion of a marly nature that prevented an adequate observation of their morphology, it was essential to remove the matrix to achieve an adequate taxonomic and anatomical identification. To achieve this, cycles of chemical and mechanical cleaning have been conducted, utilizing treatments with hydrogen chloride (HCl) diluted from 15% to 5%, overtime intervals ranging from 1 to 72 h [35], followed by matrix elimination tests with a hand engraver or vibroincisor [36], with a standard tip of 38 mm and a wider one (chisel), connected to an air compressor. Following the completion of chemical and mechanical cleaning, the fragmentary remains were assembled using a reversible concentrated adhesive (Fluoline A). All the bone remains were labelled following the standard methodology [37]. The photographs of the specimens were taken with an Olympus E-M5II camera and processed using Adobe Photoshop CS6 software. Following this study, the remains have been temporarily deposited in suitable conservation conditions at the Research Laboratory of the “PaleoIbérica” Group located in the Department of Geology, Geography and Environment of the University of Alcalá (UAH) and remain accessible to the scientific community. Following this temporary phase, the remains will be transferred to the Museum of Paleontology of Castilla-La Mancha (MUPA).
4. Systematic Paleontology
Dinosauria Owen, 1842 [38]
Ornithischia Seeley, 1887 [39]
Ornithopoda Marsh, 1881 [40]
Iguanodontia Sereno, 1986 [1]
Ankylopollexia Sereno, 1986 [1]
Styracosterna Sereno, 1986 [1]
Iguanodon Mantell, 1825 [41]
Iguanodon cf. bernissartensis Boulenger in Van Beneden, 1881 [12]
Material: three co-fused sacral centra (VD1-511), including part of a neural arch in one of them, some sacral ribs and part of the sacral yoke (VD1-507), and a sacral (possibly associated), but non fused, vertebra VD1-403.
Locality: upper Barremian (Lower Cretaceous) of Beteta Gorges (Cuenca, Spain).
The sacrum:
Sacral block: Part of the sacral material (Figure 2) comprises three firmly fused centra recovered in two blocks (VD1-511), including part of a neural arch in one of them, the transverse processes, some sacral ribs and part of the sacral yoke (VD1-507). This sacral block might be looking at the posterior end of the sacrum. The depression in the dorsal half of the posterior articular surface of the most posterior vertebra (Figure 2A,B) could be to receive a dorsal bulge in the anterior articular surface of the first caudal vertebra or an unfused caudosacral (see Section 5 below). The most posterior sacral vertebra has the highest dorsoventral height although the shortest anteroposterior length (Table 1). The ventral surfaces of the posterior and middle vertebrae are more subquadrangular compared to the anterior vertebra which is more spool shaped (Figure 2). The sacral ribs are well-developed to form a large anteroventrally inclined platform in lateral view. The sacral ribs are fused to form the sacral yoke (Figure 2B–D).

Figure 2.
Upper Barremian ornithopod remains from the Vadillos-1 fossil site, in the Beteta Gorges (Spain). Partial sacral block VD1-511, in caudal (A), dorsal (B), ventral (C), right lateral (D), and left lateral (E) views. Abbreviations: de: depression; nc: neural canal; ri: ventral ridge; sr: sacral rib; sy: sacral yoke; su: ventral sulcus; tp: transverse process. Silhouette of I. bernissartensis modified from [42].

Table 1.
Dimension of sacral vertebrae from Iguanodon cf. bernissartensis (sacral block VD1-511 and isolated sacral VD1-403) from the upper Barremian Paleontological site of Vadillos-1 (Beteta Gorges, Spain). Note: The transverse width of the articular facets of the middle vertebra of the sacral block VD1-511 have been estimated because the vertebra is firmly fused and the transverse processes emerge from the intervertebral position between the most anterior and the middle vertebra.
The most posterior vertebra shows part of the neural arch, being incomplete, located to the dorsal surface of the centrum and the sacral ribs are fused to the lateral wall of the neural arch (Figure 2A). Part of the prezygapophyses and postzygapophyses are eroded and incomplete. The transverse processes/sacral ribs are close to the centrum of this vertebra. The following vertebra, situated in the middle part of this sacral block, is wider than it is high (Table 1). The articular surfaces of this vertebra are difficult to measure due to the fusion of the vertebrae and the presence of transverse processes from the intervertebral position. The most anterior vertebra is incomplete; the anterior articular surface being eroded. In the most anterior vertebra of this block, part of the transverse processes are preserved, originating from the anterior articular area and being directed posteriorly (Figure 2C).
In ventral view, this sacral block shows a broad sagittal sulcus in the more anterior and the middle vertebrae. In the ventral view of the most anterior vertebra, the sulcus is delimited by a ridge (Figure 2C). The sulcus can also be distinguished in the most posterior vertebra, being more evident in the area in which the middle and the most posterior vertebrae are fused, in the articular area, disappearing posteriorly (Figure 2C).
Sacral non-fused-vertebra: the vertebra VD1-403 (Figure 3A–F) is represented by a centrum and the base of the neural arch (neural pedicles). It is transversely distorted. The neural canal can be distinguished (Figure 3A–C), as well as facets (eroded) for the transverse processes-sacral ribs (Figure 3D). It has a centrum with a subelliptic, slightly taller than wide, essentially flat, but slightly concave anterior articular facet (Figure 3A), and a subhexagonal, wider than tall, slightly eroded posterior articular facet (Figure 3B) (Table 1). In lateral view, large foramina for lateral nerves are present (Figure 3D,E). The ventral surface is concave in lateral view, and it shows a longitudinal ventral sulcus with a central ridge or fold, being a non-contiguous sulcus (Figure 3F). This vertebra corresponds to a sacral vertebra non fused with the sacral block because of the presence of a ventral not continuous groove and because of the presence of facets for the sacral ribs (transverse processes/sacral ribs).

Figure 3.
Upper Barremian ornithopod from the Vadillos-1 fossil site, in the Beteta Gorges (Spain). Isolated sacral vertebra VD1-403, in anterior (A), posterior (B), dorsal (C) right lateral (D), left lateral (E) and ventral (F) views. fa: facet for transverse process/sacral rib; fo: foramen; nc: neural canal; su: ventral sulcus; ri: rib. Silhouette of I. bernissartensis modified from [42].
The ischium:
Ischium: The ischium VD1-0194 is incomplete (Figure 4), with part of the shaft and the distal end preserved, although the ending of the distal end is eroded. The proximal part is absent. This bone consists of a long, slightly curved shaft, expanded towards the distal end, becoming a subtriangular shape (in cross section) with angular edges. The shaft shows a lenticular cross section in the more proximal part of the preserved bone, and a subtriangular cross section towards the distal end (Figure 4). In lateral view, the lateral edges of the shaft are twisted anteriorly and posteriorly towards the middle of the preserved bone, giving rise to this subtriangular shape. The cross section of the most distal part of the (incomplete) distal end shows asymmetric triangular section with the posterolateral part more developed and extended.
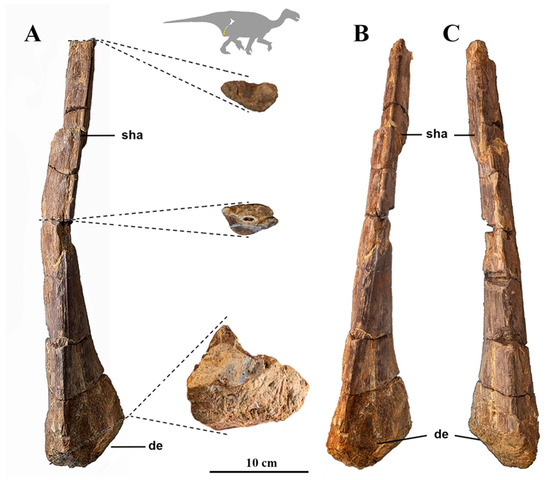
Figure 4.
Part of ischium VD1-0194 in lateral (A), with detail of the cross section, anterior (B) and posterior (C) views. Abbreviations: de: distal end; sha: shaft. Silhouette of I. bernissartensis modified from [42].
5. Discussion
The presence of a sacrum with fused vertebrae is a morphological feature that relates VD1-511 with a mature specimen [43,44,45]. Based on the vertebral size and the comparison of the vertebrae with those of the type specimen of Iguanodon bernissartensis Boulenger, 1881 (specimen IRSNB R51, [2]), an individual approximately 11 m in length [2,44], a similar length is estimated for the here described individual from the Vadillos-1 vertebrate site.
As mentioned above, the sacral block VD1-511 here studied seems to correspond to the most posterior sacral vertebrae of the sacral series and show a broad sulcus on the ventral surface. The depression in the dorsal half of the posterior articular surface of the posterior vertebra (Figure 2A,B) could be to receive a dorsal bulge in the anterior articular surface of the first caudal vertebra of perhaps an unfused caudosacral. This bulge is seen in other European styracosternans such us Brighstoneus simmondsi [11], Comptonatus chasei [46] and the Belgian specimen Dollodon bampingi [3]. Norman [2] also mentions it in Iguanodon bernissartensis so it seems to be a feature of most ‘Wealden’ iguanodontians. Considering that VD1-511 corresponds to the most posterior vertebrae, they could possibly correspond to S6, S7 and S8 (considering eight sacral vertebrae as typically occur in I. bernissartensis). In the case of the isolated sacral vertebra VD1-403, it could be the third counting from the anterior part, because of the presence of a non-continuous sulcus and a ridge similar to those described by Gasulla et al. [15] for I. bernissartensis in a vertebra from this position.
The presence of a midline broad ventral sulcus in the sacral centra VD1-511 (posterior sacral centra) leads to its comparison with the late Barremian-earliest Aptian I. bernissartensis. In the specimens of I. bernissartensis studied by Norman [2], the sacrals become progressively broader and more massive, and the ventral surface develops a shallow haemal sulcus [2] (Figure 45A,B). The sacrum of I. bernissartensis from the upper Barremian of Morella studied by Gasulla et al. [15] (Figure 3) shows a narrow midline ventral keel in sacral 2, while the more posterior sacral centra (3 to 6) has a broad midline ventral sulcus, like VD1-511. The ventral sulcus present in VD1-511 distinguishes it from other Barremian styracosternans in which this sulcus is absent, such as Morelladon beltrani [14], which have a ventral keel instead (see also [47] (Figure 7k–n)). In the sacral centra of M. atherfieldensis described by Norman [3], “the centra become at first slender and keeled, but subsequently (4–6) become again broader and develop a flattened or slightly depressed haemal surface”. However, the description of Hooley [17] of the holotype differs, showing a wide sulcus on a S3 and S4, ending at S5, and there is a slight keel on S6. According to Bonsor et al. [48], the absence of the sulcus on the holotype is difficult to qualify due to obstruction by the mount. Apart from the presence/absence of a groove or sulcus on the ventral surface of the sacral vertebrae, the skeletal proportions of the genus Mantellisaurus and Iguanodon are different, and dimensions of 8+ m are considered diagnostic of Iguanodon, while Mantellisaurus is considered a modest sized adult [49].
Comparing with the sacrum of other Barremian ornithopods, Brighstoneus simmondsi from the United Kingdom (England) apparently shows a ventral keel on the first sacral vertebra, but the rest of the sacral vertebrae are flat [44]. In the posterior sacral vertebrae of some specimens of Iguanodon galvensis Verdú et al., 2015 [20], from the lower Barremian of Spain, a deep ventral groove can also be observed (specimens MAP-8048 and MAP-8116 studied by García Cobeña et al. [47]), but this species is restricted to the lower Barremian. Verdú et al. [20,21], support an evolutionary relationship between the lower Barremian I. galvensis and the upper Barremian-lowermost Aptian I. bernissartensis.
The ischium VD1-0194 is a curved bone with a triangular cross section towards the distal end. Norman [46] regards the curvature of the ischiatic shaft as a potential diagnostic character of I. bernissartensis in combination with other features. The ‘curvature of the shaft’ as a qualitative character (curved vs. straight) is usually employed in iguanodontian systematics and phylogenetics (e.g., [14]). This character state could be difficult to assess in some European styracosternans with incomplete ischia (e.g., Brighstoneus [14], Barilium [4]). The ischiatic shaft seems to be straight in Mantellisaurus [8], Morelladon [19] or Comptonatus chasei [43]. The shaft is too incomplete in the Mantellisaurus holotype NHMUK PV R 5764 to know what shape it was. Among Early Cretaceous ornithopods, a curved shaft is well-known from I. bernissartensis and I. galvensis (see [50] (Figure 16)) According to Verdú et al. [20], I. bernissartensis and I. galvensis can be distinguished through the relative size of the distal end (small and large, respectively) of their ischia, although Verdú et al. [41] observe variability in the curvature of the ischium in adult specimens of I. bernissartensis. The ischium of I. galvensis (Verdú et al. [21]) has a rod with a D-shaped section close to the distal end, while I. bernissartensis is more triangular, like the specimen here studied VD1-0194. The size of the shaft and the extension of the distal end, with angular edges and triangular cross section close to the distal end, point to affinities with the shaft of I. bernissartensis. The features of the sacrum and ischium of other Barremian Spanish ornithopods such us the lower Barremian Magnamanus [22] and Portellsaurus [23] are still unknown.
The dimensions of the material, the groove on the ventral surface of sacral vertebrae (most posterior sacral vertebrae), the curvature of the ischium, and its triangular cross section, in combination with its stratigraphical position, point to affinities with the upper Barremian I. bernissartensis, the large styracosternan dinosaur best represented in Lower Cretaceous in the “Wealden” alluvial facies of Western Europe, already identified in other comparable vertebrate sites from Northeastern Spain. However, according to [11], the sitting of ventral keels and sulci shows intraspecific variation in some taxa, and caution is required interpreting their phylogenetic significance. This, along with the fact that the sacrum and ischium are too incomplete, makes it safer to classify the remains studied here as belonging to I. cf. bernissartensis.
6. Conclusions
New ornithopod remains from the upper Barremian (Lower Cretaceous) of Vadillos-1 fossil site, located in the Beteta Gorges (Cuenca, Spain), are described in this paper. These remains correspond to an incomplete sacrum composed of three sacral vertebral centra, fused with sacral ribs and part of the sacral yoke, a sacral vertebra not attached to the sacral block, and an incomplete ischium. The sacrum remains seem to correspond to the latest sacral vertebrae of the sacral series and shows a broad sulcus on its ventral surface. The groove on the ventral surface of sacral vertebrae, the curvature of the ischium, and its triangular cross section, in combination with its stratigraphical position, points to affinities with I. bernissartensis, being prudently classified as I. cf. bernissartensis. The ornithopod fossils identified in Vadillos-1 and described here confirm the similarities pointed out in previous papers between the vertebrate assemblage of this site and those of other Lower Cretaceous dinosaur sites from Northeastern Spain. These discoveries are key to completing the study of the dinosaur fauna in the Iberian Peninsula and Europe at a time characterized by the consolidation of new ecosystems in relation to major paleogeographical changes and land distribution areas.
Author Contributions
Conceptualization, M.B.-C. and F.B.-B.; methodology, M.B.-C., F.B.-B. and I.P.; software, M.B.-C. and F.B.-B.; validation, M.B.-C., F.B.-B., P.M.C. and R.P.; formal analysis, M.B.-C. and F.B.-B.; investigation, M.B.-C., F.B.-B., P.M.C. and R.P.; resources, M.B.-C. and F.B.-B., data curation, M.B.-C., F.B.-B., M.R.A.-F. and I.P.; writing—original draft preparation, M.B.-C. and F.B.-B.; writing—review and editing, M.B.-C., F.B.-B., M.R.A.-F., R.P., P.M.C. and I.P.; visualization, M.B.-C. and F.B.-B.; supervision, M.B.-C. and F.B.-B.; P.M.C. and R.P.; project administration, F.B.-B.; funding acquisition, M.B.-C.; F.B.-B. and P.M.C. All authors have read and agreed to the published version of the manuscript.
Funding
Postdoctoral Contract Margarita Salas UCM CT31/21 of the Complutense University of Madrid (Spain). Research Projects SBPLY/23/180801/000053, SBPLY/24/180801/000018 and “Ecosystems of the Cretaceous of Castilla-La Mancha” (SBPLY/21/180501/000242, co-financed by FEDER-ERDF, Operational Program CLM 2021–2027) of the Junta de Castilla-La Mancha and the University of Alcalá (Spain). Fundação para a Ciência e a Tecnologia through projects: UID/Multi/00611/2020 and UI/BD/150971/2021, CITEUC, Centre for Earth and Space Research of the University of Coimbra (Portugal).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The fossil remains here described are temporarily deposited in suitable conservation conditions at the Research Laboratory of the “PaleoIbérica” Research Group located in the Department of Geology, Geography and Environment of the University of Alcalá (UAH) and remain accessible to the scientific community. Following this temporary phase, the remains will be transferred to the Museum of Paleontology of Castilla-La Mancha (MUPA), Cuenca, Spain. Contact to the corresponding author for more information.
Acknowledgments
The authors would like to acknowledge Francisco Javier Verdú of the Fundación Conjunto Paleontológico de Teruel-Dinópolis (Teruel, Spain), Alfonso Calle from Carrascosa (Cuenca, Spain), and the members of the “PaleoIbérica” Research Group (University of Alcalá), especially Julia Audije-Gil and Senay Ozkaya de Juanas. The authors express their gratitude to all the palaeontologists for their participation in the field exploration and excavation campaigns at the Vadillos-1 fossil site. They are also grateful to the anonymous reviewers. These campaigns were carried out with authorization issued by the Dirección General de Patrimonio y Museos of the Junta de Castilla-La Mancha and were developed in collaboration with the municipal council of Cañizares.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sereno, P.C. Phylogeny of the bird-hipped dinosaurs (order Ornithischia). Natl. Geogr. Res. 1986, 2, 234–256. [Google Scholar]
- Norman, D.B. On the Ornithischian Dinosaur Iguanodon bernissartensis from the Lower Cretaceous of Bernissart (Belgium); Institut Royal des Sciences Naturelles de Belgique: Brussels, Belgium, 1980; Volume 178, pp. 1–103. [Google Scholar]
- Norman, D.B. On the Anatomy of Iguanodon atherfieldensis (Ornithischia: Ornithopoda); Institut Royal des Sciences Naturelles de Belgique: Brussels, Belgium, 1986; Volume 56, pp. 281–372. [Google Scholar]
- Norman, D.B. On the osteology of the lower Wealden Group (Valanginian) ornithopod Barilium dawsoni (Iguanodontia: Styracosterna). Spec. Pap. Palaeontol. 2011, 86, 165–194. [Google Scholar]
- Norman, D.B. Ornithopod dinosaurs. In English Wealden Fossils; Batten, D.J., Ed.; The Palaeontological Association: London, UK, 2011; pp. 405–475. [Google Scholar]
- Norman, D.B. On the taxonomy and diversity of Wealden iguanodontian dinosaurs (Ornithischia: Ornithopoda). Rev. Paleobiol. 2013, 32, 385–404. [Google Scholar]
- Norman, D.B. On the history, osteology, and systematic position of the Wealden (Hastings group) dinosaur Hypselospinus fittoni (Iguanodontia: Styracosterna). Zool. J. Linn. Soc. 2015, 173, 92–189. [Google Scholar] [CrossRef]
- Martin, V.; Buffetaut, E. Iguanodon (Ornithischia-Ornithopoda) du Crétacé Inférieur de la région de Saint-Dizier, Haute-Marne. Rev. Paleobiol. 1992, 11, 67–96. [Google Scholar]
- Knoll, F. A large iguanodont from the upper Barremian of the Paris Basin. Geobios 2009, 42, 755–764. [Google Scholar] [CrossRef]
- Figueiredo, S.; Rosina, P.; Figuti, L. Dinosaurs and other vertebrates from the Papo-Seco Formation (Lower Cretaceous) of southern Portugal. J. Iber. Geol. 2015, 41, 301–314. [Google Scholar] [CrossRef]
- Lockwood, J.A.F.; Martill, D.M.; Maidment, S.C.R. A new hadrosauriform dinosaur from the Wessex Formation, Wealden Group (Early Cretaceous), of the Isle of Wight, southern England. J. Syst. Palaeontol. 2021, 19, 847–888. [Google Scholar] [CrossRef]
- Boulenger, G.A. Rapport de P.J. Van Beneden “Sur l’arc pelvien chez les dinosauriens de Bernissart par M. G. A. Boulenger”. Bull. Acad. R. Sci. Lett. Beaux-Arts Belg. 1881, 1, 600–608. [Google Scholar]
- Gasulla, J.M.; Escaso, F.; Ortega, F.; Sanz, J.L. New hadrosauriform cranial remains from the Arcillas de Morella Formation (Lower Aptian) of Morella, Spain. Cretac. Res. 2014, 47, 19–24. [Google Scholar] [CrossRef]
- Gasulla, J.M. Los Dinosaurios de la Cantera del Mas de la Parreta, Morella (Formación Morella, Barremiense Superior, Cretácico Inferior): Sistemática, Análisis Filogenético e Implicaciones Paleobiológicas. Ph.D. Thesis, Universidad Autónoma de Madrid, Madrid, Spain, 2015. [Google Scholar]
- Gasulla, J.M.; Escaso, F.; Narváez, I.; Sanz, J.L.; Ortega, F. New Iguanodon bernissartensis axial bones (Dinosauria, Ornithopoda) from the Early Cretaceous of Morella, Spain. Diversity 2022, 14, 63. [Google Scholar] [CrossRef]
- Sanguino, F.; Buscalioni, A.D. The Iguanodon locality of Pata la Mona (upper Barremian, Buenache de la Sierra, Cuenca) revisited. In Proceedings of the XVI Encuentro de Jóvenes Investigadores en Paleontología, Zarautz, Spain, 11–14 April 2018; pp. 103–106. [Google Scholar]
- Hooley, R.W. On the skeleton of Iguanodon atherfieldensis sp. nov. from the Wealden Shales of Atherfield (Isle of Wight). Quart. J. Geol. Soc. Lond. 1925, 81, 1–61. [Google Scholar] [CrossRef]
- Llandres Serrano, M.L.; Vullo, R.; Marugán-Lobón, J.; Ortega, F.; Buscalioni, Á.D. An articulated hindlimb of a basal iguanodont (Dinosauria, Ornithopoda) from the Early Cretaceous Las Hoyas Lagerstätte (Spain). Geol. Mag. 2013, 150, 572–576. [Google Scholar] [CrossRef]
- Gasulla, J.M.; Escaso, F.; Narváez, I.; Ortega, F.; Sanz, J.L. A new sail-backed styracosternan (Dinosauria: Ornithopoda) from the Early Cretaceous of Morella, Spain. PLoS ONE 2015, 10, e0144167. [Google Scholar] [CrossRef]
- Verdú, F.J.; Royo-Torres, R.; Cobos, A.; Alcalá, L. Perinates of a new species of Iguanodon (Ornithischia: Ornithopoda) from the lower Barremian of Galve (Teruel, Spain). Cretac. Res. 2015, 56, 250–264. [Google Scholar] [CrossRef]
- Verdú, F.J.; Royo-Torres, R.; Cobos, A.; Alcalá, L. New systematic and phylogenetic data about the Early Barremian Iguanodon galvensis (Ornithopoda: Iguanodontoidea) from Spain. Hist. Biol. 2018, 30, 437–474. [Google Scholar] [CrossRef]
- Fuentes-Vidarte, C.; Meijide-Calvo, M.; Meijide-Fuentes, F.; Meijide-Fuentes, M. Un nuevo dinosaurio estiracosterno (Ornithopoda: Ankylopollexia) del Cretácico Inferior de España. Span. J. Palaeontol. 2016, 31, 407–446. [Google Scholar] [CrossRef]
- Santos-Cubedo, A.; de Santisteban, C.; Poza, B.; Meseguer, S. A new styracosternan hadrosauroid (Dinosauria: Ornithischia) from the Early Cretaceous of Portell, Spain. PLoS ONE 2021, 16, e0253599. [Google Scholar] [CrossRef]
- De Lapparent, A.F.; Curnelle, R.; Defaut, B.; Miroschedji, A.d.; Pallard, B. Nouveaux gisements de Dinosaures en Espagne Centrale. Estud. Geol. 1969, 25, 311–315. [Google Scholar]
- Ruiz-Omeñaca, J.I.; Canudo, J.I. Dinosaurs (Saurischia, Ornithischia) in the Barremian (Lower Cretaceous) of the Iberian Peninsula. In Dinosaurios y Otros Reptiles Mesozoicos en España; Pérez-Lorente, F., Ed.; Instituto de Estudios Riojanos: Logroño, Spain, 2003; pp. 269–312. [Google Scholar]
- Barroso-Barcenilla, F.; Berrocal-Casero, M.; Blain, H.A.; Callapez, P.M.; Cambra-Moo, O.; Escaso, F.; Martín-Closas, C.; Ortega, F.; Pérez-García, A.; Prieto, I.; et al. Geological and palaeontological context of three new Barremian vertebrate sites in the Iberian Peninsula (Lower Cretaceous, Cuenca, Spain). Proc. Geol. Assoc. 2017, 128, 256–270. [Google Scholar] [CrossRef]
- Poyato-Ariza, F.J.; Talbot, M.R.; Fregenal-Martínez, M.A.; Meléndez, N.; Wenz, S. First isotopic and multidisciplinary evidence for nonmarine coelacanths and pycnodontiform fishes: Palaeoenvironmental implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1998, 144, 65–84. [Google Scholar] [CrossRef]
- Bravo, A.M.; Sevilla, P.; Barroso-Barcenilla, F. Avian and crocodilian eggshells from the upper Barremian site of Vadillos-1 (Lower Cretaceous, Cuenca Province, Spain). Cretac. Res. 2018, 85, 28–41. [Google Scholar] [CrossRef]
- Fregenal-Martínez, M.; Meléndez, N.; Muñoz-García, M.B.; Elez, J.; Horra, R.d.l. The stratigraphic record of the Late Jurassic–Early Cretaceous rifting in the Alto Tajo–Serranía de Cuenca Region (Iberian Ranges, Spain): Genetic and structural evidences for a revision and a new lithostratigraphic proposal. Rev. Soc. Geol. Esp. 2017, 30, 113–142. [Google Scholar]
- Berrocal-Casero, M.; Alcalde-Fuentes, M.R.; Audije-Gil, J.; Sevilla, P. Theropod teeth from the upper Barremian (Lower Cretaceous) of Vadillos-1 (Spain). Cretac. Res. 2023, 142, 105392. [Google Scholar] [CrossRef]
- Díez-Somolinos, M.; Barroso-Barcenilla, F.; Berrocal-Casero, M.; Rodríguez-Lázaro, J.; Sevilla García, P.; Sames, B. New species of ostracods from the non-marine upper Barremian (Lower Cretaceous) of Vadillos-1 (Cuenca, Spain). Cretac. Res. 2025. under review. [Google Scholar]
- Sevilla, P.; Audije-Gil, J.; Berrocal-Casero, M.; Prieto, I.; Barroso-Barcenilla, F. Primeros hallazgos de restos de mamíferos en el yacimiento de Vadillos-1 (Hoces de Beteta, Cuenca). Palaeontol. Publ. 2022, 2, 3181. [Google Scholar]
- Ruiz-Galván, A.; Barroso-Barcenilla, F.; Ortega, F.; Sevilla, P. Crocodyliform dental remains from the Lower Cretaceous sites of the Beteta Gorges (Cuenca, Spain). In A Glimpse of the Past; Abstract Book XV Encuentro Jóvenes Investigadores en Paleontología/XV Encontro de Jovens Investigadores em Paleontologia; Barrios de Pedro, S., Moreno, C.B., de Celis, A., Colmenar, J., Cuesta, E., Martínez, D.G., Gascó, F., Jacinto, A., Malafaia, E., Jiménez, M.M., et al., Eds.; XV EJIP: Lisboa, Portugal, 2017; pp. 353–355. [Google Scholar]
- Prieto, I.; Barroso-Barcenilla, F.; Cambra-Moo, O.; Sevilla, P. Taphonomic approach to the vertebrate remains of the “Hoces de Beteta” Lower Cretaceous sites (Cuenca, Central Spain). In Proceedings of the XIV Encuentro de Jóvenes Investigadores en Paleontología-I International Meeting of Early-Stage Researchers in Palaeontology; Manzanares, E., Ferrón, H.G., Suñer, M., Holgado, B., Crespo, V.D., Mansino, S., Fagoaga, A., Marquina, R., García-Sanz, I., Martínez-Pérez, C., et al., Eds.; Ayuntamiento de Alpuente: Valencia, Spain, 2016; p. 7. [Google Scholar]
- Lindsay, W. A review of the acid technique. In The Care and Conservation of Palaeontological Material; Collins, C., Ed.; Butterworth-Heinemann: Oxford, UK, 1995; pp. 99–105. [Google Scholar]
- Leiggi, P.; May, P. (Eds.) Vertebrate Paleontological Techniques; Cambridge University Press: Cambridge, UK, 2005; Volume 1, 366p. [Google Scholar]
- Davidson, A.; Alderson, S.; Fox, M. Assembling an archival marking kit for paleontological specimens. J. Vertebr. Paleontol. 2006, 26, 54A. [Google Scholar]
- Owen, R. Report on British fossil reptiles, Part II. Rep. Br. Assoc. Adv. Sci. 1842, 11, 60–204. [Google Scholar]
- Seeley, H.G. On the classification of the fossil animals commonly named Dinosauria. Proc. R. Soc. Lond. 1887, 43, 165–171. [Google Scholar]
- Marsh, O.C. Principal Characters of American Jurassic Dinosaurs. Part V. Am. J. Sci. 1881, 21, 417–423. [Google Scholar] [CrossRef]
- Mantell, G.A. Notice on the Iguanodon, a newly discovered fossil reptile, from the sandstone of Tilgate Forest in Sussex. Philos. Trans. R. Soc. Lond. 1825, 115, 179–186. [Google Scholar]
- Paul, G.S. A revised taxonomy of the iguanodont dinosaur genera and species. Cretac. Res. 2008, 29, 192–216. [Google Scholar] [CrossRef]
- Horner, J.R.; Weishampel, D.B.; Forster, C.A. Hadrosauridae. In The Dinosauria, 2nd ed.; Weishampel, B., Dodson, P., Osmolska, H., Eds.; University of California Press: Berkeley, CA, USA, 2004; pp. 438–463. [Google Scholar]
- Verdú, F.J.; Godefroit, P.; Royo-Torres, R.; Cobos, A.; Alcalá, L. Individual variation in the postcranial skeleton of the Early Cretaceous Iguanodon bernissartensis (Dinosauria: Ornithopoda). Cretac. Res. 2017, 74, 65–86. [Google Scholar] [CrossRef]
- Hübner, T. The postcranial ontogeny of Dysalotosaurus lettowvorbecki (Ornithischia: Iguanodontia) and implications for the evolution of ornithopod dinosaurs. Palaeontogr. Abh. 2018, 310, 43–120. [Google Scholar] [CrossRef]
- Lockwood, J.A.F.; Martill, D.M.; Maidment, S.C.R. Comptonatus chasei, a new iguanodontian dinosaur from the Lower Cretaceous Wessex Formation of the Isle of Wight, Southern England. J. Syst. Palaeont. 2024, 22, 2346573. [Google Scholar] [CrossRef]
- García-Cobeña, J.; Verdú, F.J.; Cobos, A. Abundance of large ornithopod dinosaurs in the El Castellar Formation (Hauterivian-Barremian, Lower Cretaceous) of the Peñagolosa sub-basin (Teruel, Spain). J. Iber. Geol. 2022, 48, 107–127. [Google Scholar] [CrossRef]
- Bonsor, J.A.; Lockwood, J.A.F.; Leite, J.V.; Scott-Murray, A.; Maidment, S.C.R. The osteology of the holotype of the British iguanodontian dinosaur Mantellisaurus atherfieldensis. Monogr. Palaeont. Soc. 2023, 177, 1–63. [Google Scholar] [CrossRef]
- Norman, D.B. Iguanodontian taxa (Dinosauria: Ornithischia) from the Lower Cretaceous of England and Belgium (Chapter 15). In Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems; Godefroit, P., Ed.; Indiana University Press: Bloomington, IN, USA, 2012; pp. 175–212. [Google Scholar]
- García-Cobena, J.; Verdú, F.J.; Cobos, A. Systematic of a massively constructed specimen of Iguanodon galvensis (Ornithopoda, Iguanodontidae) from the early Barremian (Early Cretaceous) of Eastern Spain. Diversity 2024, 16, 586. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).