
When Abnormality Becomes Perennial in a Reduced Population: The Case of Altudostephanus longicostis gen. et sp. nov. (Valanginian Ammonites, South-Eastern France)


Abstract
1. Introduction
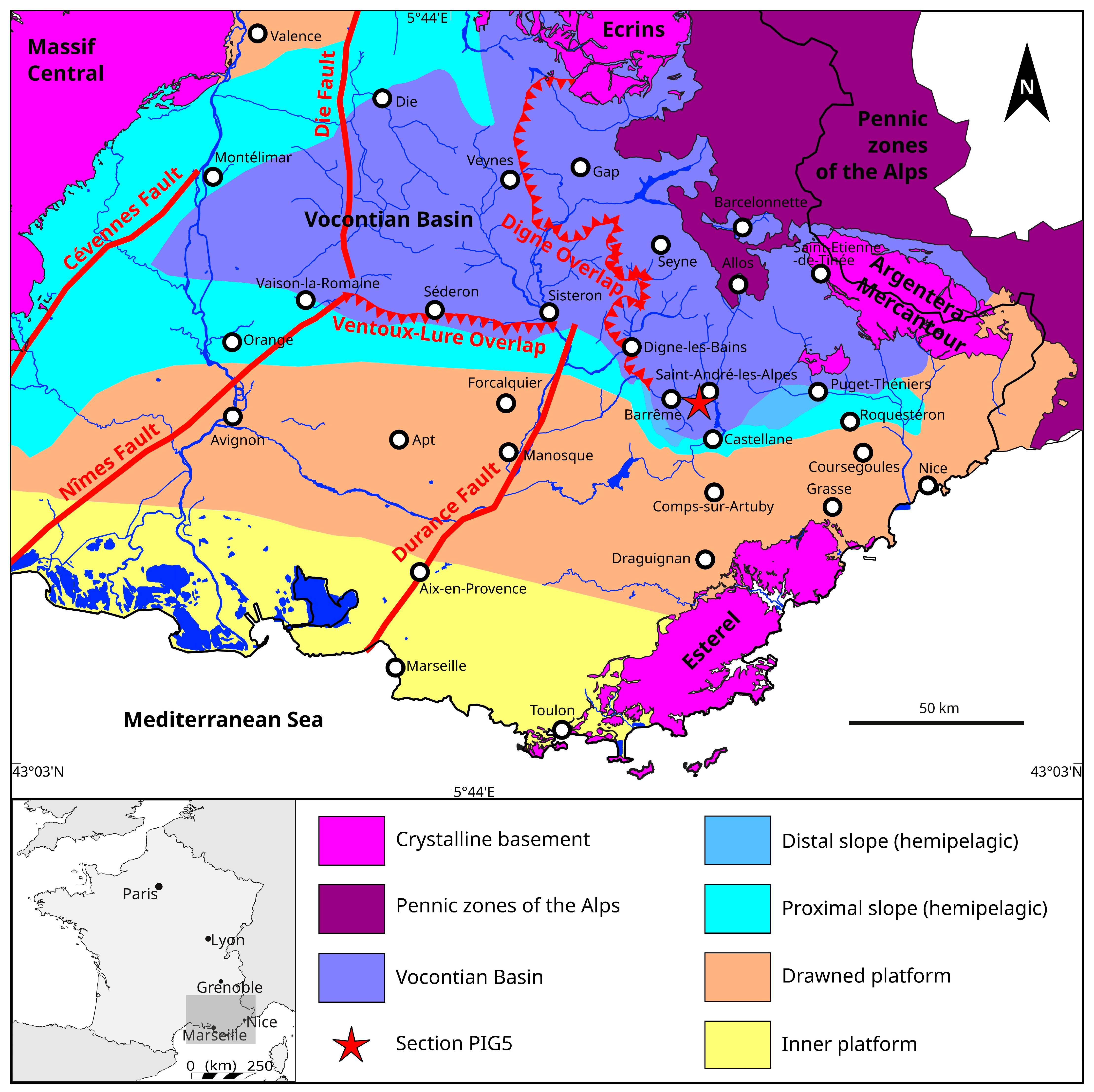
2. Geological Setting
Section PIG5
3. Material and Methods
3.1. Material and Its Conservation
- Cleaning the specimens with water makes it possible, on the one hand, to eliminate labile mineral species by conditioning with water (these are mainly oxidized species such as hydrated ferric sulphates Fe2(SO4)3, n H2O with 0 < n < 9—Sorrenti, 2007 [16]), and on the other hand, to remove mechanically any trace of powdery alteration with a fine brush. It is followed by complete passive drying;
- Neutralization of the effects of oxidation was performed using an acid (Newman, 1998 [15]): here, the choice was made to place the specimens in a bath of 20% water-diluted phosphoric acid at room temperature (H3PO4—Prytulak, 1991 [17]). The duration of the bath is 24–48 h depending on the response of the specimens (regular visual inspection). It allows the removal of the oxidation by-products; especially, the PO43− molecule reacts with Fe3+ on the pyrite surface (Evangelou, 1995 [18]; Evangelou and Zhang, 1995 [19]; Elsetinow et al., 2001 [20]), and here the phosphate molecule also acts to passivate the Fe3+. This step is followed by rinsing the specimens under running water with a fine brush, in order to remove any loosened rust and to thoroughly neutralize the excess phosphoric acid, including in the cracks. Any possible powdery parts that disintegrate are removed again. Rinsing is followed by complete passive drying;
- Passivation of the specimens: treatment by passivation with humic acid use was recommended by Belzile et al. (1997) [21], which involves coating the specimens with a substance that renders them impenetrable to oxidative attack. Passivating the surface of the ‘pyrite’ with organic molecules makes it possible to inhibit the surface chemical reactivity of the mineral (adsorption). Here, we have chosen passivation with tannic acid water diluted to 10% (Prytulak, 1991 [17]), which is a non-toxic and practical product to use. Thoroughly dry specimens are soaked for several seconds to tens of seconds in the tannic acid solution, ensuring that the product penetrates everywhere into the cracks. Tannic acid (C76H52O46), in contact with the ferric ion Fe3+, is transformed into tannate iron (III) Fe2(C14H7O9)(OH)3. This step is followed by complete passive drying by turning the specimens regularly to avoid any accumulation of tannic acid by gravity. An observation phase lasting several days, or even weeks, will ensure that the deterioration process does not resume. Any recovery must be processed again from step 2;
- Installation of the gas-proofing barrier: wax-based products, as well as liquid thermosetting resins, have yielded poor results and age poorly over time. They can crack under the effect of the contraction/expansion of the ‘pyrite’ with temperature differences, which does not make them perfectly impermeable to gases, with the risk that the degassing of these products would eventually produce oxidizers to further degrade the pyrite surface (Tacker, 2020 [13]). In addition, the thickness of the coating provides a glossy finish, coating the finest details of the specimens and making them barely visible for subsequent studies (Figure 4a; Larkin, 2011, p. 41 [12]). Treatment by encapsulation with phospholipids yields good results by protecting the specimens from oxygen, provided that they are totally immersed into the lipid fluid: the hydrophobic pocket in the bilayer lipid structure can inhibit the interaction of water with the pyrite surface and hence inhibit or suppress the deterioration process (Kargbo et al. 2004 [22]). This technique, although it stops oxidation by 90%, does not allow treatment of damaged areas on its own. It can also be complicated to be implemented in the conservation conditions of collections and hinders the subsequent handling of specimens and labels. The use of silicone seems to yield good results for stabilized specimens; it is also inert, impermeable to air and non-hazardous (Collinson et al. 2016 [23]; Newman, 1998 [15]; Fenlon and Petrera, 2019 [24]), but it dries slowly. As an alternative, we recommend treatment with residue-free Polytetrafluoroethylene (PTFE), which has the advantage of being quick-drying. Just like silicone, it has significant oxygen and moisture barrier capabilities. The use of PTFE also has a practical side by allowing the study and easy handling of treated specimens once dry. Their interest lies in the presence of fluorine in the molecules, this element providing the polymer a greater chemical stability and high resistance to ultraviolet rays, without drying shrinkage (Bromblet, 2002 [25]). Complete drying is achieved after a period of several days;
- Of course, this preventive/curative protocol does not preclude ensuring long-term conservation of the treated specimens in a place protected from humidity and temperature variations. The use of silica gel desiccants is also recommended, as well as the implementation of a collection survey protocol. Finally, producing of replicas or CT- or 3D-surface scans would ensure that the morphology is conserved, even if the specimens deteriorate despite all efforts.
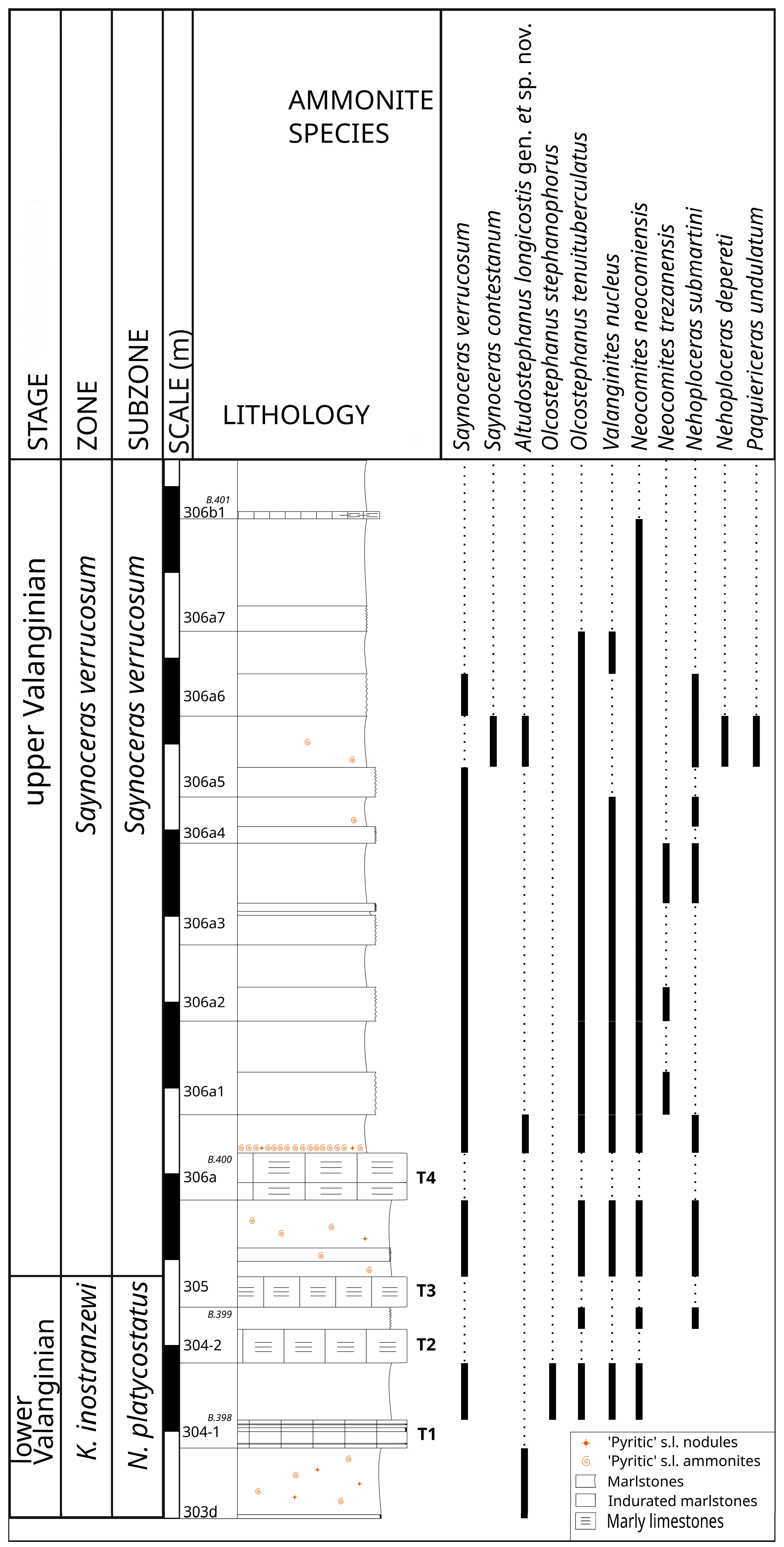
3.2. Stratigraphy
4. Systematic Palaeontology
- This latter specimen was not mentioned by Cottreau in the revision of the Prodrome types, who explicitly stated that one single specimen was attributed to this species by d’Orbigny (4861, now MNHN.F.R03115). As this is the only specimen, then this is unquestionably the holotype and there cannot be any other syntypes, according to the rules of the ICZN;
- Its morphological characters (radial primary ribs; secondary ribs thin and well expressed on the ventral area; absence of the angular shoulder forming a ledge at the termination of the primary ribs; constrictions much rarer; prominent and rounded tubercles) do not allow assigning the specimen MNHN.F.A00067 to A. josephinus;
- It is indicated that the specimen MNHN.F.A00067 comes from Saint-Julien-en-Beauchêne, a locality situated in the northern part of the Vocontian Basin (northwest of Veynes in Figure 1). This is definitely not the case: the obvious presence of glauconite in the matrix and its light beige phosphate limestone allow us to affirm without ambiguity that it comes from the area of drowned platforms surrounding the Vocontian Basin. This type of preservation is known, for example, in the Arc de Castellane to the south of the basin (Cotillon, 1971 [30]);
- Bulot (1990, 1995 [6,7]), then more recently Kenjo (2014) [27], showed that A. josephinus gen. nov. is present only in the lower Valanginian (see below the stratigraphic occurrence). However, fossils with the same preservation as specimen MNHN.F.A00067 are known on platform edges in significantly higher levels, near to the boundary between the Valanginian and the Hauterivian (e.g., Autran, 1993 [31]);
- All the previous points allow us to consider the specimen MNHN.F.A00067 as Olcostephanus balkanicus (Tzankov, 1942).
- The innermost whorls, up to approximately D = 0.7 mm, are smooth on the flanks (the venter is not visible at this diameter on the specimens studied) and have no visible ornamentation apart from the constrictions (see below);
- The rest of the known shell (Dmax = 19 mm) shows an ornamentation consisting in primary ribs, relatively frequent (9–14 per half-whorl), but irregularly spaced. On the flanks, they are concave towards the aperture in their lower part, then proverse in their upper part. At the angular ledge between the flanks and the venter, the primary ribs are reinforced by a nodular punctiform tubercle. The ledge of the angular shoulder therefore shows an irregular crenulated appearance. Very faint constrictions are visible from time to time along the same line. Their number and vigor are very irregular and they are generally associated with the primary ribs that they precede. On the venter, where they are better marked and bordered by faint ridges, they continue their proximal projection so as to present a strong concavity towards the rear of the shell at the siphonal area. Transverse striae irregular pattern is also present with the same path (DBT.BK35). Conversely, the primary ribs diminish considerably on the venter where they are only evanescent and difficult to perceive. The most striking aspect of the ornamentation consists of the presence of secondary ribs reversed perpendicularly to the primary ribs, thus forming a longitudinal ornamentation in the direction of the coiling. These ribs are slightly wavy at the intersection of the primary ribs. There are 7 longitudinal ribs per half of the ventral area in specimens where it is sufficiently preserved (i.e., 14 over the entire ventral area). This longitudinal ventral pattern appears smoothly from D = 7 mm, whereas at his diameter the constrictions, there ridges, and megastriae (when present) are fully expressed.
5. Discussion
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mironenko, A.A. The soft-tissue attachment scars in Late Jurassic ammonites from Central Russia. Acta Palaeontol. Pol. 2015, 60, 981–1000. [Google Scholar] [CrossRef]
- Ploch, I. Taxonomic interpretation and sexual dimorphism in the Early Cretaceous (Valanginian) ammonite Valanginites nucleus (ROEMER,1841). Acta Geol. Pol. 2003, 53, 201–208. [Google Scholar]
- Busnardo, R.; Thieuloy, J.-P.; Moullade, M. Hypostratotype mesogéen de l’étage Valanginien (Sud-Est de la France). Ed. Du CNRS Les Strat. Français 1979, 6, 61. [Google Scholar]
- Bert, D.; Bousquet, V.; Guiomar, M.; Bariani, F.; Hippolyte, J.-C.; Bromblet, P.; Garciaz, J.-L.; Fleury, J.; Mathieu, P.; Pagès, J.-S.; et al. The Geotouristic Project ‘the Geological Adventure’ to the Rescue of an Iconic World Heritage Geosite, the Ammonites Slab of Digne-les-Bains (National Geological Nature Reserve of Haute-Provence and Unesco Global Geopark, France). Geoheritage 2022, 14, 101. [Google Scholar] [CrossRef]
- Thieuloy, J.-P. Les ammonites boréales des formations néocomiennes du Sud-Est de la france (Province Subméditerranéenne). Géobios 1977, 10, 395–461. [Google Scholar] [CrossRef]
- Bulot, L. Evolution des Olcostephaninae (Ammonitinae, Cephalopoda) dans le Contexte Paléobiogéographique du Crétacé Inférieur (Valanginien-Hauterivien) du Sud-Est de la France; DSER, Université de Dijon: Dijon, France, 1990; 177p. [Google Scholar]
- Bulot, L. Les Formations à Ammonites du Crétacé Inférieur du Sud-Est de la France (Berriasien à Hauterivien): Biostratigraphie, Paléontologie et Cycles Sédimentaires. Ph.D. Thesis, Muséum National d’Histoire Naturelle, Paris, France, 1995; 398p. [Google Scholar]
- Reboulet, S. L’évolution des ammonites du Valanginien-Hauterivien inférieur du Bassin Vocontien et de laplate-forme provençale (Sud-Est de la France: Relations avec la stratigraphie séquentielle et implications biostratigraphiques. Doc. Lab. Géol. Lyon 1995, 137, 7–371. [Google Scholar]
- Bulot, L.; Thieuloy, J.-P. Les biohorizons du Valanginien du Sud-Est de la France: Un outil fondamental pour les corrélations au sein de la Téthys occidentale. Géologique Alp. Mémoire Hors-Série 1994, 20, 15–41. [Google Scholar]
- Martinez, M. Calibration astronomique du Valanginien et de l’Hauterivien (Crétacé inférieur): Implications Paléoclimatiques et Paléocéanographiques. Ph.D. Thesis, Université de Bourgogne, Dijon, France, 2013. [Google Scholar]
- Rouchon, V.; Badet, H.; Belhaj, O.; Bonnerot, O.; Lavédrine, B.; Michard, J.-G.; Miska, S. Raman and FTIR spectroscopy applied to the conservation report of paleontological collections: Identification of Raman and FTIR signatures of several ironsulfate species such as ferrinatrite and sideronatrite. J. Raman Spectrosc. 2011, 43, 1265–1274. [Google Scholar] [CrossRef]
- Larkin, N.R. Pyrite Decay: Cause and effect, prevention and cure. Nat. SCA News 2011, 21, 35–43. [Google Scholar]
- Tacker, R.C. A review of “pyrite disease” for paleontologists, with potential focused interventions. Palaeontol. Electron. 2020, 23, a45. [Google Scholar] [CrossRef]
- Schronk, K.R. Assessing the stability of mineralogical collections in museums. Ph.D. Thesis, University of Oxford, Oxford, UK, 2023; 222p. [Google Scholar]
- Newman, A. Pyrite oxidation and museum collections: A review of theory and conservation treatments. Geol. Curator 1998, 6, 363–371. [Google Scholar] [CrossRef]
- Sorrenti, E. Étude de la Passivation de la Pyrite: Chimie de Surface et Réactivité; Institut National Polytechnique de Lorraine: Nancy, France, 2007. [Google Scholar]
- Prytulak, G. The treatment of rusted machinery in preparation for surface coating. J. Int. Inst. Conserv. 1991, 16, 23–34. [Google Scholar]
- Evangelou, V.P. Potential Microencapsulation of Pyrite by Artificial Inducement of Ferric Phosphate Coatings. J. Environ. Qual. 1995, 24, 535–542. [Google Scholar] [CrossRef]
- Evangelou, V.P.; Zhang, Y.L. A review: Pyrite oxidation mechanisms andacid mine drainage prevention. Crit. Rev. Environ. Sci. Technol. 1995, 25, 141–199. [Google Scholar] [CrossRef]
- Elsetinow, A.R.; Schoonen, M.A.; Strongin, D.R. Aqueous geochemical and surface science investigation of the effect of phosphate on pyrite oxidation. Environ. Sci. Technol. 2001, 35, 2252–2257. [Google Scholar] [CrossRef] [PubMed]
- Belzile, N.; Maki, S.; Chen, Y.-W.; Goldsack, D. Inhibition of pyrite oxidation by surface treatment. Sci. Total Environ. 1997, 196, 177–186. [Google Scholar] [CrossRef]
- Kargbo, D.M.; Atallah, G.; Chatterjee, S. Inhibition of pyrite oxidation by a phospholipid in the presence of silicate. Environ. Sci. Technol. 2004, 38, 3432–3441. [Google Scholar] [CrossRef] [PubMed]
- Collinson, M.E.; Adam, N.F.; Manchester, S.R.; Stull, G.W.; Herrera, F.; Smith, S.Y.; Andrew, M.J.; Kenrick, P.; Sykes, D. X-ray micro-computed tomography (micro-CT) of pyrite-permineralized fruits and seeds from the Lonfon Clay Formation (Ypresian) conserved in silicon oil: A critical evaluation. Botany 2016, 94, 697–711. [Google Scholar] [CrossRef]
- Fenlon, A.; Petrera, L. Pyrite oxidation: A history of treatments at the Natural History Museum, London. Geol. Curator 2019, 11, 9–18. [Google Scholar] [CrossRef]
- Bromblet, P. Consolidation et hydrofugation de la pierre. Monum. Rev. Sci. Et Tech. Des Monum. Hist. 2002, 2002, 201–243. [Google Scholar]
- Szives, O.; Moreno-Bedmar, J.A.; Aguirre-Urreta, B.; Company, M.; Frau, C.; López-Horgue, M.; Pictet, A.; Ploch, I.; Salazar, C.; Barragán, R.; et al. Report on the 7th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the KilianGroup (Warsaw, Poland, 21st August 2022): State of the art on the current Standard Ammonite Zonation of the Western Tethyan Mediterranean Province. Cretac. Res. 2023, 153, 105716. [Google Scholar] [CrossRef]
- Kenjo, S. Biostratigraphie Intégrée à Nannofossiles Calcaires et Ammonoïdes: Développement et Implications Pour la Définition et la Valorisation Des Stratotypes d’unité et de Limite. L’exemple des étages Berriasien et Valanginien et de Leur Limite (~140 Millions d’années). Ph.D. Thesis, University Claude Bernard Lyon 1, Villeurbanne, France, 2014; 273p. [Google Scholar]
- Orbigny, A.D.D. Prodrome de paléontologie stratigraphique universelle des animaux mollusques et rayonnés, faisant suite au cours élémentaire de paléontologie et de géologie stratigraphique. Masson 1850, 2, 427. [Google Scholar]
- Cottreau, J. Types du Prodrome de paléontologie stratigraphique universelle de d’Orbigny. Ann. Paléontol. 1934, 23, 45–80. [Google Scholar]
- Cotillon, P. Le Crétacé Inférieur de l’arc Subalpin de Castellane Entre l’Asse et le Var—Stratigraphie et Sédimentologie. Ph.D. Thesis, University Claude Bernard Lyon 1, Villeurbanne, France, 1971. Volume 68. pp. 1–329. [Google Scholar]
- Autran, G. L’évolution de la marge Nord-Est provençale (Arc de Castellane) du Valanginien moyen à l’Hauterivien à travers l’analyse biostratigraphique des séries de la région de Peyroules: Séries condensées, discontinuités et indices d’une tectonogenèse distensive. Paléobiologie. Ann. Muséum D’histoire Nat. Nice 1993, 10, 240. [Google Scholar]
- Faraoni, P.; Flore, D.; Marini, A.; Pallini, G.; Pezzoni, N. Valanginian and early Hauterivian ammonite successions in the Mt Catria group (Central Apennines) and in the Lessini Mts (Southern Alps), Italy. Palaeopelagos 1997, 7, 59–100. [Google Scholar]
- De Beats, K.; Keupp, H.; Klug, C. Parasites of Ammonoids. Ammonoid Palaeobiol. 2015, 20, 837–875. [Google Scholar]
- Keupp, H. Atlas zur Paläopathologie der Cephalopoden. Berl. Paläobiol. Abh. 2012, 12, 392p. [Google Scholar]
- Hoffmann, R.; Keupp, H. Ammonoid Paleopathology. Ammonoid Palaeobiol. 2015, 21, 877–926. [Google Scholar]
- Urdy, S. Theoretical modeling of the Molluscan shell: What has been learned from the comparison among Molluscan taxa? Ammonoid Palaeobiol. 2015, 6, 207–251. [Google Scholar]
- Dommergues, J.-L.; David, B.; Marchand, D. Les relations ontogenèse-phylogenèse: Applications paléontologiques. Géobios 1986, 19, 335–356. [Google Scholar] [CrossRef]
- Rogov, M.A. Pseudoinversion of septal sutures in Middle Jurassic-Lower Cretaceous non-heteromorph ammonites. Swiss J. Palaeontol. 2019, 138, 163–176. [Google Scholar] [CrossRef]
- Lachance, J. Hardy-Weinberg equilibrium and randommating. Encycl. Evol. Biol. 2016, 208–211. [Google Scholar] [CrossRef]
- Angst, P.; Ameline, C.; Haag, C.R.; Ben-Ami, F.; Ebert, D.; Fielf, P.D. Genetic drift shapes the evolution of a highly dynamicmetapopulation. Mol. Biol. Evol. 2022, 39, msac264. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Standard Tethyan Ammonite Zonation of Szives et al., 2023 [26] | Distribution of the Species (from Bulot, 1995 [7], Kenjo, 2014 [27] and This Work) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Stages | Zones | Subzones | Barronites hirsutus (Fallot et Termier, 1923) | Altudostephanus josephinus gen. nov. | Altudostephanus longicostis gen. et sp. nov. | Olcostephanus stephanophorus (Matheron, 1878) | Olcostephanus tenuituberculatus Bulot, 1905 | Olcostephanus drumensis Kilian, 1910 | Valanginites nucleus (Roemer, 1841) | |
| Valanginian | upper | Criosarasinella furcillata | Teschenites callidiscus | |||||||
| Criosarasinella furcillata | ||||||||||
| Neocomites peregrinus | Olcostephanus (Olcostephanus) nicklesi | |||||||||
| Neocomites peregrinus | ||||||||||
| Saynoceras verrucosum | Karakashiceras pronecostatum | |||||||||
| Saynoceras verrucosum | ||||||||||
| lower | Karakashiceras inostranzewi | Neocomites platycostatus | ||||||||
| Karakashiceras inostranzewi | ||||||||||
| Neocomites neocomiensiformis | Busnardoites campylotoxus | |||||||||
| Neocomites neocomiensiformis | ||||||||||
| ‘’Thurmanniceras’’ pertransiens | Neolissoceras (Vergoliceras) salinarium | |||||||||
| Neocomites premolicus | ||||||||||
| Specimens | Section/Level | D | H | W | U | H/D | W/H | U/D | U/H | N1/2 | N2/2 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| DBT.BK35 (holotype) | PIG5/306a5m | 12.74 | 5.00 | c.8.32 | 4.28 | 0.39 | 1.66 | 0.34 | 0.86 | 9 | 7 |
| DBT.BK36 | PIG5/304-2m | c.10.80 | 3.46 | 6.26 | c.3.60 | 0.32 | 1.81 | 0.33 | 1.04 | ? | 7 |
| DBT.AU38 | C7/306a | 19.08 | 7.16 | - | 7.48 | 0.36 | - | 0.39 | 1.05 | 14 | ? |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bert, D.; Bersac, S.; Beltran, B.; Canut, L. When Abnormality Becomes Perennial in a Reduced Population: The Case of Altudostephanus longicostis gen. et sp. nov. (Valanginian Ammonites, South-Eastern France). Foss. Stud. 2024, 2, 92-109. https://doi.org/10.3390/fossils2020004
Bert D, Bersac S, Beltran B, Canut L. When Abnormality Becomes Perennial in a Reduced Population: The Case of Altudostephanus longicostis gen. et sp. nov. (Valanginian Ammonites, South-Eastern France). Fossil Studies. 2024; 2(2):92-109. https://doi.org/10.3390/fossils2020004
Chicago/Turabian StyleBert, Didier, Stéphane Bersac, Bernard Beltran, and Léon Canut. 2024. "When Abnormality Becomes Perennial in a Reduced Population: The Case of Altudostephanus longicostis gen. et sp. nov. (Valanginian Ammonites, South-Eastern France)" Fossil Studies 2, no. 2: 92-109. https://doi.org/10.3390/fossils2020004
APA StyleBert, D., Bersac, S., Beltran, B., & Canut, L. (2024). When Abnormality Becomes Perennial in a Reduced Population: The Case of Altudostephanus longicostis gen. et sp. nov. (Valanginian Ammonites, South-Eastern France). Fossil Studies, 2(2), 92-109. https://doi.org/10.3390/fossils2020004