
The Diet of Metriorhynchus (Thalattosuchia, Metriorhynchidae): Additional Discoveries and Paleoecological Implications



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Discovery of the Specimen
2. General Description
3. Systematic Paleontology
4. Geology
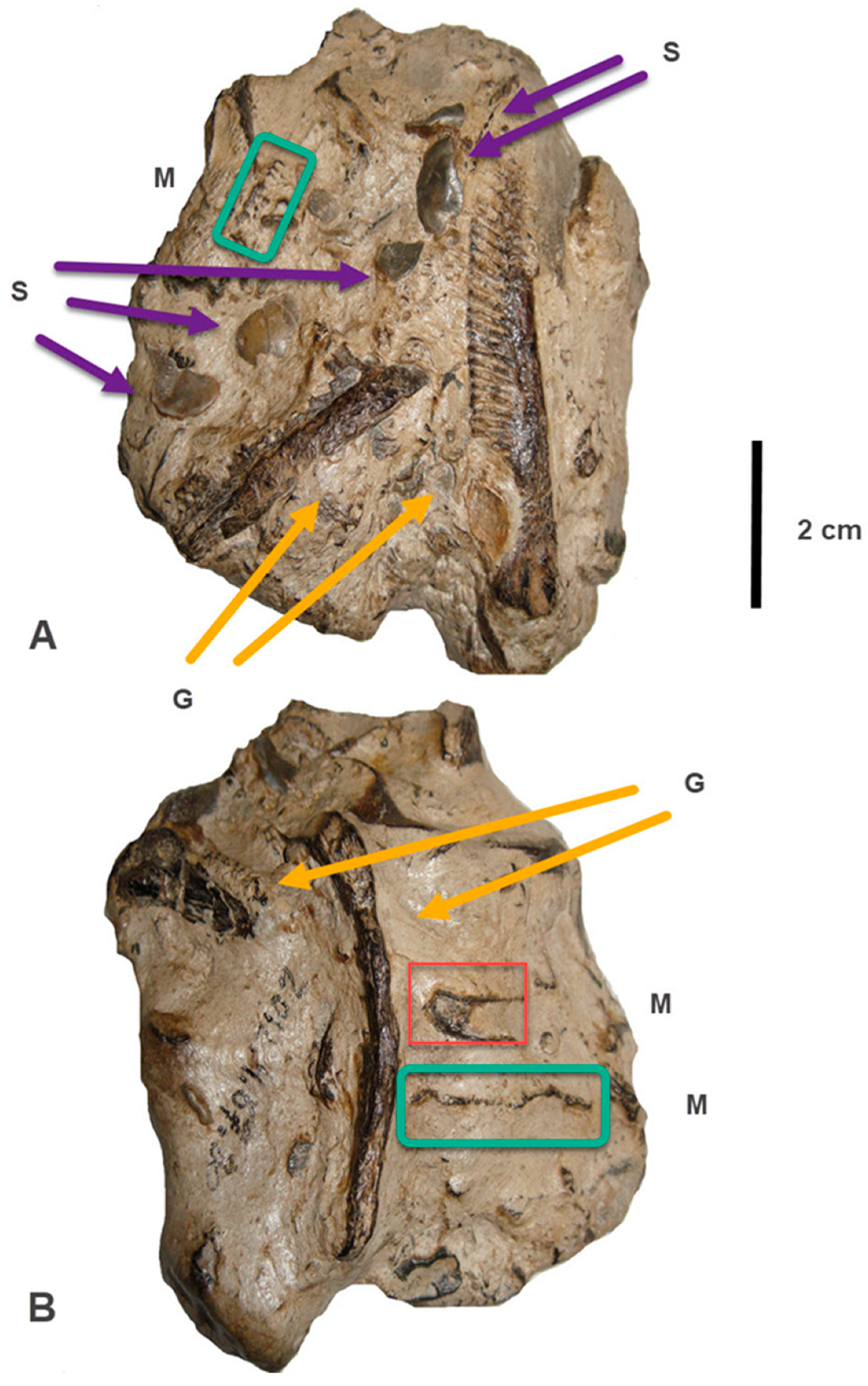
5. Description
6. Paleoecology
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hua, S. Locomotion in marine mesosuchians (Crocodylia): The contribution of the “locomotion profiles”. Neues Jahrb. Für Geol. Und Paläontologie-Abh. 2003, 227, 139–152. [Google Scholar] [CrossRef]
- Lepage, Y.; Buffetaut, E.; Hua, S.; Martin, J.E.; Tabouelle, J. Catalogue descriptif, anatomique, géologique e historique des fossiles présentés à l’exposition “Les Crocodiliens fossiles de Normandie” (6 novembre–14 décembre 2008). Bull. La Société Géologique Normandie Des Amis Du Muséum Du Havre 2008, 95, 5–152. [Google Scholar]
- Arthaber, G. Beiträge zur kenntnis der organisation und der anpassungserscheinung des genus Metriorhynchus. Beiträge Zur Paläontologie Von Osterr. 1906, 19, 287–320. [Google Scholar]
- Auer, E. Uber einige Krokodile der Juraformation. Palaeontographica 1909, 55, 217–294. [Google Scholar]
- Andrews, C.W. A Descriptive Catalogue of Marine Reptiles of the Oxford Clay. Part Two; British Museum (Natural History): London, UK, 1913; 206p. [Google Scholar]
- Le Mort, J.; Martin, J.E.; Picot, L.; Hua, S. First description of the most complete Metriorhynchus aff. superciliosus (Thalattosuchia) specimen from the Callovian of the Vaches-Noires cliffs (Normandy, France) and limitations in the classification of Metriorhynchidae. Ann. Paléontologie 2022, 108, 102539. [Google Scholar] [CrossRef]
- Young, M.T.; Brignon, A.; Sachs, S.; Hornung, J.J.; Foffa, D.; Kitson, J.J.N.; Johnson, M.M.; Steel, L. Cutting the Gordian knot: A historical and taxonomic revision of the Jurassic crocodylomorph Metriorhynchus. Zool. J. Linn. Soc. 2020, 192, 510–553. [Google Scholar] [CrossRef]
- Vignaud, P. Les Thalattosuchia, Crocodiles Marins du Mésozoique: Systématique Phylogénétique, Paléoécologie, Biochronologie et Implications Paléogéographiques. Ph.D. Thesis, University of Poitiers, Poitiers, France, 1995. [Google Scholar]
- Owen, R. Monography on the British fossil Reptilian from the Kimmeridge Clay—Sauropterygia; Paleontological Society: London, UK, 1868; Volume 3, p. 7. [Google Scholar]
- Eudes-Deslongchamps, A. Prodrôme des Téléosauriens du Calvados. In Notes paléontologiques 1867–1869; LeBlanc-Hardel (Caen) & Savy: Paris, France, 1869; Volume 1, pp. 95–354. [Google Scholar]
- Dugué, O.; Fily, G.; Rioult, M. Le Jurassique des côtes du Calvados. Biostratigraphie, Sédimentologie, Paléoécologie, Paléogéographie et Stratigraphie séquentielle. Société Géologique Normandie Des Amis Du Muséum Du Havre 1998, 85, 3–132. [Google Scholar]
- Brochu, C. Closure of neurocentral sutures during crocodilian ontogeny: Implications for maturity assessment in fossil archosaurs. J. Vertebr. Paleontol. 1996, 16, 49–62. [Google Scholar] [CrossRef]
- Liston, J.J. The occurrence of the Middle Jurassic pachycormid fish Leedsichthys. Oryctos 2010, 9, 1–36. [Google Scholar]
- Liston, J.J. Leedsichthys des Vaches Noires au peigne fin. Published (translation by M-C Buchy). L’Écho Des Falaises 2008, 12, 41–49. [Google Scholar]
- Liston, J.J.; Gendry, D. Le Python de Caen, les algues géantes d’Amblie, et autres specimens perdus de Leedsichthys d’Alexandre Bourienne, Jules Morière, Eugène Eudes-Deslongchamps et Alexandre Bigot. L’Écho Des Falaises 2015, 19, 17–33. [Google Scholar]
- Liston, J.J. The Plasticity of Gill Raker Characteristics in Suspension Feeders: Implications for Pachycormiformes; Arratia, G., Schultze, H.-P., Wilson, M.V.H., Eds.; Mesozoic Fishes 5 Global Diversity and Evolution; Verlag Dr. Friedrich Pfeil: München, Germany, 2013; pp. 121–143. [Google Scholar]
- Liston, J.J. A Fish Fit For Ozymandias?: The Ecology, Growth and Osteology of Leedsichthys (Pachycormidae, Actinopterygii). Ph.D. Thesis, University of Glasgow, Scotland, UK, 2007. [Google Scholar]
- Liston, J.J.; Newbrey, M.G.; Challands, T.J.; Adams, C.E. Growth, Age and Size of the Jurassic Pachycormid Leedsichthys problematicus (Osteichthyes: Actinopterygii); Arratia, G., Schultze, H.-P., Wilson, M.V.H., Eds.; Mesozoic Fishes 5 Global Diversity and Evolution; Verlag Dr. Friedrich Pfeil: München, Germany, 2013; pp. 145–175. [Google Scholar]
- Ferrón, H.G.; Holgado, B.; Liston, J.J.; Martínez-Pérez, C.; Botella, H. Assessing metabolic constraints on the maximum body size of actinopterygians: Locomotion energetics of Leedsichthys problematicus (Actinopterygii: Pachycormiformes). Palaeontology 2018, 61, 775–783. [Google Scholar] [CrossRef]
- Liston, J.J.; Bronzati, M.; Vernygora, O. ‘Raking illumination’: Shedding light on the utility of feeding structures as indicators of relatedness. Vertebr. Anat. Morphol. Palaeontol. 2022, 10, 24–29. [Google Scholar]
- Liston, J.J. Leedsichthys problematicus: Arthur Smith Woodward’s ‘most embarrassing enigma’. Geol. Soc. Lond. Spec. Publ. 2016, 430, 235–259. [Google Scholar] [CrossRef]
- Johanson, Z.; Liston, J.J.; Davesne, D.; Challands, T.J. Mechanisms of dermal bone repair after predatory attack in the giant stem-group teleost Leedsichthys problematicus Woodward, 1889a (Pachycormiformes). J. Anat. 2022, 241, 393–406. [Google Scholar] [CrossRef]
- Martill, D.M. The diet of Metriorhynchus, a Mesozoic marine crocodile. Neues Jahrb. Für Geol. Und Paläontologie-Monatshefte 1986, 10, 621–625. [Google Scholar] [CrossRef]
- Danise, S.; Twitchett, R.; Matts, K. Ecological succession of a Jurassic shallow-water ichthyosaur fall. Nat. Commun. 2014, 5, 4789. [Google Scholar] [CrossRef]
- Massare, J.A. Tooth morphology and prey preference of Mesozoic marine reptiles. J. Vertebr. Paleontol. 1987, 7, 121–137. [Google Scholar] [CrossRef]
- Hua, S.; Pennetier, E.; Pennetier, G. A juvenile Steneosaurus in the Callovian of Normandy (France); a genus too hastily consigned to the wastebasket? Carnets Nat. 2021, 8, 1–8. [Google Scholar]
- Vignaud, P. La morphologie dentaire des Thalattosuchia (Crocodylia, Mesosuchia). Paleaeovertebrata 1997, 26, 35–39. [Google Scholar]
- Massare, J.A. Swimming capabilities of Mesozoic Marine reptiles: Implications for method of predation. Paleobiology 1988, 14, 187–205. [Google Scholar] [CrossRef]
- Massare, J.A. Swimming capabilities of Mesozoic marine reptiles: A review. In Mechanics and Physiology of Animal Swimming; Cambridge University Press: Cambridge, UK, 1994; pp. 133–149. [Google Scholar]
- Hua, S. Hydrodynamique et modalités d’allègement chez Metriorhynchus superciliosus (Crocodylia, Thalattosuchia): Implications paléoécologiques. Neues Jahrb. Fur Paläontologie Und Geol. Abh. 1994, 193, 1–19. [Google Scholar]
- Hua, S.; De Buffrenil, V. Histology of the Thalattosuchia as a clue of the interpretation of functional adaptations in the Thalattosuchian (Reptilia, Crocodylia). J. Vertebr. Paleontol. 1996, 16, 703–717. [Google Scholar] [CrossRef]
- Gandola, R.; Buffetaut, E.; Monaghan, N.; Dyke, G. Salt glands in the fossil crocodile Metriorhynchus. J. Vertebr. Paleontol. 2006, 26, 1009–1010. [Google Scholar] [CrossRef]
- Hua, S. Adaptations des Crocodiliens Mésosuchiens au Milieu Marin. Ph.D. Thesis, Université Pierre-et-Marie-Curie, Paris, France, 1997. [Google Scholar]
- Hua, S.; Vignaud, P.; Efimov, V. First record of Metriorhynchidae (Crocodylomorpha, Mesosuchia) in the Upper Jurassic of Russia. Neues Jahrb. Für Geol. Und Paläontologie-Monatshefte 1998, 8, 475–484. [Google Scholar] [CrossRef]
- Young, M.T.; Zverkov, N.Y.; Arkhangelsky, M.S.; Ippolitov, A.P.; Meleshin, I.A.; Mirantsev, G.V.; Shmakov, A.S.; Stenshin, I.M. Thalattosuchian crocodylomorphs from European Russia, and new insights into metriorhynchid tooth serration evolution and their palaeolatitudinal distribution. PeerJ 2023, 11, e15781. [Google Scholar] [CrossRef]
- Hua, S. A propos d’un crâne de Dyrosaurus phosphaticus (Thomas, 1893) (Crocodylia, Mesosuchia): Contribution à l’interprétation paléoécologique et biogéographique des Dyrosauridae. Bull. La Société Belg. Géologie 1995, 104, 109–118. [Google Scholar]
- Andrade, M.B.; Young, M.T.; Desojo, J.; Brusatte, S.L. The evolution of extreme hypercarnivory in Metriorhynchidae (Mesoeucrocodylia: Thalattosuchia) based on evidence from microscopic denticle morphology. J. Vertebr. Paleontol. 2010, 30, 1451–1465. [Google Scholar] [CrossRef]
- Young, M.T.; Andrade, M.B.; Brusatte, S.L.; Sakamoto, M.; Liston, J.J. The oldest known metriorhynchid super-predator: A new genus and species from the Middle Jurassic of England, with implications for serration and mandibular evolution in predacious clades. J. Syst. Palaeontol. 2013, 11, 475–513. [Google Scholar] [CrossRef]
- Young, M.T.; Stephen, L.; Brusatte, S.L.; Beatty, B.L.; Brandalise De Andrade, M.; Desojo, J.B. Tooth-On-Tooth Interlocking Occlusion Suggests Macrophagy in the Mesozoic Marine Crocodylomorph Dakosaurus. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2012, 295, 1147–1158. [Google Scholar] [CrossRef]
- Seon, N.; Amiot, R.; Martin, J.A.; Young, M.T.; Middleton, H.; Fourel, F.; Picot, L.; Valentin, X.; Lecuyer, C. Thermophysiologies of Jurassic marine crocodyliforms inferred from the oxygen isotope composition of their tooth apatite. Philos. Trans. R. Soc. Lond. 2020, B375, 20190139. [Google Scholar] [CrossRef] [PubMed]
- Forrest, R. Evidence for scavenging by the marine crocodile Metriorhynchus on the carcass of a plesiosaur. Proc. Geol. Assoc. 2003, 114, 363–366. [Google Scholar] [CrossRef]
- Allen, G.R. The marine crocodile Crocodylus porosus from Ponape, Eastern Caroline Islands, with notes on food habits of crocodiles from the Palau archipelago. Copeia 1974, 2, 553. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hua, S.; Liston, J.; Tabouelle, J. The Diet of Metriorhynchus (Thalattosuchia, Metriorhynchidae): Additional Discoveries and Paleoecological Implications. Foss. Stud. 2024, 2, 66-76. https://doi.org/10.3390/fossils2010002
Hua S, Liston J, Tabouelle J. The Diet of Metriorhynchus (Thalattosuchia, Metriorhynchidae): Additional Discoveries and Paleoecological Implications. Fossil Studies. 2024; 2(1):66-76. https://doi.org/10.3390/fossils2010002
Chicago/Turabian StyleHua, Stéphane, Jeff Liston, and Jérôme Tabouelle. 2024. "The Diet of Metriorhynchus (Thalattosuchia, Metriorhynchidae): Additional Discoveries and Paleoecological Implications" Fossil Studies 2, no. 1: 66-76. https://doi.org/10.3390/fossils2010002
APA StyleHua, S., Liston, J., & Tabouelle, J. (2024). The Diet of Metriorhynchus (Thalattosuchia, Metriorhynchidae): Additional Discoveries and Paleoecological Implications. Fossil Studies, 2(1), 66-76. https://doi.org/10.3390/fossils2010002