
Taxonomic Status of Nanotyrannus lancensis (Dinosauria: Tyrannosauroidea)—A Distinct Taxon of Small-Bodied Tyrannosaur



Abstract
1. Introduction
1.1. The Tyrannosaurs, the Most Iconic of All Dinosaurs
1.2. Abbreviations
1.3. Tyrannosaurus rex
1.4. Nanotyrannus
1.5. Purpose and Approach of This Paper
- Patterns of diversity in Tyrannosauridae and other predators. The frequent coexistence of two species of tyrannosaur suggests that this pattern is the rule; the existence of a distinct taxon alongside Tyrannosaurus is expected from known patterns of dinosaur diversity.
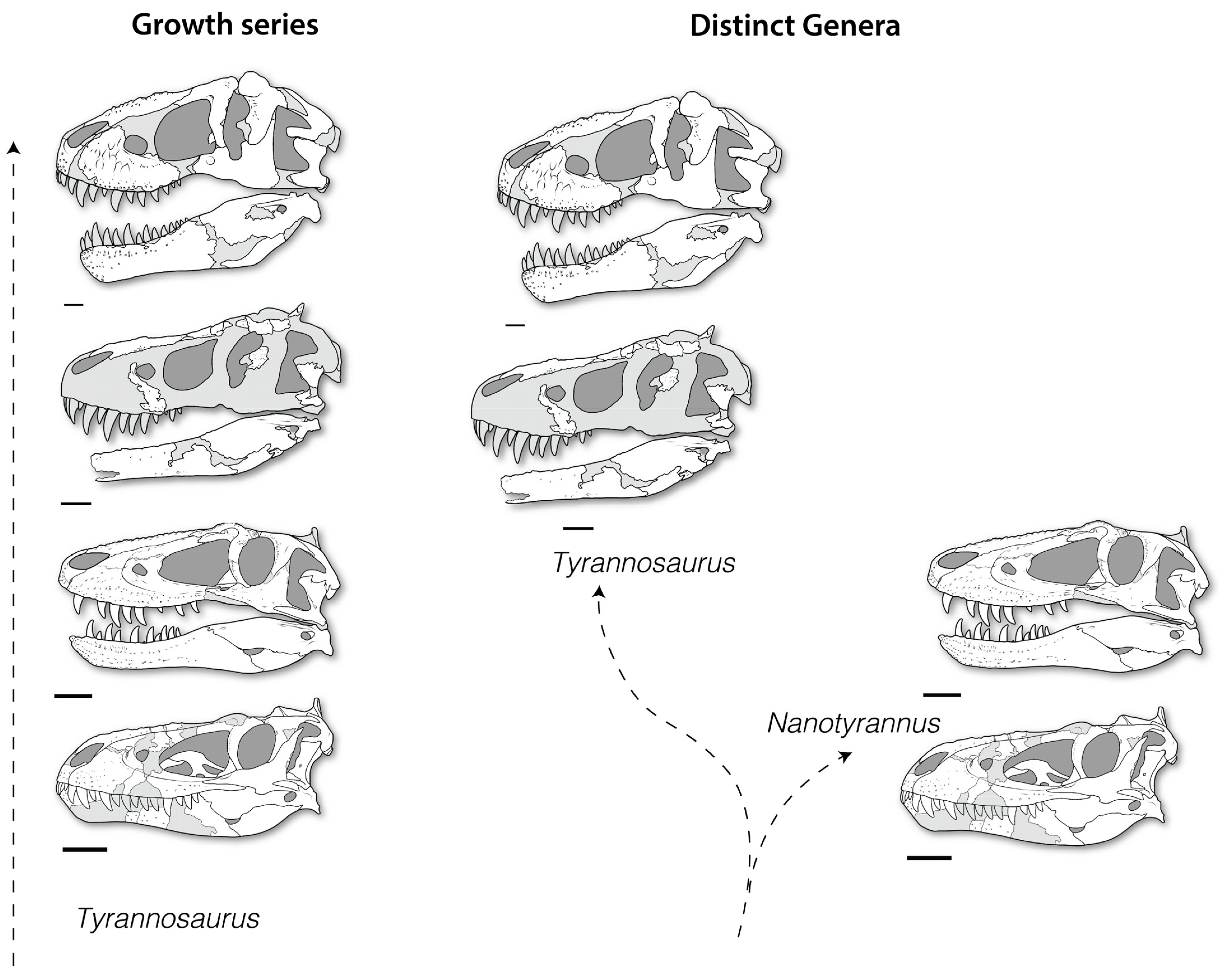
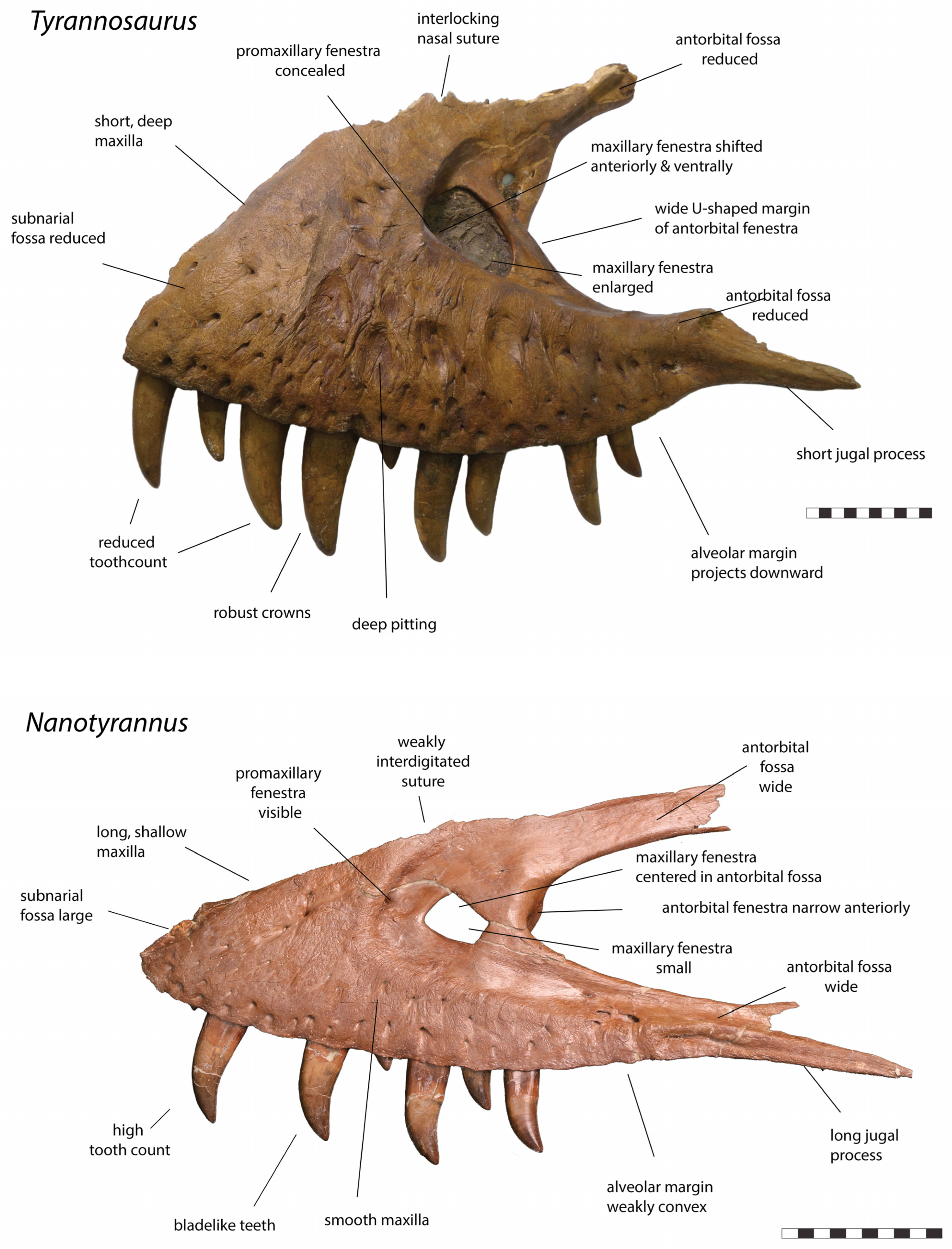
- Morphological differences between Nanotyrannus and Tyrannosaurus and lack of intermediates. The hypothesis that the animals represent a growth series of a single species predicts intermediate forms linking the two. However, dozens of characters in almost every skull bone distinguish Nanotyrannus and Tyrannosaurus, while no intermediate forms are known.
- Developmental patterns seen in other tyrannosaurs. The hypothesized synonymy of Nanotyrannus and Tyrannosaurus suggests that Nanotyrannus will resemble the juveniles of other tyrannosaur species. However, the major changes in morphology proposed for the Nanotyrannus–Tyrannosaurus growth series are not seen in tyrannosaurs such as Tarbosaurus and Gorgosaurus, arguing against ontogeny as an explanation for morphological differences.
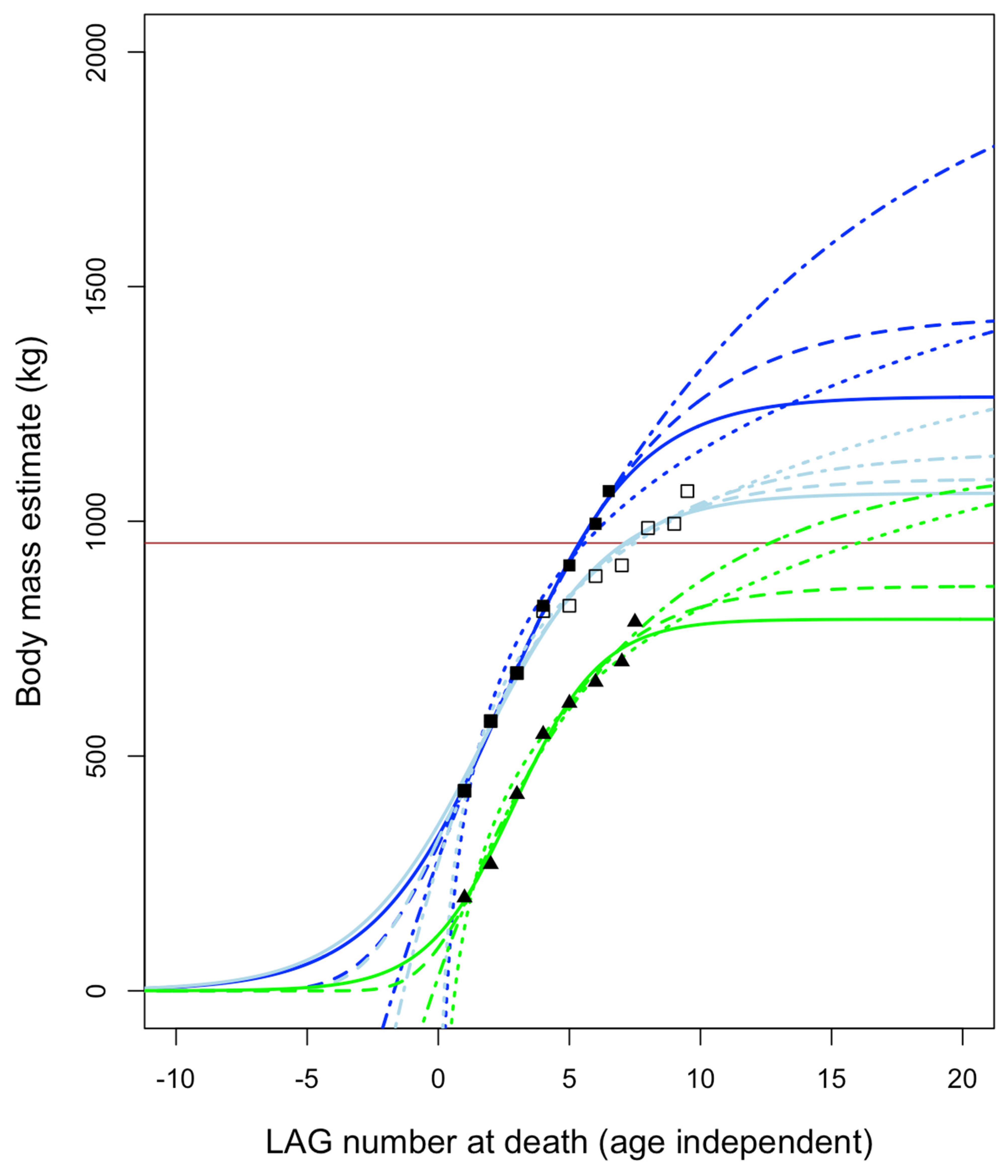
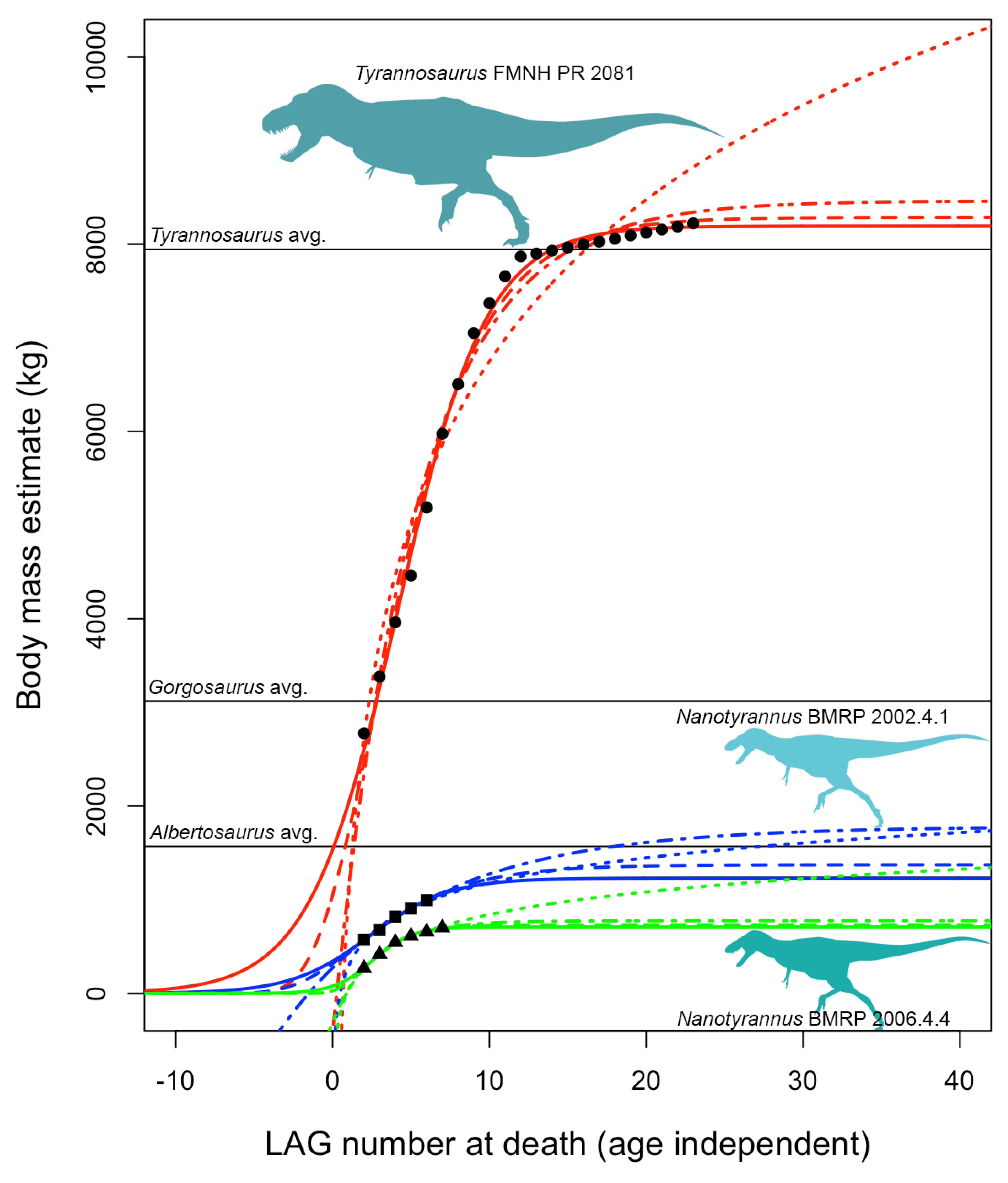
- Adult specimens referable to Nanotyrannus. The hypothesis that Nanotyrannus is a juvenile Tyrannosaurus predicts that all Nanotyrannus will show features associated with immaturity, including immature bone texture and lack of fusion between skeletal elements. Histology is also predicted to show Nanotyrannus specimens rapidly growing to reach adult sizes on the order of ~8000 kg. Nanotyrannus specimens instead show patterns of bone fusion and bone texture consistent with maturity, are slow growing compared to maximal growth rates of T. rex, and show a pattern of decelerating growth in their final years of life, consistent with fairly mature animals. Reconstructed growth curves predict small adult body mass (<2000 kg, more likely ~900–1500 kg).
- Juvenile tyrannosaurs showing diagnostic features of T. rex. The hypothesized Nanotyrannus–Tyrannosaurus growth series requires that small specimens showing features of Tyrannosaurus do not exist. A juvenile skull, slightly larger than Nanotyrannus, and a juvenile frontal, smaller than the holotype, represent juveniles of Tyrannosaurus.
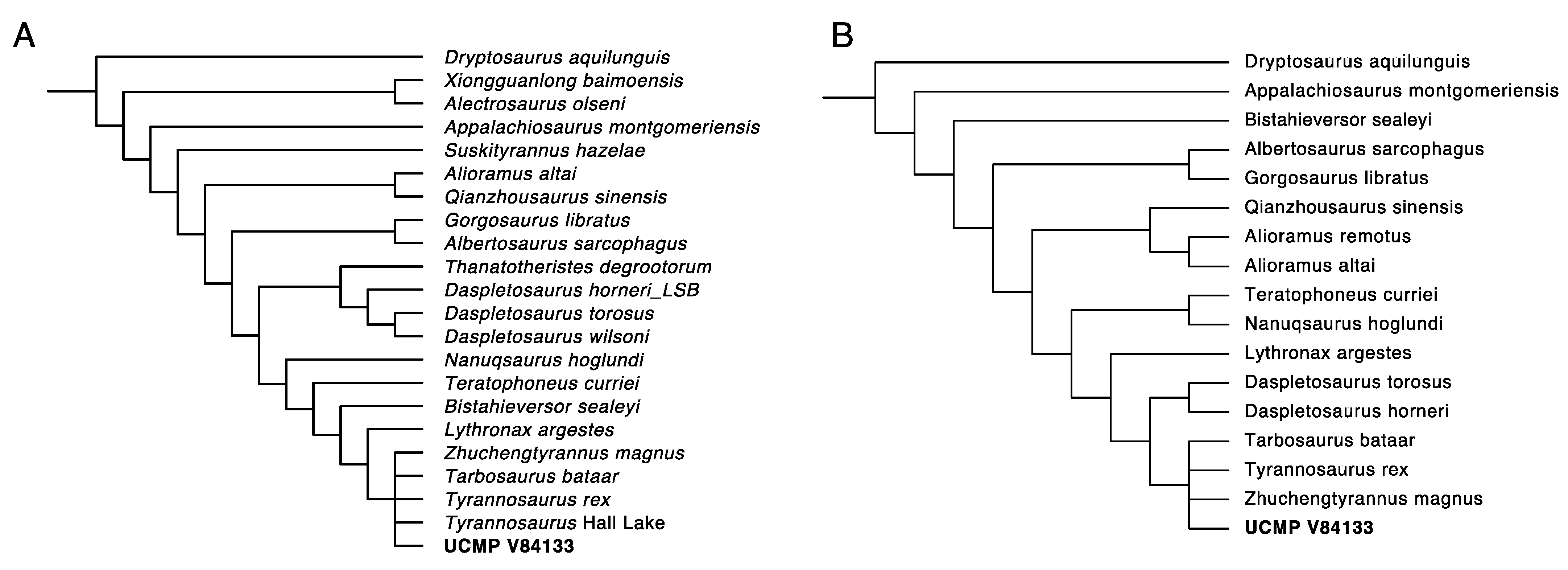
- Phylogenetic analysis of Nanotyrannus. If Nanotyrannus is a juvenile Tyrannosaurus, then phylogenetic analysis with ontogenetically variable characters removed should cause Nanotyrannus to cluster with Tyrannosaurus. The morphology of Nanotyrannus instead places it outside of Tyrannosaurinae and Tyrannosauridae, even when restricting analysis to characters known to be stable during ontogeny.
2. Materials and Methods
2.1. Clustering Analysis
2.2. Histology and Growth Curves
2.3. Phylogenetic Analysis
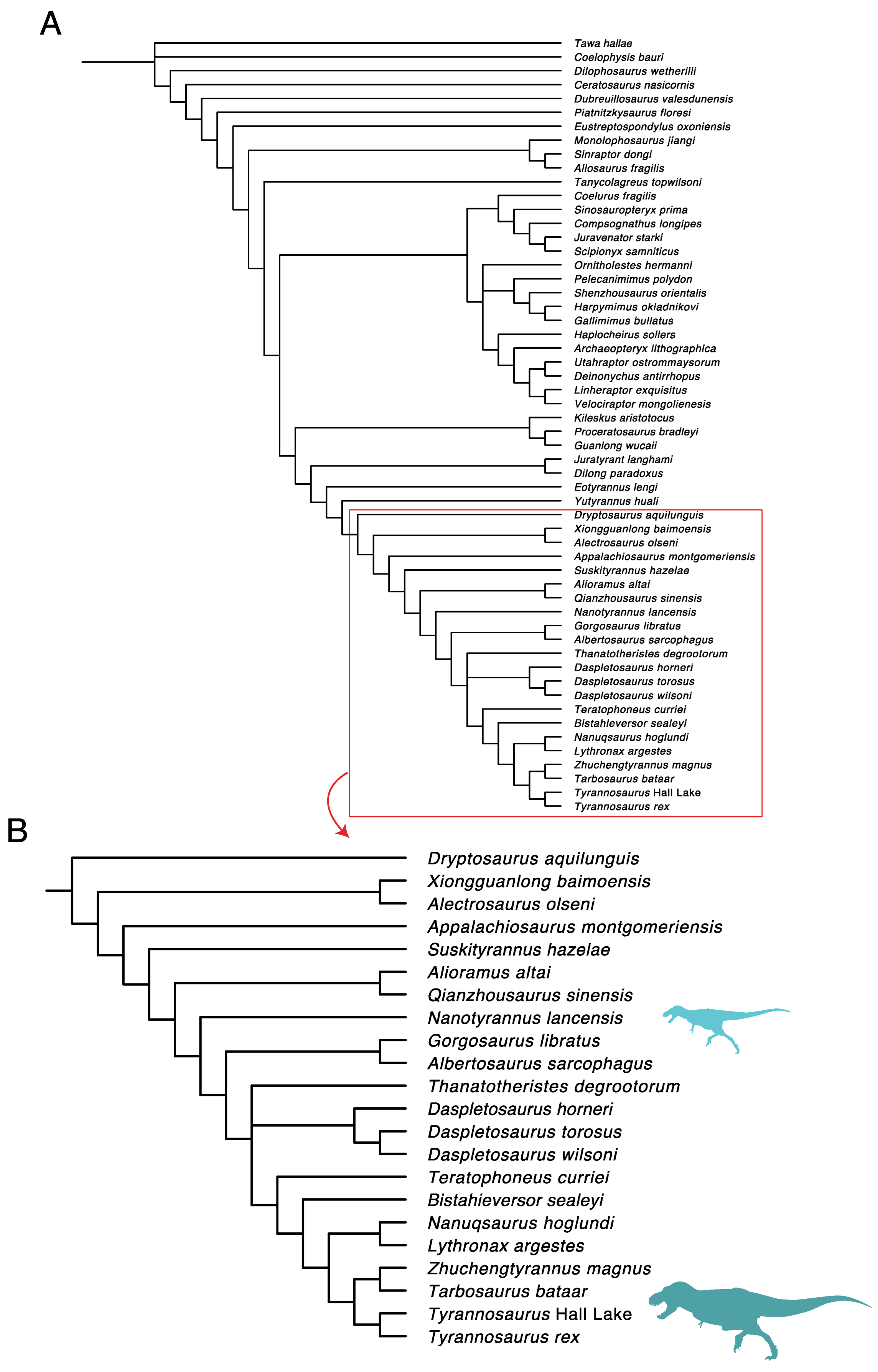
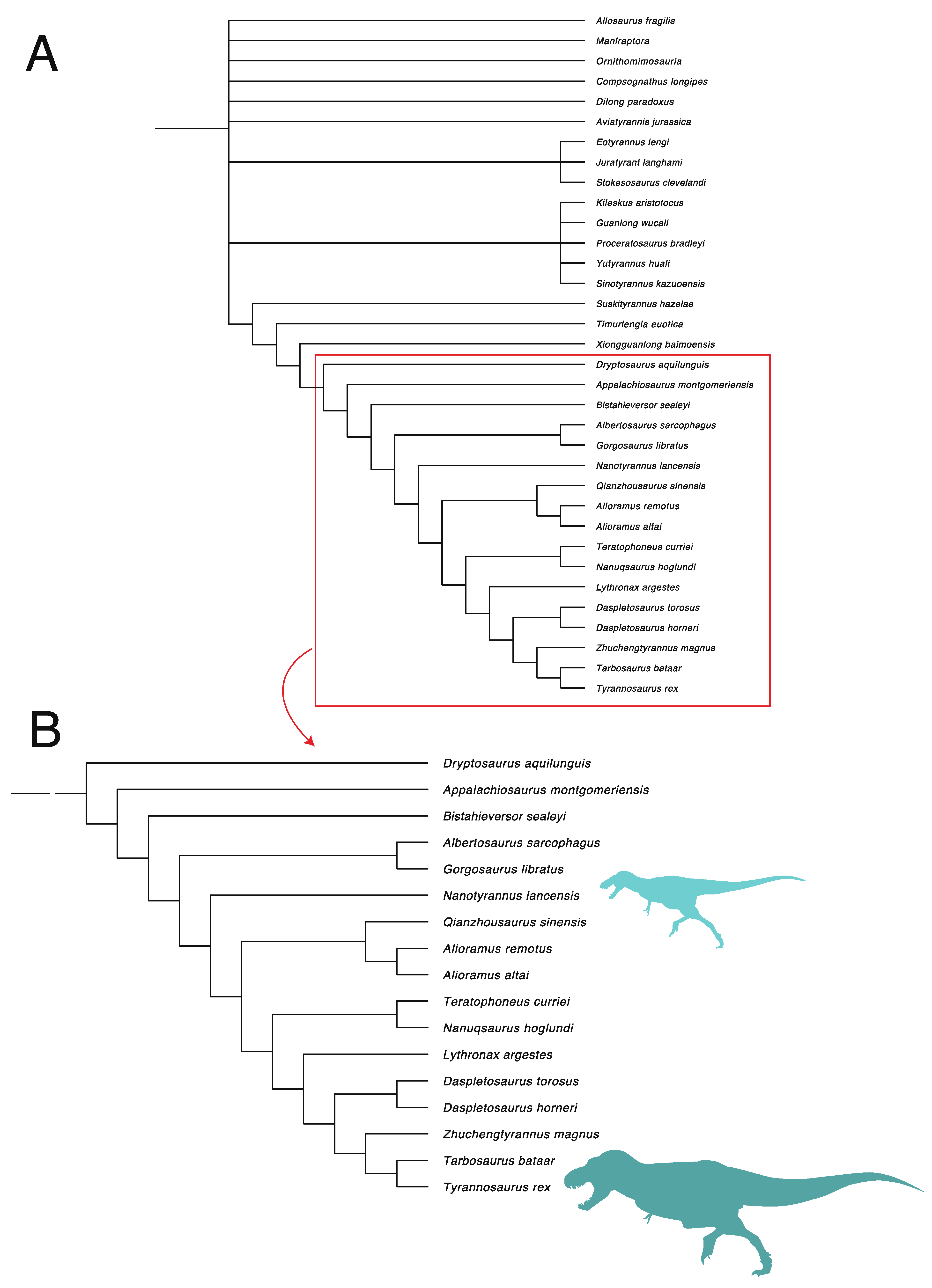
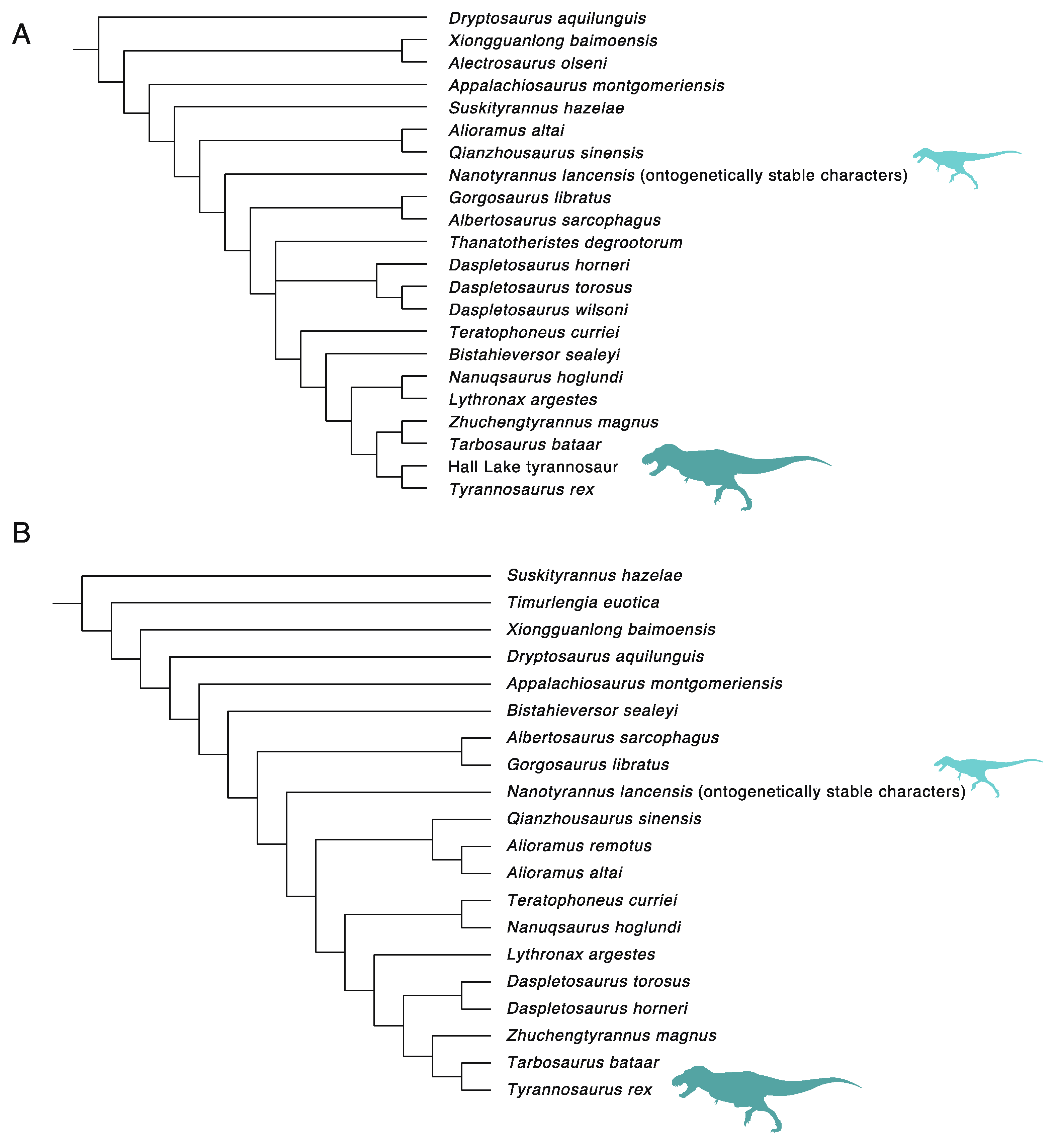
3. Results
3.1. Diversity Patterns of Tyrannosaurs and Apex Predators

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Age | Formation | Locality | Describer |
|---|---|---|---|---|
| Dryptosauridae | ||||
| Dryptosaurus aquilunguis | Late Maastrichtian | Hornerstown Formation | New Jersey, USA | [94] |
| Appalachiosaurus montgomeriensis (?) | Early Campanian | Demopolis Formation | Alabama, USA | [95] |
| Alioraminae | ||||
| Alioramus altai | Maastrichtian | Nemegt Formation | Mongolia | [96] |
| Alioramus remotus | Maastrichtian | Nemegt Formation | Mongolia | [97] |
| Qianzhousaurus sinensis | Maastrichtian | Nanxiong Formation | China | [98] |
| Albertosaurinae | ||||
| Albertosaurus sarcophagus | Early Maastrichtian | Horseshoe Canyon Formation | Alberta, Canada | [6] |
| Gorgosaurus libratus | Late Campanian | Dinosaur Park Formation | Alberta, Canada | [66] |
| Tyrannosaurinae | ||||
| Thanatotheristes degrootorum | Early Campanian | Foremost Formation | Alberta, Canada | [57] |
| Daspletosaurus torosus | Middle Campanian | Oldman Formation | Alberta, Canada | [29] |
| Daspletosaurus wilsoni | Middle Campanian | Judith River Fm. | Montana, USA | [60] |
| Daspletosaurus sp. | Late Campanian | Dinosaur Park Formation | Alberta, Canada | [39] |
| Daspletosaurus horneri | Late Campanian | Two Medicine Fm. | Montana, USA | [61] |
| Dynamoterror dynastes | Early Campanian | Menefee Fm. | New Mexico, USA | [99] |
| Labocania anomala | Late Campanian | La Bocana Roja Formation | Baja California, Mexico | [62] |
| Lythronax argestes | Early Campanian | Wahweap Formation | Utah, USA | [56] |
| Teratophoneus curriei | Late Campanian | Kaiparowits Formation | Utah, USA | [58] |
| Bistahieversor sealeyi | Late Campanian | Kirtland Formation | New Mexico, USA | [59] |
| Nanuqsaurus hoglundi | Middle Maastrichtian | Prince Creek Fm. | Alaska, USA | [68] |
| Shanshanosaurus huoyanshanensis | Late Cretaceous | Subashi Formation | Xinjiang, China | [100] |
| Tarbosaurus bataar | Maastrichtian | Nemegt Formation | Mongolia | [101] |
| Zhuchengtyrannus magnus | Campanian | Hongtuya Formation, Wangshi Group | Shandong, China | [102] |
| Tyrannosaurus rex | Late Maastrichtian | Hell Creek, Lance, Frenchman, Scollard, North Horn Fms | Alberta and Saskatchewan, Canada; Montana, Wyoming, North Dakota, South Dakota, Colorado, Utah, USA | [6] |
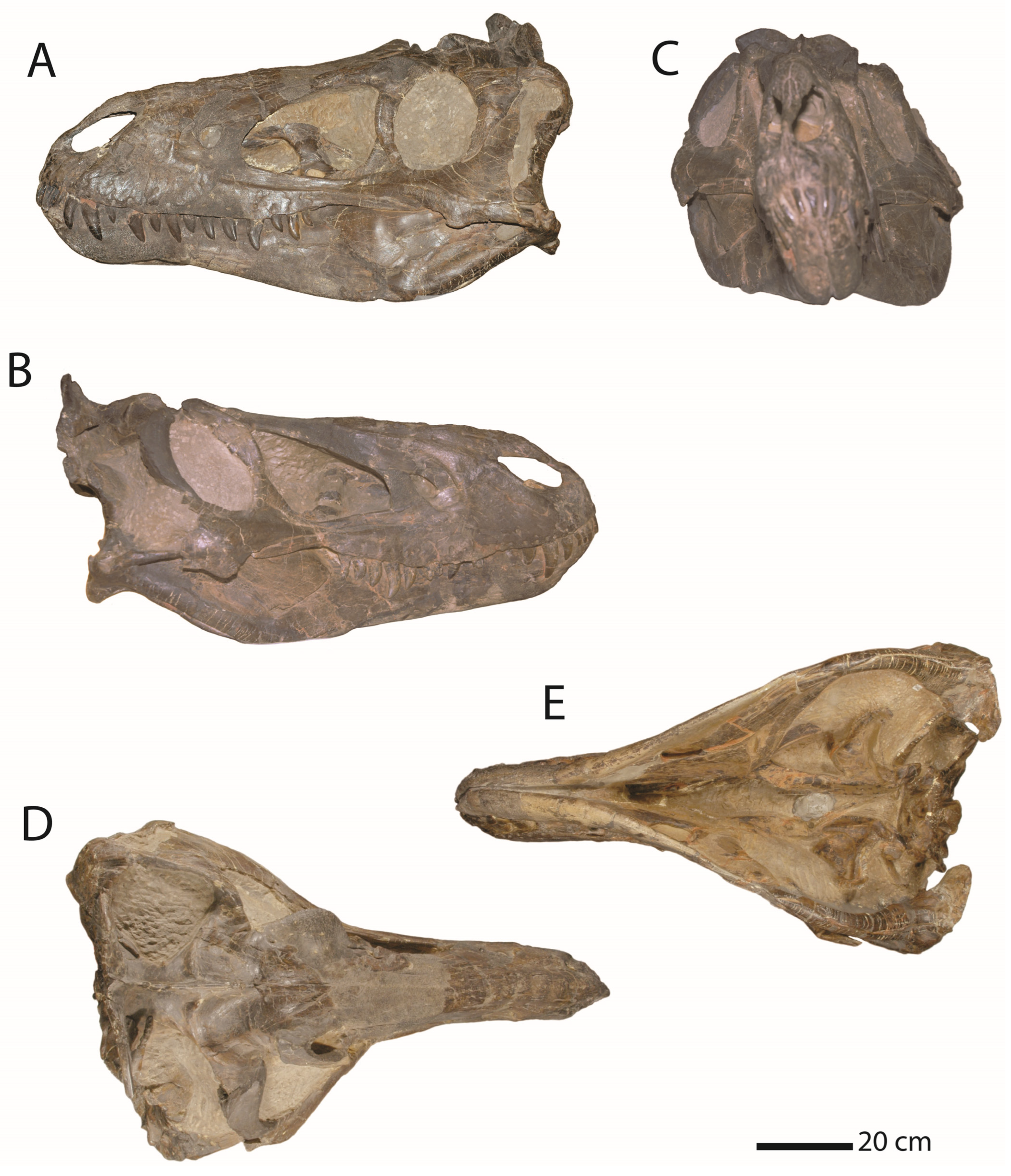
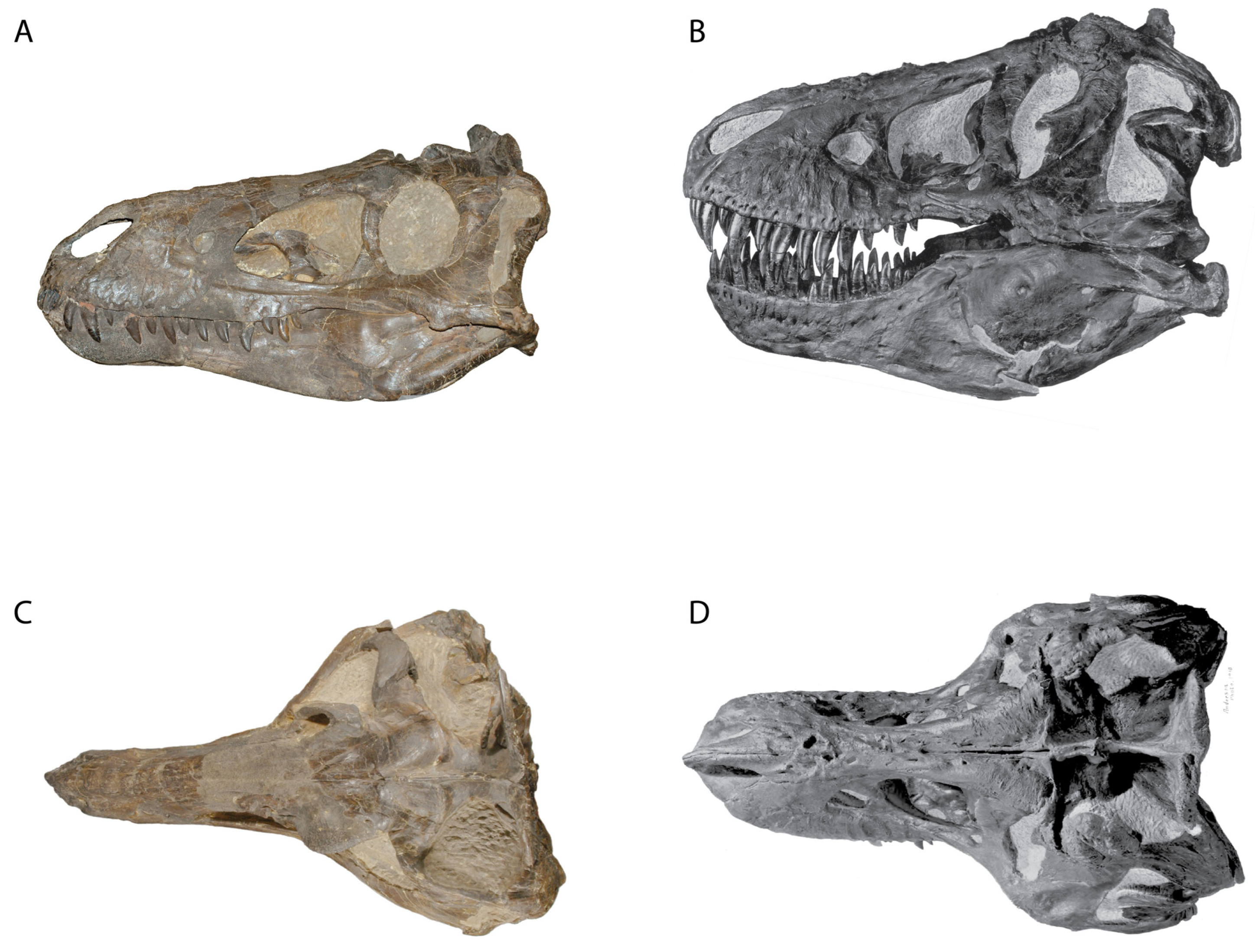
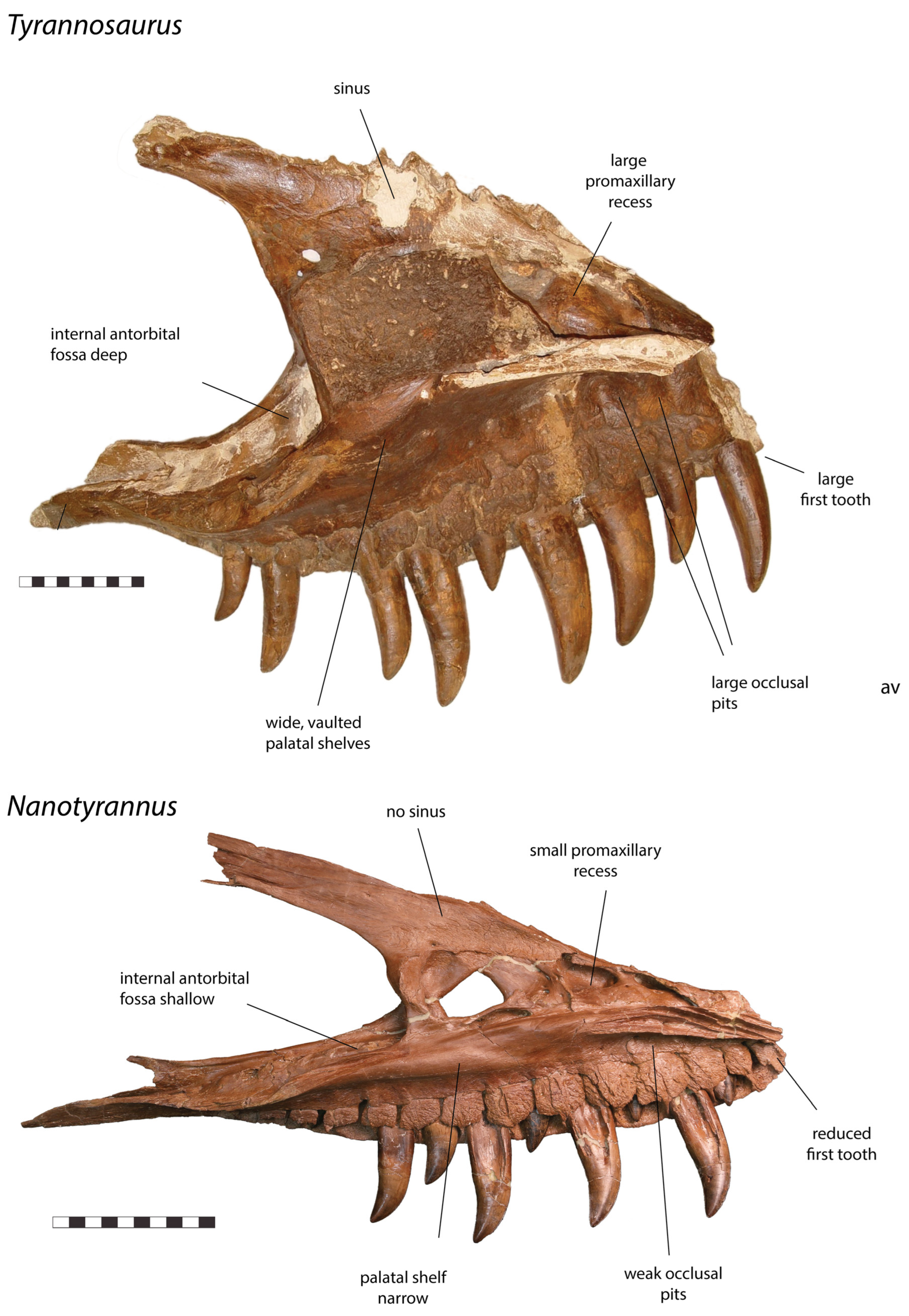
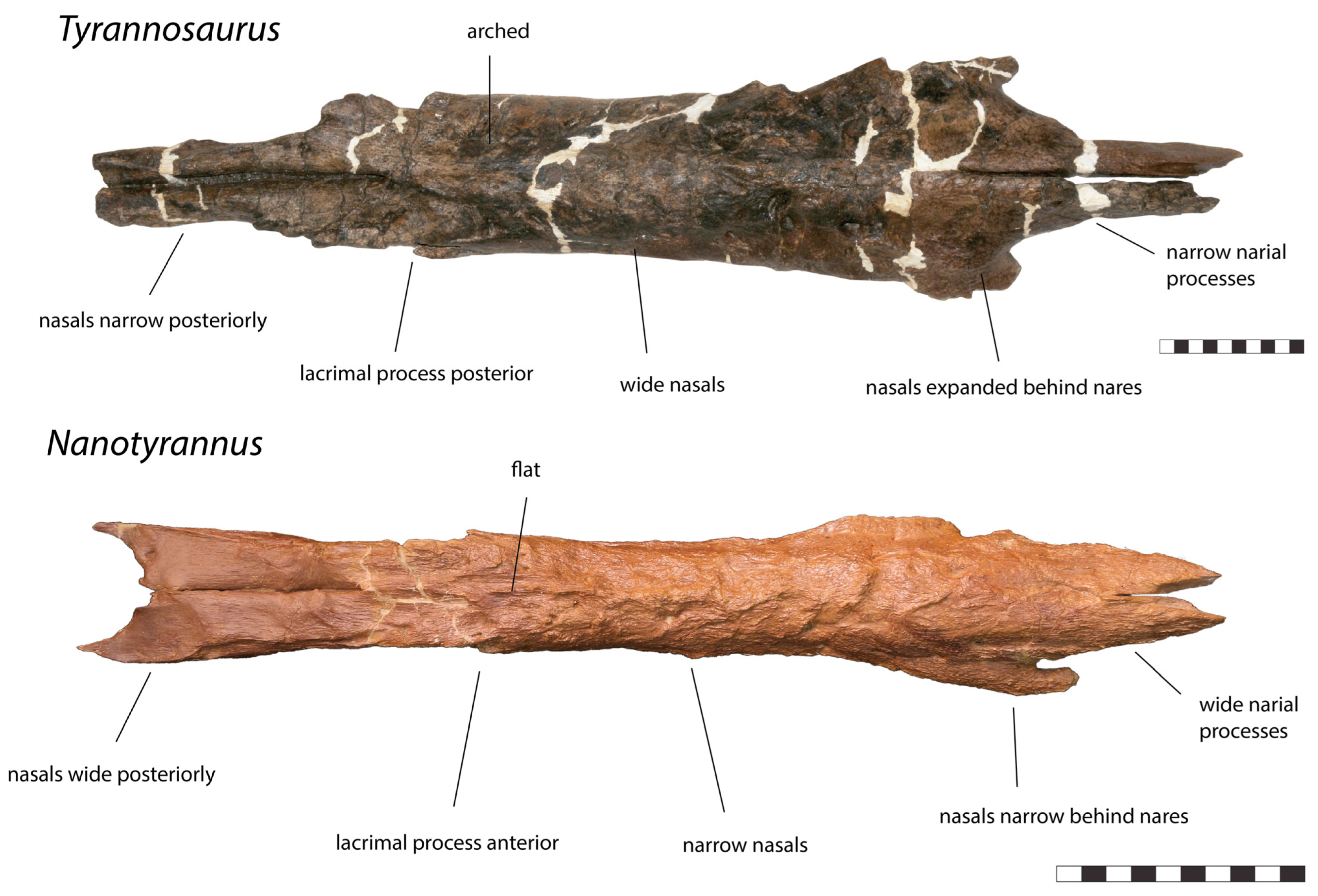
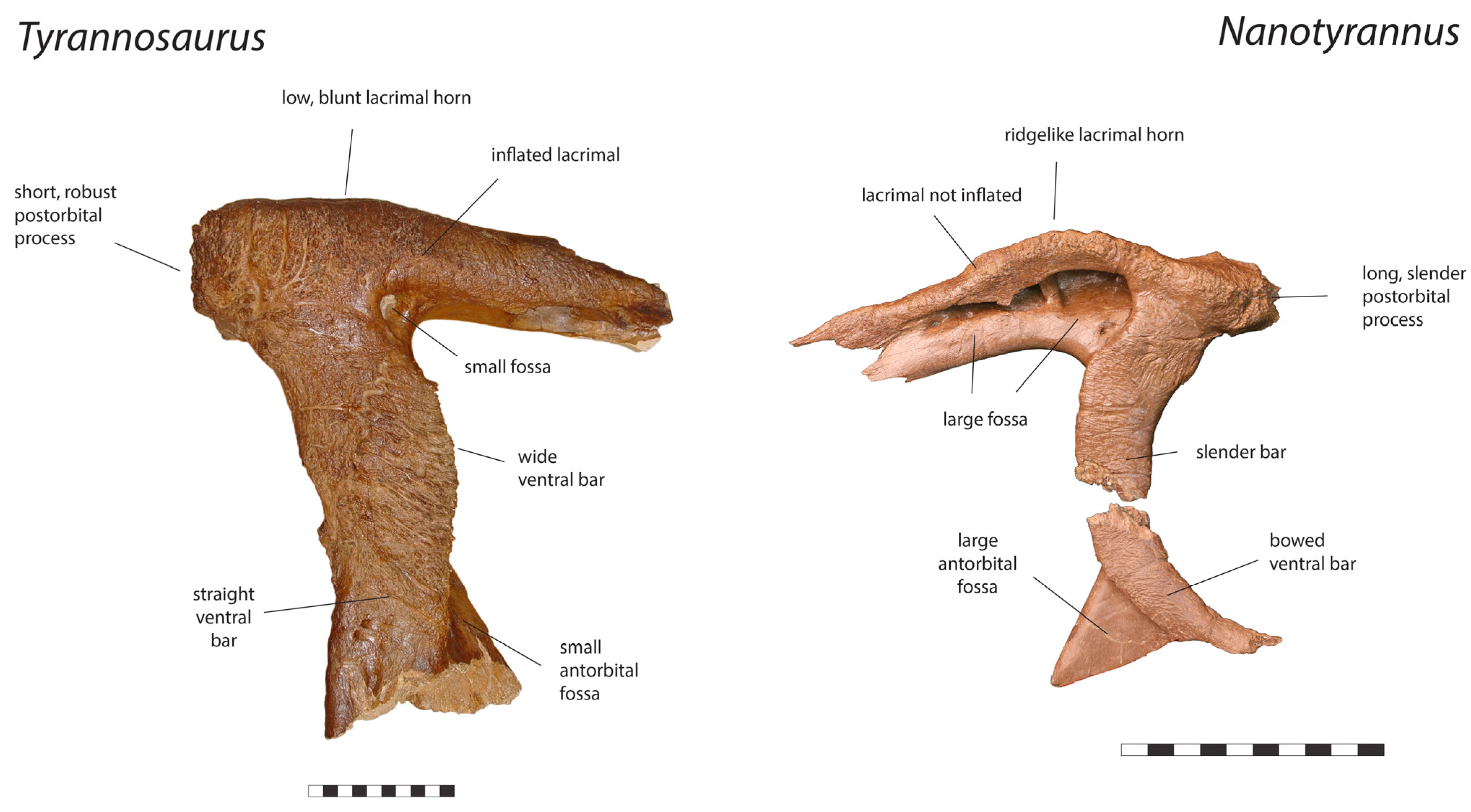
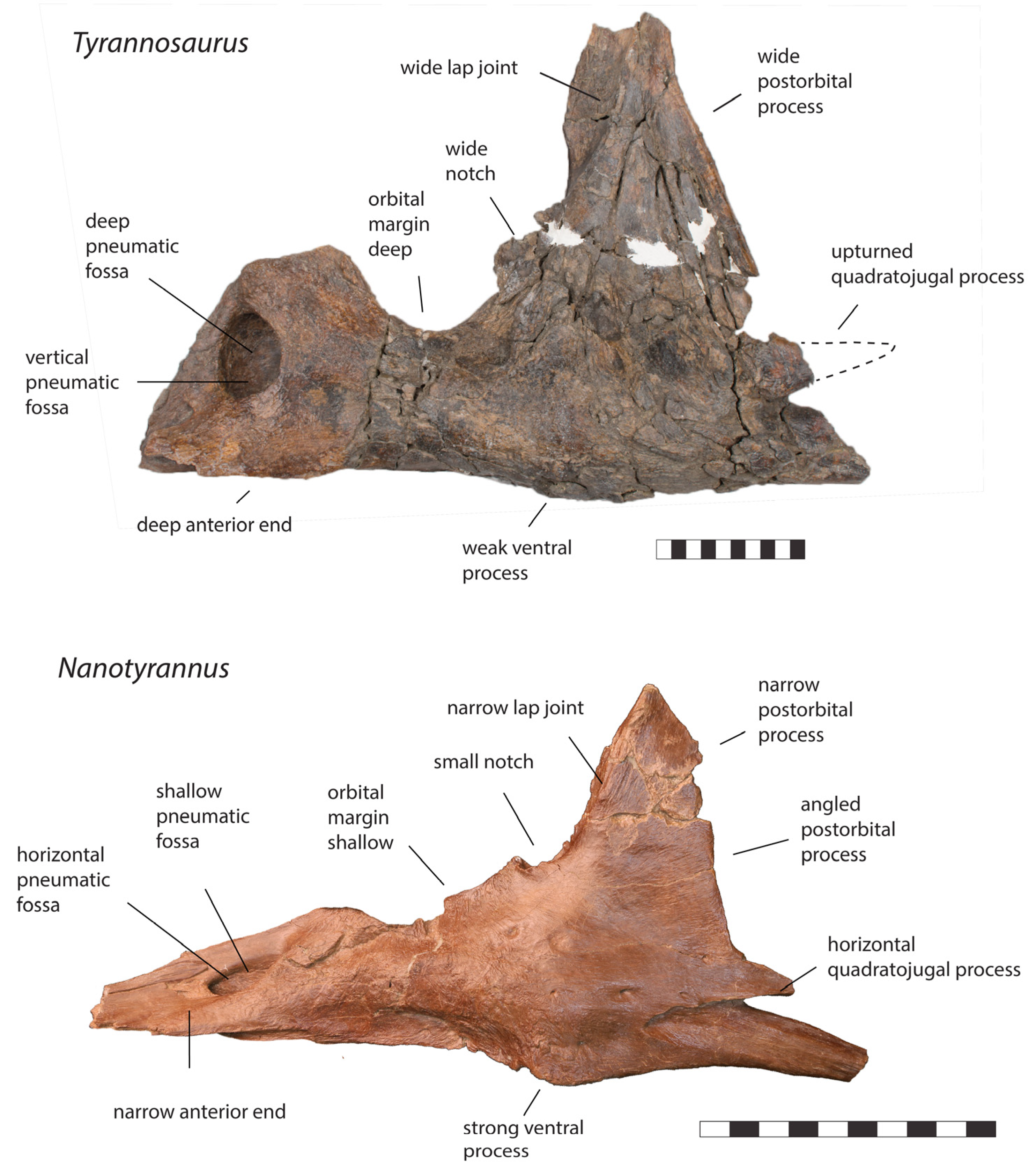
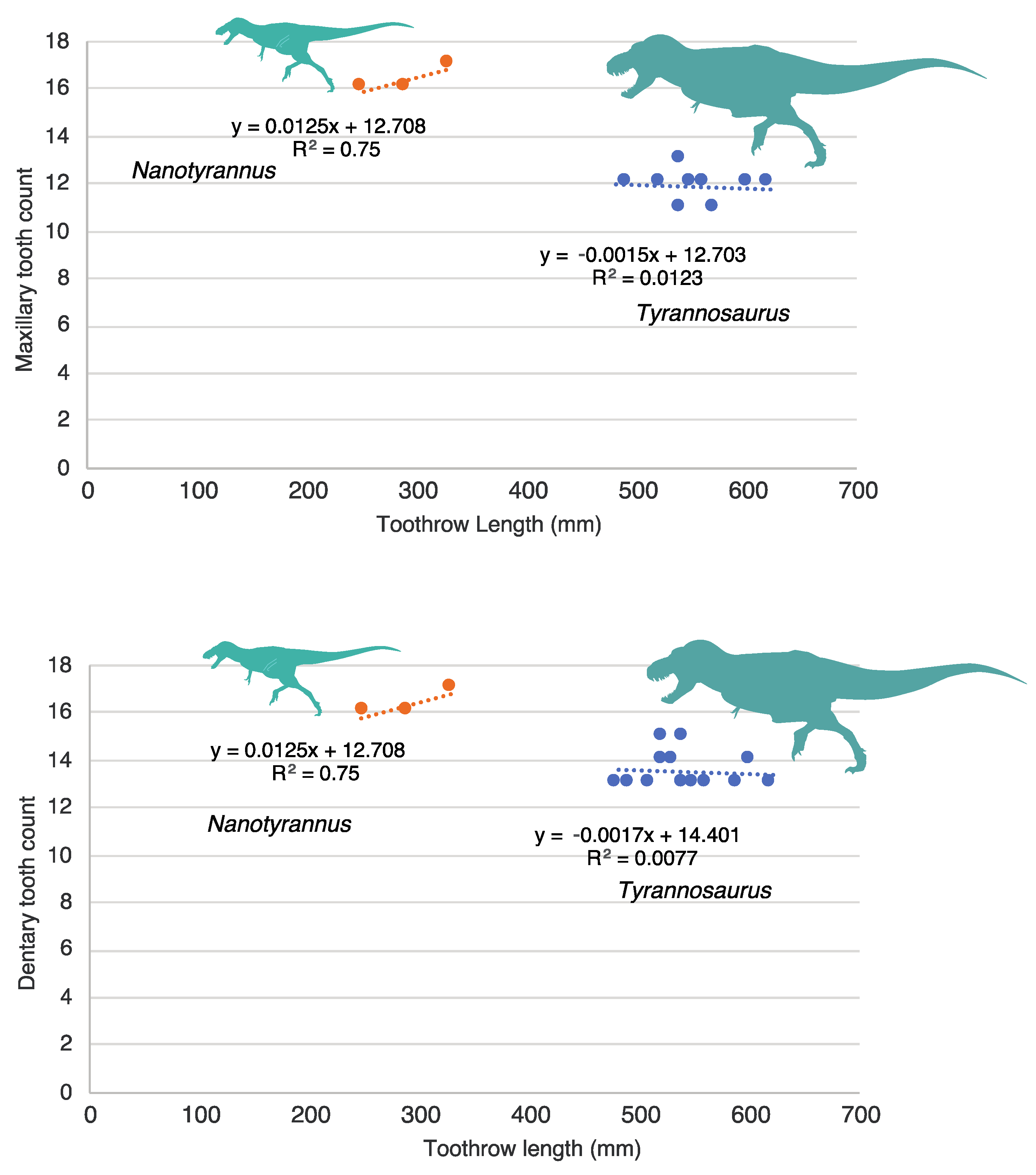
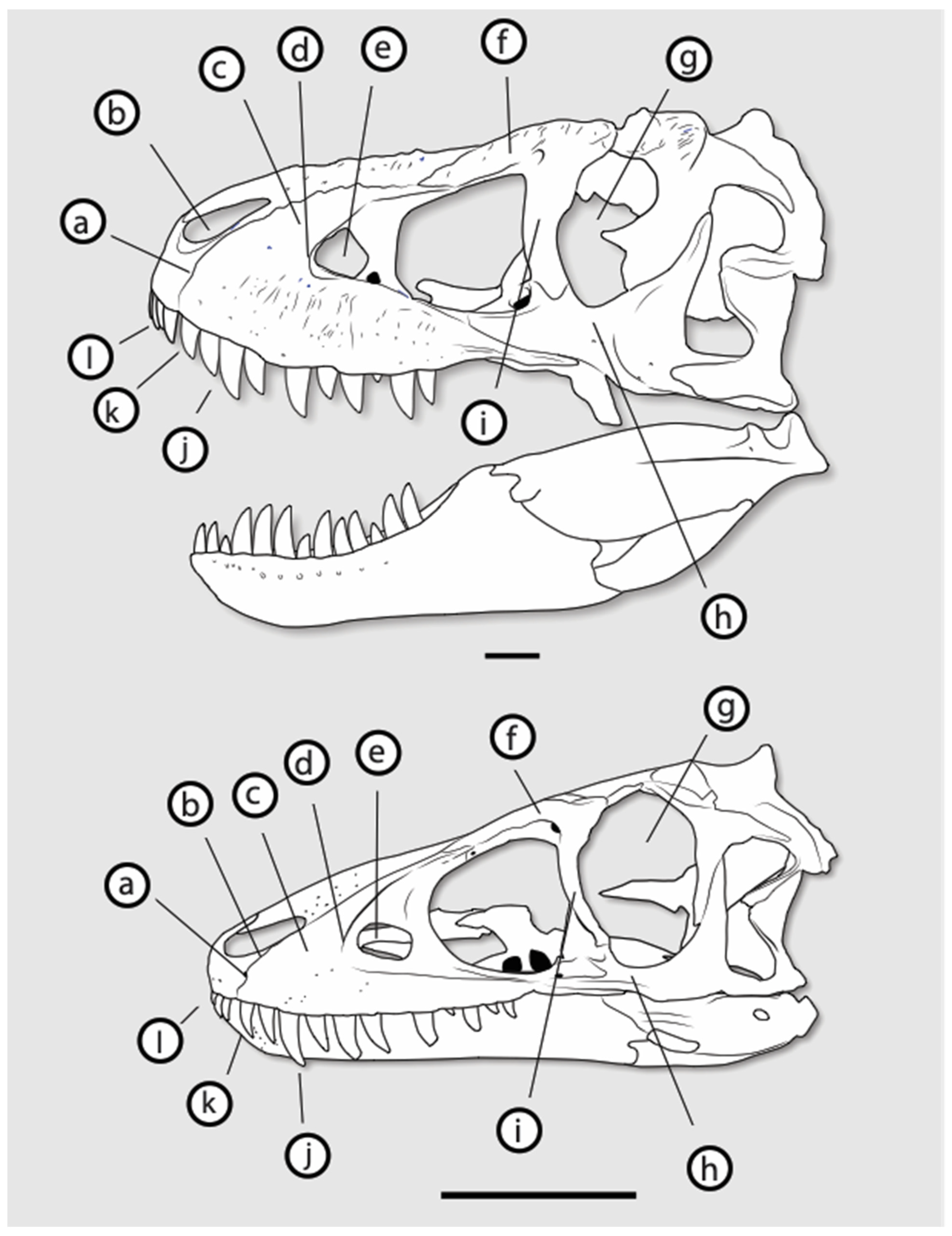
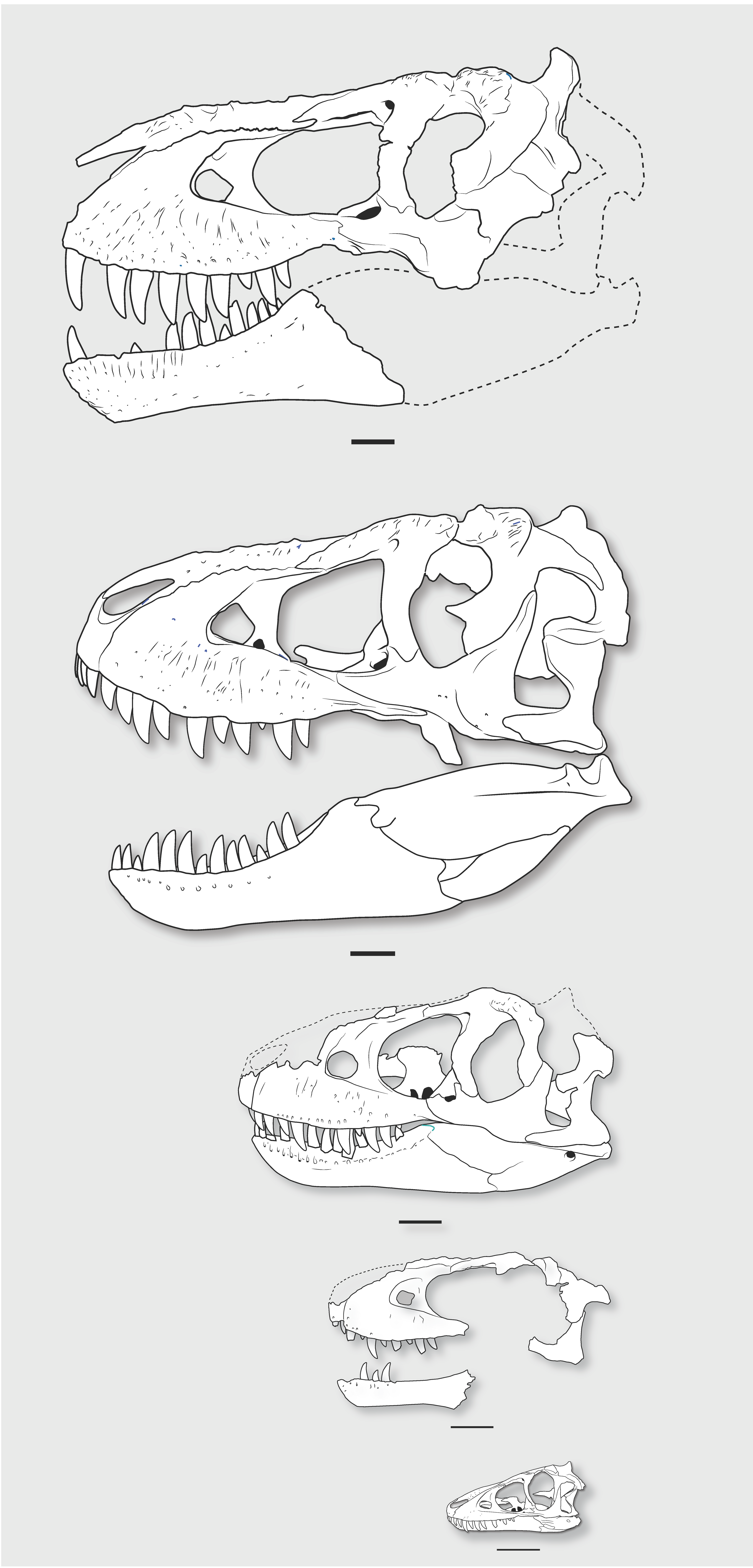
3.2. Morphology of Nanotyrannus and Tyrannosaurus
3.2.1. Characters Differentiating Nanotyrannus and Tyrannosaurus
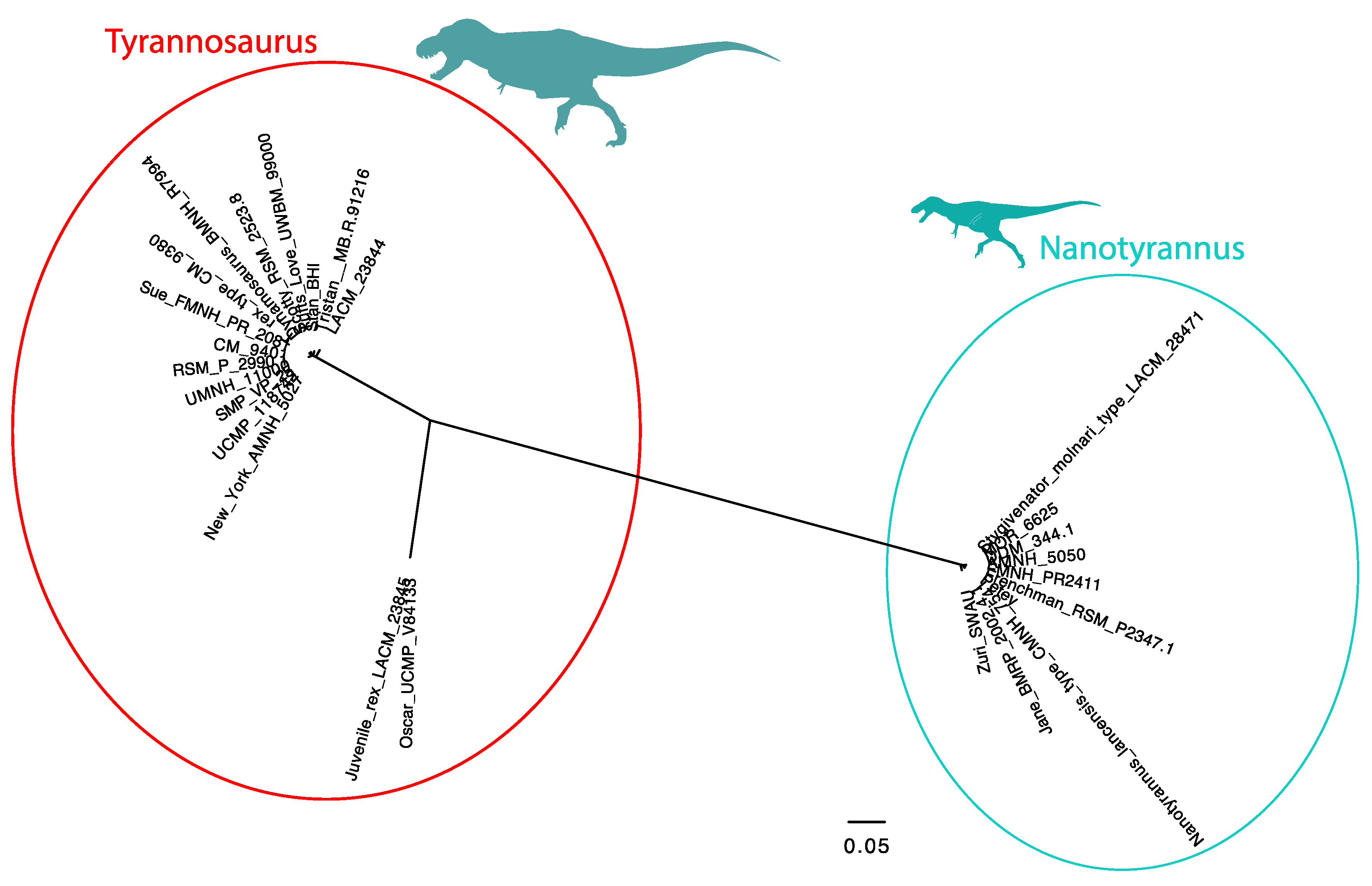
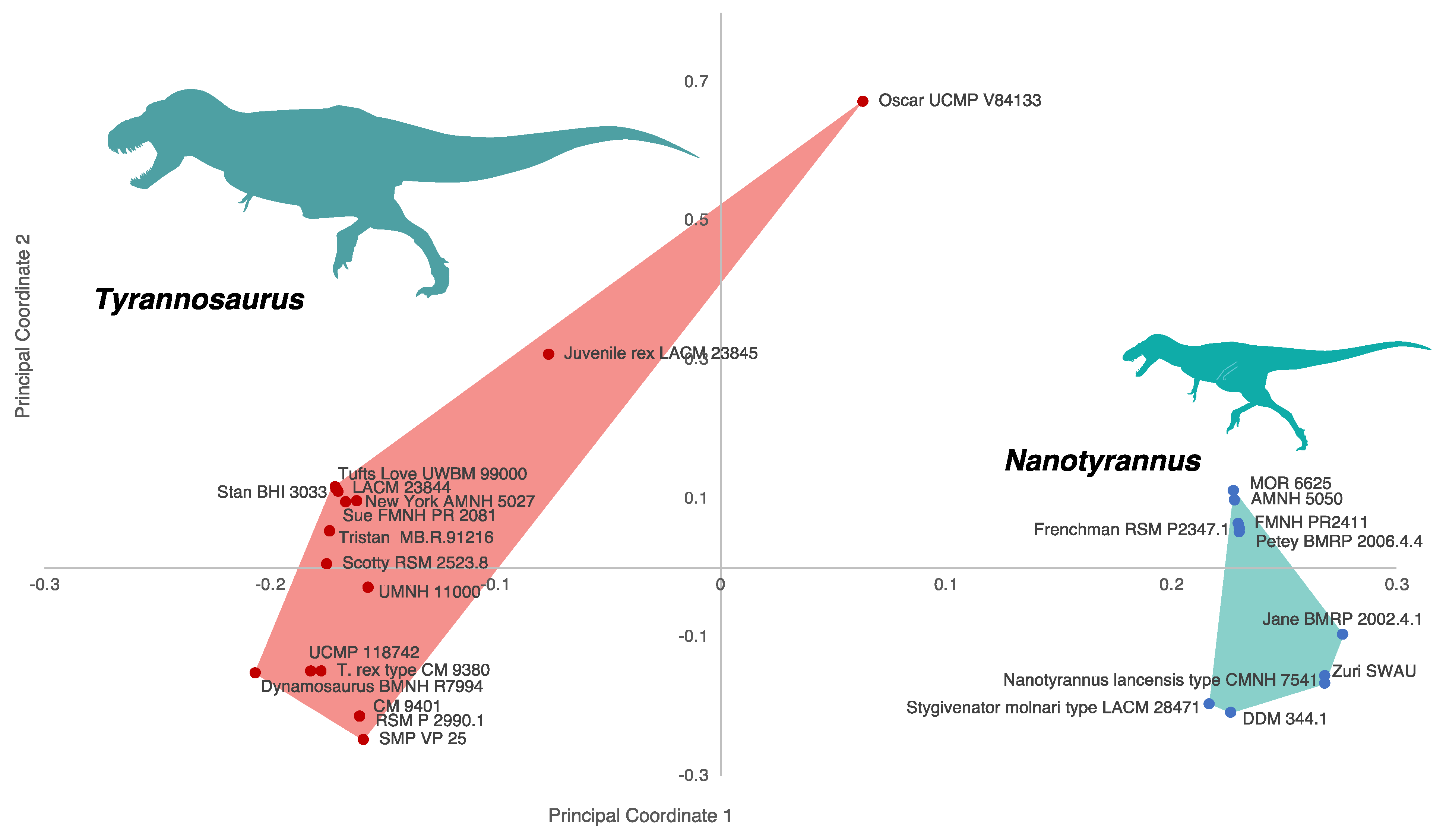
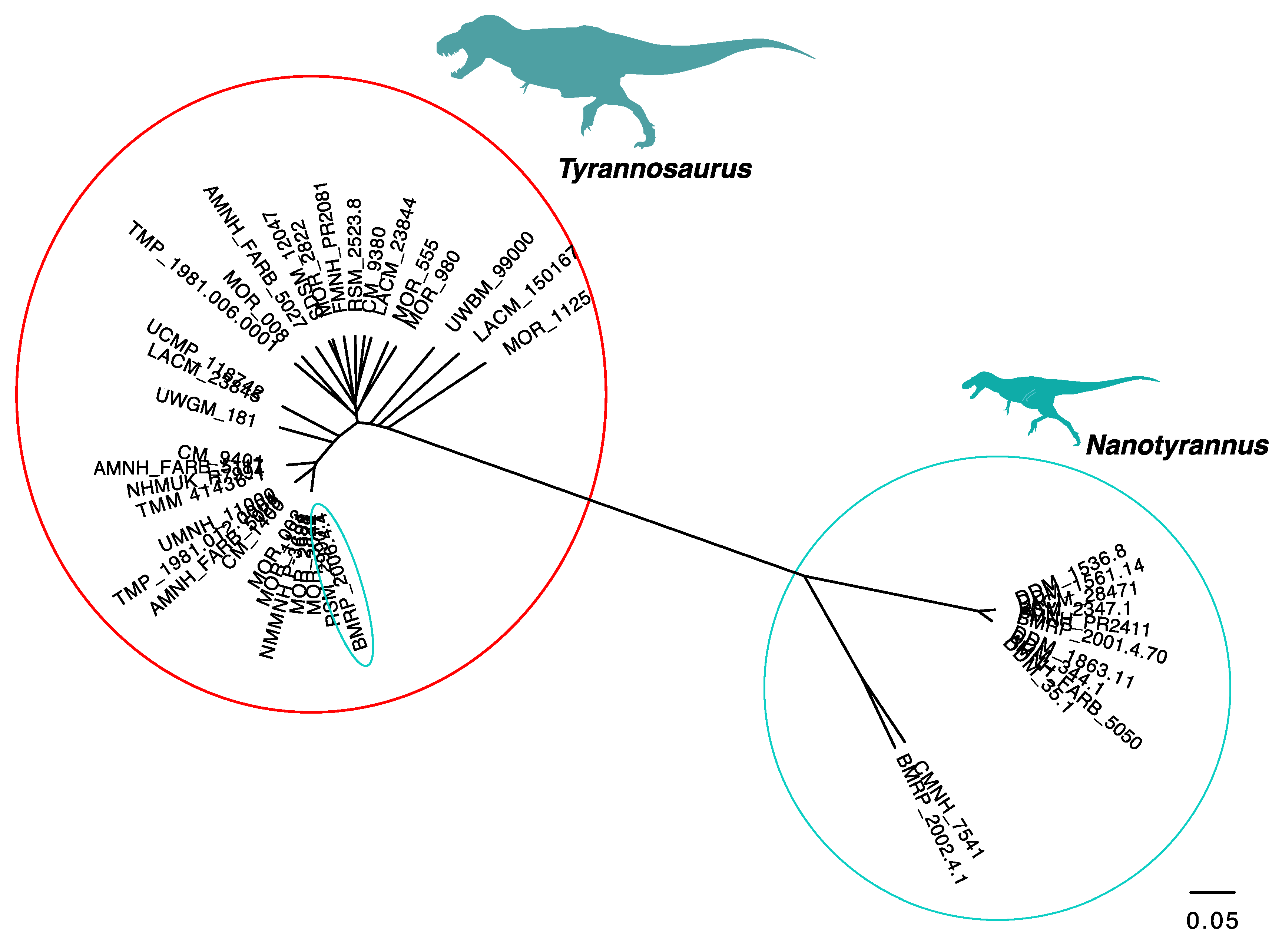
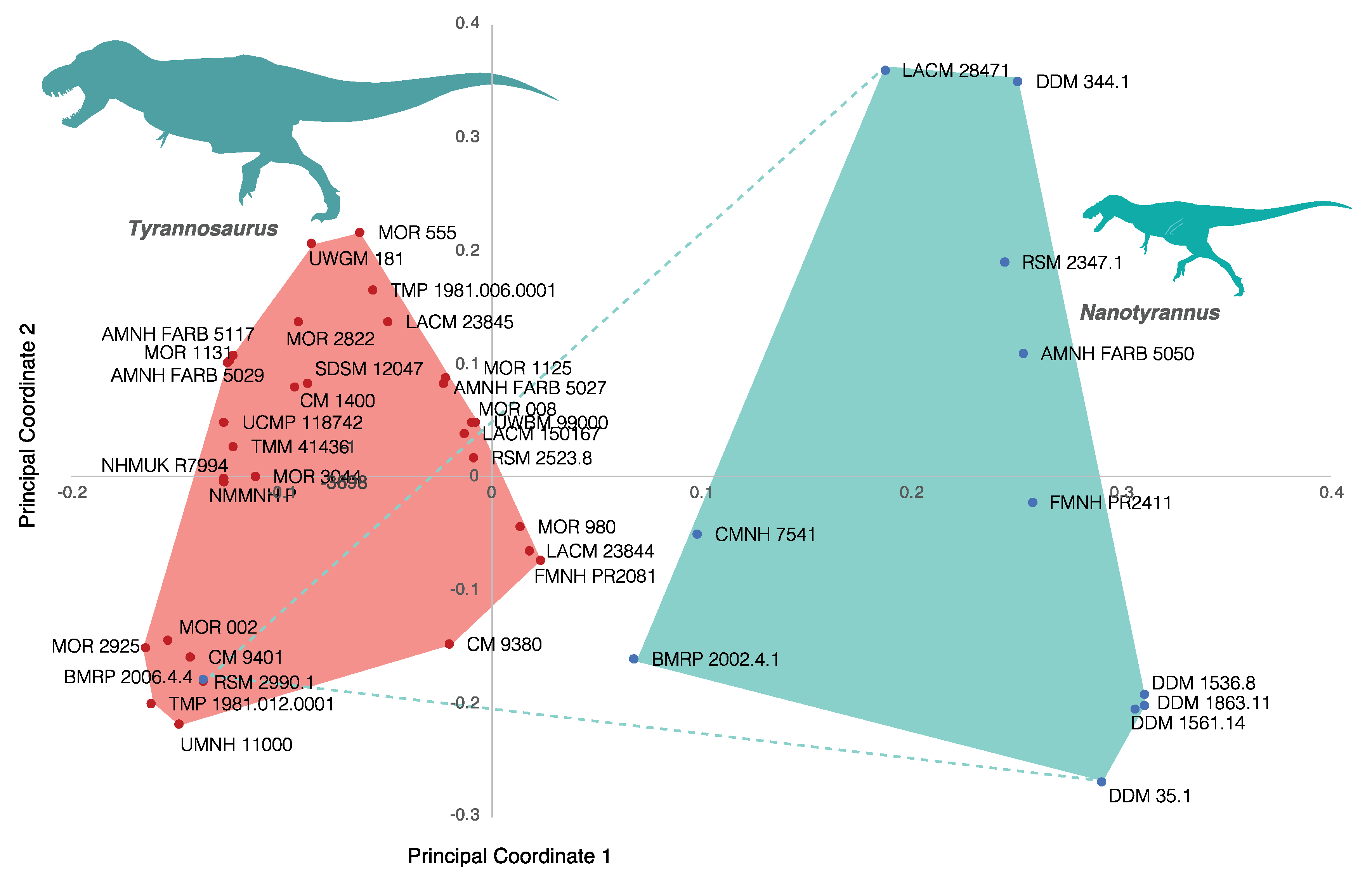
3.2.2. Character Distribution and Clustering Analysis
3.3. Nanotyrannus Morphology Inconsistent with Predicted Morphology of Juvenile Tyrannosaurinae
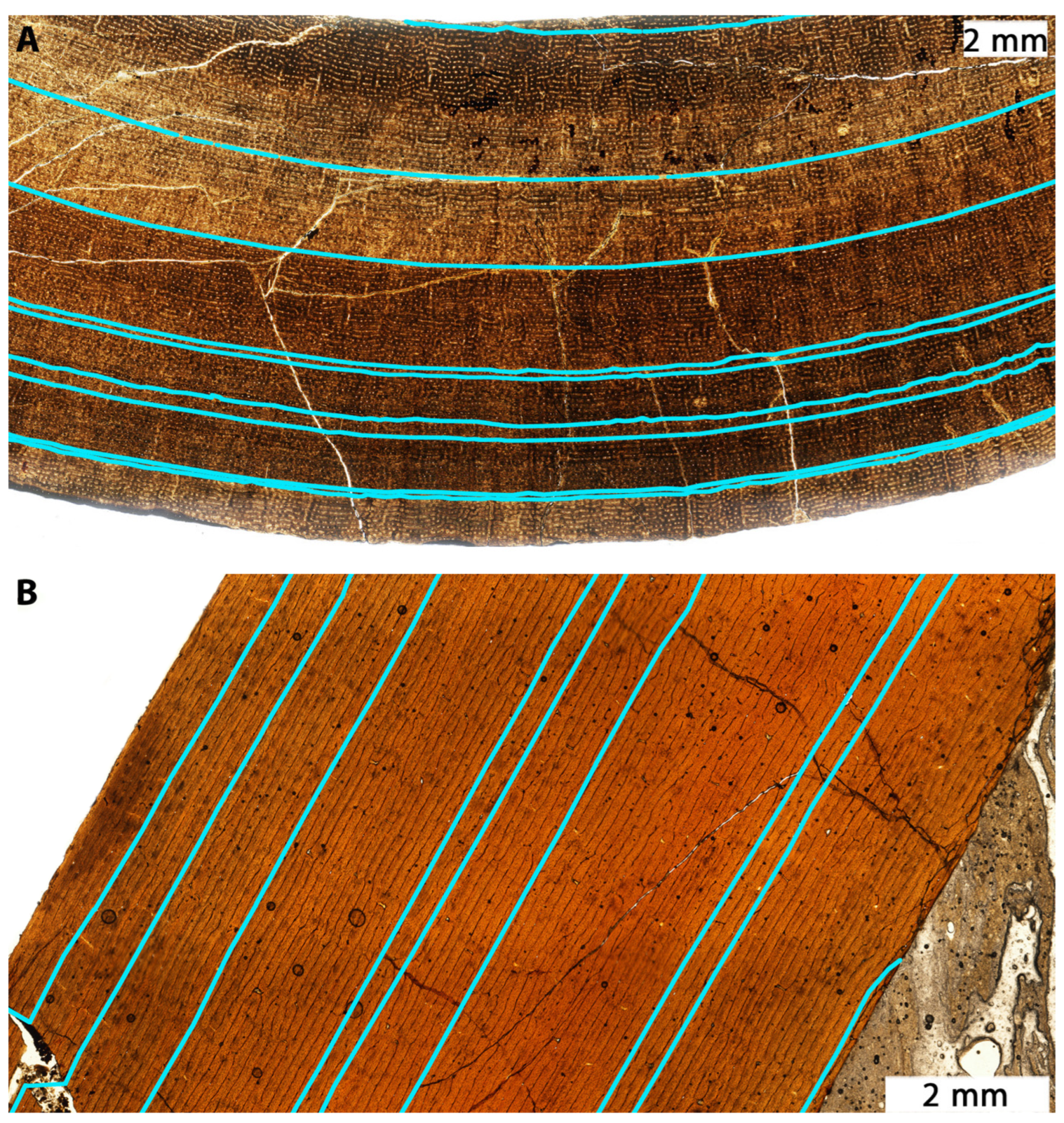
3.4. Histology Supports Existence of Mature Nanotyrannus
3.4.1. Use of Histology to Test the Two Hypotheses
- (i)
- Patterns of skeletal fusion;
- (ii)
- Bone surface texture;
- (iii)
- Presence/absence of an external fundamental system (EFS);
- (iv)
- Patterns of annual growth rates, either in terms of measures of bone deposition or kilograms of mass;
- (v)
- Predicted adult mass, extrapolated from growth curves.
3.4.2. Skeletal Fusion
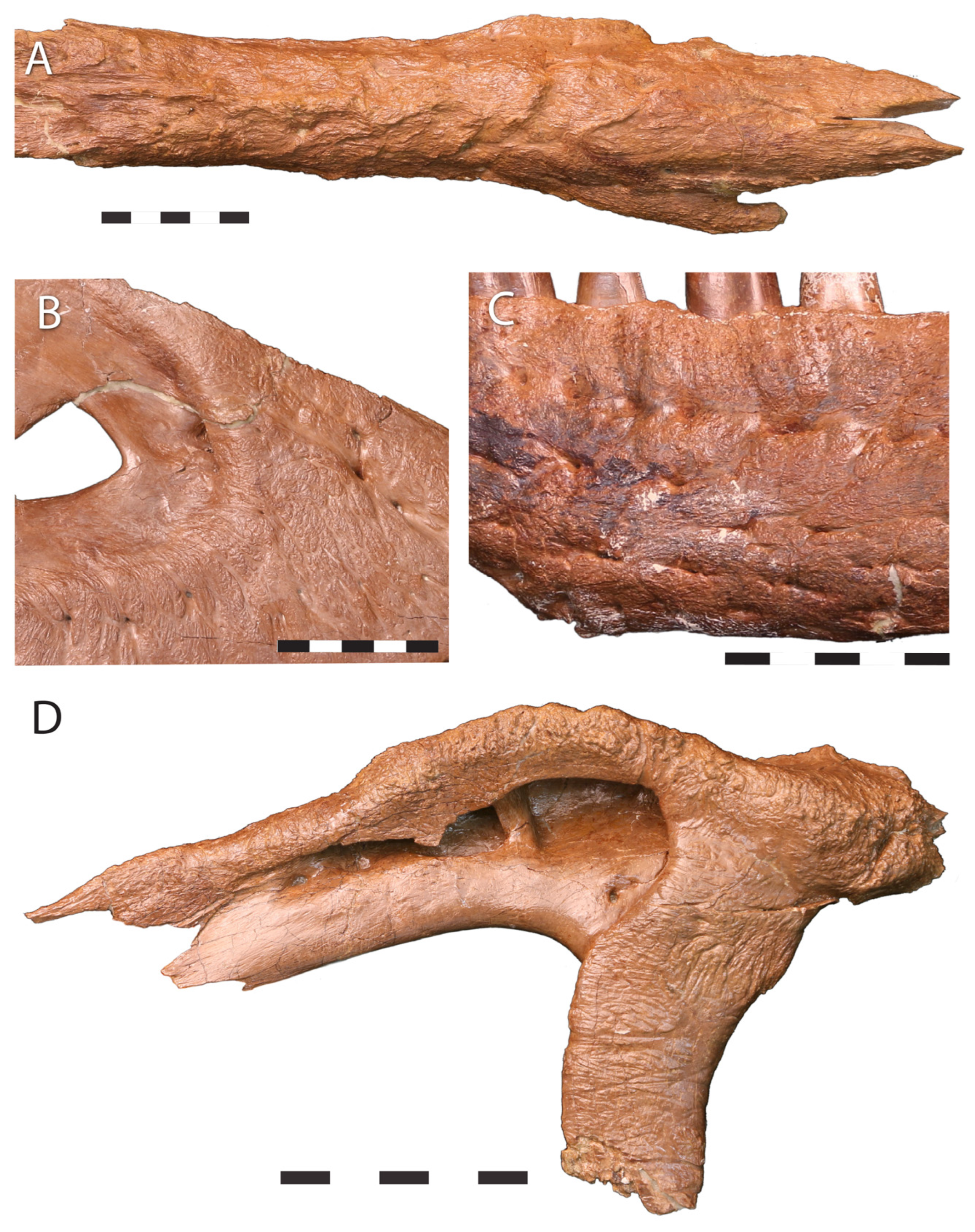
3.4.3. Surface Texture
3.4.4. External Fundamental System
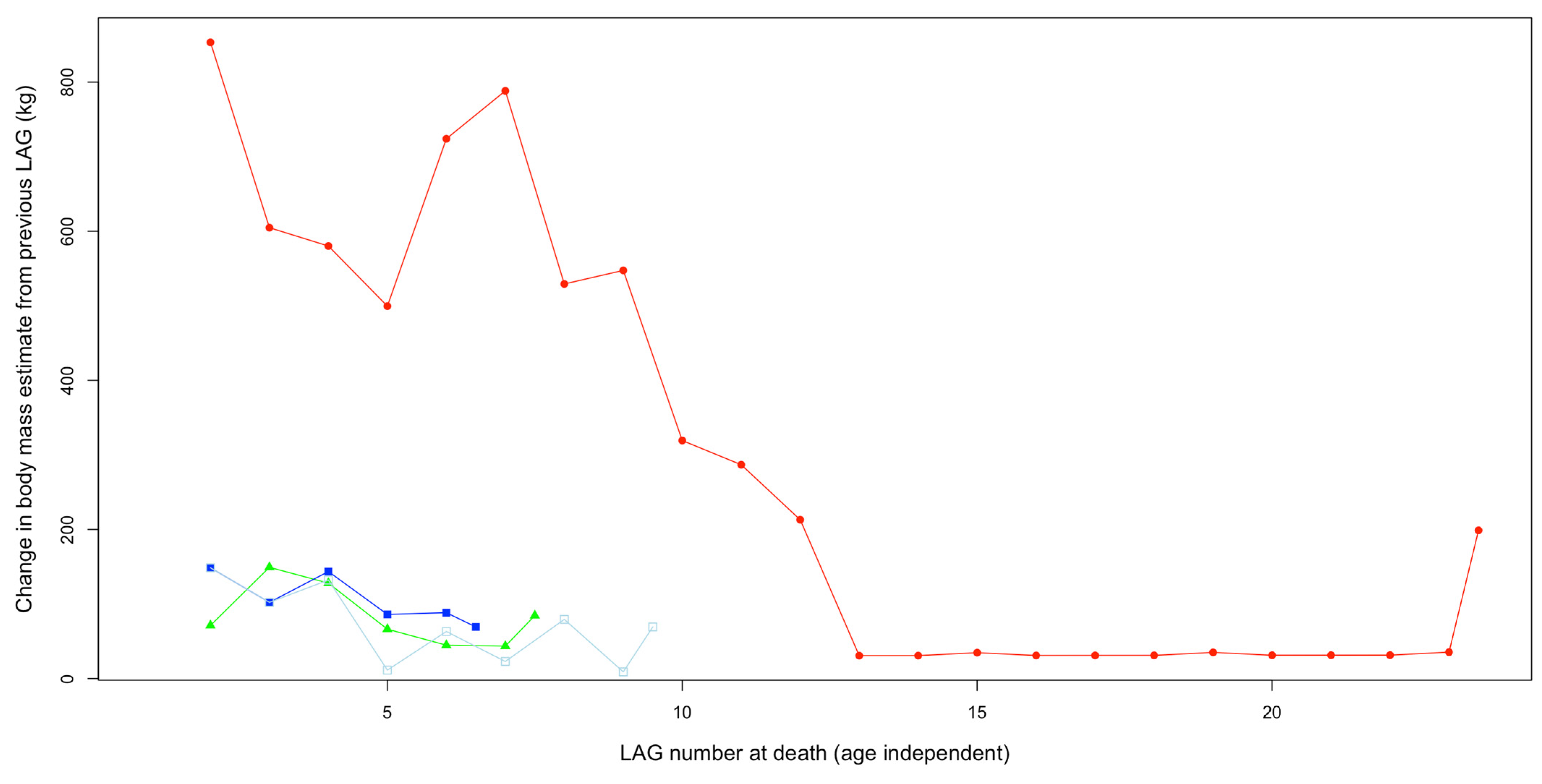
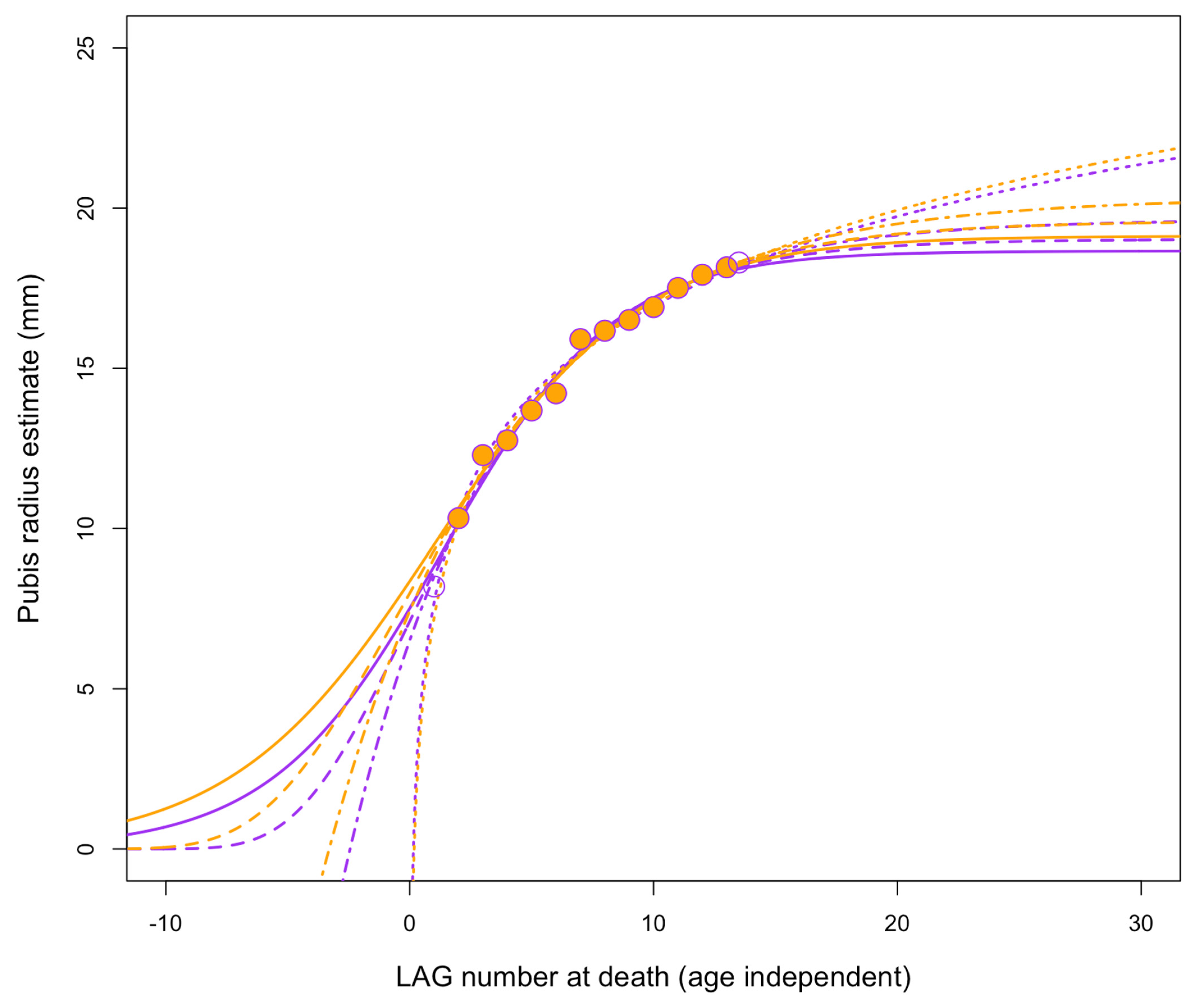
3.4.5. Growth Rates
3.4.6. Growth Trends
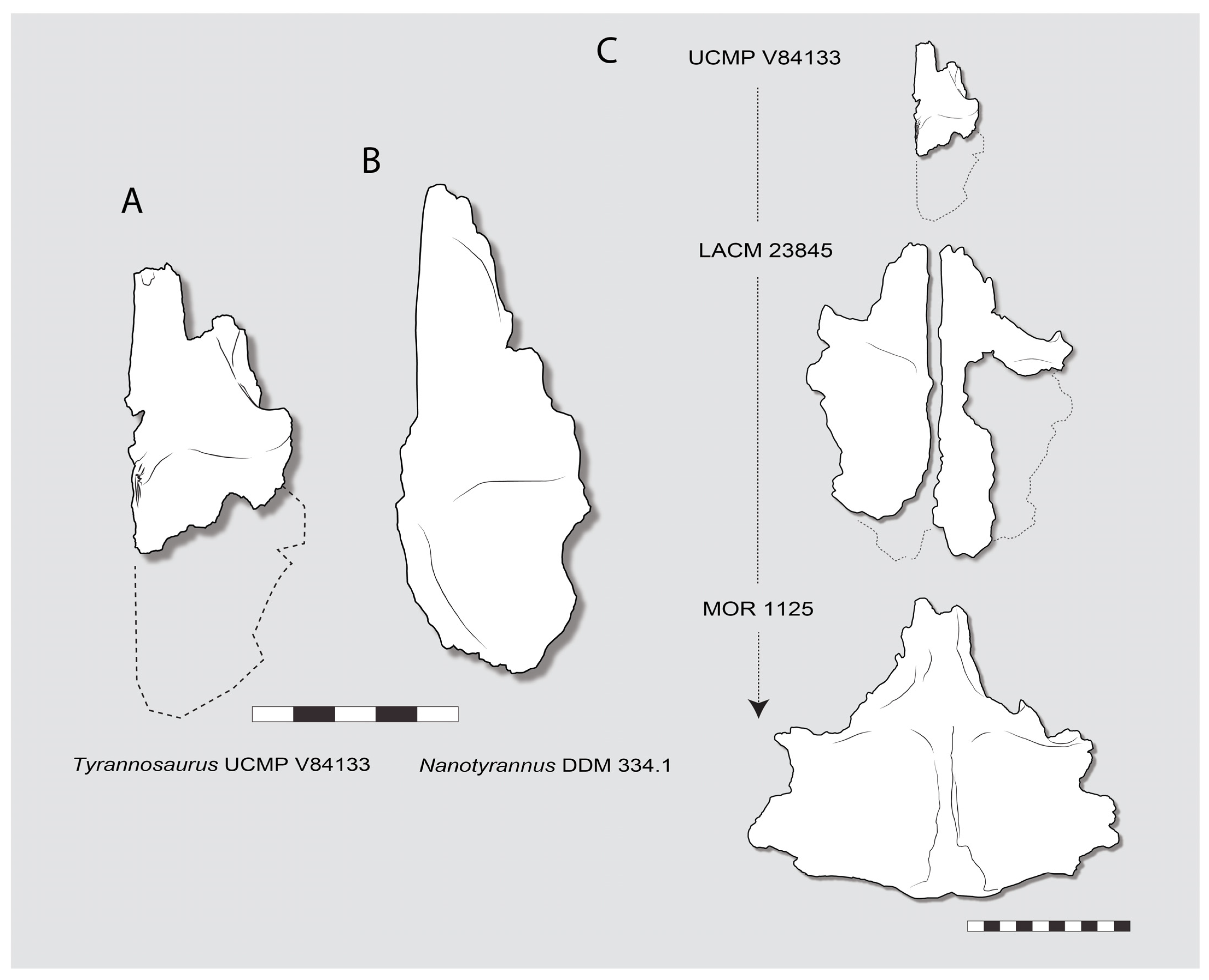
3.5. Existence of Juvenile Tyrannosaurus Refutes Identification of Nanotyrannus as Juvenile Tyrannosaurus
3.6. Phylogenetic Analysis Suggests Nanotyrannus Is Not a Member of Tyrannosauridae
4. Discussion
4.1. Summary of Evidence for the Validity of Nanotyrannus
4.2. Critique of Previous Work Synonymizing Nanotyrannus and Tyrannosaurus
4.3. Affinities of Nanotyrannus-like Dinosaurs
4.4. Weaknesses in the Hypothesis
4.5. Systematics and The Evolution of Development
4.6. Implications
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brusatte, S.L.; Carr, T.D. The phylogeny and evolutionary history of tyrannosauroid dinosaurs. Sci. Rep. 2016, 6, 20252. [Google Scholar] [CrossRef]
- Zanno, L.E.; Makovicky, P.J. Neovenatorid theropods are apex predators in the Late Cretaceous of North America. Nat. Commun. 2013, 4, 2827. [Google Scholar] [CrossRef]
- Brusatte, S.L.; Benson, R.B.; Hutt, S. The osteology of Neovenator salerii (Dinosauria: Theropoda) from the Wealden group (Barremian) of the Isle of Wight. Paleontogr. Soc. Monogr. 2008, 162, 1–75. [Google Scholar]
- Holtz, T.R. Tyrannosauroidea. In The Dinosauria, 2nd ed.; Weishampel, D.B., Dodson, P., Osmolska, H., Eds.; University of California Press: Berkeley, CA, USA, 2004; pp. 111–136. [Google Scholar]
- Brusatte, S.L.; Norell, M.A.; Carr, T.D.; Erickson, G.M.; Hutchinson, J.R.; Balanoff, A.M.; Bever, G.S.; Choiniere, J.N.; Makovicky, P.J.; Xu, X. Tyrannosaur paleobiology: New research on ancient exemplar organisms. Science 2010, 329, 1481–1485. [Google Scholar] [CrossRef]
- Osborn, H.F. Tyrannosaurus and other Cretaceous carnivorous dinosaurs. Bull. Am. Mus. Nat. Hist. 1905, 35, 733–771. [Google Scholar]
- Brochu, C.A. Osteology of Tyrannosaurus rex: Insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull. J. Vertebr. Paleontol. Mem. 2003, 7, 1–138. [Google Scholar] [CrossRef]
- Erickson, G.M.; Makovicky, P.J.; Currie, P.J.; Norell, M.A.; Yerby, S.A.; Brochu, C.A. Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs. Nature 2004, 430, 772–775. [Google Scholar] [CrossRef]
- Carr, T.D. Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria). J. Vertebr. Paleontol. 1999, 19, 497–520. [Google Scholar] [CrossRef]
- Carr, T.D. A high-resolution growth series of Tyrannosaurus rex obtained from multiple lines of evidence. PeerJ 2020, 8, e9192. [Google Scholar] [CrossRef]
- Hutchinson, J.R.; Garcia, M. Tyrannosaurus was not a fast runner. Nature 2002, 415, 1018–1021. [Google Scholar] [CrossRef]
- DePalma, R.A.; Burnham, D.A.; Martin, L.D.; Rothschild, B.M.; Larson, P.L. Physical evidence of predatory behavior in Tyrannosaurus rex. Proc. Natl. Acad. Sci. USA 2013, 110, 12560–12564. [Google Scholar] [CrossRef]
- Longrich, N.R.; Horner, J.R.; Erickson, G.M.; Currie, P.J. Cannibalism in Tyrannosaurus rex. PLoS ONE 2010, 5, e13419. [Google Scholar] [CrossRef]
- Erickson, G.M.; Van Kirk, S.D.; Su, J.; Levenston, M.E.; Caler, W.E.; Carter, D.R. Bite-force estimation for Tyrannosaurus rex from tooth-marked bones. Nature 1996, 382, 706–708. [Google Scholar] [CrossRef]
- Chin, K.; Tokaryk, T.T.; Erickson, G.M.; Calk, L.C. A king-sized theropod coprolite. Nature 1998, 393, 680–682. [Google Scholar] [CrossRef]
- Larson, P.L. Variation and Sexual Dimorphism in Tyrannosaurus rex. In Tyrannosaurus rex, the Tyrant King; Larson, P.L., Carpenter, K., Eds.; Indiana University Press: Bloomington, IN, USA, 2008; pp. 103–128. [Google Scholar]
- Carr, T.D.; Williamson, T.E. Diversity of late Maastrichtian Tyrannosauridae (Dinosauria: Theropoda) from western North America. Zool. J. Linn. Soc. 2004, 142, 479–523. [Google Scholar] [CrossRef]
- Paul, G.S.; Persons, W.S.; Van Raalte, J. The Tyrant Lizard King, Queen and Emperor: Multiple Lines of Morphological and Stratigraphic Evidence Support Subtle Evolution and Probable Speciation Within the North American Genus Tyrannosaurus. Evol. Biol. 2022, 49, 156–179. [Google Scholar] [CrossRef]
- Carr, T.D.; Napoli, J.G.; Brusatte, S.L.; Holtz, T.R.; Hone, D.W.; Williamson, T.E.; Zanno, L.E. Insufficient Evidence for Multiple Species of Tyrannosaurus in the Latest Cretaceous of North America: A Comment on “The Tyrant Lizard King, Queen and Emperor: Multiple Lines of Morphological and Stratigraphic Evidence Support Subtle Evolution and Probable Speciation Within the North American Genus Tyrannosaurus”. Evol. Biol. 2022, 39, 327–341. [Google Scholar]
- Breithaup, B.H.; Southwell, E.H.; Matthews, N.A. Dynamosaurus imperiosus and the earliest discoveries of Tyrannosaurus rex in Wyoming and the West. New Mex. Mus. Nat. Hist. Sci. Bull. 2006, 35, 257–258. [Google Scholar]
- Marsh, O.C. Description of new dinosaurian reptiles. Am. J. Sci. 1890, 39, 81–86. [Google Scholar] [CrossRef]
- Cope, E.D. Fourth note on the Dinosauria of the Laramie. Am. Nat. 1892, 26, 756–758. [Google Scholar]
- Paul, G.S. Predatory Dinosaurs of the World; Simon and Schuster: New York, NY, USA, 1988; p. 464. [Google Scholar]
- Olshevsky, G. The origin and evolution of the tyrannosaurids. Kyoryugaku Saizensen 1995, 9, 92–119. [Google Scholar]
- Osborn, H.F. Tyrannosaurus, Upper Cretaceous carnivorous dinosaur (second communication). Bull. Am. Mus. Nat. Hist. 1906, 22, 281–296. [Google Scholar]
- Scannella, J.B. Anagenesis in Triceratops: Evidence from a newly resolved stratigraphic framework for the Hell Creek Formation. Cincinnati. Mus. Cent. Sci. Contrib. 2009, 3, 148–149. [Google Scholar]
- Gilmore, C.W. A new carnivorous dinosaur from the Lance Formation. Smithson. Misc. Collect. 1946, 106, 1–19. [Google Scholar]
- Rozhdestvensky, A.K. Growth changes in Asian dinosaurs and some problems of their taxonomy. Paleontol. Žurnal 1965, 3, 95–109. [Google Scholar]
- Russell, D.A. Tyrannosaurs from the Late Cretaceous of Western Canada. Natl. Mus. Nat. Sci. Publ. Palaeontol. 1970, 1, 1–34. [Google Scholar]
- Bakker, R.; Williams, M.; Currie, P.J. Nanotyrannus, a new genus of pygmy tyrannosaur, from the latest Cretaceous of Montana. Hunteria 1988, 1, 1–30. [Google Scholar]
- Molnar, R. A new theropod dinosaur from the Upper Cretaceous of central Montana. J. Paleontol. 1978, 52, 73–82. [Google Scholar]
- Molnar, R.E.; Carpenter, K. The Jordan theropod (Maastrichtian, Montana, USA) referred to the genus Aublysodon. Geobios 1989, 22, 445–454. [Google Scholar] [CrossRef]
- Leidy, J.F. Remarks on a jaw fragment of Megalosaurus. Proc. Phila. Acad. Nat. Sci. 1868, 20, 197–200. [Google Scholar]
- Carpenter, K. Tyrannosaurids (Dinosauria) of Asia and North America. In Aspects of Nonmarine Cretaceous Geology; Mateer, N.J., Chen, P.-J., Eds.; China Ocean Press: Beijing, China, 1992; pp. 250–268. [Google Scholar]
- Tumarkin-Deratzian, A.R. Evaluation of long bone surface textures as ontogenetic in centrosaurine ceratopsids. Anat. Rec. 2009, 292, 1485–1500. [Google Scholar] [CrossRef] [PubMed]
- Tumarkin-Deratzian, A.R.; Vann, D.R.; Dodson, P. Bone surface texture as an ontogenetic indicator in long bones of the Canada goose Branta canadensis (Anseriformes: Anatidae). Zool. J. Linn. Soc. 2006, 148, 133–168. [Google Scholar] [CrossRef]
- Holtz, T.R., Jr. The phylogeny and taxonomy of the Tyrannosauridae. In Mesozoic Vertebrate Life; Tanke, D.H., Carpenter, K., Eds.; Indiana University Press: Bloomington, IN, USA, 2001; pp. 64–83. [Google Scholar]
- Brusatte, S.L.; Carr, T.D.; Williamson, T.E.; Holtz, T.R., Jr.; Hone, D.W.; Williams, S.A. Dentary groove morphology does not distinguish ‘Nanotyrannus’ as a valid taxon of tyrannosauroid dinosaur. Comment on:“Distribution of the dentary groove of theropod dinosaurs: Implications for theropod phylogeny and the validity of the genus Nanotyrannus Bakker et al. 1988”. Cretac. Res. 2016, 65, 232–237. [Google Scholar]
- Currie, P.J. Cranial anatomy of tyrannosaurid dinosaurs from the Late Cretaceous of Alberta, Canada. Acta Palaeontol. Pol. 2003, 48, 191–226. [Google Scholar]
- Larson, P. The case for Nanotyrannus. In Tyrannosaurid Paleobiology; Parrish, J.M., Molnar, R.E., Currie, P.J., Koppelhus, E.B., Eds.; Indiana University Press: Bloomington, IN, USA, 2013; pp. 15–53. [Google Scholar]
- Henderson, M.D.; Harrison, W.H. Taphonomy and environment of deposition of juvenile tyrannosaurid skeleton from the Hell Creek Formation (latest Maastrichtian) of southeastern Montana. In Tyrannosaurus rex, the Tyrant King; Indiana University Press: Bloomington, IN, USA, 2008; pp. 82–90. [Google Scholar]
- Larson, P. The Validity of Nanotyrannus lancensis (Theropoda, Lancian—Upper Maastrichtian of North America). In Proceedings of the Society of Vertebrate Paleontology, Los Angeles, CA, USA, 30 October–2 November 2013. [Google Scholar]
- Witmer, L.M.; Ridgely, R.C. New insights into the brain, braincase, and ear region of tyrannosaurs (Dinosauria, Theropoda), with implications for sensory organization and behavior. Anat. Rec. 2009, 292, 1266–1296. [Google Scholar] [CrossRef]
- Witmer, L.M.; Ridgely, R.C. The Cleveland tyrannosaur skull (Nanotyrannus or Tyrannosaurus): New findings based on CT scanning, with special reference to the braincase. Kirtlandia 2010, 57, 61–81. [Google Scholar]
- Schmerge, J.D.; Rothschild, B.M. Distribution of the dentary groove of theropod dinosaurs: Implications for theropod phylogeny and the validity of the genus Nanotyrannus Bakker et al., 1988. Cretac. Res. 2016, 61, 26–33. [Google Scholar] [CrossRef]
- Woodward, H.N.; Tremaine, K.; Williams, S.A.; Zanno, L.E.; Horner, J.R.; Myhrvold, N. Growing up Tyrannosaurus rex: Osteohistology refutes the pygmy “Nanotyrannus” and supports ontogenetic niche partitioning in juvenile Tyrannosaurus. Sci. Adv. 2020, 6, eaax6250. [Google Scholar] [CrossRef]
- Jevnikar, E.; Zanno, L. Bimodal trajectories and unresolved early growth stages in Tyrannosaurus rex growth. In Proceedings of the Society of Vertebrate Paleontology 2021 Annual Meeting, Minneapolis, MN, USA, 3–7 November 2021; p. 151. [Google Scholar]
- Swofford, D.L. PAUP *. Phylogenetic Analysis Using Parsimony (* and Other Methods); 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Cullen, T.M.; Canale, J.I.; Apesteguía, S.; Smith, N.D.; Hu, D.; Makovicky, P.J. Osteohistological analyses reveal diverse strategies of theropod dinosaur body-size evolution. Proc. R. Soc. B 2020, 287, 20202258. [Google Scholar] [CrossRef]
- Campione, N.E.; Evans, D.C.; Brown, C.M.; Carrano, M.T. Body mass estimation in non-avian bipeds using a theoretical conversion to quadruped stylopodial proportions. Methods Ecol. Evol. 2014, 5, 913–923. [Google Scholar] [CrossRef]
- Griffin, C. Using osteohistology to determine the taxonomic validity of the Late Cretaceous dinosaur Nanotyrannus lancensis Bakker et al, 1988 (Theropoda: Tyrannosauridae). In Proceedings of the The Research and Scholarship Symposium, Cedarville, OH, USA, 16 April 2014. [Google Scholar]
- Dalman, S.G.; Loewen, M.A.; Pyron, R.A.; Jasinski, S.E.; Malinzak, D.E.; Lucas, S.G.; Fiorillo, A.R.; Currie, P.J.; Longrich, N.R. A giant tyrannosaur from the Campanian–Maastrichtian of southern North America and the evolution of tyrannosaurid gigantism. Sci. Rep. 2024, in press. [Google Scholar]
- Hurum, J.H.; Sabath, K. Giant theropod dinosaurs from Asia and North America: Skulls of Tarbosaurus bataar and Tyrannosaurus rex compared. Acta Palaeontol. Pol. 2003, 48, 161–190. [Google Scholar]
- Tsuihiji, T.; Watabe, M.; Tsogtbaatar, K.; Tsubamoto, T.; Barsbold, R.; Suzuki, S.; Lee, A.H.; Ridgely, R.C.; Kawahara, Y.; Witmer, L.M. Cranial osteology of a juvenile specimen of Tarbosaurus bataar (Theropoda, Tyrannosauridae) from the Nemegt Formation (Upper Cretaceous) of Bugin Tsav, Mongolia. J. Vertebr. Paleontol. 2011, 31, 497–517. [Google Scholar] [CrossRef]
- Fowler, D.W.; Woodward, H.N.; Freedman, E.A.; Larson, P.L.; Horner, J.R. Reanalysis of “Raptorex kriegsteini”: A juvenile tyrannosaurid dinosaur from Mongolia. PLoS ONE 2011, 6, e21376. [Google Scholar] [CrossRef]
- Loewen, M.A.; Irmis, R.B.; Sertich, J.J.; Currie, P.J.; Sampson, S.D. Tyrant dinosaur evolution tracks the rise and fall of Late Cretaceous oceans. PLoS ONE 2013, 8, e79420. [Google Scholar] [CrossRef] [PubMed]
- Voris, J.T.; Therrien, F.; Zelenitsky, D.K.; Brown, C.M. A new tyrannosaurine (Theropoda: Tyrannosauridae) from the Campanian Foremost Formation of Alberta, Canada, provides insight into the evolution and biogeography of tyrannosaurids. Cretac. Res. 2020, 110, 104388. [Google Scholar] [CrossRef]
- Carr, T.D.; Williamson, T.E.; Britt, B.B.; Stadtman, K. Evidence for high taxonomic and morphologic tyrannosauroid diversity in the Late Cretaceous (Late Campanian) of the American Southwest and a new short-skulled tyrannosaurid from the Kaiparowits Formation of Utah. Naturwissenschaften 2011, 98, 241–246. [Google Scholar] [CrossRef]
- Carr, T.D.; Williamson, T.E. Bistahieversor sealeyi, gen. et sp. nov., a new tyrannosauroid from New Mexico and the origin of deep snouts in Tyrannosauroidea. J. Vertebr. Paleontol. 2010, 30, 1–16. [Google Scholar] [CrossRef]
- Warshaw, E.A.; Fowler, D.W. A transitional species of Daspletosaurus Russell, 1970 from the Judith River Formation of eastern Montana. PeerJ 2022, 10, e14461. [Google Scholar] [CrossRef]
- Carr, T.D.; Varricchio, D.J.; Sedlmayr, J.C.; Roberts, E.M.; Moore, J.R. A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Molnar, R.E. A distinctive theropod dinosaur from the Upper Cretaceous of Baja California (Mexico). J. Paleontol. 1974, 48, 1009–1017. [Google Scholar]
- Zanno, L.E.; Tucker, R.T.; Canoville, A.; Avrahami, H.M.; Gates, T.A.; Makovicky, P.J. Diminutive fleet-footed tyrannosauroid narrows the 70-million-year gap in the North American fossil record. Commun. Biol. 2019, 2, 64. [Google Scholar] [CrossRef] [PubMed]
- Nesbitt, S.J.; Denton Jr, R.K.; Loewen, M.A.; Brusatte, S.L.; Smith, N.D.; Turner, A.H.; Kirkland, J.I.; McDonald, A.T.; Wolfe, D.G. A mid-Cretaceous tyrannosauroid and the origin of North American end-Cretaceous dinosaur assemblages. Nat. Ecol. Evol. 2019, 3, 892–899. [Google Scholar] [CrossRef] [PubMed]
- Brusatte, S.L.; Averianov, A.; Sues, H.-D.; Muir, A.; Butler, I.B. New tyrannosaur from the mid-Cretaceous of Uzbekistan clarifies evolution of giant body sizes and advanced senses in tyrant dinosaurs. Proc. Natl. Acad. Sci. USA 2016, 113, 3447–3452. [Google Scholar] [CrossRef] [PubMed]
- Lambe, L.M. On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon. Ott. Nat. 1914, 28, 13–20. [Google Scholar]
- Currie, P.J. Theropods, including birds. In Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed; Currie, P.J., Koppelhus, E.B., Farlow, J.O., Eds.; Indiana University Press: Bloomington, IN, USA, 2005; pp. 367–397. [Google Scholar]
- Fiorillo, A.R.; Tykoski, R.S. A diminutive new tyrannosaur from the top of the world. PLoS ONE 2014, 9, e91287. [Google Scholar] [CrossRef] [PubMed]
- Carr, T.D. Significant Geographic Range Extension for the Sympatric Tyrannosaurids Albertosaurus Libratus and Daspletosaurus Torosus from the Judith River Formation (Late Campanian) of Northern Montana. In Proceedings of The Society of Vertebrate Paleontology 2018 Annual Meeting, Albuquerque, NM, USA, 17–20 October 2018. [Google Scholar]
- Brusatte, S.L.; Carr, T.D.; Norell, M.A. The osteology of Alioramus, a gracile and long-snouted tyrannosaurid (Dinosauria: Theropoda) from the Late Cretaceous of Mongolia. Bull. Am. Mus. Nat. Hist. 2012, 2012, 1–197. [Google Scholar] [CrossRef]
- Mallon, J.C.; Bura, J.R.; Schumann, D.; Currie, P.J. A problematic tyrannosaurid (Dinosauria: Theropoda) skeleton and its implications for tyrannosaurid diversity in the Horseshoe Canyon Formation (Upper Cretaceous) of Alberta. Anat. Rec. 2020, 303, 673–690. [Google Scholar] [CrossRef]
- Longrich, N.R.; Isasmendi, E.; Pereda-Suberbiola, X.; Jalil, N.-E. New fossils of Abelisauridae (Dinosauria: Theropoda) from the upper Maastrichtian of Morocco, North Africa. Cretac. Res. 2023, 152, 105677. [Google Scholar] [CrossRef]
- Gasparini, Z.; Sterli, J.; Parras, A.; O’Gorman, J.P.; Salgado, L.; Varela, J.; Pol, D. Late Cretaceous reptilian biota of the La Colonia Formation, central Patagonia, Argentina: Occurrences, preservation and paleoenvironments. Cretac. Res. 2015, 54, 154–168. [Google Scholar] [CrossRef]
- Novas, F.E.; Chatterjee, S.; Rudra, D.K.; Datta, P. Rahiolisaurus gujaratensis, n. gen. n. sp., a New Abelisaurid Theropod from the Late Cretaceous of India. In New Aspects of Mesozoic Biodiversity; Bandyopadhyay, S., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 45–62. [Google Scholar]
- Wilson, J.A.; Sereno, P.C.; Srivastava, S.; Bhatt, D.K.; Khosla, A.; Sahni, A. A new abelisaurid (Dinosauria, Theropoda) from the Lameta Formation (Cretaceous, Maastrichtian) of India. Contrib. Mus. Paleontol. Univ. Mich. 2003, 31, 1–42. [Google Scholar]
- Chatterjee, S. Indosuchus and Indosaurus, Cretaceous carnosaurs from India. J. Paleontol. 1978, 52, 570–580. [Google Scholar]
- Huene, F.R.; Matley, C.A. The Cretaceous Saurischia and Ornithischia of the Central provinces. Palaeontol. Indica (New Ser.) Mem. Geol. Surv. India 1933, 21, 1–74. [Google Scholar]
- Cau, A.; Dalla Vecchia, F.M.; Fabbri, M. A thick-skulled theropod (Dinosauria, Saurischia) from the Upper Cretaceous of Morocco with implications for carcharodontosaurid cranial evolution. Cretac. Res. 2013, 40, 251–260. [Google Scholar] [CrossRef]
- Sereno, P.C.; Dutheil, D.B.; Iarochene, M.; Larsson, H.C. Predatory dinosaurs from the Sahara and Late Cretaceous faunal differentiation. Science 1996, 272, 986. [Google Scholar] [CrossRef] [PubMed]
- Calvo, J.O.; Rubilar-Rogers, D.; Moreno, K. A new Abelisauridae (Dinosauria: Theropoda) from northwest Patagonia. Ameghiniana 2004, 41, 555–563. [Google Scholar]
- Coria, R.A.; Salgado, L. A new giant carnivorous dinosaur from the Cretaceous of Patagonia. Nature 1995, 377, 224–226. [Google Scholar] [CrossRef]
- Mateus, O. Late Jurassic dinosaurs from the Morrison Formation (USA), the Lourinha and Alcobaça formations (Portugal), and the Tendaguru Beds (Tanzania): A comparison. New Mex. Mus. Nat. Hist. Sci. Bull. 2006, 36, 223–231. [Google Scholar]
- Van Valkenburgh, B.; Hayward, M.W.; Ripple, W.J.; Meloro, C.; Roth, V.L. The impact of large terrestrial carnivores on Pleistocene ecosystems. Proc. Natl. Acad. Sci. USA 2016, 113, 862–867. [Google Scholar] [CrossRef]
- Barnett, R.; Barnes, I.; Phillips, M.J.; Martin, L.D.; Harington, C.R.; Leonard, J.A.; Cooper, A. Evolution of the extinct Sabretooths and the American cheetah-like cat. Curr. Biol. 2005, 15, R589–R590. [Google Scholar] [CrossRef]
- Nowak, R.M. Walker’s Mammals of the World, 6th ed.; JHU Press: Baltimore, MD, USA, 1999. [Google Scholar]
- Bibi, F.; Kiessling, W. Continuous evolutionary change in Plio-Pleistocene mammals of eastern Africa. Proc. Natl. Acad. Sci. USA 2015, 112, 10623–10628. [Google Scholar] [CrossRef] [PubMed]
- Diamond, J. The Third Chimpanzee: The Evolution and Future of the Human Animal; Harper Perennial: New York, NY, USA, 1992. [Google Scholar]
- Bardet, N.; Falconnet, J.; Fischer, V.; Houssaye, A.; Jouve, S.; Pereda-Suberbiola, X.; Perez-García, A.; Rage, J.-C.; Vincent, P. Mesozoic marine reptile palaeobiogeography in response to drifting plates. Gondwana Res. 2014, 26, 869–887. [Google Scholar] [CrossRef]
- Longrich, N.R.; Jalil, N.-E.; Khaldoune, F.; Yazami, O.K.; Pereda-Suberbiola, X.; Bardet, N. Thalassotitan atrox, a giant predatory mosasaurid (Squamata) from the Upper Maastrichtian Phosphates of Morocco. Cretac. Res. 2022, 140, 105315. [Google Scholar] [CrossRef]
- Lambert, O.; Bianucci, G.; Post, K.; De Muizon, C.; Salas-Gismondi, R.; Urbina, M.; Reumer, J. The giant bite of a new raptorial sperm whale from the Miocene epoch of Peru. Nature 2010, 466, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Hart, J.L. Pacific Fishes of Canada; Fisheries Research Board of Canada Bulletin: Ottowa, IL, USA, 1973; Volume 180, p. 741. [Google Scholar]
- Doughty, P.; Kealley, L.; Fitch, A.; Donnellan, S.C. A new diminutive species of Varanus from the Dampier Peninsula, western Kimberley region, Western Australia. Rec. West. Aust. Mus. 2014, 29, 128–140. [Google Scholar] [CrossRef]
- Dunning, J.B. Handbook of Avian Body Masses, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2007; p. 672. [Google Scholar]
- Cope, E.D. Discovery of a gigantic dinosaur in the Cretaceous of New Jersey. Proc. Acad. Nat. Sci. Phila. 1866, 18, 275–279. [Google Scholar]
- Carr, T.D.; Williamson, T.E.; Schwimmer, D.R. A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama. J. Vertebr. Paleontol. 2005, 25, 119–143. [Google Scholar] [CrossRef]
- Brusatte, S.L.; Carr, T.D.; Erickson, G.M.; Bever, G.S.; Norell, M. A long-snouted, multihorned tyrannosaurid from the Late Cretaceous of Mongolia. Proc. Natl. Acad. Sci. USA 2009, 106, 17251–17266. [Google Scholar] [CrossRef]
- Kurzanov, S. New carnosaur from the Late Cretaceous of Nogon− Tsav, Mongolia. Sovmest. Sov. Mong. Paleon Tologičeskaâ Eksped. Tr. 1976, 3, 93–104. [Google Scholar]
- Lü, J.; Yi, L.; Brusatte, S.L.; Yang, L.; Li, H.; Chen, L. A new clade of Asian Late Cretaceous long-snouted tyrannosaurids. Nat. Commun. 2014, 5, 3788. [Google Scholar] [CrossRef]
- McDonald, A.T.; Wolfe, D.G.; Dooley, A.C., Jr. A new tyrannosaurid (Dinosauria: Theropoda) from the Upper Cretaceous Menefee Formation of New Mexico. PeerJ 2018, 6, e5749. [Google Scholar] [CrossRef]
- Currie, P.J.; Zhiming, D. New information on Shanshanosaurus huoyanshanensis, a juvenile tyrannosaurid (Theropoda, Dinosauria) from the Late Cretaceous of China. Can. J. Earth Sci. 2001, 38, 1729–1737. [Google Scholar] [CrossRef]
- Maleev, E.A. New carnivorous dinosaurs from the Upper Cretaceous of Mongolia. Dokl. Akad. Nauk. SSSR 1955, 104, 779–782. [Google Scholar]
- Hone, D.W.; Wang, K.; Sullivan, C.; Zhao, X.; Chen, S.; Li, D.; Ji, S.; Ji, Q.; Xu, X. A new, large tyrannosaurine theropod from the Upper Cretaceous of China. Cretac. Res. 2011, 32, 495–503. [Google Scholar] [CrossRef]
- Persons IV, W.S.; Currie, P.J.; Erickson, G.M. An older and exceptionally large adult specimen of Tyrannosaurus rex. Anat. Rec. 2020, 303, 656–672. [Google Scholar] [CrossRef]
- Hutchinson, J.R.; Bates, K.T.; Molnar, J.; Allen, V.; Makovicky, P.J. A computational analysis of limb and body dimensions in Tyrannosaurus rex with implications for locomotion, ontogeny, and growth. PLoS ONE 2011, 6, e26037. [Google Scholar] [CrossRef]
- DePalma, R.A.; Burnham, D.A.; Martin, L.D.; Larson, P.L.; Bakker, R.T. The first giant raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation. Paleontol. Contrib. 2015, 14, 1–16. [Google Scholar]
- Evans, D.C.; Larson, D.W.; Currie, P.J. A new dromaeosaurid (Dinosauria: Theropoda) with Asian affinities from the latest Cretaceous of North America. Naturwissenschaften 2013, 100, 1041–1049. [Google Scholar] [CrossRef]
- Longrich, N.R.; Field, D. Torosaurus is not Triceratops: Ontogeny in Chasmosaurine Ceratopsids as a Case Study in Dinosaur Taxonomy. PLoS ONE 2012, 7, e32623. [Google Scholar] [CrossRef]
- Maleev, E.A. Giant carnosaurs of the family Tyrannosauridae. Jt. Sov. Mong. Paleontol. Exped. 1974, 1, 132–191. [Google Scholar]
- Voris, J.T.; Zelenitsky, D.K.; Therrien, F.; Ridgely, R.C.; Currie, P.J.; Witmer, L.M. Two exceptionally preserved juvenile specimens of Gorgosaurus libratus (Tyrannosauridae, Albertosaurinae) provide new insight into the timing of ontogenetic changes in tyrannosaurids. J. Vertebr. Paleontol. 2021, 41, e2041651. [Google Scholar] [CrossRef]
- Voris, J.T.; Zelenitsky, D.K.; Therrien, F.; Currie, P.J. Reassessment of a juvenile Daspletosaurus from the Late Cretaceous of Alberta, Canada with implications for the identification of immature tyrannosaurids. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Brochu, C.A. Closure of neurocentral sutures during crocodylian ontogeny: Implication for maturity assessment in fossil archosaurs. J. Vertebr. Paleontol. 1996, 16, 49–62. [Google Scholar] [CrossRef]
- Goodwin, M.B.; Clemens, W.A.; Horner, J.R.; Padian, K. The smallest known Triceratops skull: New observations on ceratopsid cranial anatomy and ontogeny. J. Vertebr. Paleontol. 2006, 26, 103–112. [Google Scholar] [CrossRef]
- Sereno, P.C.; Tan, L.; Brusatte, S.L.; Kriegstein, H.J.; Zhao, X.; Cloward, K. Tyrannosaurid skeletal design first evolved at small body size. Science 2009, 326, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Sereno, P.C. The pectoral girdle and forelimb of the basal theropod Herrerasaurus ischigualastensis. J. Vertebr. Paleontol. 1994, 13, 425–450. [Google Scholar] [CrossRef]
- Burch, S.H.; Carrano, M.T. An articulated pectoral girdle and forelimb of the abelisaurid theropod Majungasaurus crenatissimus from the Late Cretaceous of Madagascar. J. Vertebr. Paleontol. 2012, 32, 1–16. [Google Scholar] [CrossRef]
- Norell, M.A.; Makovicky, P.J. Important features of the dromaeosaurid skeleton II: Information from newly collected specimens of Velociraptor mongoliensis. Am. Mus. Novit. 1999, 3282, 1–45. [Google Scholar]
- Perle, A.; Norell, M.A.; Clark, J.M. A new maniraptoran theropod Achillobator giganticus (Dromaeosauridae) from the Upper Cretaceous of Burkhant, Mongolia. Dep. Geol. Natl. Univ. Mong. Ulaanbaatar 1999, 101, 1–105. [Google Scholar]
- Jasinski, S.E.; Sullivan, R.M.; Lucas, S.G. Taxonomic composition of the Alamo Wash local fauna from the Upper Cretaceous Ojo Alamo Formation (Naashoibito Member), San Juan Basin, New Mexico. New Mex. Mus. Nat. Hist. Sci. Bull. 2011, 53, 216–265. [Google Scholar]
- Longrich, N.R.; Currie, P.J. A microraptorine (Dinosauria-Dromaeosauridae) from the Late Cretaceous of North America. PNAS 2009, 106, 5002–5008. [Google Scholar] [CrossRef] [PubMed]
- Raath, M.A.; Carpenter, K.; Currie, P.J. Morphological variation in small theropods and its meaning in systematics: Evidence from Syntarsus. In Dinosaur Systematics: Perspectives and Approaches; Currie, P.J., Carpenter, K., Eds.; Cambridge University Press: Cambridge, UK, 1990; pp. 91–105. [Google Scholar]
- Iori, F.V.; de Araújo-Júnior, H.I.; Tavares, S.A.S.; da Silva Marinho, T.; Martinelli, A.G. New theropod dinosaur from the Late Cretaceous of Brazil improves abelisaurid diversity. J. South Am. Earth Sci. 2021, 112, 103551. [Google Scholar] [CrossRef]
- Pol, D.; Rauhut, O.W. A Middle Jurassic abelisaurid from Patagonia and the early diversification of theropod dinosaurs. Proc. R. Soc. B: Biol. Sci. 2012, 279, 3170–3175. [Google Scholar] [CrossRef] [PubMed]
- Cerroni, M.A.; Baiano, M.A.; Canale, J.I.; Agnolín, F.L.; Otero, A.; Novas, F.E. Appendicular osteology of Skorpiovenator bustingorryi (Theropoda, Abelisauridae) with comments on phylogenetic features of abelisaurids. J. Syst. Palaeontol. 2022, 20, 1–32. [Google Scholar] [CrossRef]
- Macdonald, I.; Currie, P.J. Description of a partial Dromiceiomimus (Dinosauria: Theropoda) skeleton with comments on the validity of the genus. Can. J. Earth Sci. 2019, 56, 129–157. [Google Scholar] [CrossRef]
- Osborn, H.F. Skeletal adaptations of Ornitholestes, Struthiomimus, Tyrannosaurus. Bull. Am. Mus. Nat. Hist. 1917, 35, 733–771. [Google Scholar]
- Yao, X.; Sullivan, C.; Tan, Q.; Xu, X. New ornithomimosaurian (Dinosauria: Theropoda) pelvis from the Upper Cretaceous Erlian Formation of Nei Mongol, North China. Cretac. Res. 2022, 137, 105234. [Google Scholar] [CrossRef]
- Xu, L.; Kobayashi, Y.; Lü, J.; Lee, Y.-N.; Liu, Y.; Tanaka, K.; Zhang, X.; Jia, S.; Zhang, J. A new ornithomimid dinosaur with North American affinities from the Late Cretaceous Qiupa Formation in Henan Province of China. Cretac. Res. 2011, 32, 213–222. [Google Scholar] [CrossRef]
- Brown, C.M.; Russell, A.P.; Ryan, M.J. Pattern and transition of surficial bone texture of the centrosaurine frill and their ontogenetic and taxonomic implications. J. Vertebr. Paleontol. 2009, 29, 132–141. [Google Scholar] [CrossRef]
- Sampson, S.D.; Ryan, M.J.; Tanke, D.H. Craniofacial ontogeny in centrosaurine dinosaurs (Ornithischia: Ceratopsidae): Taxonomic and behavioral implications. Zooogical J. Linn. Soc. 1997, 121, 293–337. [Google Scholar] [CrossRef]
- Horner, J.R.; de Ricqlès, A.; Padian, K. Variation in dinosaur skeletochronology indicators: Implications for age assessment and physiology. Paleobiology 1999, 25, 295–304. [Google Scholar] [CrossRef]
- Erickson, G.M.; Tumanova, T.A. Growth curve of Psittacosaurus mongoliensis Osborn (Ceratopsia: Psittacosauridae) inferred from long bone histology. Zool. J. Linn. Soc. 2000, 130, 551–566. [Google Scholar] [CrossRef]
- Griebeler, E.M.; Klein, N.; Sander, P.M. Aging, maturation and growth of sauropodomorph dinosaurs as deduced from growth curves using long bone histological data: An assessment of methodological constraints and solutions. PLoS ONE 2013, 8, e67012. [Google Scholar] [CrossRef]
- Erickson, G.M. On dinosaur growth. Annu. Rev. Earth Planet. Sci. 2014, 42, 675–697. [Google Scholar] [CrossRef]
- Saitta, E.T.; Stockdale, M.T.; Longrich, N.R.; Bonhomme, V.; Benton, M.J.; Cuthill, I.C.; Makovicky, P.J. An effect size statistical framework for investigating sexual dimorphism in non-avian dinosaurs and other extinct taxa. Biol. J. Linn. Soc. 2020, 131, 231–273. [Google Scholar] [CrossRef]
- Ralls, K.; Mesnick, S. Sexual Dimorphism. In Encyclopedia of Marine Mammals; Elsevier: Amsterdam, The Netherlands, 2009; pp. 1005–1011. [Google Scholar]
- Laws, R.M. The elephant seal (Mirounga leonina, Linn.): I. Growth and age; HMSO: London, UK, 1953; p. 62. [Google Scholar]
- McCann, T.S.; Fedak, M.A.; Harwood, J. Parental investment in southern elephant seals, Mirounga leonina. Behav. Ecol. Sociobiol. 1989, 25, 81–87. [Google Scholar] [CrossRef]
- Molnar, R.E. The cranial morpholgy of Tyrannosaurus rex Palaeontographica. Abt. A Paläozoologie Stratigr. 1991, 217, 137–176. [Google Scholar]
- Yun, C.-G.; Peters, G.F.; Currie, P.J. Allometric growth in the frontals of the Mongolian theropod dinosaur Tarbosaurus bataar. Acta Palaeontol. Pol. 2022, 67, 601–615. [Google Scholar] [CrossRef]
- Forster, C.A. Species resolution in Triceratops: Cladistic and morphometric approaches. J. Vertebr. Paleontol. 1996, 16, 259–270. [Google Scholar] [CrossRef]
- Scannella, J.B.; Fowler, D.W.; Goodwin, M.B.; Horner, J.R. Evolutionary trends in Triceratops from the Hell Creek Formation, Montana. Proc. Natl. Acad. Sci. USA 2014, 111, 10245–10250. [Google Scholar] [CrossRef]
- Pringle, H. Selling America’s fossil record. Science 2014, 343, 364–367. [Google Scholar] [CrossRef]
- Currie, P.J.; Rigby, K.J.; Sloan, R.E. Theropod teeth from the Judith River Formation of southern Alberta, Canada. In Dinosaur Systematics: Perspectives and Approaches; Currie, P.J., Carpenter, K., Eds.; Cambridge University Press: Cambridge, UK, 1990; pp. 107–125. [Google Scholar]
- Hone, D.W.; Farke, A.A.; Wedel, M.J. Ontogeny and the fossil record: What, if anything, is an adult dinosaur? Biol. Lett. 2016, 12, 20150947. [Google Scholar] [CrossRef]
- Erickson, G.M.; Currie, P.J.; Inouye, B.D.; Winn, A.A. Tyrannosaur life tables: An example of nonavian dinosaur population biology. Science 2006, 313, 213–217. [Google Scholar] [CrossRef]
- Darwin, C. The Descent of Man and Selection in Relation to Sex; Murray: London, UK, 1871. [Google Scholar]
- Horner, J.R.; Goodwin, M.B. Major cranial changes during Triceratops ontogeny. Proc. R. Soc. B 2006, 273, 2757–2761. [Google Scholar]
- Brown, B.; Schlaikjer, E.M. The structure and relationships of Protoceratops. Ann. N. Y. Acad. Sci. 1940, 40, 133–266. [Google Scholar] [CrossRef]
- Carr, T.D. A reappraisal of tyrannosauroid fossils from the Iren Dabasu Formation (Coniacian–Campanian), Inner Mongolia, People’s Republic of China. J. Vertebr. Paleontol. 2022, 42, e2199817. [Google Scholar] [CrossRef]
- De Beer, G.R. Embryos and Ancestors, 2nd ed.; Clarendon Press: Oxford, UK, 1951; p. 159. [Google Scholar]
- Bhullar, B.-A.S.; Marugán-Lobón, J.; Racimo, F.; Bever, G.S.; Rowe, T.B.; Norell, M.A.; Abzhanov, A. Birds have paedomorphic dinosaur skulls. Nature 2012, 487, 223–226. [Google Scholar]
- Naish, D.; Cau, A. The osteology and affinities of Eotyrannus lengi, a tyrannosauroid theropod from the Wealden Supergroup of southern England. PeerJ 2022, 10, e12727. [Google Scholar] [CrossRef]
- Funston, G.F.; Powers, M.J.; Whitebone, S.A.; Brusatte, S.L.; Scannella, J.B.; Horner, J.R.; Currie, P.J. Baby tyrannosaurid bones and teeth from the Late Cretaceous of western North America. Can. J. Earth Sci. 2021, 58, 756–777. [Google Scholar]
- Gilmore, C.W. A new dinosaur from the Lance Formation of Wyoming. Smithson. Misc. Collect. 1913, 61, 1–5. [Google Scholar]
- Gilmore, C.W. A new coelurid dinosaur from the Belly River Cretaceous of Alberta. Can. Geol. Surv. Bull. 1924, 38, 210639511. [Google Scholar]
- Gilmore, C.W. On the dinosaurian fauna of the Iren Dabasu Formation. Bull. Am. Mus. Nat. Hist. 1933, 67, 23–78. [Google Scholar]
- Gilmore, C.W. A new sauropod dinosaur from the Ojo Alamo Formation of New Mexico. Smithson. Misc. Collect. 1922, 72, 1–9. [Google Scholar]
- Gilmore, C.W. A new ceratopsian dinosaur from the Upper Cretaceous of Montana, with note on Hypacrosaurus. Smithson. Misc. Collect. 1914, 63, 1–10. [Google Scholar]
- Larson, N.L.; Larson, P.L.; Carpenter, K. One hundred years of Tyrannosaurus rex: The skeletons. Tyrannosaurus Rex Tyrant King 2008, 1, 1–56. [Google Scholar]
- Mallon, J.C.; Evans, D.C.; Ryan, M.J.; Anderson, J.S. Megaherbivorous dinosaur turnover in the Dinosaur Park Formation (upper Campanian) of Alberta, Canada. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 350-352, 124–138. [Google Scholar] [CrossRef]
- Sampson, S.D.; Loewen, M.A.; Farke, A.A.; Roberts, E.M.; Forster, C.A.; Smith, J.A.; Titus, A.L. New horned dinosaurs from Utah provide evidence for intracontinental dinosaur endemism. PLoS ONE 2010, 5, e12292. [Google Scholar] [CrossRef]
- Longrich, N.R. Titanoceratops ouranos, a giant horned dinosaur from the Late Campanian of New Mexico. Cretac. Res. 2011, 32, 264–276. [Google Scholar] [CrossRef]
- Hanai, T.; Tsuihiji, T. Description of tooth ontogeny and replacement patterns in a juvenile Tarbosaurus bataar (Dinosauria: Theropoda) using CT-scan data. Anat. Rec. 2019, 302, 1210–1225. [Google Scholar] [CrossRef]
- Longrich, N.R.; Scriberas, J.; Wills, M.A. Severe extinction and rapid recovery of mammals across the Cretaceous-Paleogene boundary, and the effects of rarity on patterns of extinction and recovery. J. Evol. Biol. 2016, 29, 1495–1512. [Google Scholar] [CrossRef]
- Gao, K.-Q.; Fox, R.C. Taxonomy and evoluton of Late Cretaceous lizards (Reptilia: Squamata) from Western Canada. Bull. Carnegie Mus. Nat. Hist. 1996, 33, 1–107. [Google Scholar] [CrossRef]
- Longrich, N.R.; Bhullar, B.-A.S.; Gauthier, J. Mass extinction of lizards and snakes at the Cretaceous-Paleogene boundary. Proc. Natl. Acad. Sci. USA 2012, 109, 21396–21401. [Google Scholar] [CrossRef]
- Sweet, A.R.; Braman, D.R. Cretaceous–Tertiary palynofloral perturbations and extinctions within the Aquilapollenites Phytogeographic Province. Can. J. Earth Sci. 2001, 38, 249–269. [Google Scholar] [CrossRef]
- Johnson, K.R. Megaflora of the Hell Creek and lower Fort Union Formations in the western Dakotas: Vegetational response to climate change, the Cretaceous-Tertiary boundary event, and rapid marine transgression. Geol. Soc. Am. Spec. Pap. 2002, 361, 329–391. [Google Scholar]
- Sibley, D.A. The Sibley Guide to Birds; Knopf: New York, NY, USA, 2000; p. 545. [Google Scholar]
- Hekkala, E.; Shirley, M.H.; Amato, G.; Austin, J.D.; Charter, S.; Thorbjarnarson, J.; Vliet, K.A.; Houck, M.L.; Desalle, R.; Blum, M.J. An ancient icon reveals new mysteries: Mummy DNA resurrects a cryptic species within the Nile crocodile. Mol. Ecol. 2011, 20, 4199–4215. [Google Scholar] [CrossRef] [PubMed]
- Rohland, N.; Reich, D.; Mallick, S.; Meyer, M.; Green, R.E.; Georgiadis, N.J.; Roca, A.L.; Hofreiter, M. Genomic DNA sequences from mastodon and woolly mammoth reveal deep speciation of forest and savanna elephants. PLoS Biol. 2010, 8, e1000564. [Google Scholar] [CrossRef] [PubMed]
- Rosel, P.E.; Wilcox, L.A.; Yamada, T.K.; Mullin, K.D. A new species of baleen whale (Balaenoptera) from the Gulf of Mexico, with a review of its geographic distribution. Mar. Mammal Sci. 2021, 37, 577–610. [Google Scholar] [CrossRef]
- Wada, S.; Oishi, M.; Yamada, T.K. A newly discovered species of living baleen whale. Nature 2003, 426, 278–281. [Google Scholar] [CrossRef]
- Fennessy, J.; Bidon, T.; Reuss, F.; Kumar, V.; Elkan, P.; Nilsson, M.A.; Vamberger, M.; Fritz, U.; Janke, A. Multi-locus analyses reveal four giraffe species instead of one. Curr. Biol. 2016, 26, 2543–2549. [Google Scholar] [CrossRef]
- Schott, R.K.; Evans, D.C.; Goodwin, M.B.; Horner, J.R.; Brown, C.M.; Longrich, N.R. Cranial Ontogeny in Stegoceras validum (Dinosauria: Pachycephalosauria): A Quantitative Model of Pachycephalosaur Dome Growth and Variation. PLoS ONE 2011, 6, e21092. [Google Scholar] [CrossRef]
- Longrich, N.R.; Sankey, J.T.; Tanke, D.H. Texacephale langstoni, a new genus of pachycephalosaurid (Dinosauria: Ornithischia) from the upper Campanian Aguja Formation, southern Texas, USA. Cretac. Res. 2010, 31, 274–284. [Google Scholar] [CrossRef]
- Scannella, J.; Horner, J.R. Torosaurus Marsh, 1891 is Triceratops, Marsh, 1889 (Ceratopsidae: Chasmosaurinae) synonymy through ontogeny. J. Vertebr. Paleontol. 2010, 30, 1157–1168. [Google Scholar] [CrossRef]
- Horner, J.R.; Goodwin, M.B. Extreme cranial ontogeny in the Upper Cretaceous dinosaur Pachycephalosaurus. PLoS ONE 2009, 4, 27626. [Google Scholar] [CrossRef] [PubMed]
- Longrich, N.R. Mojoceratops perifania, a new chasmosaurine ceratopsid from the Late Campanian of Western Canada. J. Paleontol. 2010, 84, 681–694. [Google Scholar] [CrossRef]
- van der Reest, A.J.; Currie, P.J. Troodontids (Theropoda) from the Dinosaur Park Formation, Alberta, with a description of a unique new taxon: Implications for deinonychosaur diversity in North America. Can. J. Earth Sci. 2017, 54, 919–935. [Google Scholar] [CrossRef]
- Darwin, C.R. The Origin of Species; John Murray: London, UK, 1859. [Google Scholar]
- Raup, D.M.; Sepkoski, J.J., Jr. Mass extinctions in the marine fossil record. Science 1982, 215, 1501–1503. [Google Scholar] [CrossRef]


































| Taxon | Publication | Specimen | Status |
|---|---|---|---|
| Ornithomimus grandis | Marsh 1890 [21] | USNM 2110 | Tyrannosaurus indet. |
| Manospondylus gigas | Cope 1892 [22] | AMNH 3982 | Tyrannosaurus indet. |
| Dynamosaurus imperiosus | Osborn 1905 [6] | AMNH 5866/BMNH R7995 | Tyrannosaurus rex |
| Tyrannosaurus rex Albertosaurus megagracilis | Osborn 1905 [6] Paul 1988 [23] | AMNH 973/CM 9380 | Tyrannosaurus rex Tyrannosaurus indet. |
| Dinotyrannus megagracilis | Olshevsky 1995 [24] | Tyrannosaurus indet. | |
| Tyrannosaurus “x” | Larson 2008 [16] | AMNH 5027 | T. rex or T. imperator? |
| Tyrannosaurus imperator | Paul et al. 2022 [18] | FMNH PR 2081 | T. rex or T. imperator? |
| Tyrannosaurus regina | Paul et al. 2022 [18] | USNM 555000 | Tyrannosaurus indet. |
| Taxon | Publication | Specimen | Status |
|---|---|---|---|
| Gorgosaurus lancensis | Gilmore, 1946 [27] | CMNH 7541 | Nanotyrannus lancensis or Tyrannosaurus (?) |
| Albertosaurus lancensis | Paul, 1988 [23] | CMNH 7541 | Nanotyrannus lancensis or Tyrannosaurus (?) |
| Nanotyrannus lancensis | Bakker et al., 1988 [30] | CMNH 7541 | Nanotyrannus lancensis or Tyrannosaurus (?) |
| Aublysodon molnari | Paul, 1988 [23] | LACM 28471 | Nanotyrannus lancensis, Stygivenator molnari, or Tyrannosaurus (?) |
| Stygivenator molnari | Olshevsky, 1995 [24] | LACM 28471 | Nanotyrannus lancensis, Stygivenator molnari, or Tyrannosaurus (?) |
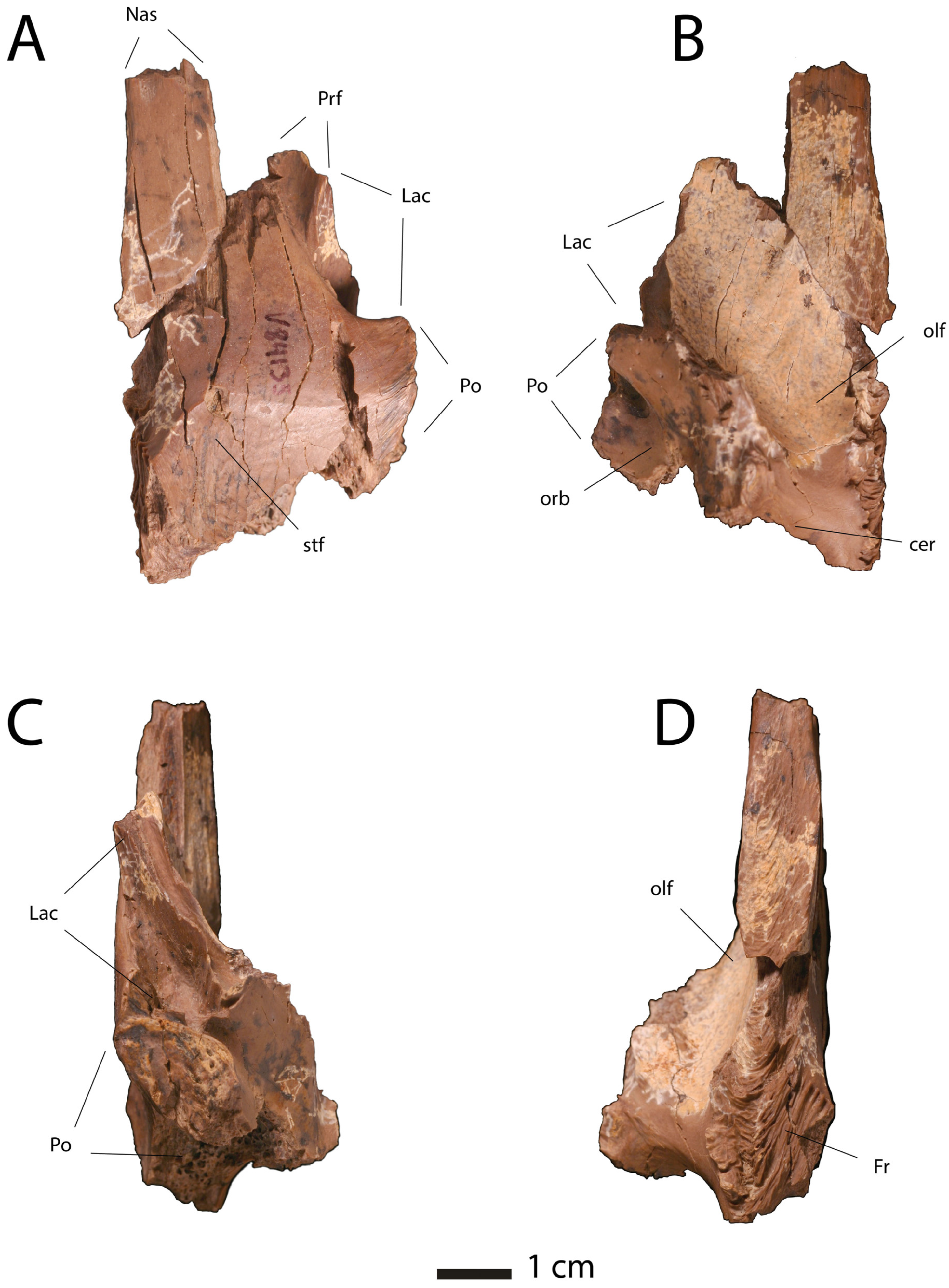
| Premaxilla |
|
| Maxilla |
|
| Nasals |
|
| Lacrimal |
|
| Frontals |
|
| Parietal |
|
| Postorbital |
|
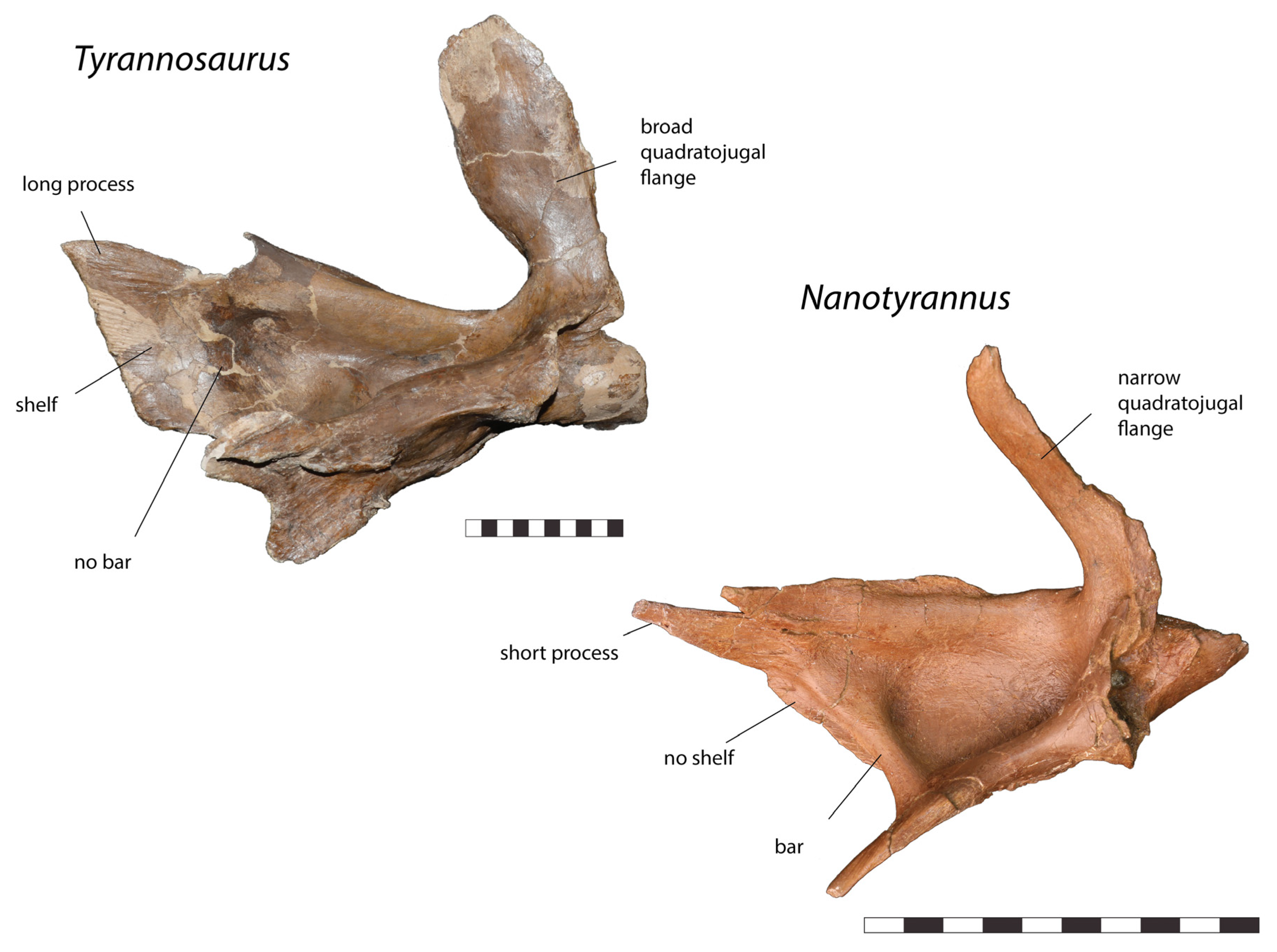
| Jugal |
|
| Quadratojugal |
|
| Squamosals |
|
| Vomer |
|
| Palatine |
|
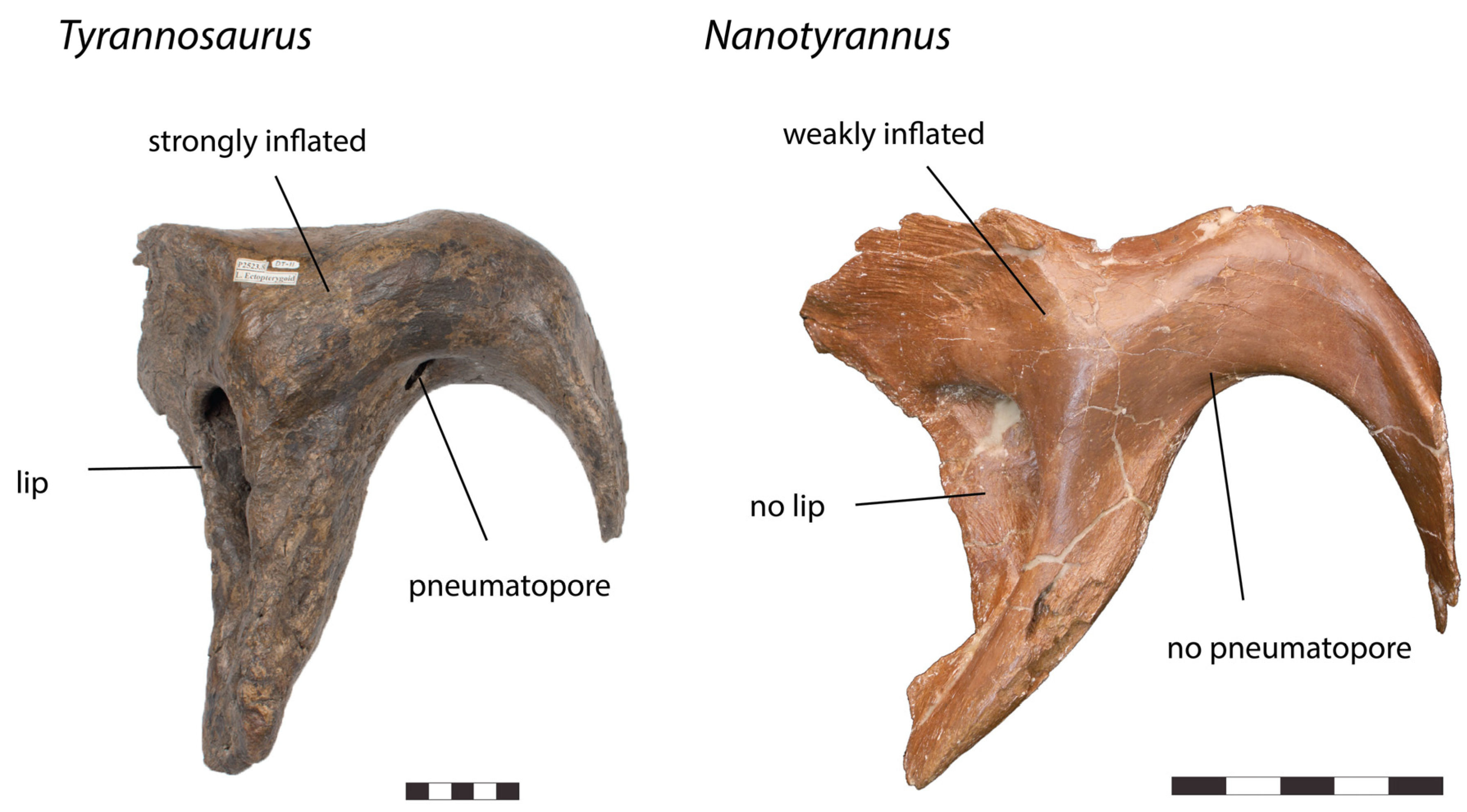
| Ectopterygoid |
|
| Braincase |
|
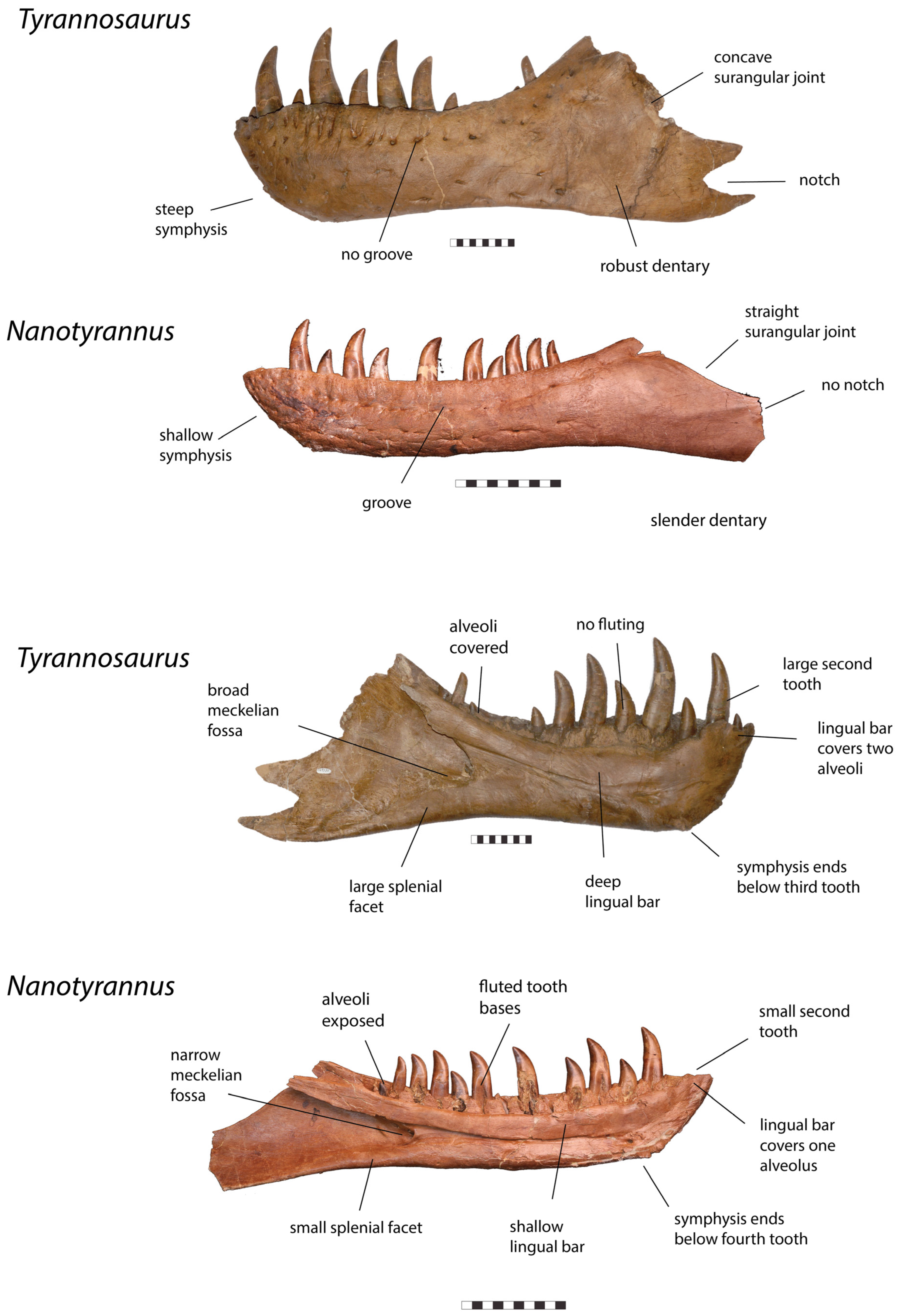
| Dentary |
|
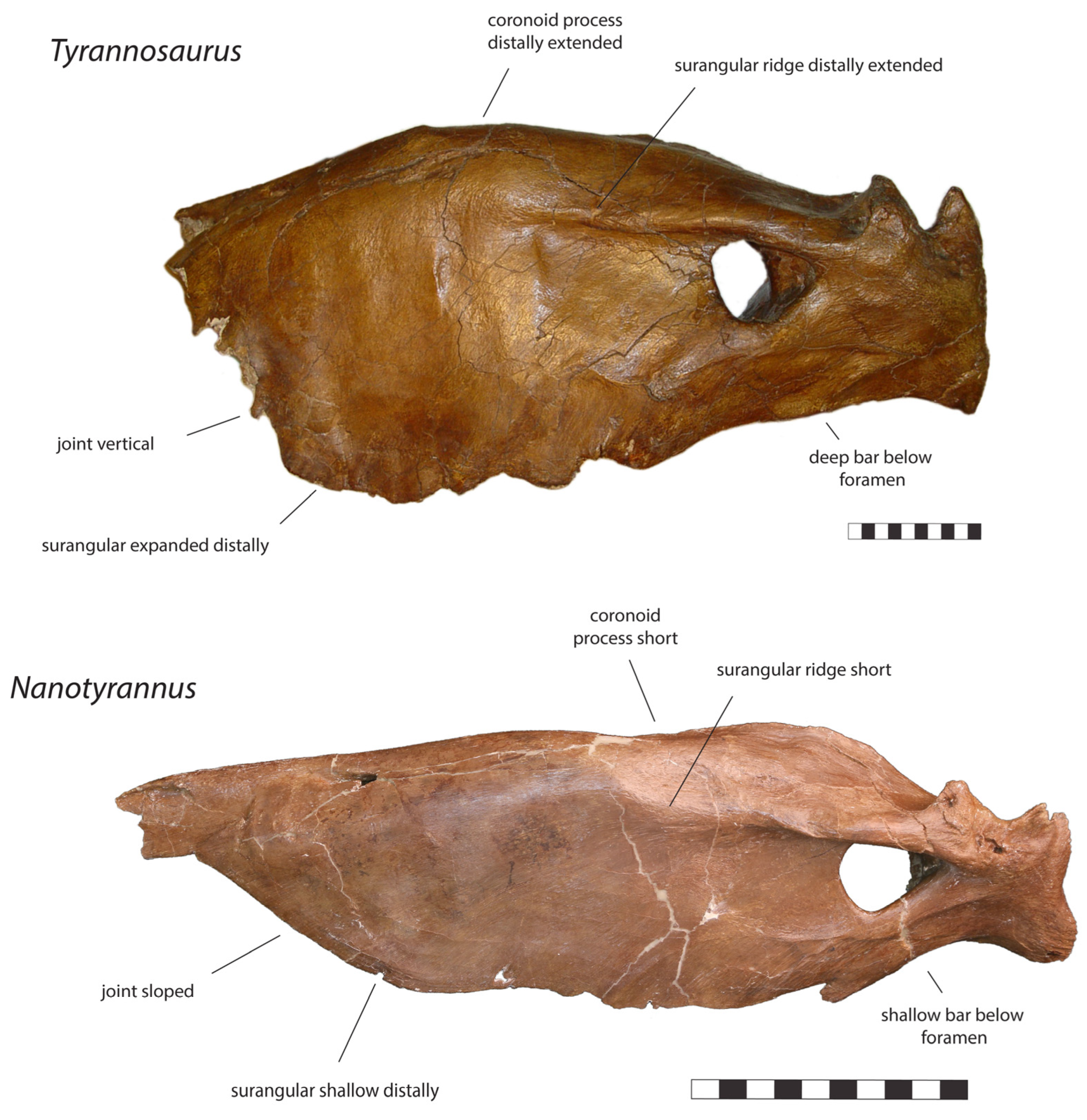
| Posterior Mandible |
|
| Dentition |
|
|
| Body Size |
|
| Specimen | Nickname | Taxon | LAG No. | Circumference (mm) | Mass (kg) (Campione et al., 2014), Equation (7)) | Mass Increase from Previous LAG (kg) |
|---|---|---|---|---|---|---|
| FMNH PR 2081 | Sue | Tyrannosaurus | 1 | 338.7 | 1924 | NA |
| FMNH PR 2081 | Sue | Tyrannosaurus | 2 | 387 | 2777 | 853 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 3 | 415.7 | 3382 | 605 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 4 | 440.3 | 3962 | 580 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 5 | 459.7 | 4461 | 500 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 6 | 485.5 | 5185 | 724 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 7 | 511.1 | 5973 | 788 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 8 | 527.1 | 6503 | 529 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 9 | 542.8 | 7050 | 547 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 10 | 551.6 | 7369 | 319 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 11 | 559.3 | 7656 | 287 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 12 | 564.9 | 7869 | 213 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 13 | 565.7 | 7900 | 31 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 14 | 566.5 | 7931 | 31 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 15 | 567.4 | 7965 | 35 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 16 | 568.2 | 7996 | 31 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 17 | 569 | 8027 | 31 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 18 | 569.8 | 8059 | 31 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 19 | 570.7 | 8094 | 35 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 20 | 571.5 | 8125 | 31 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 21 | 572.3 | 8156 | 31 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 22 | 573.1 | 8188 | 31 |
| FMNH PR 2081 | Sue | Tyrannosaurus | 23 | 574 | 8223 | 35 |
| FMNH PR 2081 | Sue | Tyrannosaurus | Periosteum | 579 | 8422 | 199 |
| BMRP 2006.4.4 | Petey | Nanotyrannus | 1 | 148.4 | 198 | NA |
| BMRP 2006.4.4 | Petey | Nanotyrannus | 2 | 165.9 | 269 | 71 |
| BMRP 2006.4.4 | Petey | Nanotyrannus | 3 | 194.7 | 419 | 149 |
| BMRP 2006.4.4 | Petey | Nanotyrannus | 4 | 214.5 | 547 | 128 |
| BMRP 2006.4.4 | Petey | Nanotyrannus | 5 | 223.6 | 613 | 66 |
| BMRP 2006.4.4 | Petey | Nanotyrannus | 6 | 229.4 | 658 | 45 |
| BMRP 2006.4.4 | Petey | Nanotyrannus | 7 | 234.8 | 701 | 44 |
| BMRP 2006.4.4 | Petey | Nanotyrannus | Periosteum | 244.7 | 786 | 84 |
| BMRP 2002.4.4 | Jane | Nanotyrannus | 1 | 195.9 | 426 | NA |
| BMRP 2002.4.4 | Jane | Nanotyrannus | 2 | 218.4 | 575 | 149 |
| BMRP 2002.4.4 | Jane | Nanotyrannus | 3 | 231.8 | 677 | 102 |
| BMRP 2002.4.4 | Jane | Nanotyrannus | 4 | 247.3 | 809 | 132 |
| BMRP 2002.4.4 | Jane | Nanotyrannus | 5 | 248.6 | 820 | 11 |
| BMRP 2002.4.4 | Jane | Nanotyrannus | 6 | 255.3 | 884 | 63 |
| BMRP 2002.4.4 | Jane | Nanotyrannus | 7 | 257.7 | 906 | 23 |
| BMRP 2002.4.4 | Jane | Nanotyrannus | 8 | 265.7 | 986 | 79 |
| BMRP 2002.4.4 | Jane | Nanotyrannus | 9 | 266.6 | 995 | 9 |
| BMRP 2002.4.4 | Jane | Nanotyrannus | Periosteum | 273.2 | 1064 | 69 |
| BMRP 2002.4.4 | Jane (corrected) | Nanotyrannus | 1 | 195.9 | 426 | NA |
| BMRP 2002.4.4 | Jane (corrected) | Nanotyrannus | 2 | 218.4 | 575 | 149 |
| BMRP 2002.4.4 | Jane (corrected) | Nanotyrannus | 3 | 231.8 | 677 | 102 |
| BMRP 2002.4.4 | Jane (corrected) | Nanotyrannus | 4 | 248.6 | 820 | 144 |
| BMRP 2002.4.4 | Jane (corrected) | Nanotyrannus | 5 | 257.7 | 906 | 86 |
| BMRP 2002.4.4 | Jane (corrected) | Nanotyrannus | 6 | 266.6 | 995 | 88 |
| BMRP 2002.4.4 | Jane (corrected) | Nanotyrannus | Periosteum | 273.2 | 1064 | 69 |
| Specimen | Identification | Formation | Locality | Notes |
|---|---|---|---|---|
| CMNH 7541 | Nanotyrannus lancensis | Hell Creek | Montana | Nanotyrannus lancensis holotype skull; subadult or young adult |
| AMNH 5050 | aff. Nanotyrannus | Partial dentary | ||
| BMRP 2002.4.1 | Nanotyrannus lancensis (or aff Stygivenator?) | Hell Creek | Montana | “Jane”; skull and skeleton, young adult |
| BMRP 2006.4.4 | cf. Nanotyrannus | Hell Creek | Montana | “Petey”; partial post-cranial skeleton young adult |
| FMNH PR 2411 | cf. Nanotyrannus | Hell Creek | South Dakota | lacrimal |
| HRS specimen | Nanotyrannus lancensis (or aff Stygivenator?) | Lance | Wyoming | “Zuri”; partial skull and skeleton, young adult; HRS 08423, 08502, 08607, 08486, 08496, |
| RSM P2347.1 | aff. Nanotyrannus | Frenchman Formation | Saskatchewan | Maxilla |
| KU 155809 | Nanotyrannus lancensis (or aff Stygivenator?) | Hell Creek | Montana | Skull and partial skeleton |
| LACM 28471 | Nanotyrannus lancensis (or Stygivenator molnari) | Hell Creek | Montana | Stygivenator lancensis holotype; partial skull, juvenile |
| MOR 6625 | aff. Nanotyrannus | Partial jaws | ||
| DDM 344.1 | aff. Nanotyrannus | Frontal | ||
| “Dueling Dinosaurs” | Nanotyrannus lancensis (or Stygivenator molnari?) | Hell Creek | Montana | Skull and skeleton (Larson, 2013b) |
| Character | Assessment |
|---|---|
| (1) Nasal processes of the premaxillae tightly appressed throughout their entire length | Not diagnostic of Tyrannosaurus; present in Tarbosaurus, Bistahieversor, Daspletosaurus |
| (2) Restricted exposure of the jugal within the antorbital fenestra | Not diagnostic of Tyrannosaurus; present in Gorgosaurus libratus |
| (3) Antorbital fossa reaches the nasal suture caudodorsally | Not diagnostic of Tyrannosaurus; present in Gorgosaurus libratus, Alioramus altai |
| (4) Transversely broad jugal pneumatic recess | Not shared by Tyrannosaurus and Nanotyrannus; recess broad in Tyrannosaurus, narrow in Nanotyrannus |
| (5) Elongate frontal sagittal crest | Not shared by Tyrannosaurus and Nanotyrannus; frontal sagittal crest short in Nanotyrannus |
| (6) Strongly divergent and short basal tubers | Not shared by Tyrannosaurus and Nanotyrannus; basal tubera more elongate in Nanotyrannus than Tyrannosaurus |
| (7) Rostroventrally-oriented caudal occipital plate | Not shared by Tyrannosaurus and Nanotyrannus; caudally oriented in Tyrannosaurus, faces posteroventrally in Nanotyrannus |
| (8) Shallow subcondylar recess | Not shared by Tyrannosaurus and Nanotyrannus; subcondylar recess well-developed in Nanotyrannus |
| (9) Rostroventrally deep basisphenoid plate and rostrocaudally-restricted basisphenoid recess | Not shared by Tyrannosaurus and Nanotyrannus; anteroposteriorly narrow basispenoid recess in Tyrannosaurus; anteroposteriorly wide basisphenoid recess in Nanotyrannus |
| (10) Inflated ectopterygoid | Not shared by Tyrannosaurus and Nanotyrannus; ectopterygoid weakly inflated in Nanotyrannus |
| (11) Strongly convex rostral plate of the surangular | Not diagnostic of Tyrannosaurus; seen in Daspletosaurus |
| (12) Transversely narrow snout and broad temporal region relative to other tyrannosaurids | Not shared by Tyrannosaurus and Nanotyrannus; snout narrow in Nanotyrannus and broad in Tyrannosaurus; temporal region broader in Tyrannosaurus |
| (13) Deep mandible relative to other tyrannosaurids | Not diagnostic of Tyrannosaurus; seen in Tarbosaurus, Bistahieversor, Daspletosaurus; absent in some specimens of Nanotyrannus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Longrich, N.R.; Saitta, E.T. Taxonomic Status of Nanotyrannus lancensis (Dinosauria: Tyrannosauroidea)—A Distinct Taxon of Small-Bodied Tyrannosaur. Foss. Stud. 2024, 2, 1-65. https://doi.org/10.3390/fossils2010001
Longrich NR, Saitta ET. Taxonomic Status of Nanotyrannus lancensis (Dinosauria: Tyrannosauroidea)—A Distinct Taxon of Small-Bodied Tyrannosaur. Fossil Studies. 2024; 2(1):1-65. https://doi.org/10.3390/fossils2010001
Chicago/Turabian StyleLongrich, Nicholas R., and Evan T. Saitta. 2024. "Taxonomic Status of Nanotyrannus lancensis (Dinosauria: Tyrannosauroidea)—A Distinct Taxon of Small-Bodied Tyrannosaur" Fossil Studies 2, no. 1: 1-65. https://doi.org/10.3390/fossils2010001
APA StyleLongrich, N. R., & Saitta, E. T. (2024). Taxonomic Status of Nanotyrannus lancensis (Dinosauria: Tyrannosauroidea)—A Distinct Taxon of Small-Bodied Tyrannosaur. Fossil Studies, 2(1), 1-65. https://doi.org/10.3390/fossils2010001