Abstract
The birch tree-derived pentacyclic lupine type-triterpenoid Betulinic acid has demonstrated a variety of biological activities BetA is known for its harmlessness on normal healthy cells. However, recent investigations have indicated that BetA can cause cellular changes in mouse normal embryonic fibroblasts even with a minimal concentration. This report cautioned the use of BetA at the clinical level, which encouraged us to examine whether BetA could produce any key effect on normal healthy cells of any organs in mice. The present study extended its investigation to evaluate whether BetA could induce any changes in the renal system and the expression pattern of NADPH-diaphorase an indirect marker of the enzyme nitric oxide synthase in mice. Our results indicated that BetA exposure induced NADPH-d expression in both organs without causing any significant morphological changes. Moreover, NADPH-d activity patterns in the organs of BetA-treated animals tremendously increased (from day 4 until day 12) when compared to controls. The expression of NADPH-d in both the kidney and bladder implies that NADPH-d-mediated nitric oxide signaling could be a mechanism involved in BetA-induced nephroprotection. These outcomes are of direct clinical importance and could pay the way for the improvement of BetA as an important pharmaceutical product.
1. Introduction
The reactive oxygen species (ROS) play a prominent role in human well-being and are beneficial when combating numerous human diseases [1,2]. The following free radicals such as OH, O2− H2O2, O3, HOCI, RO2, and RO produced during various metabolic activities [3]. Numerous urinary diseases including chronic kidney/renal failure (CRF) have been linked c with redox imbalance, but the mechanism responsible remains unknown [4,5]). It is established that NADPH oxidase is the major source of O2− ion formation and superoxide dismutase (SOD) participates in the elimination [5].
Nitric oxide (NO) is a ubiquitous gaseous radical species [6,7] present in the immune [8], and urinary system [9] of various species and endocrine tissues [10,11] among others [12,13,14]. Many studies [15,16] have suggested various contributing factors for chronic renal failure (CRF) like protein nitration, nitric oxide (NO) inactivation, hypertension, and functional NO deficiency. Nitric oxide is a recognized mediator in numerous therapeutic as well as immunomodulatory functions, suggesting its role in immune organs [8,17]. Three different forms of nitric oxide synthases (NOSs) exist they are NOS1, NOS2, and NOS3, which participate in NO and L-citrulline synthesis using cellular oxygen (O2) from the l-arginine amino acid, between this NOS1 and NOS3 is constitutive, whereas NOS2 is inducible one [18]. It is known that all three types of NOSs express NADPH-d enzyme activity of NADPH-d [10,12,14,19], suggesting NADPH-d expression pattern is an indirect presence of the enzyme NOS or NO [17,18]. Since the beginning, NADPH-d considered an indicator of NOS [10].
It is well known that NO is the main homeostatic regulator of renal hemodynamics [20,21] and it has been determined as a key player in the pathogenesis of metabolic diseases [22]. To support this, previous studies have established NO deficiency in numerous renal injury experimental models [14,23]. These animal investigations reveal that abnormalities in endogenous NO production are responsible for renal injury, due to the disparity between the production of eNOS and iNOS [24]. Although NOS2 is known to play a considerable role in the urinary system specifically in the kidney, however, its localization in the kidney is not known. However, [25] demonstrated inducible-NOS-positivity in the rat kidneys using immune- and enzyme histochemistry with NADPH-d, further confirmed by RT-PCR. The results have shown the expression not only in the following cell types like interstitial, and glomerular parietal epithelial cells but also in the proximal part of the short-looped and the upper and middle papillary parts of the long-looped descending thin limb. Besides, some inner medullary collecting duct cells and calyceal and papillary epithelial cells also have been shown.
In line with this, numerous pieces of evidence have supported the role of NO in the pathogenesis and cellular changes in diabetic nephropathy (DN), which is facilitated by an increase/decrease in renal NO production and/or its action [14,26]. Despite the above, many studies have raised the question that which isoform of NOS is responsible for NO generation and which type of cells expresses which form of NOS isoform in the DN [27,28]. It is obvious that as of now there is no effective treatment or preventive mechanisms exist for DN to stop its advancement [29]. However, numerous biomedical data indicate that some herbal extracts have beneficial attribution on certain processes with reduced renal function in DM [29,30].
Many renal ailments including cancer need either immune modulation [31] or protection for disease management and care [32] under such circumstances, the host defense system needs activation, which can offer alternates for the existing chemotherapeutics [33]. In this regard, few potential immunomodulatory agents [34,35] have been isolated from several herbs [2,36] or various protective plant-derived natural products (NPs). NPs are small molecules produced either by medicinal plants, microorganisms, or marine sources [30,37,38]. They play a vital role in pharmaceutical drug development as they exhibit a variety of biological properties and deliver different targets throughout the drug discovery process [39]. As chemicals, NPs contain classes of compounds such as terpenoids, amino acids, polyketides, peptides, lipids, proteins, carbohydrates, nucleic acid bases, deoxyribonucleic acid, and ribonucleic acid [40]. Although all NPs have served to inspire intellectual inquiry, one of the most interesting classes of molecules is the terpenes [41,42].
Terpenoids are groups of hydrocarbons that contain terpenes, which include oxygen-containing groups [43,44]. They are usually found in plants and can form cyclic structures such as sterols [8]. Triterpenes compounds were long considered biological inactive phytocompounds until numerous recent investigations have shown their variety of potential biological properties [45]. Normally, pentacyclic triterpenes’ dry weight accounts for less than 0.1% of the plant [46]. However, the bark of white birch is one of the exceptions as it contains 34% of triterpene pentacyclic betulin (BE) (Figure 1) [47]. Additional results have further demonstrated that it is possible to obtain about 70 to 90% of active betulin compound [48,49,50]. They are also present in various other natural sources, which include animal products and microorganisms [51,52].
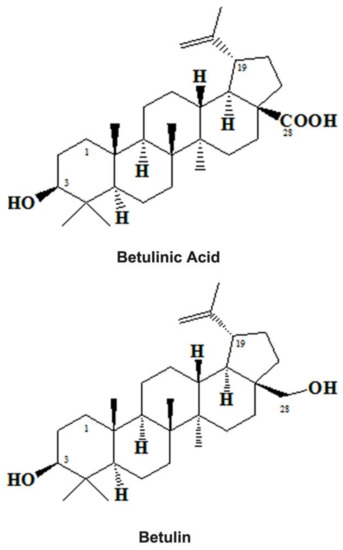
Figure 1.
Chemical structures of Betulinic Acid and Betulin.
The ubiquitous presence of triterpenoids in the environment resulted in a variety of medicinal properties [43]. BetA (3b-hydroxy-lup-20(29)-en-28-oic acid) is one of the pentacyclic lupine forms of triterpenoids (Figure 1) and botulin derivatives (betulin oxidation product) [53]. It is widely found in the outer bark of the birch tree (Betula spp.) [54,55] but also presents significant amounts of free BetA in the underground parts of the swampy plant Menyanthes trifoliata [56]. For decades, BetA has been employed in various traditional practices [57]. Besides, it can be obtained from other plants source also like Sarracenia flava (Sarraceniaceae) [58] trees and shrubs such as Inga punctata (Fabaceae), Diospyros spp. (Ebenaceae) [59], Vauquelinia corymbosa (Rosaceae), Ziziphus spp. (Rhamnaceae) [60] and Syzygium spp. [41,61].
Many studies have demonstrated that both BE and BetA from various sources displayed their potential through different biological activities such as anticancer [62], anti-inflammatory [63], anti-HIV [64], and anti-bacteria [65]. Besides, it has shown further actions like anti-malarial [66], anthelmintic [67], anti-platelet [68], cardioprotective [69], immunomodulation [8], and so on. It targets various parts of cells like mitochondria [70] and enzyme aminopeptidase N [71], topoisomerase [72], acetyl-coA [73], DGAT [74], NF-kB [75], cell cycle [76,77] and proteasome [78].
In line with this, previous investigations [70,79] have recognized the differential effects or selective toxicity of BetA on cancer cells. These results have been identified as a potential cancer inhibitor in many in vitro studies using various human metastatic skin cancer cell lines like MEL-1 to 4 [80] and animal models but this compound spared normal healthy cells. This property is unique and different from the other conventional anticancer agents such as vinblastine, taxol, vincristine, etoposide, and camptothecin as they exhibit very toxic and cause damage to both cancer and normal cells [81]. To support further, no major toxic effects of BetA have been witnessed in rodents [70]. Despite the above fact BetA is still considered a weak anticancer agent because its requirements to control cell growth at the in vitro level is µM concentrations, but higher concentrations (2.5 gm/kg/b.wt) are required to inhibit cancer formation in immunodeficient nude mice l [82].
Despite its poor potency and no toxicity on normal cells, still BetA faces challenges in its clinical usage [70] the reason may be that the existing data are not sufficient to support its action on normal healthy cells. When compared to previous reports [70,80] that 10 µM BetA can induce metabolic changes in normal cells without morphological changes [8,82]. The above concerns especially the use of BetA at the clinical level motivated the present study to examine whether BetA could produce any major effect on healthy tissues and cells, more specifically on the urinary organs of mice.
Therefore, the present study designed to determine whether minimal concentrations (10 to 20 µM) of BetA could induce NO production or cause any changes in kidney and urinary bladder cells. NADPH-d normally considered a marker for nitric oxide synthase done by histochemical approach [83]. Thus, in this study, we have investigated the effect of BetA on the NADPH-d or NOS expression pattern of kidneys and urinary bladders of mice by using NADPH-d staining reaction.
2. Results
Based on our observation, in the normal standard and DMSO-treated vehicle control group, mild to moderately stained NADPH-d cellular structures (mean ± SD of 0.1479 ± 0.0408; p < 0.001) were observed both in the cortico-medullary parts of the kidney (Figure 2a,d,g). While in the GTN group, the cortical radiate arteries as well as a renal pyramid in the medulla region (mean ± SD of 0.2553 ± 0.0780; p < 0.001) showed only moderate NADPH-d expression on day 4 after GTN treatment, that continued day 8th and 12th (Figure 2b,e,h) compared to DMSO control and BetA test group. A similar NADPH-d staining pattern observed in the urinary bladder, however, there is mild or reduced staining in the GTN group, but they showed no cellular changes in the kidney or urinary bladder. This shows that GTN induces a time-dependent fashion of NADPH-d activity in the kidney. Although NADPH-d distribution was like that of control groups at the beginning i.e., on day 4, however, its distribution extends to the entire kidney (both cortex and medulla) for the rest of the treatment periods (i.e., 8th and 12th day of treatment).
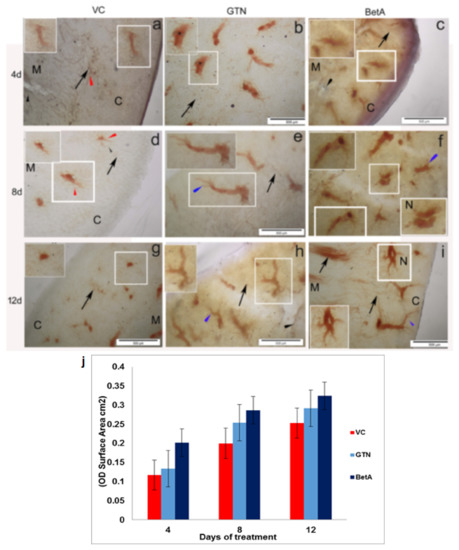
Figure 2.
Representative photomicrographs demonstrating Kidney sections of the (a–g) VC (dimethyl sulfoxide), (b–h) positive (GTN), and (c–i) BetA-treated groups on days 4, 8, and 12 after treatment. The inset is representative of the area in the figure outlined in white. Black arrows identify the NADPH-d-stained medullary rays and black arrow indicates blood vessels, the red arrowhead points to the perivascular nerve fibers and the blue arrowhead demonstrates the NADPH-d positive radiate arteries in the cortex (denoted by the letter C) and medulla (denoted by the letter M). The letter ‘N’ indicates the nephron. Scale bar, 100 µm. (j) The intensity (OD) of the NADPH-d staining was quantified using Olympus Soft Imaging cell Sens software version 1.6. Data expressed as means ± standard deviation of mice (n = 6 per group) p < 0.001 vs. the VC group, p < 0.001 vs. the GTN group. p < 0.01 vs. the VC group. GTN, Goniothalamin; BetA, Betulinic acid; NADPH-d, nicotinamide adenine dinucleotide phosphate diaphorase; VC, vehicle control; OD, optical density.
A similar but gradual steady increase of NADPH-d expression was evident in the cortical radiate arteries and nephron in the kidney cortex as well as in the medullary pyramids (mean ± SD of 0.3237 ± 0.1340; p < 0.001) of the BetA group (Figure 2c,f,i). The NADPH-d activity increased along with the BetA treatment period, which means longer the BetA exposure, the intensity of NADPH-d expression found to increase (Figure 2c,f,i). It is interesting to note that on days 8 (Figure 2f) and 12 (Figure 2i) of BetA treated kidney showed strong NADPH-d staining, however that occurred only in the peripheral cortical radiate arteries (arrow) but not in the medullary arteries as they exhibit only mild staining. It is noteworthy to mention that the cortical area of the kidney, where blood vasculatures are highly oriented, so intense NADPH-d staining in this area (Figure 2i inset) likely to support that NO has a prominent role in vascular physiology [84]. The graphical image in Figure 2j shows the intensity (OD)/staining area (cm2) of the kidney tissue sections obtained from 4, 8, and 12 days of treatment. In brief, increased NADPH-d activity implies that the effect of BetA on the kidney depends on its exposure time.
3. Urinary Bladder
Interestingly, a similar NADPH-d expression pattern was observed in the urinary bladder. Both negative and DMSO-treated vehicle control group animals sacrificed on days 4, 8, and 12 have shown NADPH-d staining in various cellular structures. Structures include neuron-like bodies and blood vessels mostly in the outer larger areas of the bladder; however, the intensity of the staining found extremely low (mean ± SD of 0.0827 ± 0.0035; p < 0.001) (Figure 3a,d,g). The positive-control (GTN) group on day 4 showed less to mild staining (Figure 3b) but from day 8th and 12th after treatment, NADPH-d activity continuously increased (Figure 3e,h). This shows that GTN induces NADPH-d in the urinary bladder in a time-dependent fashion like that of the kidney (mean ± SD of 0.1164 ± 0.1120; p < 0.001).
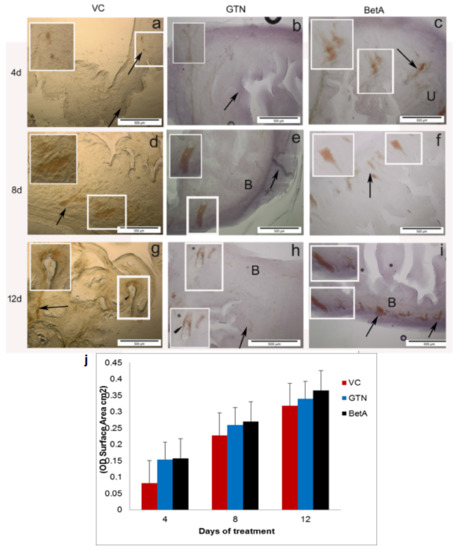
Figure 3.
Representative photomicrographs demonstrating urinary bladder sections of the (a–g) VC (dimethyl sulfoxide), (b–h) positive (GTN), and (c–i) BetA-treated groups on days 4, 8, and 12 after treatment. Inset (IS) is representative of the area in the figure outlined in white. Black arrows identify the NADPH-d-stained neuron-like cell bodies and black arrowhead indicate NADPH-d positive blood vessels lining. ‘B’ indicates the bladder’s outer region. ‘U’ indicates the bladder’s inner region. Scale bar, 100 µm. (j) The intensity (OD) of the NADPH-d staining was quantified using Olympus Soft Imaging cell Sens software version 1.6. Data expressed as means ± standard deviation of mice (n = 6 per group) p < 0.001 vs. the VC group, p < 0.001 vs. the GTN group, p < 0.01 vs. the VC group. GTN, Goniothalamin; BetA, Betulinic acid; NADPH-d, nicotinamide adenine dinucleotide phosphate diaphorase; VC, vehicle control; OD, optical density.
A steady increase of NADPH-d expression and distribution witnessed in every area of the urinary bladder with the higher intensity (p < 0.001) in the test group because its expression increased together with the BetA exposure (Figure 3c,f,i). Like the kidney, the bladder also displayed mild to modest NADPH-d staining 4 days following the treatment (Figure 3c) throughout the bladder but from days 8 (Figure 3f) and 12 (Figure 3i) onwards. BetA-treated bladder showed moderate to stronger NADPH-d staining (mean ± SD of 0.2488 ± 0.1220; p < 0.001). Neither did we find any substantial structural changes in the positive nor in the test agent-treated urinary bladder. The graphical image in (Figure 3j) shows the intensity (OD)/staining area (cm2) of the kidney tissue sections obtained from 4, 8, and 12 days of treatment. In brief, increased NADPH-d activity implies that the effect of BetA on the kidney depends on its exposure time.
4. Discussion
The current study examined the distribution of NADPH-d expression in the GTN and BetA-treated mice kidney and urinary bladder using NADPH-d histochemistry. The results showed varied expression and diverse distribution of NADPH-d in these organs for the entire study period. There are no phenotypical/structural changes have observed both in the kidney as well as in the urinary bladder, however, NADPH-d expression was abundant in the kidney, but less in the urinary bladder. The cortical region of the kidney expressed stronger NADPH-d activity than the medulla region. Although the expression of NADPH-d has seen only after 4 days of BetA treatment, however, significant activity witnessed after 8 and 12 days, suggesting NO production occurred throughout the complete study period. A similar but constant increase of NADPH-d activity also found in the urinary bladder, starting from day 4 may require an interactive outcome of NO in the vascular and other regions of the kidney suggesting it may play a protective role for the nephron [9,85,86]. Although the role of NO in the kidney and urinary bladder not justified, the results of this study corroborated that there is an association between the nephroprotective properties attributed to BetA. The prime effect of BetA-mediated NO engages in the regulation of renal organs [87,88,89] and our results support the statement in the occurrence of NO demonstrated in the renal system.
Moreover, the present study’s histochemical approach revealed the strongest expression for NADPH-d in the renal capsules (comprised of macula densa cells), whereas vascular endothelial showed weaker expression. Our results suggest that renal capsules may produce a substantial quantity of NO. It is probable that in NO-produced target cells, it stimulates soluble guanylate cyclase to generate cGMP from GTP [90,91] and it established that NO/cGMP acts as a vaso-relaxant in the afferent branch of the artery [92]. Previous investigations have also revealed the key outcome of NOS in the efferent branch artery [93]. The obvious discrepancy in inhibition [94] and stimulation [95,96] raises concern about the impact of NO on renin production from granular cells. Since no or limited evidence is available on the potential influence of cGMP on the extra-glomerular mesangial cells, the present results hypothesize and support that NO produced from renal capsules diffuses across the avascular space of the extraglomerular mesangium and may impact the vascular tone of the afferent and/or the efferent arteriole. In addition, NO may enhance renin secretions from granular cells [96].
Since nitric oxide is a gaseous messenger molecule, which has a role in various vital functions including the communication between cells, involved in signaling for vasodilation, neurotransmission, and platelet aggregation inhibition. Physiologically, a constitutive, calcium-dependent isoform of NOS enzymes such as neuronal and endothelial NOS produces a meager quantity of NO for short period, whereas, the inducible isoforms of NOS, which is calcium-independent and require de novo protein synthesis, and produce more amount of NO [97,98,99].
In addition, NO can be found both in the peripheral as well as in the CNS [100] and in other systems like the endocrine [101,102], immune [103], and renal systems [104,105]. NO exhibits numerous physiological roles in the kidney, which include the regulation of renal and glomerular hemodynamics [106] and natriuretic pressure [107,108], and regulation of medullary perfusion [109]. Furthermore, blunting of tubule-glomerular feedback (TGF) [110], inhibition of tubular sodium reabsorption [111], and involvement in renal sympathetic neural activity modulation also involved [108]. Besides the above, a significant deficiency in NO production in response to increased dietary salt intake has been implicated in the pathogenesis of hypertension [112]. Additionally, NO also plays a significant role in tissue injury as a mediator and portrayed NO as a main r causative factor for end-stage renal failure inflammation [113].
Previous studies have also established that NO deficiency is responsible for the formation and development of chronic kidney disease (CKD) and cardiovascular diseases (CVDs) [114]. Since NO acts locally, the exact NO deficiency location dictates the nephron pathology of renal disease, mediated by multiple molecular mechanisms [115,116]. In agreement with this, previous results have demonstrated that the inhibition of induced chronic NOS in vivo experiments caused various complications such as systemic, glomerular hypertension and ischemia, glomerulosclerosis, tubule-interstitial injury, and proteinuria [116,117]. To support the key role of NO deficiency in renal disease formation, various experimental models have also demonstrated that NO depletion by administration of nitrite (NO2) or nitrate (NO3) delays kidney disease development and decreases blood pressure [89,118]. In addition, routine consumption of NO3 from vegetables and fruits may contribute to cardiovascular protection [119].
It is more evident that several chronic kidney diseases (CKDs) require an improved as well as an effective therapeutic option for their management of them [120]. Pentacyclic triterpene is a triterpenoid found in various medicinal plants. Both BE and BetA are birch tree-derived secondary metabolites found in the leaves, stem bark, and fruit peels [121,122]. These NPs are the main important components employed in the oriental as well as in the traditional medicine systems throughout the world [46,123]. These NPs possess and have shown a variety of beneficial attributions against various disease conditions like anticancer [62], anti-inflammatory [63], and anti-microbial including viruses and bacteria [64,65]. Besides, it has shown further actions like anti-malarial [66], anthelmintic [67], anti-platelet [68], cardio-protective [69], immunomodulation [8], and so on. Besides, its additional targets are mitochondria [70] and the enzyme aminopeptidase N [71], topoisomerase [72], acetyl-CoA [73], DGAT [74], NF-kB [75], cell cycle [76] (Chen et al. 2008; Rzeski et al. 2006) and proteasome [78]. The above targets are currently under drug development for pipeline anti-cancer drugs [124]. However, its efficacy against various renal diseases is unknown or not fully explored [125].
On the other hand, previous studies have demonstrated the selective toxicity of BetA on cancer cells [70,79]. Since BetA considered as potential inhibitor for its anti-cancer action on various metastatic skin cancer cell lines (e.g., MEL-1-4) [80] as well as on animal models [126], this compound specifically spares normal healthy cells [70]. This property is unique and different from the other conventional anticancer agents like vinblastine, taxol, vincristine, etoposide, and camptothecin as they exhibit very toxic and cause damage to both cancer and normal cells [124]. To support further, no systemic toxic actions were also observed in rodents [70] using BetA. Despite the above fact BetA is still considered a weak anti-cancer agent as it requires µM concentrations for the inhibition of cell growth, whereas higher concentrations (250 mg/kg b.wt) are required to control skin cancer in an immunodeficient mouse model [82].
Despite its no of toxicity on normal cells, BetA faces hindrances in its clinical applications [70]. The reason may be due to the non-availability of much in vivo data to support its toxic effect on healthy cells. On contrary, previous reports by [82] observed changes in normal cellular metabolism together with morphological changes. A plausible explanation provided by [82] aimed that the reason for the observed morphological changes by BetA in normal cells may be due to its effect on decreasing oxidative capacity, eliciting increased expression of mitochondrial uncoupling proteins 1 and 2 and triggering liver kinase B1-dependent AMPK activation in mouse embryonic fibroblasts. This enzyme activation further led to an increase in glucose uptake and the glycolysis process [127]. Unfortunately, no in vivo or animal data are available to support the above claim. However, the investigation by [8] described that a minimal concentration of BetA can induce NADPH-d expression, without causing any morphological changes. These contrary reports raised concern about the pharmaceutical significance of BetA and encouraged us to examine whether it could produce any impact on urinary structures like the kidney and urinary bladder of mice, especially on the NADPH-d expression.
Recently, NO has been identified as the main mediator of renal hemodynamics and homeostasis [23]. NO deficiency has also been demonstrated in several renal injury animal models, including during cardiopulmonary bypass (CPB) surgery. Evidence from various experimental data proposes that endogenous NO production abnormality linked with renal injury, due to an imbalance in the production of inducible vs endothelial NOS [24]. Amino acid L-arginine-NO signaling is associated with many physiological roles in the kidney and the disruption in this signaling lead to renal injury [20]. Various investigations have revealed that the macula densa of the kidney produces a significant amount of nNOS/NOS1 [95,110,128,129], where NO has been demonstrated as a key player in the TGF response [110]. Besides the above, [130] have reported that the expression of NOS1 in various structures like non-adrenergic, non-cholinergic neurons within the renal arteries of the hilus, arcuate and interlobular arteries, and rarely in the pre-glomerular afferent branches of the artery. To support this, polymerase chain reaction (PCR) on micro-dissected nephron segments has also shown an elevated level of NOS1 RNA in various regions like inner and outer medullary and cortical collecting ducts (IMCD) [131]. Besides the above, NO produced by eNOS/NOS3 has also shown protective nature in renal vasculature. Increased angiotensin II activity is one of the causes of ROS through NADPH-dependent superoxide generation in the damaged kidney [132,133]. In addition, many other oxidases like Xanthine oxidase, cyclooxygenases, and uncoupled NOS are also involved in the generation of oxidative species, however, NADPH oxidase is the most crucial renal oxidase [5].
It is well known that factors like bradykinin (BK), acetylcholine (ACh), and various other endothelium-dependent ones are promoting water and salt loss through the production of cGMP and the formation of NO [134,135]. Previous studies have demonstrated that these factors influence renal blood vessels by resulting in the alteration in glomerular hemodynamics. In the kidney, like choroid plexus and ciliary processes, cGMP alters fluid secretion and stimulates protein phosphorylation in the secretory epithelium [136,137]. In line with this, the present study results also support the possible role of NO and cGMP.
Previous studies have demonstrated that BetA upregulates eNOS but reduces NADPH oxidase induction in human endothelial cells through PKC-independent mechanisms [121,122]. Triterpenoids have the potential to reverse eNOS uncoupling and augment eNOS enzyme activity by phosphorylation of eNOS at serine 1177 and dephosphorylation of eNOS at threonine 495 [138]. However, the effect of BetA on eNOS expression remains unclear [139] so as in the renal organs, hence it needs further investigation.
In line with this, emerging evidence indicates that BetA has shown renal-protective properties [46,70]. Renal fibrosis considered as an end-stage renal failure that progresses from CKD. It is because of the abnormal increase of extracellular matrix (ECM), as a result, it leads to kidney tissue loss and function [140]. However, in the experimental model of CKD, BetA treatment reversed the loss of functions [125] and up-regulate pro-fibrotic proteins like TGF-β, CTGF, hydroxyl proline, type I collagen, and fibronectin. Besides, tubule dilation, degeneration of glomerulus, and vacuolation with the deposition of collagen fibers also attenuated [69,125]. If there is an impact in the renal tissues due to BetA exposure, it predicted that NO might play regulatory roles in these tissues. In addition to the above, BetA exhibits protection against myocardial ischemia-reperfusion injury mouse model by increasing blood flow but reducing oxidative and nitrosative stress [141].
It is well known that NO function as a vasodilator following its release [142,143] from endothelial cells even from the BetA-treated ones [138,144]. The innervation of NO-positive perivascular nerves has been established in numerous vascular tissues [145,146]. The distribution of such neural structures reported also inside the thymus [8,147,148]. Besides, NADPH-d expression also demonstrated in several regions of the mammalian brain [149,150,151]. The present study results agree with the earlier investigation that NADPH-d-positive cells exist in the mammalian kidney including rats and mice [25,136]. Although the present study witnessed only NADPH-d, positive nerve fibers not neuronal body-like structures in the perivascular area of the kidney. It is interesting to note that the reasonable distribution of NO-positive nerves traveling alongside the blood vessels could reflect the role of neuronal NO may be controlling blood flow through both the kidney and urinary bladder. Both NO-positive nerves and blood vessel endothelium may produce NO to influence bloodstream has been reported in the neural system [150] and endocrine organs like the pancreas [152], thyroid [11,12,13], and a series of other organs. These findings allow if NO can participate in neurotransmission in the kidney and urinary bladder. It is interesting to note that apart from blood vessels, NO may also involve in the regulation of regulatory activity of cortical and medullary cells of the kidney by its production in these cells [103]. Thus, BetA may be a promising pharmaceutical or biological response modifier and may reinforce the renal protection of a host. Although the present study employed a simple but most reliable NADPH-diaphorase histochemical method to demonstrate the presence of NO in the renal system, however, authors admit that involving combined NOS indirect immunofluorescence or immunohistochemistry, immunoelectron microscopy techniques along with more advanced techniques like next-generation sequencing or by microarray analysis at the mRNA level could have been adopted to confirm these interesting results obtained in the current study as they may reproduced NO presence and provided more detailed information and new insights to identify more pharmacological targets, which may benefit for future investigations.
5. Materials and Methods
5.1. Experimental Animals and Chemical Requirements
All animal model investigations conducted in this study were under the Universiti Kebangsaan Malaysia (UKM) Animal Ethics Committee (UKMAEC; FF/2020/ALI/20-MAY/685-JUNE-2020) guidelines. Female BALB/c mice (six weeks old) subjected to the study monitored under appropriate conditions. Betulinic acid (Sigma-Aldrich, St. Louis, MO, USA) used as a test (BetA 10 µM) drug, and Goniothalamin (GTN 50 µM) used as a positive control drug obtained from Merck—Life Science, Malaysia, whereas DMSO (Merck, Darmstadt, Germany) (0.05% DMSO) used as vehicle control.
5.2. Animal Treatment and Sample Collection
Animals used in this study broadly divided into four separate groups and each contains 48 animals: (1) test, (2) positive, (3) negative and (4) a normal control groups. The above-categorized mice were further subdivided equally into three subgroups with each having 6 animals following the treatment regimen (4 days, 8 days, and 12 days). The present study followed the protocol described by [12].
5.3. NADPH-Diaphorase Histochemistry and Tissue Morphology Analysis
Cryo-protected tissue sections collected from each group thawed at normal RT for 30 min, washed twice in PBS, and subjected to NADPH-d staining. β-NADPH (Sigma-Aldrich (M) Sdn Bhd, Subang Jaya, Malaysia) used as a substrate with the addition of nitro blue tetrazolium (NBT) (FISHER SCIENTIFIC (M) SDN BHD), a salt that produces an insoluble blue formazan precipitate visible under the light microscope [8,12].
5.4. Statistical Analysis
One-way ANOVA followed by Bonferroni multiple comparison tests (GraphPad PRISM v. 4.0, San Diego, CA, USA) used to perform intensity data analysis. All numerical data expressed as mean ± SEM and the differences considered statistically significant at p < 0.05.
6. Conclusions
In summary, our findings indicate that NADPH-d activity correlates with NOS activity. Although the present study employed a simple but most reliable NADPH-diaphorase histochemistry to demonstrate the presence of NO, still it requires NOS indirect immunofluorescence/immunohistochemistry or immunoelectron microscopy and more advanced techniques like next-generation sequencing or by microarray analysis at the mRNA level to confirm the NO presence. These additional techniques may provide more detailed information and identified new pharmacological targets. However, the present results reveal that BetA treatment induces NADPH-d expression in both the kidney and urinary bladder without producing any substantial morphological changes in these renal structures. The outcome of the present study has direct pharmaceutical importance, which may contribute to the development of new novel drugs to improve the quality of human health and life. Based on the available literature and our understanding of BetA in the renal system, the present results are particularly important as it describes BetA-induced NADPH-d-mediated nitric oxide signaling in the kidney and urinary bladder, which could be the potential molecular mechanism underlying BetA-elicited renal protection in the treated animals.
Author Contributions
M.A.S. inscribed a major part of the manuscript contributed to the guiding and configuring of the manuscript. S.Y.Y. executed most of the bench work and initial data analysis. S.A., C.P., F.M.A. and A.I.A. contributed to the writing of the manuscript in various segments and in preparing figures. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
This animal study obtained ethical approval from The National University of Malaysia (UKM) and already provided the approval number under experimental procedure section (All animal model investigations conducted in this study were under the Universiti Kebangsaan Malaysia (UKM) Animal Ethics Committee (UKMAEC; FF/2020/ALI/20-MAY/685-JUNE-2020) guidelines).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| BetA | Betulinic Acid |
| BE | Betulin |
| NADPH-d | NADPH-diaphorase |
| µM | micro-Molar |
| NO | Nitric Oxide |
| NOS | Nitric Oxide Synthase |
| eNOS | endothelial nitric oxide synthase |
| iNOS | Inducible nitric oxide synthase |
| OH | Hydroxyl radical |
| O2− | Super Oxide |
| H2O2 | Hydrogen Peroxide |
| O3 | Ozone |
| HOCI | Hypochlorous acid |
| RO2 | Alkoxyl radical 2 |
| RO | Alkoxyl radical |
| CR | Chronic Renal |
| CRF | Chronic Renal Failure |
| SOD | Superoxide Dismutase |
| NOS1 | Neuronal nitric oxide synthase |
| NOS3 | Endothelial nitric oxide synthase |
| NOS2 | Inducible nitric oxide synthase |
| DN | Diabetic nephropathy |
| NPs | Natural products |
| HIV | Human Immunodeficiency Virus |
| DGAT | Diglyceride acyltransferase |
| NF-kB | Nuclear factor kappa |
| MEL-1 | Human metastatic Skin Cancer cell line 1 |
| UKM | Universiti Kebangsaan Malaysia |
| UKMAEC | Universiti Kebangsaan Malaysia (UKM) Animal Ethics Committee |
| GTN | Goniothalamin |
| DMSO | Dimethyl sulfoxide |
| ANOVA | Analysis of variance |
| cGMP | Cyclic guanosine monophosphate |
| cGTP | Cyclic guanosine diphosphate |
| CNS | Central Nervous System |
| TGF | Tubule-glomerular feedback |
| CKD | Chronic kidney disease |
| NO2 | Nitrite |
| NO3 | Nitrate |
| b. wt. | body weight |
| AMPK | 5’ AMP-activated protein kinas |
| CPB | Cardiopulmonary bypass |
| PCR | Polymerase chain reaction |
| IMCD | Inner medullary cortical collecting ducts |
| ROS | Reactive Oxygen Species |
| BK | Bradykinin |
| Ach | Acetylcholine |
| PKC | Protein kinase C |
| ECM | Extracellular matrix |
| TGF | Transforming growth factor |
| CTGF | Connective tissue growth factor |
References
- Ali, S.M.; Olivo, M. Nitric oxide mediated photo-induced cell death in human malignant cells. Int. J. Oncol. 2003, 22, 751–756. [Google Scholar] [CrossRef] [PubMed]
- Vijayaraghavan, K.; Rajkumar, J.; Seyed, M.A. Phytochemical screening, free radical scavenging, and antimicrobial potential of Chromolaena odorata leaf extracts against pathogenic bacterium in wound infections—A multispectrum perspective. Biocatal. Agric. Biotech. 2008, 15, 103–112. [Google Scholar] [CrossRef]
- Phaniendra, A.; Babu, D.J.; Latha, P. Free Radicals: Properties, Sources, Targets, and Their Implication in Various Diseases. Indian J. Clin. Biochem. 2015, 30, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Griendling, K.K.; Sorescu, D.; Ushio-Fukai, M. NAD(P)H oxidase: Role in cardiovascular biology and disease. Circ. Res. 2000, 86, 494–501. [Google Scholar] [CrossRef]
- Vaziri, N.D.; Dicus, M.; Ho, N.D.; Boroujerdi-Rad, L.; Sindhu, R.K. Oxidative stress and dysregulation of superoxide dismutase and NADPH oxidase in renal insufficiency. Kidney Int. 2003, 63, 179–185. [Google Scholar] [CrossRef]
- Moncada, S.; Palmer, R.M.G.; Higgs, A. Nitric oxide: Physiology, pathophysiology, and pharmacology. Pharm. Rev. 1991, 43, 109–142. [Google Scholar]
- Knowles, R.G.; Moncada, S. Nitric oxide synthase in mammals. Biochem. J. 1994, 298, 249–258. [Google Scholar] [CrossRef]
- Pang, K.L.; Vijayaraghavan, K.; Badr, A.S.; Ali, S.M. Betulinic acid-induced expression of nicotinamide adenine dinucleotide phosphate-diaphorase in the immune organs of mice: A role of nitric oxide in immunomodulation. Mol. Med. Rep. 2018, 17, 3035–3041. [Google Scholar] [CrossRef]
- Khorashadi, M.; Bokoch, M.P.; Legrand, M. Is nitric oxide the forgotten nephroprotective treatment during cardiac surgery? Ann. Intensive Care 2020, 10, 22. [Google Scholar] [CrossRef]
- Ali, S.M.; Chan, A.S.; Leong, S.K. Localization of nitrergic neuronal and non-neuronal cells in the ultimobranchial glands of the chicken. Anat. Embryol. 1996, 193, 161–168. [Google Scholar] [CrossRef]
- Akbari, Z.; Rohani, M.H.; Behzadi, G. NADPH-d/NOS reactivity in the lumbar dorsal horn of congenitally hypothyroid pups before and after formalin pain induction. Int. J. Dev. Neurosci. 2009, 27, 779–787. [Google Scholar] [CrossRef]
- Syed, M.A.; Leong, S.K.; Chan, A.S. Localization of NADPH-diaphorase reactivity in the chick and mouse thyroid gland. Thyroid 1994, 4, 475–478. [Google Scholar] [CrossRef]
- Gulati, P.; Leong, S.K.; Chan, A.S. Ontogeny of NADPH-d expression in the thymic microenvironment of the chick embryo. Cell Tissue Res. 1998, 294, 335–343. [Google Scholar] [CrossRef]
- Kerschbaum, H.H.; Huang, S.; Xie, M.; Hermann, A. NADPH-diaphorase activity and nitric oxide synthase activity in the kidney of the clawed frog, Xenopus laevis. Cell Tissue Res. 2000, 301, 405–411. [Google Scholar] [CrossRef]
- Vaziri, N.D.; Ni, Z.; Oveisi, F.; Liang, K. Enhanced nitric oxide inacti vation and protein nitration by reactive oxygen species in chronic renal insufficiency. Hypertension 2002, 39, 135–141. [Google Scholar] [CrossRef]
- Deng, G.; Vaziri, N.D.; Jabbari, B.; Yan, X.X. Increased tyrosine nitration of the brain in chronic renal insufficiency: Reversal by antioxidant therapy. J. Am. Soc. Nephrol. 2001, 12, 1892–1898. [Google Scholar] [CrossRef]
- Dorko, F.; Špakovská, T.; Lovasová, K.; Patlevič, P.; Kluchová, D. NADPH-d activity in rat thymus after the application of retinoid acid. Eur. J. Histochem. 2012, 56, e7. [Google Scholar] [CrossRef]
- Förstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart. J. 2012, 33, 829–837. [Google Scholar] [CrossRef]
- Andronowska, A.; Chruściel, M. Expression, and cellular distribution of NADPH-diaphorase and nitric oxide synthases in the porcine uterus during early pregnancy. Folia Histochem. Cytobiol. 2007, 45, 375–380. [Google Scholar]
- Lee, J. Nitric oxide in the kidney: Its physiological role and pathophysiological implications. Electrolyte Blood Press. 2008, 6, 27–34. [Google Scholar] [CrossRef]
- Ahmad, A.; Dempsey, S.K.; Daneva, Z.; Azam, M.; Li, N.; Li, P.L.; Ritter, K.J. Role of Nitric Oxide in the Cardiovascular and Renal Systems. Int. J. Mol. Sci. 2018, 19, 2605. [Google Scholar] [CrossRef] [PubMed]
- Martin, B.; Caron, N.; Jadot, I.; Colombaro, V.; Federici, G.; Depommier, C.; Declèves, A.É. Evaluation of inducible nitric oxide synthase inhibition on kidney function and structure in high-fat diet-induced kidney disease. Exp. Physiol. 2018, 103, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, W.I.C.; de Wit, N.C.J.; Sertorio, J.T.C.; van Bijnen, A.A.; Ganushchak, Y.M.; Heijmans, J.H.; Tanus-Santos, J.E.; Jacobs, M.J.; Maessen, J.G.; Buurman, W.A. Hemolysis during cardiac surgery is associated with increased intravascular nitric oxide consumption and perioperative kidney and intestinal tissue damage. Front. Physiol. 2014, 5, 340. [Google Scholar] [CrossRef] [PubMed]
- Legrand, M.; Mik, E.G.; Johannes, T.; Payen, D.; Ince, C. Renal hypoxia and dysoxia after reperfusion of the ischemic kidney. Mol. Med. 2008, 14, 502–516. [Google Scholar] [CrossRef]
- Choi, J.Y.; Nam, S.A.; Jin, D.C.; Kim, J.; Cha, J.H. Expression, and cellular localization of inducible nitric oxide synthase in lipopolysaccharide-treated rat kidneys. J. Histochem. Cytochem. 2012, 60, 301–315. [Google Scholar] [CrossRef]
- Goligorsky, M.S.; Brodsky, S.V.; Noiri, E. Nitric oxide in acute renal failure: NOS versus NOS. Kidney Int. 2002, 61, 855–861. [Google Scholar] [CrossRef]
- Bank, N.; Aynedjian, H.S. Role of EDRF (nitric oxide) in diabetic renal hyperfiltration. Kidney. Int. 1993, 43, 1306–1312. [Google Scholar] [CrossRef]
- Komers, R.; Allen, T.J.; Cooper, M.E. Role of endothelium-derived nitric oxide in the pathogenesis of the renal hemodynamic changes of experimental diabetes. Diabetes 1993, 43, 1190–1197. [Google Scholar] [CrossRef]
- Musabayane, C.T. The effects of medicinal plants on renal function and blood pressure in diabetes mellitus. Cardiovasc. J. Afr. 2012, 23, 462–468. [Google Scholar] [CrossRef]
- Seyed, M.A.; Ayesha, S. Modern phytomedicine in treating Diabetic foot ulcer: Progress and opportunities. In Diabetic Foot Ulcer; Zubair, M., Ahmad, J., Malik, A., Talluri, M.R., Eds.; Springer: Singapore, 2021. [Google Scholar] [CrossRef]
- Disis, M.L. Immune regulation of cancer. J. Clin. Oncol. 2010, 28, 4531–4538. [Google Scholar] [CrossRef]
- Geetha, S.; Singh, V.; Ram, M.S.; Ilavazhagan, G.; Banerjee, P.K.; Sawhney, R.C. Immunomodulatory effects of seabuckthorn (Hippophae rhamnoides L.) against chromium (VI) induced immunosuppression. Mol. Cell. Biochem. 2005, 278, 101–109. [Google Scholar] [CrossRef]
- K Aravindaram, N.S. Anti-inflammatory plant natural products for cancer therapy. Planta Med. 2010, 76, 1103–1117. [Google Scholar] [CrossRef]
- Winkler, C.; Wirleitner, B.; Schroecksnadel, K.; Schennach, H.; Mur, E.; Fuchs, D. In vitro effects of two extracts and two pure alkaloid preparations of Uncaria tomentosa on peripheral blood mononuclear cells. Planta Med. 2004, 70, 205–210. [Google Scholar]
- B Patwardhan, M.G. Botanical immunodrugs: Scope and opportunities. Drug Discov. Today 2005, 10, 495–502. [Google Scholar] [CrossRef]
- Oveissi, V.; Ram, M.; Bahramsoltani, R.; Ebrahimi, F.; Rahimi, R.; Naseri, R.; Belwal, T.; Devkota, H.P.; Abbasabadi, Z.; Farzaei, M.H. Medicinal plants and their isolated phytochemicals for the management of chemotherapy-induced neuropathy: Therapeutic targets and clinical perspective. Daru 2019, 27, 389–406. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Quinn, M.T. Botanical polysaccharides: Macrophage immunomodulation and therapeutic potential. Int. Immunopharmacol. 2006, 6, 317–333. [Google Scholar] [CrossRef]
- Seyed, M.A.; Siddiqua, A. Calotropis—A multi-potential plant to humankind: Special focus on its wound healing efficacy. Biocatal. Agric. Biotechnol. 2020, 28, 101725. [Google Scholar] [CrossRef]
- Lahlou, M. The Success of Natural Products in Drug Discovery. Pharmacol. Pharm. 2013, 4, 7–31. [Google Scholar] [CrossRef]
- Bruce, J. The role of natural products in evolution. In Recent Advances in Phytochemisty; Elsevier: Oxford, UK; Volume 34, pp. 1–24. 2022. [Google Scholar]
- Yogeeswari, P.; Sriram, D. Betulinic acid and its derivatives: A review on their biological properties. Curr. Med. Chem. 2005, 12, 657–666. [Google Scholar] [CrossRef]
- McChesney, J.D.; Venkataraman, S.K.; Henri, J.T. Plant natural products: Back to the future or into extinction? Phytochemistry 2007, 68, 2015–2022. [Google Scholar] [CrossRef]
- James, J.T.; Dubery, I.A. Pentacyclic Triterpenoids from the Medicinal Herb, Centella asiatica (L.) Urban. Molecules 2009, 14, 3922–3941. [Google Scholar] [CrossRef] [PubMed]
- Perveen, S. Introductory chapter: Terpenes and terpenoids. In Terpenes and Terpenoids; Perveen, S., Al-Taweel, A., Eds.; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef]
- Hordyjewska, A.; Ostapiuk, A.; Horecka, A.; Kurzepa, J. Betulin and betulinic acid: Triterpenoids derivatives with a powerful biological potential. Phytochem. Rev. 2019, 18, 929–951. [Google Scholar] [CrossRef]
- Jäger, S.; Trojan, H.; Kopp, T.; Laszczyk, M.N.; Scheffler, A. Pentacyclic triterpene distribution in various plants—Rich sources for a new group of multi-potent plant extracts. Molecules 2009, 14, 2016–2631. [Google Scholar] [CrossRef]
- Laszczyk, M.N. Pentacyclic triterpenes of the lupane, oleanane and ursane group as tools in cancer therapy. Planta Med. 2009, 75, 1549–1560. [Google Scholar] [CrossRef] [PubMed]
- Dehelean, C.A.; Feflea, S.; Molnár, J.; Zupko, I.; Soica, C. Betulin as an antitumor agent tested in vitro on A431, HeLa and MCF7, and as an angiogenic inhibitor in vivo in the CAM assay. Nat. Prod. Commun. 2012, 7, 981–985. [Google Scholar] [PubMed]
- Dehelean, C.A.; Soica, C.; Ledeţi, I.; Aluaş, M.; Zupko, I.G.; Luşcan, A.; Cinta-Pinzaru, S.; Munteanu, M. Study of the betulin enriched birch bark extracts effects on human carcinoma cells and ear inflammation. Chem. Cent. J. 2012, 6, 137. [Google Scholar] [CrossRef]
- Kovalenko, L.P.; Balakshin, V.V.; Presnova, G.A.; Chistyakov, A.N.; Shipaeva, E.V.; Alekseeva, S.V.; Durnev, A.D. Immunotoxicity and allergenic properties of betulin-containing birch bark dry extract. Pharm. Chem. J. 2007, 41, 17–19. [Google Scholar] [CrossRef]
- Silchenko, A.S.; Kalinovsky, A.I.; Avilov, S.A.; Andryjaschenko, P.V.; Dmitrenok, P.S.; Martyyas, E.A.; Kalinin, V.I. Triterpene Glycosides from the Sea Cucumber Eupentacta fraudatrix. Structure and Biological Action of Cucumariosides A1, A3, A4, A5, A6, A12 and A15, Seven New Minor Non-sulfated Tetraosides and Unprecedented 25-Keto, 27-Norholostane Aglycone. Nat. Prod. Commun. 2012, 7, 517–525. [Google Scholar]
- Ragasa, C.Y.; Cornelio, K.B. Triterpenes from Euphorbia hirta and their cytotoxicity. Chin. J. Nat. Med. 2013, 11, 528–533. [Google Scholar] [CrossRef]
- Moghaddam, M.G.; Ahmad, F.B.H.; Kermani, A.S. Biological Activity of Betulinic Acid: A Review. Pharmacol. Pharm. 2012, 3, 119–123. [Google Scholar] [CrossRef]
- Drag-Zalesinska, M.; Kulbacka, J.; Saczko, J.; Wysocka, T.; Zabel, M.; Surowiak, P.; Drag, M. Esters of betulin and betulinic acid with amino acids have improved water solubility and are selectively cytotoxic toward cancer cells. Bioorg. Med. Chem. Lett. 2009, 19, 4814–4817. [Google Scholar] [CrossRef]
- Drag, M.; Surowiak, P.; Drag-Zalesinska, M.; Dietel, M.; Lage, H.; Oleksyszy, J. Comparision of the cytotoxic effects of birch bark extract, betulin and betulinic acid towards human gastric carcinoma and pancreatic carcinoma drug-sensitive and drug-resistant cell lines. Molecules 2009, 14, 1639–1651. [Google Scholar] [CrossRef]
- Zdzisińska, B.; Rzeski, W.; Paduch, R.; Szuster-Ciesielska, A.; Kaczor, J.; Wejksza, K.; Kandefer-Szerszeń, M. Differentialeffect of betulin and betulinic acid on cytokine productionin human whole blood cell cultures. Pol. J. Pharmacol. 2003, 55, 235–238. [Google Scholar]
- Tsai, J.C.; Peng, W.H.; Chiu, T.H.; Lai, S.C.; Lee, C.Y. Anti-inflammatory Effects of Scoparia dulcis L. and Betulinic Acid. Am. J. Chin. Med. 2011, 39, 943–956. [Google Scholar] [CrossRef]
- Bhattacharyya, J.; Kokpol, U.; Miles, D.H. The isolation from Sarracenia flava and partial synthesis of betulinaldehyde. Phytochemistry 1976, 15, 432–433. [Google Scholar] [CrossRef]
- Kingston, D.G.; Munjal, R.C. Plant anticancer agents. VIII. Constituents of Inga punctata. Lloydia 1978, 41, 499–500. [Google Scholar]
- Trumbull, E.R.; Bianchi, E.; Wiedhopf, D.J.E.R.M.; Cole, J.R. Tumor Inhibitory Agents from Vauquelinia corymbosa Rosaceae. J. Pharm. Sci. 1976, 65, 1407–1408. [Google Scholar] [CrossRef]
- Eiznhamer, D.A.; Xu, Z.Q. Betulinic acid: A promising anticancer candidate. IDrugs 2004, 7, 359–373. [Google Scholar]
- Patlolla, J.M.; Rao, C.V. Triterpenoids for cancer prevention and treatment: Status and future prospects. Curr. Pharm. Biotechnol. 2012, 13, 147–155. [Google Scholar] [CrossRef]
- Costa, J.F.; Barbosa-Filho, J.M.; Maia, G.L.; Guimarães, E.T.; Meira, C.S.; Ribeiro-dos-Santos, R.; de Carvalho, L.C.; Soares, M.B. Potent anti-inflammatory activity of betulinic acid treatment in a model of lethal endotoxemia. Int. Immunopharmacol. 2014, 23, 469–474. [Google Scholar] [CrossRef]
- Fujioka, T.; Kashiwada, Y.; Kilkuskie, R.E.; Cosentino, L.M.; Ballas, L.M.; Jiang, J.B.; Janzen, W.B.; Chen, I.S.; Lee, K.H. Anti-AIDS Agents, 11. Betulinic Acid and Platanic Acid as Anti-HIV Principles from Syzigium claviflorum, and the Anti-HIV Activity of Structurally Related Triterpenoids. J. Nat. Prod. 1994, 57, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Chandramu, C.; Manohar, R.D.; Krupadanam, D.G.; Dashavantha, R.V. Isolation, characterization, and biological activity of betulinic acid and ursolic acid from Vitex negundo L. Phytother. Res. 2003, 17, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Steele, J.C.; Warhurst, D.C.; Kirby, G.C.; Simmonds, M.S. In vitro and in vivo evaluation of betulinic acid as an antimalarial. Phytother. Res. 1999, 13, 115–119. [Google Scholar] [CrossRef]
- Enwerem, N.M.; Okogun, J.I.; Wambebe, C.O.; Okorie, D.A.; Akah, P.A. Anthelmintic activity of the stem bark extracts of Berlina grandiflora and one of its active principles, Betulinic acid. Phytomedicine 2001, 8, 112–114. [Google Scholar] [CrossRef] [PubMed]
- Tzakos, A.G.; Kontogianni, V.G.; Tsoumani, M.; Kyriakou, E.; Hwa, J.; Rodrigues, F.A.; Tselepis, A.D. Exploration of the Antiplatelet Activity Profile of Betulinic Acid on Human Platelets. J. Agric. Food Chem. 2012, 60, 6977–6983. [Google Scholar] [CrossRef]
- Xia, A.; Xue, Z.; Li, Y.; Wang, W.; Xia, J.; Wei, T.; Cao, J.; Zhou, W. Cardioprotective effect of betulinic Acid on myocardial ischemia reperfusion injury in rats. Evid. Based Complem. Altern. Med. 2014, 2014, 573745. [Google Scholar] [CrossRef]
- Mullauer, F.B.; Kessler, J.H.; Medema, J.P. Betulinic Acid, a Natural Compound with Potent Anticancer Effects. Anticancer Drug 2010, 21, 215–227. [Google Scholar] [CrossRef]
- Melzig, M.F.; Bormann, H. Betulinic acid inhibits aminopeptidase N activity. Planta Med. 1998, 64, 655–657. [Google Scholar] [CrossRef]
- Chowdhury, A.R.; Mandal, S.; Mittra, B.; Sharma, S.; Mukhopadhyay, S.; Majumder, H.K. Betulinic acid, a potent inhibitor of eukaryotic topoisomerase I: Identification of the inhibitory step, the major functional group responsible and development of more potent derivatives. Med. Sci. Monit. 2002, 8, BR254–BR265. [Google Scholar]
- Lee, S.H.; Stephens, J.L.; Paul, K.S.; Englund, P.T. Fatty acid synthesis by elongases in trypanosomes. Cell 2006, 126, 691–699. [Google Scholar] [CrossRef]
- De Melo, C.L.; Queiroz, M.G.R.; Filho, A.C.V.A.; Rodrigues, A.M.; de Sousa, D.F.; Almeida, J.G.L.; Pessoa, O.D.L.; Silveira, E.R.; Menezes, D.B.; Melo, T.S.; et al. Betulinic Acid, a Natural Pentacyclic Triterpenoid, Prevents Abdominal Fat Accumulation in Mice Fed a High-Fat Die. J. Agric. Food Chem. 2009, 57, 8776–8781. [Google Scholar] [CrossRef]
- Takada, Y.; Mukhopadhyay, A.; Kundu, G.C.; Mahabeleshwar, G.H.; Singh, S.; Aggarwal, B.B. Hydrogen peroxide activates NF-kappa B through tyrosine phosphorylation of I kappa B alpha and serine phosphorylation of p65: Evidence for the involvement of I kappa B alpha kinase and Syk protein-tyrosine kinase. J. Biol. Chem. 2003, 278, 24233–24241. [Google Scholar] [CrossRef]
- Chen, X.; Xu, H.; Yuan, P.; Fang, F.; Huss, M.; Vega, V.B.; Wong, F.; Orlov, Y.L.; Zhang, W.; Jiang, J.; et al. Integration of external signaling pathways with the core transcriptional network in embryonic stem cells. Cell 2008, 133, 1106–1117. [Google Scholar] [CrossRef]
- Rzeski, W.; Stepulak, A.; Szymański, M.; Sifringer, M.; Kaczor, J.; Wejksza, K.; Zdzisińska, B.; Kandefer-Szerszeń, M. Betulinic acid decreases expression of bcl-2 and cyclin D1, inhibits proliferation, migration and induces apoptosis in cancer cells. Naunyn Schmiedebergs Arch. Pharm. 2006, 374, 11–20. [Google Scholar] [CrossRef]
- Huang, L.; Ho, P.; Chen, C.H. Activation, and inhibition of the proteasome by betulinic acid and its derivatives. FEBS Lett. 2007, 581, 4955–4959. [Google Scholar] [CrossRef]
- Zuco, V.; Supino, R.; Righetti, S.C.; Cleris, L.; Marchesi, E.; Gambacorti-Passerini, C.; Formelli, F. Selective cytotoxicity of betulinic acid on tumor cell lines, but not on normal cells. Cancer Lett. 2002, 175, 17–25. [Google Scholar] [CrossRef]
- Pisha, E.; Chai, H.; Lee, I.S.; Chagwedera, T.E.; Farnsworth, N.R.; Cordell, G.A.; Beecher, C.W.W.; Fong, H.H.S.; Kinghorn, A.D.; Brown, D.M.; et al. Discovery of betulinic acid as a selective inhibitor of human melanoma that functions by induction of apoptosis. Nat. Med. 1995, 1, 1046–1051. [Google Scholar] [CrossRef]
- Desai, A.G.; Qazi, G.N.; Ganju, R.K.; El-Tamer, M.; Singh, J.; Saxena, A.K.; Bedi, Y.S.; Taneja, S.C.; Bhat, H.K. Medicinal plants and cancer chemoprevention. Curr. Drug Metab. 2008, 9, 581–591. [Google Scholar] [CrossRef]
- Heiss, E.H.; Kramer, M.P.; Atanasov, A.G.; Beres, H.; Schachner, D.; Dirsch, V.M. Glycolytic switch in response to betulinic acid in non-cancer cells. PLoS ONE 2014, 9, e115683. [Google Scholar] [CrossRef]
- Hope, B.T.; Michael, G.J.; Knigge, K.M.; Vincent, S.R. Neuronal NADPH diaphorase is a nitric oxide synthase. Proc. Natl. Acad. Sci. USA 1991, 88, 2811–2814. [Google Scholar] [CrossRef]
- Moncada, S.; Higgs, E.A. The discovery of nitric oxide and its role in vascular biology. Br. J. Pharm. 2006, 147 (Suppl. S1), S193–S201. [Google Scholar] [CrossRef] [PubMed]
- Pramod, K.; Devala, R.G.; Lakshmayya; Ramachandra, S.S. Nephroprotective and Nitric Oxide Scavenging Activity of Tubers of Momordica tuberosa in Rats. Avicenna J. Med. Biotechnol. 2011, 3, 87–93. [Google Scholar] [PubMed]
- Wang, P.; Li, Q.; Li, K.; Zhang, X.; Han, Z.; Wang, J.; Gao, D.; Li, J. Betulinic acid exerts immunoregulation and anti-tumor effect on cervical carcinoma (U14) tumor-bearing mice. Pharmazie 2012, 67, 733–739. [Google Scholar] [PubMed]
- Herrera, M.; Garvin, J.L. Recent Advances in the Regulation of Nitric Oxide in the Kidney. Hypertension 2005, 45, 1062–1067. [Google Scholar] [CrossRef]
- Ahmed, A.; Campbell, R.C. Epidemiology of chronic kidney disease in heart failure. Heart Fail. Clin. 2008, 4, 387–399. [Google Scholar] [CrossRef]
- Carlström, M.; Persson, A.E.; Larsson, E.; Hezel, M.; Scheffer, P.G.; Teerlink, T.; Weitzberg, E.; Lundberg, J.O. Dietary nitrate attenuates oxidative stress, prevents cardiac and renal injuries, and reduces blood pressure in salt-induced hypertension. Cardiovasc. Res. 2011, 89, 574–585. [Google Scholar] [CrossRef]
- Derbyshire, E.R.; Marletta, M.A. Biochemistry of soluble guanylate cyclase. Handb. Exp. Pharm. 2009, 191, 17–31. [Google Scholar]
- Stasch, J.P.; Pacher, P.; Evgenov, O.V. Soluble guanylate cyclase as an emerging therapeutic target in cardiopulmonary disease. Circulation 2011, 123, 2263–2273. [Google Scholar] [CrossRef]
- Sausbier, M.; Schubert, R.; Voigt, V.; Hirneiss, C.; Pfeifer, A.; Korth, M.; Kleppisch, T.; Ruth, P.; Hofmann, F. Mechanisms of NO/cGMP-dependent vasorelaxation. Circ. Res. 2009, 87, 825–830. [Google Scholar] [CrossRef]
- Dalal, R.; Bruss, Z.S.; Sehdev, J.S. Physiology, renal blood flow and filtration. In StatPearls; Updated 26 July 2021; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK482248/ (accessed on 25 July 2022).
- Kurtz, A.; Eckardt, K.U.; Neumann, R.; Kaissling, B.; le Hir, M.; Bauer, C. Site of erythropoietin formation. Contrib. Nephrol. 1989, 76, 14–23. [Google Scholar]
- Mundel, P.; Bachmann, S.; Bader, M.; Fischer, A.; Kummer, W.; Mayer, B.; Kriz, W. Expression of nitric oxide synthase in kidney macula densa cells. Kidney Int. 1992, 42, 1017–1019. [Google Scholar] [CrossRef]
- Münter, K.; Hackenthal, E. The effects of endothelin on renovascular resistance and renin release. J. Hypertens. 1989, 7, S276–S277. [Google Scholar] [CrossRef]
- Szabo, C. Alterations in nitric oxide production in various forms of circulatory shock. New Horiz. 1995, 3, 2–32. [Google Scholar]
- Mulligan, M.S.; Hovel, J.M.; Marletta, M.A.; Ward, P.A. Tissue injury caused by deposition of immune complexes is L-arginine dependent. Proc. Natl. Acad. Sci. USA 1991, 88, 6338–6342. [Google Scholar] [CrossRef]
- Kolb, H.; Kolb-Bachofen, V. Nitric oxide: A pathogenetic factor in autoimmunity. Immunol. Today 1992, 13, 157–160. [Google Scholar] [CrossRef]
- Chachlaki, K.; Prevot, V. Nitric oxide signalling in the brain and its control of bodily functions. Br. J. Pharm. 2020, 177, 5437–5458. [Google Scholar] [CrossRef]
- Ali, S.M.; Chan, A.S.; Leong, S.K. Histochemical and immunohistochemical localisation of nitrergic neuronal and non-neuronal cells in the bursa of Fabricius of the chicken. Cell Tissue Res. 1996, 285, 273–279. [Google Scholar] [CrossRef]
- Clark, K.E.; Myatt, L. Uterine effects of nitric oxide. In Nitric Oxide and the Regulation of the Peripheral Circulation; Nitric Oxide in Biology and Medicine; Kadowitz, P.J., McNamara, D.B., Eds.; Birkhäuser: Boston, MA, USA, 2000; Volume 1. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric oxide synthase in innate and adaptive immunity: An update. Trends Immunol. 2015, 36, 161–178. [Google Scholar] [CrossRef]
- Birder, L.A.; Nakamura, Y.; Kiss, S.; Nealen, M.L.; Barrick, S.; Kanai, A.J.; Wang, E.; Ruiz, G.; de Groat, W.C.; Apodaca, G.; et al. Altered urinary bladder function in mice lacking the vanilloid receptor TRPV1. Nat. Neurosci. 2002, 5, 856–860. [Google Scholar] [CrossRef]
- Studeny, S.; Cheppudira, B.P.; Meyers, S.; Balestreire, E.M.; Apodaca, G.; Birder, L.A.; Braas, K.M.; Waschek, J.A.; May, V.; Vizzard, M.A. Urinary bladder function and somatic sensitivity in vasoactive intestinal polypeptide (VIP)−/− mice. J. Mol. Neurosci. 2008, 36, 175–187. [Google Scholar] [CrossRef]
- Majid, D.S.; Navar, L.G. Nitric oxide in the control of renal hemodynamics and excretory function. Am. J. Hypertens. 2001, 146 Pt 2, 74S–82S. [Google Scholar] [CrossRef] [PubMed]
- Majid, D.S.; Williams, D.; Navar, L.G. Inhibition of nitric oxide synthesis attenuates pressure-induced natriuretic responses in anesthetized dogs. Am. J. Physiol. Ren. Physiol. 1993, 264, F79–F87. [Google Scholar] [CrossRef] [PubMed]
- Eppel, G.A.; Bergstrom, G.; Anderson, W.P.; Evans, R.G. Autoregulation of renal medullary blood flow in rabbits. Am. J. Physiol. Regul. Int. Reg. Comp. Physiol. 2003, 284, R233–R244. [Google Scholar] [CrossRef] [PubMed]
- Mattson, D.L.; Wu, F. Nitric Oxide Synthase Activity and Isoforms in Rat Renal Vasculature. Hypertension 2000, 35, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, C.S. Role of macula densa NOS in tubuloglomerular feedback. Curr. Opin. Nephrol. Hypertens. 1998, 7, 443–449. [Google Scholar]
- Ortiz, P.A.; Garvin, J.L. Superoxide stimulates NaCl absorption by the thick ascending limb. Am. J. Physiol. Ren. Physiol. 2002, 283, F957–F962. [Google Scholar] [CrossRef]
- Pallone, T.L.; Mattson, D.L. Role of nitric oxide in regulation of the renal medulla in normal and hypertensive kidneys. Curr. Opin. Nephrol. Hypertens. 2002, 11, 93–98. [Google Scholar] [CrossRef]
- Locatelli, F.; Canaud, B.; Eckardt, E.U.; Stenvinkel, P.; Wanner, C.; Zoccali, C. Oxidative stress in end-stage renal disease: An emerging threat to patient outcome. Nephrol. Dial. Transpl. 2003, 18, 1272–1280. [Google Scholar] [CrossRef]
- Reddy, Y.S.; Kiranmayi, V.S.; Bitla, A.R.; Krishna, G.S.; Rao, P.V.L.N.S.; Sivakumar, V. Nitric oxide status in patients with chronic kidney disease. Indian J. Nephrol. 2015, 25, 287–291. [Google Scholar]
- Martens, C.R.; Edwards, D.G. Peripheral vascular dysfunction in chronic kidney disease. Cardiol. Res. Pract. 2011, 2011, 257–267. [Google Scholar] [CrossRef]
- Zatz, R.; Baylis, C. Chronic nitric oxide inhibition model six years on. Hypertension 1998, 32, 958–964. [Google Scholar] [CrossRef]
- Baylis, C. Nitric oxide synthase derangements and hypertension in kidney disease. Curr. Opin. Nephrol. Hypertens. 2012, 21, 1–6. [Google Scholar] [CrossRef]
- Tsuchiya, K.; Tomita, S.; Ishizawa, K.; Abe, S.; Ikeda, Y.; Kihira, Y.; Tamaki, T. Dietary nitrite ameliorates renal injury in L-NAME-induced hypertensive rats. Nitric Oxide 2010, 22, 98–103. [Google Scholar] [CrossRef]
- Gilchrist, M.; Shore, A.C.; Benjamin, N. Inorganic nitrate and nitrite and control of blood pressure. Cardiovasc. Res. 2011, 89, 492–498. [Google Scholar] [CrossRef]
- Lou, H.; Li, H.; Zhang, S.; Lu, H.; Chen, Q. A Review on Preparation of Betulinic Acid and Its Biological Activities. Molecules 2021, 26, 5583. [Google Scholar] [CrossRef]
- Steinkamp-Fenske, K.; Bollinger, L.; Völler, N.; Xu, H.; Yao, Y.; Bauer, R.; Förstermann, U.; Li, H. Ursolic acid from the Chinese herb danshen Salvia miltiorrhiza L. upregulates eNOS and downregulates Nox4 expression in human endothelial cells. Atherosclerosis 2007, 195, e104–e111. [Google Scholar] [CrossRef]
- Steinkamp-Fenske, K.; Bollinger, L.; Xu, H.; Yao, Y.; Horke, S.; Förstermann, U.; Li, H. Reciprocal regulation of endothelial nitric-oxide synthase and NADPH oxidase by betulinic acid in human endothelial cells. J. Pharm. Exp. 2007, 322, 836–842. [Google Scholar] [CrossRef]
- Ovesna, Z.; Vachalkova, A.; Horvathova, K.; Tothova, D. Pentacyclic triterpenoic acids: New chemoprotective compounds. Minireview. Neoplasma 2004, 51, 327–333. [Google Scholar]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M. The International Natural Product Sciences Taskforce, C.T. Supuran. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Sharma, A.; Thakur, R.; Lingaraju, M.C.; Kumar, D.; Mathesh, K.; Telang, A.G.; Singh, T.U.; Kumar, D. Betulinic acid attenuates renal fibrosis in rat chronic kidney disease model. Biomed. Pharm. 2017, 89, 796–804. [Google Scholar] [CrossRef]
- Zeng, A.Q.; Yu, Y.; Yao, Y.Q.; Yang, F.F.; Liao, M.; Song, L.J.; Li, Y.L.; Yu, Y.; Li, Y.J.; Deng, Y.L.; et al. Betulinic acid impairs metastasis and reduces immunosuppressive cells in breast cancer models. Oncotarget 2017, 9, 3794–3804. [Google Scholar] [CrossRef]
- O’Neill, H.M. AMPK and Exercise: Glucose Uptake and Insulin Sensitivity. Diabetes Metab. J. 2013, 37, 1–21. [Google Scholar] [CrossRef]
- Komers, R.; Lindsley, J.N.; Oyama, T.T.; Allison, K.M.; Anderson, S. Role of neuronal nitric oxide synthase NOS1 in the pathogenesis of renal hemodynamic changes in diabetes. Am. J. Physiol. Ren. Physiol. 2000, 279, F573–F583. [Google Scholar] [CrossRef] [PubMed]
- Mount, P.F.; Power, D.A. Nitric oxide in the kidney: Functions and regulation of synthesis. Acta Physiol. 2006, 187, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, S.; Oberbäumer, H. Structural and molecular dissection of the juxtaglomerular apparatus: New aspects for the role of nitric oxide. Kidney Int. 1998, 54, S29–S33. [Google Scholar] [CrossRef] [PubMed]
- Terada, Y.; Tomita, K.; Nonoguchi, H.; Yang, T.; Marumo, F. Different localization and regulation of two types of vasopressin receptor messenger RNA in microdissected rat nephron segments using reverse transcription polymerase chain reaction. J. Clin. Investig. 1993, 92, 2339–2345. [Google Scholar] [CrossRef]
- Datla, S.R.; Griendling, K.K. Reactive oxygen species, NADPH oxidases, and hypertension. Hypertension 2010, 56, 325–330. [Google Scholar] [CrossRef]
- Jia, Y.; Kang, X.; Tan, L.; Ren, Y.; Qu, L.; Tang, J.; Liu, G.; Wang, S.; Xiong, Z.; Yang, L. Nicotinamide Mononucleotide Attenuates Renal Interstitial Fibrosis After AKI by Suppressing Tubular DNA Damage and Senescence. Front. Physiol. 2021, 12, 649547. [Google Scholar] [CrossRef]
- Romero-Puertas, M.C.; Perazzolli, M.; Zago, E.D.; Delledonne, M. Nitric oxide signalling functions in plant–pathogen interactions. Cell. Microbiol. 2004, 6, 795–803. [Google Scholar] [CrossRef]
- Lincoln, T.M.; Komalavilas, P.; Boerth, N.J.; MacMillan-Crow, L.A.; Cornwell, T.L. cGMP Signaling through cAMP- and cGMP-Dependent Protein Kinases. Adv. Pharmacol. 1995, 34, 305–322. [Google Scholar]
- McKee, M.; Scavone, C.; Nathanson, J. Nitric oxide, cGMP, and hormone regulation of active sodium transport. Proc. Natl. Acad. Sci. USA 1995, 91, 12056–12060. [Google Scholar] [CrossRef]
- Snyder, S.H. Nitric oxide: First in a new class of neurotransmitters. Science 1992, 257, 494–496. [Google Scholar] [CrossRef]
- Hohmann, N.; Xia, N.; Steinkamp-Fenske, K.; Förstermann, U.; Li, H. Estrogen Receptor Signaling and the PI3K/Akt Pathway Are Involved in Betulinic Acid-Induced eNOS Activation. Molecules 2016, 21, 973. [Google Scholar] [CrossRef]
- Lee, G.H.; Park, J.S.; Jin, S.W.; Pham, T.H.; Thai, T.N.; Kim, J.Y.; Kim, C.Y.; Choi, J.H.; Han, E.H.; Jeong, H.G. Betulinic Acid Induces eNOS Expression via the AMPK-Dependent KLF2 Signaling Pathway. J. Agric. Food Chem. 2020, 68, 14523–14530. [Google Scholar] [CrossRef]
- Wynn, T.A. Common and unique mechanisms regulate fibrosis in various fibroproliferative diseases. J. Clin. Investig. 2007, 117, 524–529. [Google Scholar] [CrossRef]
- Lu, Q.; Xia, N.; Xu, H.; Guo, L.; Wenzel, P.; Daiber, A.; Munzel, T.; Forstermann, U.; Li, H. Betulinic acid protects against cerebral ischemia-reperfusion injury in mice by reducing oxidative and nitrosative stress. Nitric Oxide Biol. Chem. Off. J. Nitric Oxide Soc. 2011, 24, 132–138. [Google Scholar] [CrossRef]
- Son, Y.; Lee, J.H.; Cheong, Y.K.; Jung, H.C.; Jeong, S.O.; Park, S.H.; Pae, H.O. Piceatannol, a natural hydroxylated analog of resveratrol, promotes nitric oxide release through phosphorylation of endothelial nitric oxide synthase in human endothelial cells. Eur. Rev. Med. Pharm. Sci. 2015, 19, 3125–3132. [Google Scholar]
- Tillery, L.C.; Epperson, T.A.; Eguchi, S.; Motley, E.D. Differential regulation of endothelial nitric oxide synthase phosphorylation by protease-activated receptors in adult human endothelial cells. Exp. Biol. Med. 2016, 241, 569–580. [Google Scholar] [CrossRef]
- Jin, S.W.; Choi, C.Y.; Hwang, Y.P.; Kim, H.G.; Kim, S.J.; Chung, Y.C.; Lee, K.J.; Jeong, T.C.; Jeong, H.G. Betulinic acid increases eNOS phosphorylation and no synthesis via the calcium-signaling pathway. J. Agric. Food Chem. 2016, 64, 785–791. [Google Scholar] [CrossRef]
- Koyama, T.; Hatanaka, Y.; Jin, X.; Yokomizo, A.; Fujiwara, H.; Goda, M.; Hobara, N.; Zamami, Y.; Kitamura, Y.; Kawasaki, H. Altered function of nitrergic nerves inhibiting sympathetic neurotransmission in mesenteric vascular beds of renovascular hypertensive rats. Hypertens. Res. 2020, 33, 485–491. [Google Scholar] [CrossRef]
- Shimada, S.; Todoki, K.; Omori, Y.; Toyama, T.; Matsuo, M.; Wada-Takahashi, S.; Takahashi, S.S.; Lee, M.C. Contribution of nitrergic nerve in canine gingival reactive hyperemia. J. Clin. Biochem. Nutr. 2015, 56, 98–104. [Google Scholar] [CrossRef][Green Version]
- Mignini, F.; Sabbatini, M.; D’Andrea, V.; Cavallotti, C. Intrinsic innervation and dopaminergic markers after experimental denervation in rat thymus. Eur. J. Histochem. 2010, 54, e17. [Google Scholar] [CrossRef]
- Dorko, F.; Danko, J.; Flešárová, S.; Boroš, E.; Sobeková, A. Effect of pesticide bendiocar-bamate on distribution of acetylcholine- and butyrylcholine-positive nerves in rabbit’s thymus. Eur. J. Histochem. 2011, 55, e37. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Na, C.; Huh, Y. Alterations in nitric oxide synthase in the aged CNS. Oxid. Med. Cell. Longev. 2012, 2012, 718976. [Google Scholar] [CrossRef]
- Cossenza, M.; Socodato, R.; Portugal, C.C.; Domith, I.C.; Gladulich, L.F.; Encarnação, T.G.; Calaza, K.C.; Mendonça, H.R.; Campello-Costa, P.; Paes-de-Carvalho, R. Nitric oxide in the nervous system: Biochemical, developmental, and neurobiological aspects. Vitam. Horm. 2014, 96, 79–125. [Google Scholar] [PubMed]
- Liu, C.; Yang, Y.; Hu, X.; Li, J.M.; Zhang, X.M.; Cai, Y.; Li, Z.; Yan, X.X. Ontogenesis of NADPH-diaphorase positive neurons in guinea pig neocortex. Front. Neuroanat. 2015, 9, 11. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, C.; Giulivi, C. Subcellular and cellular locations of nitric oxide synthase isoforms as determinants of health and disease. Free Radic. Biol. Med. 2010, 49, 307–316. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).