
Expression of the Purinergic P2X7 Receptor in Murine MOPC315.BM Myeloma Cells

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Calcium Influx Assay
2.3. Pore Formation
2.4. TaqMan Gene Expression
2.5. Statistics
3. Results
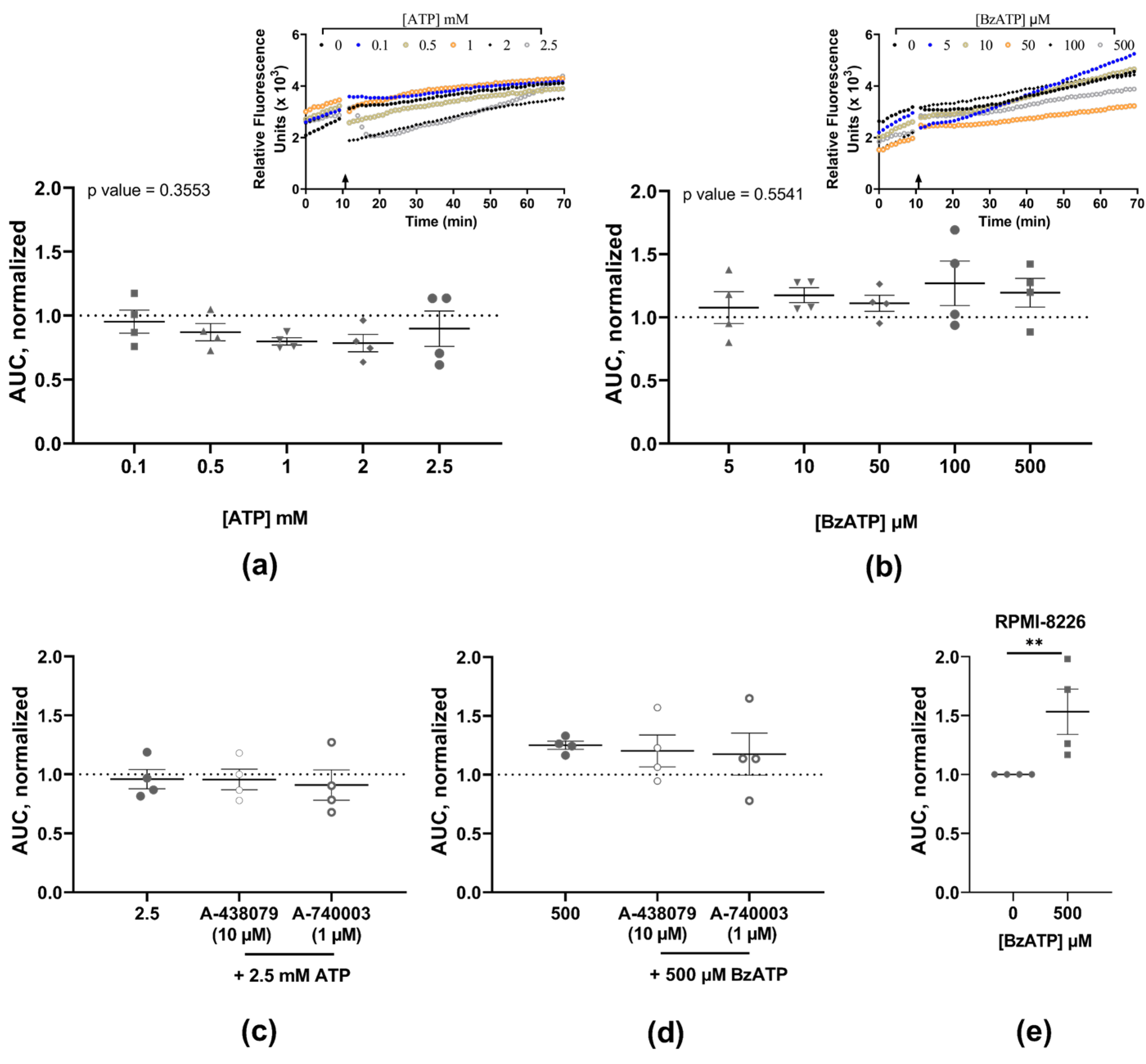
3.1. Neither ATP nor BzATP Induces Calcium Influx in MOPC315.BM
3.2. Neither ATP nor BzATP Induces Pore Formation in MOPC315.BM
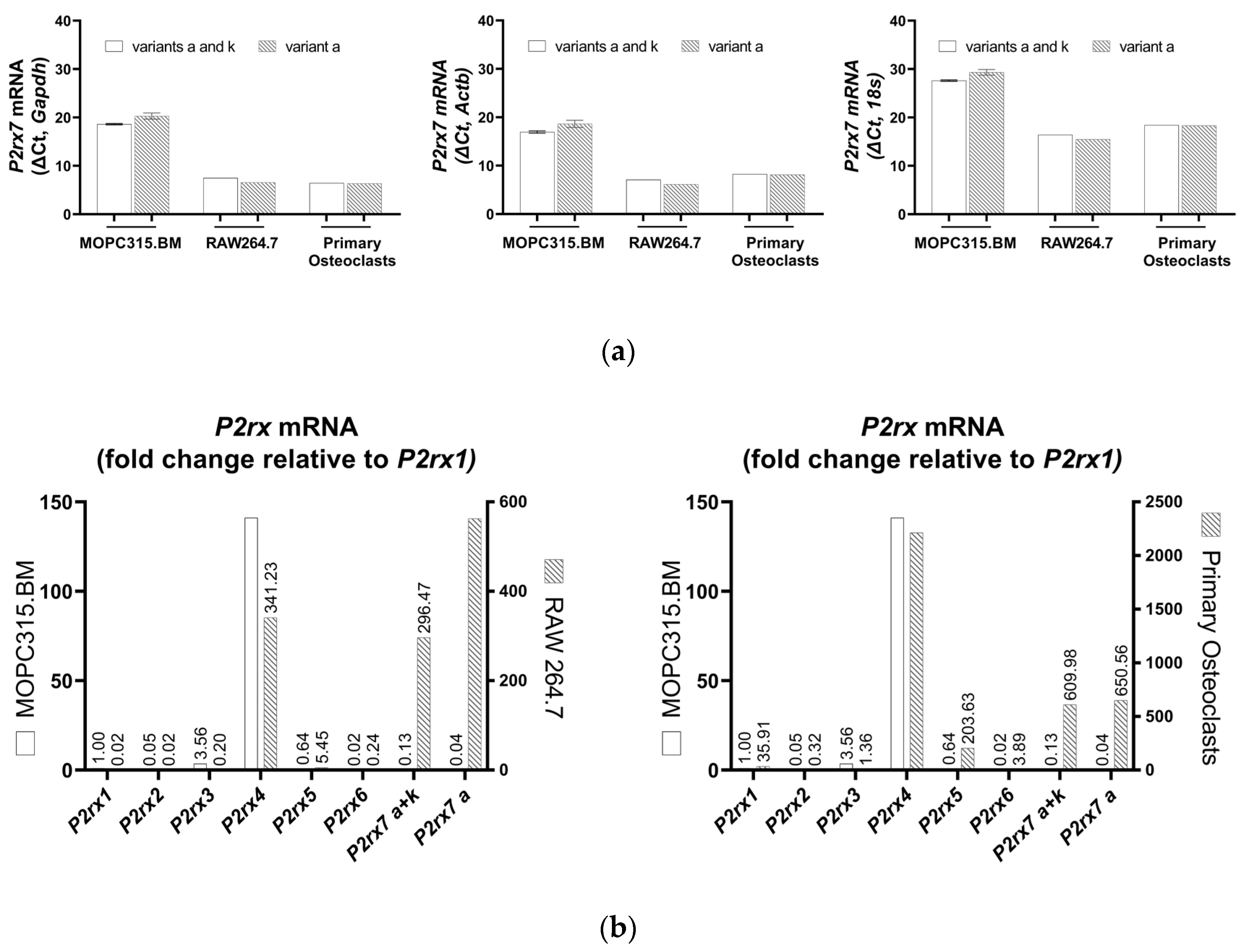
3.3. Expression of P2X7 Receptor Variants in MOPC315.BM
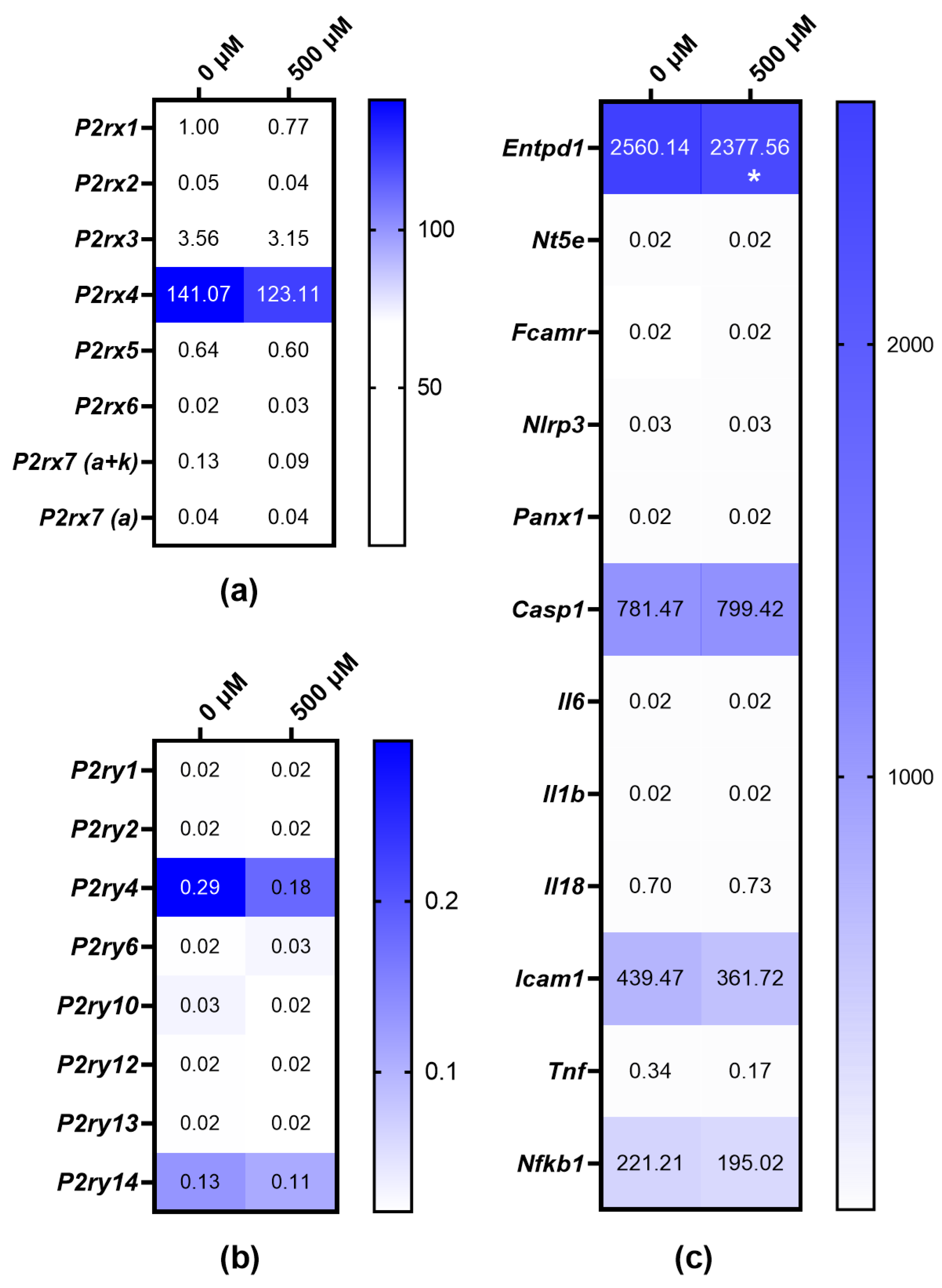
3.4. Expressions of P2XR and P2YR Subtypes in MOPC315.BM and BzATP-Induced Genetic Regulation
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Surprenant, A.; Rassendren, F.; Kawashima, E.; North, R.A.; Buell, G. The cytolytic P2Z receptor for extracellular ATP identified as a P2X receptor (P2X7). Science 1996, 272, 735–738. [Google Scholar] [CrossRef] [PubMed]
- Lara, R.; Adinolfi, E.; Harwood, C.A.; Philpott, M.; Barden, J.A.; Di Virgilio, F.; McNulty, S. P2X7 in Cancer: From Molecular Mechanisms to Therapeutics. Front. Pharmacol. 2020, 11, 793. [Google Scholar] [CrossRef] [PubMed]
- Khakh, B.S.; North, R.A. P2X receptors as cell-surface ATP sensors in health and disease. Nature 2006, 442, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G.; Kennedy, C. P2X receptors in health and disease. Adv. Pharmacol. 2011, 61, 333–372. [Google Scholar] [CrossRef]
- Di Virgilio, F.; Schmalzing, G.; Markwardt, F. The Elusive P2X7 Macropore. Trends Cell Biol. 2018, 28, 392–404. [Google Scholar] [CrossRef]
- Di Virgilio, F.; Giuliani, A.L.; Vultaggio-Poma, V.; Falzoni, S.; Sarti, A.C. Non-nucleotide Agonists Triggering P2X7 Receptor Activation and Pore Formation. Front. Pharmacol. 2018, 9, 39. [Google Scholar] [CrossRef]
- Di Virgilio, F. P2RX7: A receptor with a split personality in inflammation and cancer. Mol. Cell. Oncol. 2016, 3, e1010937. [Google Scholar] [CrossRef]
- Di Virgilio, F.; Sarti, A.C.; Falzoni, S.; De Marchi, E.; Adinolfi, E. Extracellular ATP and P2 purinergic signalling in the tumour microenvironment. Nat. Rev. Cancer 2018, 18, 601–618. [Google Scholar] [CrossRef]
- Burnstock, G.; Di Virgilio, F. Purinergic signalling and cancer. Purinergic Signal. 2013, 9, 491–540. [Google Scholar] [CrossRef]
- Sluyter, R.; Adriouch, S.; Fuller, S.J.; Nicke, A.; Sophocleous, R.A.; Watson, D. Animal Models for the Investigation of P2X7 Receptors. Int. J. Mol. Sci. 2023, 24, 8225. [Google Scholar] [CrossRef]
- Muller, C.E.; Baqi, Y.; Namasivayam, V. Agonists and Antagonists for Purinergic Receptors. Methods Mol. Biol. 2020, 2041, 45–64. [Google Scholar] [CrossRef]
- Kumar, S.K.; Rajkumar, V.; Kyle, R.A.; van Duin, M.; Sonneveld, P.; Mateos, M.V.; Gay, F.; Anderson, K.C. Multiple myeloma. Nat. Rev. Dis. Primers 2017, 3, 17046. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, S.V. Multiple myeloma: 2018 update on diagnosis, risk-stratification, and management. Am. J. Hematol. 2018, 93, 981–1114. [Google Scholar] [CrossRef] [PubMed]
- Marino, S.; Roodman, G.D. Multiple Myeloma and Bone: The Fatal Interaction. Cold Spring Harb. Perspect. Med. 2018, 8, a031666. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.C.; Carrasco, R.D. Pathogenesis of myeloma. Annu. Rev. Pathol. 2011, 6, 249–274. [Google Scholar] [CrossRef]
- Sperling, A.S.; Anderson, K.C. Facts and Hopes in Multiple Myeloma Immunotherapy. Clin. Cancer Res. 2021, 27, 4468–4477. [Google Scholar] [CrossRef]
- Robak, P.; Drozdz, I.; Szemraj, J.; Robak, T. Drug resistance in multiple myeloma. Cancer Treat. Rev. 2018, 70, 199–208. [Google Scholar] [CrossRef]
- Pinto, V.; Bergantim, R.; Caires, H.R.; Seca, H.; Guimaraes, J.E.; Vasconcelos, M.H. Multiple Myeloma: Available Therapies and Causes of Drug Resistance. Cancers 2020, 12, 407. [Google Scholar] [CrossRef]
- Hofgaard, P.O.; Jodal, H.C.; Bommert, K.; Huard, B.; Caers, J.; Carlsen, H.; Schwarzer, R.; Schunemann, N.; Jundt, F.; Lindeberg, M.M.; et al. A novel mouse model for multiple myeloma (MOPC315.BM) that allows noninvasive spatiotemporal detection of osteolytic disease. PLoS ONE 2012, 7, e51892. [Google Scholar] [CrossRef]
- Agrawal, A.; Kruse, L.S.; Vangsted, A.J.; Gartland, A.; Jorgensen, N.R. Human P2X7 Receptor Causes Cycle Arrest in RPMI-8226 Myeloma Cells to Alter the Interaction with Osteoblasts and Osteoclasts. Cells 2020, 9, 2341. [Google Scholar] [CrossRef]
- Agrawal, A.; Ellegaard, M.; Haanes, K.A.; Wang, N.; Gartland, A.; Ding, M.; Praetorius, H.; Jorgensen, N.R. Absence of P2Y(2) Receptor Does Not Prevent Bone Destruction in a Murine Model of Muscle Paralysis-Induced Bone Loss. Front. Endocrinol. 2022, 13, 850525. [Google Scholar] [CrossRef] [PubMed]
- Donnelly-Roberts, D.L.; Namovic, M.T.; Han, P.; Jarvis, M.F. Mammalian P2X7 receptor pharmacology: Comparison of recombinant mouse, rat and human P2X7 receptors. Br. J. Pharmacol. 2009, 157, 1203–1214. [Google Scholar] [CrossRef] [PubMed]
- Nicke, A.; Kuan, Y.H.; Masin, M.; Rettinger, J.; Marquez-Klaka, B.; Bender, O.; Gorecki, D.C.; Murrell-Lagnado, R.D.; Soto, F. A functional P2X7 splice variant with an alternative transmembrane domain 1 escapes gene inactivation in P2X7 knock-out mice. J. Biol. Chem. 2009, 284, 25813–25822. [Google Scholar] [CrossRef]
- Inoue, K.; Tsuda, M. Nociceptive signaling mediated by P2X3, P2X4 and P2X7 receptors. Biochem. Pharmacol. 2021, 187, 114309. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, X.; Wang, C.; Li, G.; Gu, Y.; Huang, L.Y. Activation of P2X7 receptors in glial satellite cells reduces pain through downregulation of P2X3 receptors in nociceptive neurons. Proc. Natl. Acad. Sci. USA 2008, 105, 16773–16778. [Google Scholar] [CrossRef]
- Gonzalez-Rodriguez, S.; Pevida, M.; Roques, B.P.; Fournie-Zaluski, M.C.; Hidalgo, A.; Menendez, L.; Baamonde, A. Involvement of enkephalins in the inhibition of osteosarcoma-induced thermal hyperalgesia evoked by the blockade of peripheral P2X3 receptors. Neurosci. Lett. 2009, 465, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Hansen, R.R.; Nasser, A.; Falk, S.; Baldvinsson, S.B.; Ohlsson, P.H.; Bahl, J.M.; Jarvis, M.F.; Ding, M.; Heegaard, A.M. Chronic administration of the selective P2X3, P2X2/3 receptor antagonist, A-317491, transiently attenuates cancer-induced bone pain in mice. Eur. J. Pharmacol. 2012, 688, 27–34. [Google Scholar] [CrossRef]
- Kaan, T.K.; Yip, P.K.; Patel, S.; Davies, M.; Marchand, F.; Cockayne, D.A.; Nunn, P.A.; Dickenson, A.H.; Ford, A.P.; Zhong, Y.; et al. Systemic blockade of P2X3 and P2X2/3 receptors attenuates bone cancer pain behaviour in rats. Brain 2010, 133, 2549–2564. [Google Scholar] [CrossRef]
- Xiang, Z.; Lv, J.; Jiang, P.; Chen, C.; Jiang, B.; Burnstock, G. Expression of P2X receptors on immune cells in the rat liver during postnatal development. Histochem. Cell Biol. 2006, 126, 453–463. [Google Scholar] [CrossRef]
- Burnstock, G. Blood cells: An historical account of the roles of purinergic signalling. Purinergic Signal. 2015, 11, 411–434. [Google Scholar] [CrossRef]
- Vargas-Martinez, E.M.; Gomez-Coronado, K.S.; Espinosa-Luna, R.; Valdez-Morales, E.E.; Barrios-Garcia, T.; Barajas-Espinosa, A.; Ochoa-Cortes, F.; Montano, L.M.; Barajas-Lopez, C.; Guerrero-Alba, R. Functional expression of P2X1, P2X4 and P2X7 purinergic receptors in human monocyte-derived macrophages. Eur. J. Pharmacol. 2020, 888, 173460. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Unresolved issues and controversies in purinergic signalling. J. Physiol. 2008, 586, 3307–3312. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.; Prudic, K.; Pippel, A.; Klapperstuck, M.; Braam, U.; Muller, C.E.; Schmalzing, G.; Markwardt, F. Interaction of Purinergic P2X4 and P2X7 Receptor Subunits. Front. Pharmacol. 2017, 8, 860. [Google Scholar] [CrossRef]
- Craigie, E.; Birch, R.E.; Unwin, R.J.; Wildman, S.S. The relationship between P2X4 and P2X7: A physiologically important interaction? Front. Physiol. 2013, 4, 216. [Google Scholar] [CrossRef] [PubMed]
- Weinhold, K.; Krause-Buchholz, U.; Rodel, G.; Kasper, M.; Barth, K. Interaction and interrelation of P2X7 and P2X4 receptor complexes in mouse lung epithelial cells. Cell. Mol. Life Sci. 2010, 67, 2631–2642. [Google Scholar] [CrossRef]
- Adamiak, M.; Bujko, K.; Thapa, A.; Pensato, V.; Brzezniakiewicz-Janus, K.; Ratajczak, J.; Davies, D.L.; Ulrich, H.; Kucia, M.; Ratajczak, M.Z. The P2X4 purinergic receptor has emerged as a potent regulator of hematopoietic stem/progenitor cell mobilization and homing-a novel view of P2X4 and P2X7 receptor interaction in orchestrating stem cell trafficking. Leukemia 2022, 36, 248–256. [Google Scholar] [CrossRef]
- Vanderkerken, K.; Asosingh, K.; Croucher, P.; Van Camp, B. Multiple myeloma biology: Lessons from the 5TMM models. Immunol. Rev. 2003, 194, 196–206. [Google Scholar] [CrossRef]
- Paton-Hough, J.; Chantry, A.D.; Lawson, M.A. A review of current murine models of multiple myeloma used to assess the efficacy of therapeutic agents on tumour growth and bone disease. Bone 2015, 77, 57–68. [Google Scholar] [CrossRef]
- Diaz-del Castillo, M.; Kamstrup, D.; Olsen, R.B.; Hansen, R.B.; Pembridge, T.; Simanskaite, B.; Jimenez-Andrade, J.M.; Lawson, M.A.; Heegaard, A.M. Differential Pain-Related Behaviors and Bone Disease in Immunocompetent Mouse Models of Myeloma. JBMR Plus 2020, 4, e10252. [Google Scholar] [CrossRef]
- Vanderkerken, K.; De Raeve, H.; Goes, E.; Van Meirvenne, S.; Radl, J.; Van Riet, I.; Thielemans, K.; Van Camp, B. Organ involvement and phenotypic adhesion profile of 5T2 and 5T33 myeloma cells in the C57BL/KaLwRij mouse. Br. J. Cancer 1997, 76, 451–460. [Google Scholar] [CrossRef]
- Cheung, W.C.; Kim, J.S.; Linden, M.; Peng, L.; Van Ness, B.; Polakiewicz, R.D.; Janz, S. Novel targeted deregulation of c-Myc cooperates with Bcl-X(L) to cause plasma cell neoplasms in mice. J. Clin. Investig. 2004, 113, 1763–1773. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, D.R.; Sukhdeo, K.; Protopopova, M.; Sinha, R.; Enos, M.; Carrasco, D.E.; Zheng, M.; Mani, M.; Henderson, J.; Pinkus, G.S.; et al. The differentiation and stress response factor XBP-1 drives multiple myeloma pathogenesis. Cancer Cell 2007, 11, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Chesi, M.; Robbiani, D.F.; Sebag, M.; Chng, W.J.; Affer, M.; Tiedemann, R.; Valdez, R.; Palmer, S.E.; Haas, S.S.; Stewart, A.K.; et al. AID-dependent activation of a MYC transgene induces multiple myeloma in a conditional mouse model of post-germinal center malignancies. Cancer Cell 2008, 13, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Winkler, W.; Farre Diaz, C.; Blanc, E.; Napieczynska, H.; Langner, P.; Werner, M.; Walter, B.; Wollert-Wulf, B.; Yasuda, T.; Heuser, A.; et al. Mouse models of human multiple myeloma subgroups. Proc. Natl. Acad. Sci. USA 2023, 120, e2219439120. [Google Scholar] [CrossRef] [PubMed]
- Mitsiades, C.S.; Anderson, K.C.; Carrasco, D.R. Mouse models of human myeloma. Hematol. Oncol. Clin. N. Am. 2007, 21, 1051–1069. [Google Scholar] [CrossRef]
- Paton-Hough, J.; Tazzyman, S.; Evans, H.; Lath, D.; Down, J.M.; Green, A.C.; Snowden, J.A.; Chantry, A.D.; Lawson, M.A. Preventing and Repairing Myeloma Bone Disease by Combining Conventional Antiresorptive Treatment With a Bone Anabolic Agent in Murine Models. J. Bone Miner. Res. 2019, 34, 783–796. [Google Scholar] [CrossRef]
- Adinolfi, E.; Raffaghello, L.; Giuliani, A.L.; Cavazzini, L.; Capece, M.; Chiozzi, P.; Bianchi, G.; Kroemer, G.; Pistoia, V.; Di Virgilio, F. Expression of P2X7 receptor increases in vivo tumor growth. Cancer Res. 2012, 72, 2957–2969. [Google Scholar] [CrossRef]
- Adinolfi, E.; Capece, M.; Franceschini, A.; Falzoni, S.; Giuliani, A.L.; Rotondo, A.; Sarti, A.C.; Bonora, M.; Syberg, S.; Corigliano, D.; et al. Accelerated tumor progression in mice lacking the ATP receptor P2X7. Cancer Res. 2015, 75, 635–644. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbols | Assay IDs | Gene Symbols | Assay IDs | Gene Symbols | Assay IDs | Gene Symbols | Assay IDs |
|---|---|---|---|---|---|---|---|
| 18s | Hs99999901_s1 | P2rx1 | Mm00435460_m1 | P2ry1 | Mm00435471_m | Entpd1 | Mm00515447_m1 |
| Gapdh | Mm99999915_g1 | P2rx2 | Mm00462952_m1 | P2ry2 | Mm00435472_m1 | Nt5e | Mm00501910_m1 |
| Actb | Mm01205647_g1 | P2rx3 | Mm00523699_m1 | P2ry4 | Mm00445136_s1 | Fcamr | Mm03047928_m1 |
| Hprt | Mm03024075_m1 | P2rx4 | Mm00501787_m1 | P2ry6 | Mm01275473_m1 | Nlrp3 | Mm00840904_m1 |
| P2rx5 | Mm00473677_m1 | P2ry10 | Mm00549811_m1 | Panx1 | Mm00450900_m1 | ||
| P2rx6 | Mm00440591_m1 | P2ry12 | Mm00446026_m1 | Casp1 | Mm00438023_m1 | ||
| P2rx7 * | Mm01199500_m1 | P2ry13 | Mm00546978_m1 | Il6 | Mm00446190_m1 | ||
| P2rx7 ** | Mm00440578_m1 | P2ry14 | Mm01289602_m1 | Il1b | Mm00434228_m1 | ||
| Il18 | Mm00434226_m1 | ||||||
| Icam1 | Mm00516023_m1 | ||||||
| Tnf | Mm00443258_m1 | ||||||
| Nfkb1 | Mm00476361_m1 |
| [BzATP] | 0 μM | 10 μM | 50 μM | 100 μM | 200 μM | 300 μM | ||
|---|---|---|---|---|---|---|---|---|
| Fmax/F0, mean ± SEM | 1.000 ± 0.000 | 1.201 ± 0.086 | 1.214 ± 0.109 | 0.873 ± 0.175 | 1.333 ± 0.103 | 1.357 ± 0.153 | ||
| (n) | (4) | (4) | (4) | (4) | (4) | (4) | ||
| p-value | - | 0.803 | 0.6638 | >0.999 | 0.1373 | 0.1559 | ||
| [BzATP] | 0 μM | 300 μM | 300 μM | 300 μM | ||||
| + 10 µM A-438079 | 1 µM A-740003 | |||||||
| Fmax/F0, mean ± SEM | 1.000 ± 0.000 | 1.020 ± 0.088 | 1.170 ± 0.222 | 0.866 ± 0.225 | ||||
| (n) | (4) | (4) | (4) | (4) | ||||
| p-value | - | 0.3143 | 0.857 | >0.999 | ||||
| [ATP] | 0 mM | 0.5 mM | 1 mM | 2 mM | 3 mM | 4 mM | ||
| Fmax/F0, mean ± SEM | 1.000 ± 0.000 | 1.553 ± 0.458 | 1.260 ± 0.242 | 1.935 ± 0.616 | 1.864 ± 0.453 | 1.056 ± 0.409 | ||
| (n) | (4) | (4) | (4) | (4) | (4) | (4) | ||
| p-value | - | >0.999 | >0.999 | 0.535 | 0.5726 | >0.999 | ||
| [ATP] | 0 mM | 2.5 mM | 2.5 mM | 2.5 mM | ||||
| + 10 µM A-438079 | 1 µM A-740003 | |||||||
| Fmax/F0, mean ± SEM | 1.000 ± 0.000 | 1.841 ± 0.539 | 2.299 ± 1.126 | 1.533 ± 0.405 | ||||
| (n) | (4) | (4) | (4) | (4) | ||||
| p-value | - | 0.9333 | 0.6857 | 0.8857 | ||||
| [BzATP] | 0 μM | 5 μM | 10 μM | 50 μM | 100 μM | 500 μM | ||
|---|---|---|---|---|---|---|---|---|
| AUC, mean ± SEM | 1.000 ± 0.000 | 1.077 ± 0.126 | 1.175 ± 0.059 | 1.111 ± 0.064 | 1.269 ± 0.176 | 1.195 ± 0.1134 | ||
| (n) | (4) | (4) | (4) | (4) | (4) | (4) | ||
| p-value | - | >0.999 | 0.5441 | >0.999 | 0.6638 | 0.803 | ||
| [BzATP] | 0 μM | 500 μM | 500 μM | 500 μM | ||||
| + 10 µM A-438079 | 1 µM A-740003 | |||||||
| AUC, mean ± SEM | 1.000 ± 0.000 | 1.250 ± 0.033 | 1.202 ± 0.135 | 1.175 ± 0.179 | ||||
| (n) | (4) | (4) | (4) | (4) | ||||
| p-value | - | 0.0286 | 0.4857 | 0.3429 | ||||
| [ATP] | 0 mM | 0.1 mM | 0.5 mM | 1 mM | 2 mM | 2.5 mM | ||
| AUC, mean ± SEM | 1.000 ± 0.000 | 0.953 ± 0.089 | 0.870 ± 0.067 | 0.798 ± 0.028 | 0.786 ± 0.068 | 0.898 ± 0.139 | ||
| (n) | (4) | (4) | (4) | (4) | (4) | (4) | ||
| p-value | - | >0.999 | >0.999 | 0.491 | 0.2534 | >0.999 | ||
| [ATP] | 0 mM | 2.5 mM | 2.5 mM | 2.5 mM | ||||
| + 10 µM A-438079 | 1 µM A-740003 | |||||||
| AUC, mean ± SEM | 1.000 ± 0.000 | 0.959 ± 0.164 | 0.956 ± 0.087 | 0.909 ± 0.129 | ||||
| (n) | (4) | (4) | (4) | (4) | ||||
| p-value | - | 0.3143 | 0.8857 | 0.6857 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Høyer, E.R.; Demir, M.; Bak, L.K.; Jørgensen, N.R.; Agrawal, A. Expression of the Purinergic P2X7 Receptor in Murine MOPC315.BM Myeloma Cells. Receptors 2023, 2, 191-203. https://doi.org/10.3390/receptors2030013
Høyer ER, Demir M, Bak LK, Jørgensen NR, Agrawal A. Expression of the Purinergic P2X7 Receptor in Murine MOPC315.BM Myeloma Cells. Receptors. 2023; 2(3):191-203. https://doi.org/10.3390/receptors2030013
Chicago/Turabian StyleHøyer, Eva Risborg, Melisa Demir, Lasse Kristoffer Bak, Niklas Rye Jørgensen, and Ankita Agrawal. 2023. "Expression of the Purinergic P2X7 Receptor in Murine MOPC315.BM Myeloma Cells" Receptors 2, no. 3: 191-203. https://doi.org/10.3390/receptors2030013
APA StyleHøyer, E. R., Demir, M., Bak, L. K., Jørgensen, N. R., & Agrawal, A. (2023). Expression of the Purinergic P2X7 Receptor in Murine MOPC315.BM Myeloma Cells. Receptors, 2(3), 191-203. https://doi.org/10.3390/receptors2030013