

The NK-1 Receptor Signaling: Distribution and Functional Relevance in the Eye


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. The Tachykinin Peptide Family and Its Receptors
1.2. NK1R Structure
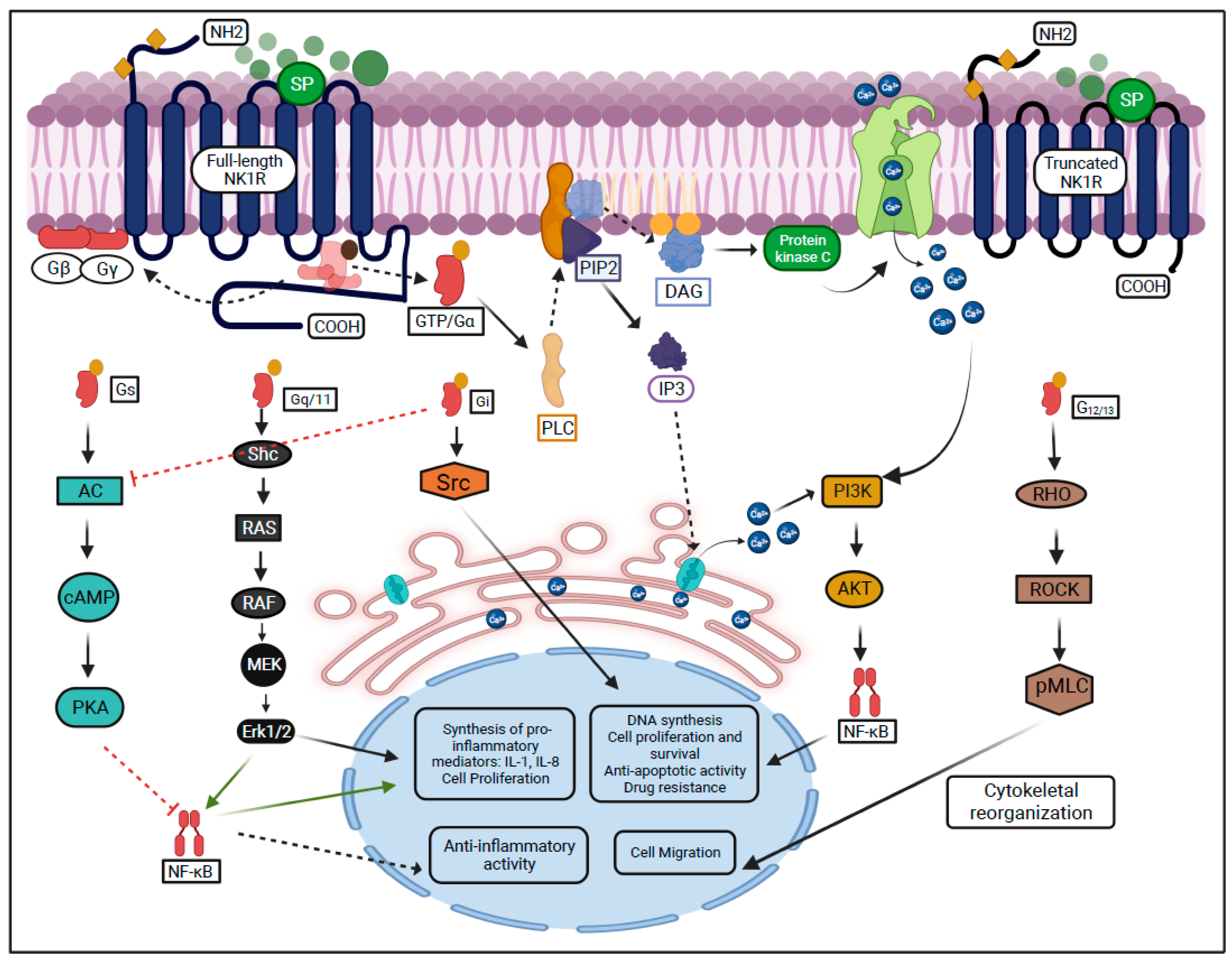
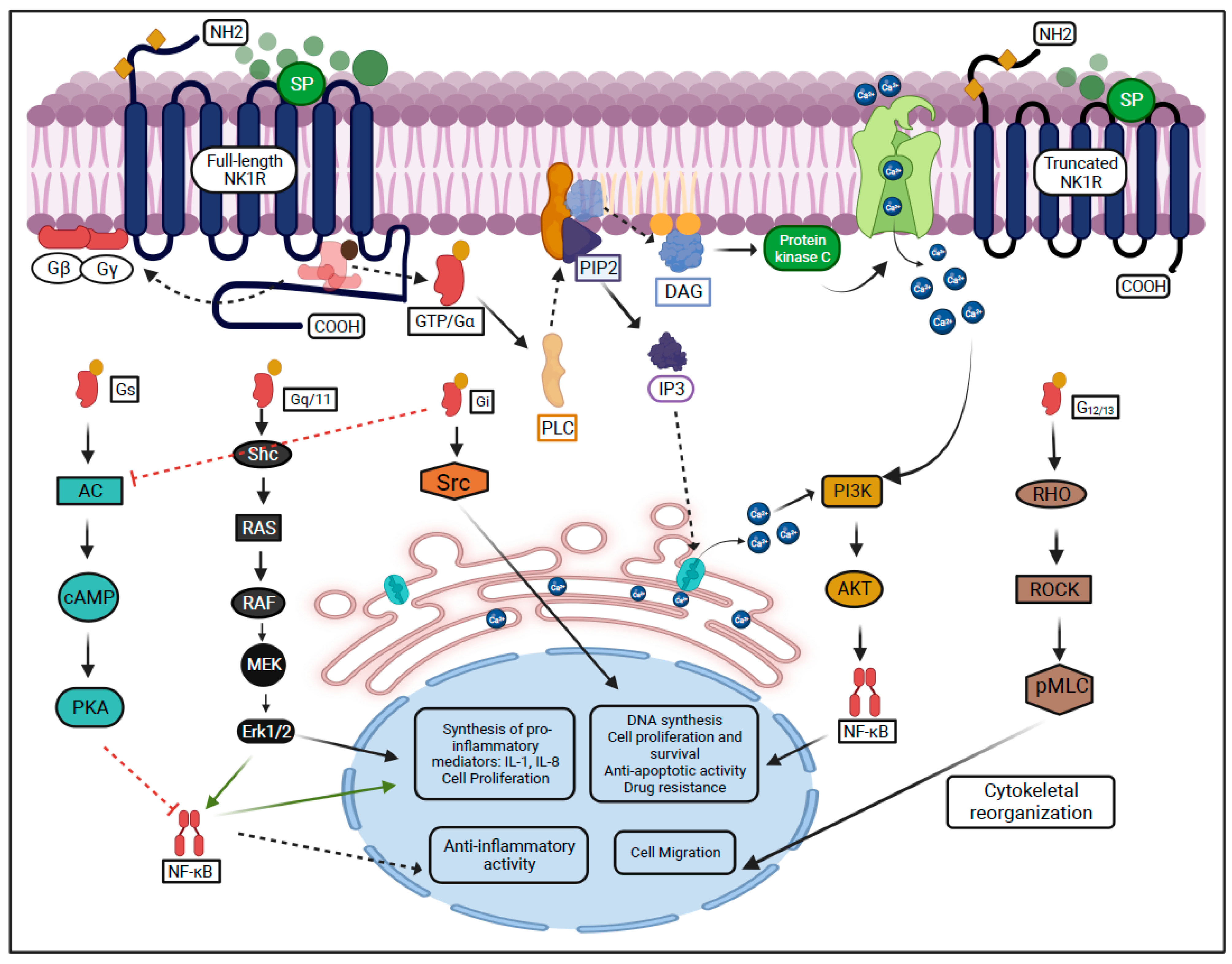
1.3. NK1R Signaling Activity
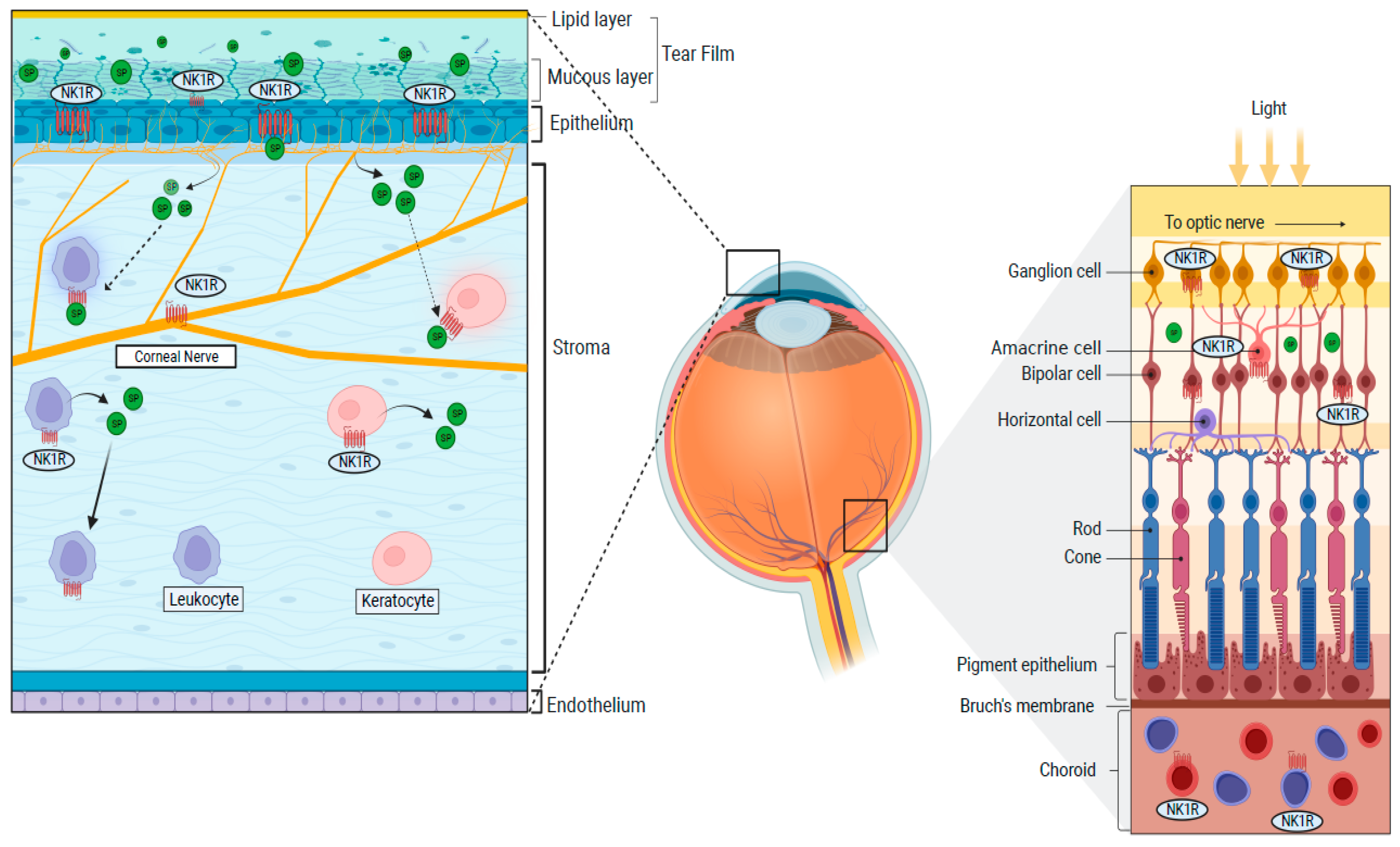
1.4. Distribution of NK1R in the Eye
1.5. NK1R in Wound Healing, Inflammation, and Pain
1.5.1. NK1R and Corneal Epithelial Wound Healing
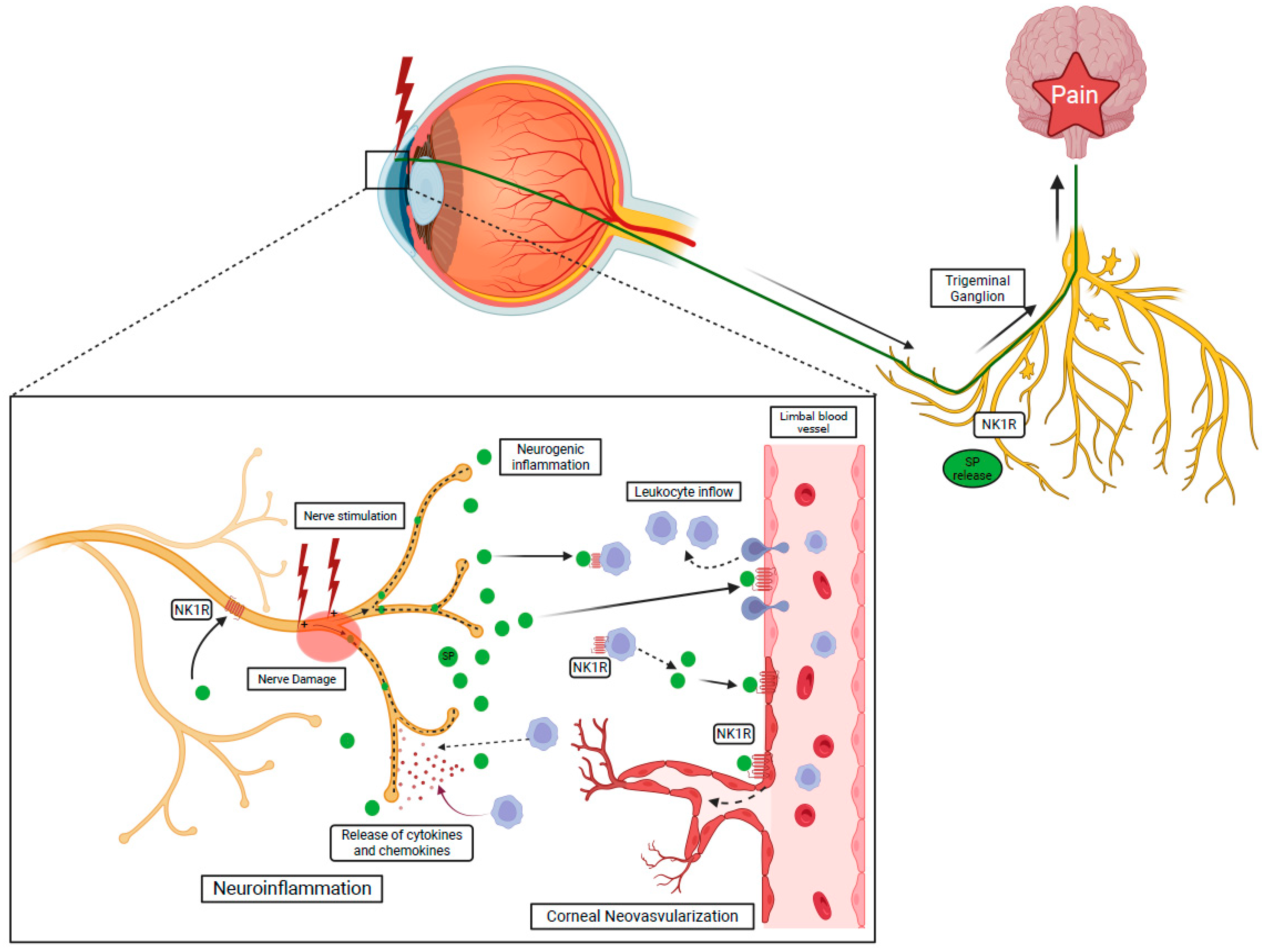
1.5.2. NK1R and Ocular Inflammation
1.5.3. NK1R and Ocular Pain
1.6. NK1R Antagonists and Their Potential in Eye Diseases
2. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Pennefather, J.N.; Lecci, A.; Candenas, M.L.; Patak, E.; Pinto, F.M.; Maggi, C.A. Tachykinins and Tachykinin Receptors: A Growing Family. Life Sci. 2004, 74, 1445–1463. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Berger, A.; Milne, C.D.; Paige, C.J. Tachykinins in the Immune System. Curr. Drug Targets 2006, 7, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Lasagni Vitar, R.M.; Rama, P.; Ferrari, G. The Two-Faced Effects of Nerves and Neuropeptides in Corneal Diseases. Prog. Retin. Eye Res. 2022, 86, 100974. [Google Scholar] [CrossRef] [PubMed]
- Pinto, F.M.; Pintado, C.O.; Pennefather, J.N.; Patak, E.; Candenas, L. Ovarian Steroids Regulate Tachykinin and Tachykinin Receptor Gene Expression in the Mouse Uterus. Reprod. Biol. Endocrinol. 2009, 7, 77. [Google Scholar] [CrossRef] [Green Version]
- Gerard, N.P.; Garraway, L.A.; Eddy, R.L.; Shows, T.B.; Iijima, H.; Paquet, J.L.; Gerard, C. Human Substance P Receptor (NK-1): Organization of the Gene, Chromosome Localization, and Functional Expression of CDNA Clones. Biochemistry 1991, 30, 10640–10646. [Google Scholar] [CrossRef]
- Blasco, V.; Pinto, F.M.; González-Ravina, C.; Santamaría-López, E.; Candenas, L.; Fernández-Sánchez, M. Tachykinins and Kisspeptins in the Regulation of Human Male Fertility. J. Clin. Med. 2019, 9, 113. [Google Scholar] [CrossRef] [Green Version]
- Keringer, P.; Rumbus, Z. The Interaction between Neurokinin-1 Receptors and Cyclooxygenase-2 in Fever Genesis. Temp. Austin 2019, 6, 4–6. [Google Scholar] [CrossRef] [Green Version]
- Almeida, T.A.; Rojo, J.; Nieto, P.M.; Pinto, F.M.; Hernandez, M.; Martín, J.D.; Candenas, M.L. Tachykinins and Tachykinin Receptors: Structure and Activity Relationships. Curr. Med. Chem. 2004, 11, 2045–2081. [Google Scholar] [CrossRef]
- Douglas, S.D.; Leeman, S.E. Neurokinin-1 Receptor: Functional Significance in the Immune System in Reference to Selected Infections and Inflammation. Ann. N. Y. Acad. Sci. 2011, 1217, 83–95. [Google Scholar] [CrossRef] [Green Version]
- Maggi, C.A. The Mammalian Tachykinin Receptors. Gen. Pharmacol. Vasc. Syst. 1995, 26, 911–944. [Google Scholar] [CrossRef]
- Kolorz, J.; Demir, S.; Gottschlich, A.; Beirith, I.; Ilmer, M.; Lüthy, D.; Walz, C.; Dorostkar, M.M.; Magg, T.; Hauck, F.; et al. The Neurokinin-1 Receptor Is a Target in Pediatric Rhabdoid Tumors. Curr. Oncol. Tor. Ont 2021, 29, 94–110. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.-P.; Lai, S.; Tuluc, F.; Tansky, M.F.; Kilpatrick, L.E.; Leeman, S.E.; Douglas, S.D. Differences in the Length of the Carboxyl Terminus Mediate Functional Properties of Neurokinin-1 Receptor. Proc. Natl. Acad. Sci. USA 2008, 105, 12605–12610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitsin, S.; Pappa, V.; Douglas, S.D. Truncation of Neurokinin-1 Receptor-Negative Regulation of Substance P Signaling. J. Leukoc. Biol. 2018, 103, 1043–1051. [Google Scholar] [CrossRef] [PubMed]
- Eapen, P.M.; Rao, C.M.; Nampoothiri, M. Crosstalk between Neurokinin Receptor Signaling and Neuroinflammation in Neurological Disorders. Rev. Neurosci. 2019, 30, 233–243. [Google Scholar] [CrossRef]
- Markovic, D.; Challiss, R.A.J. Alternative Splicing of G Protein-Coupled Receptors: Physiology and Pathophysiology. Cell. Mol. Life Sci. 2009, 66, 3337–3352. [Google Scholar] [CrossRef] [Green Version]
- Tansky, M.F.; Pothoulakis, C.; Leeman, S.E. Functional Consequences of Alteration of N-Linked Glycosylation Sites on the Neurokinin 1 Receptor. Proc. Natl. Acad. Sci. USA 2007, 104, 10691–10696. [Google Scholar] [CrossRef] [Green Version]
- Redkiewicz, P. The Regenerative Potential of Substance P. Int. J. Mol. Sci. 2022, 23, 750. [Google Scholar] [CrossRef]
- Singh, R.B.; Naderi, A.; Cho, W.; Ortiz, G.; Musayeva, A.; Dohlman, T.H.; Chen, Y.; Ferrari, G.; Dana, R. Modulating the Tachykinin: Role of Substance P and Neurokinin Receptor Expression in Ocular Surface Disorders. Ocul. Surf. 2022, 25, 142–153. [Google Scholar] [CrossRef]
- Bignami, F.; Rama, P.; Ferrari, G. Substance P and Its Inhibition in Ocular Inflammation. Curr. Drug Targets 2016, 17, 1265–1274. [Google Scholar] [CrossRef]
- Maggi, C.A.; Patacchini, R.; Giachetti, A.; Meli, A. Tachykinin Receptors in the Circular Muscle of the Guinea-Pig Ileum. Br. J. Pharmacol. 1990, 101, 996–1000. [Google Scholar] [CrossRef]
- Satake, H.; Kawada, T. Overview of the Primary Structure, Tissue-Distribution, and Functions of Tachykinins and Their Receptors. Curr. Drug Targets 2006, 7, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Bost, K.L. Tachykinin-Mediated Modulation of the Immune Response. Front. Biosci. 2004, 9, 3331–3332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koon, H.-W.; Zhao, D.; Na, X.; Moyer, M.P.; Pothoulakis, C. Metalloproteinases and Transforming Growth Factor-α Mediate Substance P-Induced Mitogen-Activated Protein Kinase Activation and Proliferation in Human Colonocytes. J. Biol. Chem. 2004, 279, 45519–45527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Hirschfeld, J.; Lopez-Briones, L.G.; Belmonte, C. Neurotrophic Influences on Corneal Epithelial Cells. Exp. Eye Res. 1994, 59, 597–605. [Google Scholar] [CrossRef]
- Słoniecka, M.; Le Roux, S.; Zhou, Q.; Danielson, P. Substance P Enhances Keratocyte Migration and Neutrophil Recruitment through Interleukin-8. Mol. Pharmacol. 2016, 89, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Morelli, A.E.; Sumpter, T.L.; Rojas-Canales, D.M.; Bandyopadhyay, M.; Chen, Z.; Tkacheva, O.; Shufesky, W.J.; Wallace, C.T.; Watkins, S.C.; Berger, A.; et al. Neurokinin-1 Receptor Signaling Is Required for Efficient Ca2+ Flux in T-Cell-Receptor-Activated T Cells. Cell Rep. 2020, 30, 3448–3465.e8. [Google Scholar] [CrossRef] [Green Version]
- García-Aranda, M.; Téllez, T.; McKenna, L.; Redondo, M. Neurokinin-1 Receptor (NK-1R) Antagonists as a New Strategy to Overcome Cancer Resistance. Cancers 2022, 14, 2255. [Google Scholar] [CrossRef]
- O’Hayre, M.; Degese, M.S.; Gutkind, J.S. Novel Insights into G Protein and G Protein-Coupled Receptor Signaling in Cancer. Curr. Opin. Cell Biol. 2014, 27, 126–135. [Google Scholar] [CrossRef] [Green Version]
- Morishita, R.; Ueda, H.; Ito, H.; Takasaki, J.; Nagata, K.-I.; Asano, T. Involvement of Gq/11 in Both Integrin Signal-Dependent and -Independent Pathways Regulating Endothelin-Induced Neural Progenitor Proliferation. Neurosci. Res. 2007, 59, 205–214. [Google Scholar] [CrossRef]
- Guard, S.; Watson, S.P. Tachykinin Receptor Types: Classification and Membrane Signalling Mechanisms. Neurochem. Int. 1991, 18, 149–165. [Google Scholar] [CrossRef]
- Ye, R.D. Regulation of Nuclear Factor KappaB Activation by G-Protein-Coupled Receptors. J. Leukoc. Biol. 2001, 70, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Tsybko, A.S.; Ilchibaeva, T.V.; Popova, N.K. Role of Glial Cell Line-Derived Neurotrophic Factor in the Pathogenesis and Treatment of Mood Disorders. Rev. Neurosci. 2017, 28, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Schwindinger, W.F.; Robishaw, J.D. Heterotrimeric G-Protein Βγ-Dimers in Growth and Differentiation. Oncogene 2001, 20, 1653–1660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raddatz, R.; Crankshaw, C.L.; Snider, R.M.; Krause, J.E. Similar Rates of Phosphatidylinositol Hydrolysis Following Activation of Wild-Type and Truncated Rat Neurokinin-1 Receptors. J. Neurochem. 2002, 64, 1183–1191. [Google Scholar] [CrossRef]
- Torrens, Y.; Daguet De Montety, M.C.; el Etr, M.; Beaujouan, J.C.; Glowinski, J. Tachykinin Receptors of the NK1 Type (Substance P) Coupled Positively to Phospholipase C on Cortical Astrocytes from the Newborn Mouse in Primary Culture. J. Neurochem. 1989, 52, 1913–1918. [Google Scholar] [CrossRef]
- Sun, L.; Yu, F.; Ullah, A.; Hubrack, S.; Daalis, A.; Jung, P.; Machaca, K. Endoplasmic Reticulum Remodeling Tunes IP3-Dependent Ca2+ Release Sensitivity. PLoS ONE 2011, 6, e27928. [Google Scholar] [CrossRef] [Green Version]
- Meshki, J.; Douglas, S.D.; Lai, J.-P.; Schwartz, L.; Kilpatrick, L.E.; Tuluc, F. Neurokinin 1 Receptor Mediates Membrane Blebbing in HEK293 Cells through a Rho/Rho-Associated Coiled-Coil Kinase-Dependent Mechanism. J. Biol. Chem. 2009, 284, 9280–9289. [Google Scholar] [CrossRef] [Green Version]
- Kelly, P.; Moeller, B.J.; Juneja, J.; Booden, M.A.; Der, C.J.; Daaka, Y.; Dewhirst, M.W.; Fields, T.A.; Casey, P.J. The G12 Family of Heterotrimeric G Proteins Promotes Breast Cancer Invasion and Metastasis. Proc. Natl. Acad. Sci. USA 2006, 103, 8173–8178. [Google Scholar] [CrossRef] [Green Version]
- Kelly, P.; Stemmle, L.N.; Madden, J.F.; Fields, T.A.; Daaka, Y.; Casey, P.J. A Role for the G12 Family of Heterotrimeric G Proteins in Prostate Cancer Invasion. J. Biol. Chem. 2006, 281, 26483–26490. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Recio, S.; Gascón, P. Biological and Pharmacological Aspects of the NK1-Receptor. BioMed Res. Int. 2015, 2015, 495704. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Kugimiya, T.; Miyazaki, T. Substance P Receptor in U373 MG Human Astrocytoma Cells Activates Mitogen-Activated Protein Kinases ERK1/2 through Src. Brain Tumor Pathol. 2005, 22, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wong, Y.H.; Federman, A.; Pace, A.M.; Zachary, I.; Evans, T.; Pouysségur, J.; Bourne, H.R. Mutant Alpha Subunits of Gi2 Inhibit Cyclic AMP Accumulation. Nature 1991, 351, 63–65. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.C.; Huang, J.; Ali, S.; Lowry, W.; Huang, X.Y. Src Tyrosine Kinase Is a Novel Direct Effector of G Proteins. Cell 2000, 102, 635–646. [Google Scholar] [CrossRef] [Green Version]
- Ding, H.; Zhang, X.; Su, Y.; Jia, C.; Dai, C. GNAS Promotes Inflammation-Related Hepatocellular Carcinoma Progression by Promoting STAT3 Activation. Cell. Mol. Biol. Lett. 2020, 25, 8. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.N.; Druey, K.M. Heterotrimeric G Protein Signaling: Role in Asthma and Allergic Inflammation. J. Allergy Clin. Immunol. 2002, 109, 592–602. [Google Scholar] [CrossRef]
- He, X.; Zhang, L.; Chen, Y.; Remke, M.; Shih, D.; Lu, F.; Wang, H.; Deng, Y.; Yu, Y.; Xia, Y.; et al. The G Protein α Subunit Gαs Is a Tumor Suppressor in Sonic Hedgehog-Driven Medulloblastoma. Nat. Med. 2014, 20, 1035–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, L.; Herrmann, V.; Hofer, J.K.; Insel, P.A. Beta-Adrenergic Receptor/CAMP-Mediated Signaling and Apoptosis of S49 Lymphoma Cells. Am. J. Physiol. Cell Physiol. 2000, 279, C1665–C1674. [Google Scholar] [CrossRef] [Green Version]
- Khawaja, A.M.; Rogers, D.F. Tachykinins: Receptor to Effector. Int. J. Biochem. Cell Biol. 1996, 28, 721–738. [Google Scholar] [CrossRef]
- Nakajima, Y.; Tsuchida, K.; Negishi, M.; Ito, S.; Nakanishi, S. Direct Linkage of Three Tachykinin Receptors to Stimulation of Both Phosphatidylinositol Hydrolysis and Cyclic AMP Cascades in Transfected Chinese Hamster Ovary Cells. J. Biol. Chem. 1992, 267, 2437–2442. [Google Scholar] [CrossRef]
- Ng, C.Y.; Kannan, S.; Chen, Y.J.; Tan, F.C.K.; Ong, W.Y.; Go, M.L.; Verma, C.S.; Low, C.-M.; Lam, Y. A New Generation of Arachidonic Acid Analogues as Potential Neurological Agent Targeting Cytosolic Phospholipase A2. Sci. Rep. 2017, 7, 13683. [Google Scholar] [CrossRef]
- Azzi, A.; Boscoboinik, D.; Hensey, C. The Protein Kinase C Family. Eur. J. Biochem. 1992, 208, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Schaechter, J.; Benowitz, L. Activation of Protein Kinase C by Arachidonic Acid Selectively Enhances the Phosphorylation of GAP-43 in Nerve Terminal Membranes. J. Neurosci. 1993, 13, 4361–4371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonelli, F.; Lasagni Vitar, R.M.; Merlo Pich, F.G.; Fonteyne, P.; Rama, P.; Mondino, A.; Ferrari, G. Corneal Endothelial Cell Reduction and Increased Neurokinin-1 Receptor Expression in a Graft-versus-Host Disease Preclinical Model. Exp. Eye Res. 2022, 220, 109128. [Google Scholar] [CrossRef] [PubMed]
- Roux, S.L.; Borbely, G.; Słoniecka, M.; Backman, L.J.; Danielson, P. Transforming Growth Factor Beta 1 Modulates the Functional Expression of the Neurokinin-1 Receptor in Human Keratocytes. Curr. Eye Res. 2016, 41, 1035–1043. [Google Scholar] [CrossRef] [Green Version]
- Gaddipati, S.; Rao, P.; Jerome, A.D.; Burugula, B.B.; Gerard, N.P.; Suvas, S. Loss of Neurokinin-1 Receptor Alters Ocular Surface Homeostasis and Promotes an Early Development of Herpes Stromal Keratitis. J. Immunol. 2016, 197, 4021–4033. [Google Scholar] [CrossRef] [Green Version]
- Edvinsson, J.C.; Reducha, P.V.; Sheykhzade, M.; Warfvinge, K.; Haanes, K.A.; Edvinsson, L. Neurokinins and Their Receptors in the Rat Trigeminal System: Differential Localization and Release with Implications for Migraine Pain. Mol. Pain 2021, 17, 17448069211059400. [Google Scholar] [CrossRef]
- Lee, S.J.; Im, S.-T.; Wu, J.; Cho, C.S.; Jo, D.H.; Chen, Y.; Dana, R.; Kim, J.H.; Lee, S.-M. Corneal Lymphangiogenesis in Dry Eye Disease Is Regulated by Substance P/Neurokinin-1 Receptor System through Controlling Expression of Vascular Endothelial Growth Factor Receptor 3. Ocul. Surf. 2021, 22, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Lasagni Vitar, R.M.; Bonelli, F.; Atay, A.; Triani, F.; Fonteyne, P.; Di Simone, E.; Rama, P.; Mondino, A.; Ferrari, G. Topical Neurokinin-1 Receptor Antagonist Fosaprepitant Ameliorates Ocular Graft-versus-Host Disease in a Preclinical Mouse Model. Exp. Eye Res. 2021, 212, 108825. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Ogata, M.; Kawai, M.; Mashima, Y.; Nishida, T. Substance P in Human Tears. Cornea 2003, 22, S48–S54. [Google Scholar] [CrossRef]
- Yamada, M.; Ogata, M.; Kawai, M.; Mashima, Y.; Nishida, T. Substance P and Its Metabolites in Normal Human Tears. Investig. Ophthalmol. Vis. Sci. 2002, 43, 2622–2625. [Google Scholar]
- Suvas, S. Role of Substance P Neuropeptide in Inflammation, Wound Healing, and Tissue Homeostasis. J. Immunol. 2017, 199, 1543–1552. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Lal, G. Neurokinin Receptors and Their Implications in Various Autoimmune Diseases. Curr. Res. Immunol. 2021, 2, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, H.; Kobayashi, M.; Wakita, D.; Nishimura, T. Neuropeptide Signaling Activates Dendritic Cell-Mediated Type 1 Immune Responses through Neurokinin-2 Receptor. J. Immunol. Baltim. Md. 1950 2012, 188, 4200–4208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, J.M.; Mitchell, D.; Morton, I.K.M. Typical and Atypical NK1 Tachykinin Receptor Characteristics in the Rabbit Isolated Iris Sphincter. Br. J. Pharmacol. 1994, 112, 985–991. [Google Scholar] [CrossRef] [Green Version]
- Catalani, E.; Gangitano, C.; Bosco, L.; Casini, G. Expression of the Neurokinin 1 Receptor in the Mouse Retina. Neuroscience 2004, 128, 519–530. [Google Scholar] [CrossRef]
- Casini, G.; Rickman, D.W.; Sternini, C.; Brecha, N.C. Neurokinin 1 Receptor Expression in the Rat Retina. J. Comp. Neurol. 1997, 389, 496–507. [Google Scholar] [CrossRef]
- Casini, G.; Dal Monte, M.; Fornai, F.; Bosco, L.; Willems, D.; Yang, Q.; Zhou, Z.J.; Bagnoli, P. Neurokinin 1 Receptor Expression and Substance P Physiological Actions Are Developmentally Regulated in the Rabbit Retina. Neuroscience 2004, 124, 147–160. [Google Scholar] [CrossRef]
- Hwang, D.D.-J.; Lee, S.-J.; Kim, J.-H.; Lee, S.-M. The Role of Neuropeptides in Pathogenesis of Dry Dye. J. Clin. Med. 2021, 10, 4248. [Google Scholar] [CrossRef]
- Yu, M.; Lee, S.-M.; Lee, H.; Amouzegar, A.; Nakao, T.; Chen, Y.; Dana, R. Neurokinin-1 Receptor Antagonism Ameliorates Dry Eye Disease by Inhibiting Antigen-Presenting Cell Maturation and T Helper 17 Cell Activation. Am. J. Pathol. 2020, 190, 125–133. [Google Scholar] [CrossRef]
- Liu, L.; Dana, R.; Yin, J. Sensory Neurons Directly Promote Angiogenesis in Response to Inflammation via Substance P Signaling. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2020, 34, 6229–6243. [Google Scholar] [CrossRef] [Green Version]
- Bignami, F.; Giacomini, C.; Lorusso, A.; Aramini, A.; Rama, P.; Ferrari, G. NK1 Receptor Antagonists as a New Treatment for Corneal Neovascularization. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6783–6794. [Google Scholar] [CrossRef] [PubMed]
- Ljubimov, A.V.; Saghizadeh, M. Progress in Corneal Wound Healing. Prog. Retin. Eye Res. 2015, 49, 17–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labetoulle, M.; Baudouin, C.; Calonge, M.; Merayo-Lloves, J.; Boboridis, K.G.; Akova, Y.A.; Aragona, P.; Geerling, G.; Messmer, E.M.; Benítez-del-Castillo, J. Role of Corneal Nerves in Ocular Surface Homeostasis and Disease. Acta Ophthalmol. 2019, 97, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Zoukhri, D. Effect of Inflammation on Lacrimal Gland Function. Exp. Eye Res. 2006, 82, 885–898. [Google Scholar] [CrossRef] [Green Version]
- Lasagni Vitar, R.M.; Barbariga, M.; Fonteyne, P.; Bignami, F.; Rama, P.; Ferrari, G. Modulating Ocular Surface Pain Through Neurokinin-1 Receptor Blockade. Investig. Opthalmol. Vis. Sci. 2021, 62, 26. [Google Scholar] [CrossRef]
- Nishida, T.; Chikama, T.-I.; Morishige, N.; Yanai, R.; Yamada, N.; Saito, J. Persistent Epithelial Defects Due to Neurotrophic Keratopathy Treated with a Substance P-Derived Peptide and Insulin-like Growth Factor 1. Jpn. J. Ophthalmol. 2007, 51, 442–447. [Google Scholar] [CrossRef]
- Nishida, T.; Inui, M.; Nomizu, M. Peptide Therapies for Ocular Surface Disturbances Based on Fibronectin–Integrin Interactions. Prog. Retin. Eye Res. 2015, 47, 38–63. [Google Scholar] [CrossRef] [Green Version]
- Bonini, S.; Rama, P.; Olzi, D.; Lambiase, A. Neurotrophic Keratitis. Eye 2003, 17, 989–995. [Google Scholar] [CrossRef] [Green Version]
- Dua, H.S.; Said, D.G.; Messmer, E.M.; Rolando, M.; Benitez-Del-Castillo, J.M.; Hossain, P.N.; Shortt, A.J.; Geerling, G.; Nubile, M.; Figueiredo, F.C.; et al. Neurotrophic Keratopathy. Prog. Retin. Eye Res. 2018, 66, 107–131. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, M.; Nagano, T.; Chikama, T.; Nishida, T. Up-Regulation of Phosphorylation of Focal Adhesion Kinase and Paxillin by Combination of Substance P and IGF-1 in SV-40 Transformed Human Corneal Epithelial Cells. Biochem. Biophys. Res. Commun. 1998, 242, 16–20. [Google Scholar] [CrossRef]
- Nishida, T.; Nakamura, M.; Ofuji, K.; Reid, T.W.; Mannis, M.J.; Murphy, C.J. Synergistic Effects of Substance P with Insulin-like Growth Factor-1 on Epithelial Migration of the Cornea. J. Cell. Physiol. 1996, 169, 159–166. [Google Scholar] [CrossRef]
- Palmade, F.; Sechoy-Chambon, O.; Coquelet, C.; Bonne, C. Insulin-like Growth Factor-1 (IGF-1) Specifically Binds to Bovine Lens Epithelial Cells and Increases the Number of Fibronectin Receptor Sites. Curr. Eye Res. 1994, 13, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Lasagni Vitar, R.; Triani, F.; Barbariga, M.; Fonteyne, P.; Rama, P.; Ferrari, G. Substance P/Neurokinin-1 Receptor Pathway Blockade Ameliorates Limbal Stem Cell Deficiency by Modulating MTOR Pathway and Preventing Cell Senescence. Stem Cell Rep. 2022, 17, 849–863. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Nakayasu, K.; Iwatsu, M.; Kanai, A. Endogenous Substance P in Corneal Epithelial Cells and Keratocytes. Jpn. J. Ophthalmol. 2002, 46, 616–620. [Google Scholar] [CrossRef]
- Mansoor, H.; Tan, H.C.; Lin, M.T.-Y.; Mehta, J.S.; Liu, Y.-C. Diabetic Corneal Neuropathy. J. Clin. Med. 2020, 9, 3956. [Google Scholar] [CrossRef]
- Zhou, T.; Lee, A.; Lo, A.C.Y.; Kwok, J.S.W.J. Diabetic Corneal Neuropathy: Pathogenic Mechanisms and Therapeutic Strategies. Front. Pharmacol. 2022, 13, 816062. [Google Scholar] [CrossRef]
- Dogru, M.; Katakami, C.; Inoue, M. Tear Function and Ocular Surface Changes in Noninsulin-Dependent Diabetes Mellitus. Ophthalmology 2001, 108, 586–592. [Google Scholar] [CrossRef]
- Choi, J.E.; Di Nardo, A. Skin Neurogenic Inflammation. Semin. Immunopathol. 2018, 40, 249–259. [Google Scholar] [CrossRef]
- Corrigan, F.; Vink, R.; Turner, R.J. Inflammation in Acute CNS Injury: A Focus on the Role of Substance P: Neurogenic Inflammation in Acute CNS Injury. Br. J. Pharmacol. 2016, 173, 703–715. [Google Scholar] [CrossRef] [Green Version]
- Barbariga, M.; Fonteyne, P.; Ostadreza, M.; Bignami, F.; Rama, P.; Ferrari, G. Substance P Modulation of Human and Murine Corneal Neovascularization. Investig. Opthalmology Vis. Sci. 2018, 59, 1305–1312. [Google Scholar] [CrossRef]
- Ziche, M.; Morbidelli, L.; Pacini, M.; Geppetti, P.; Alessandri, G.; Maggi, C.A. Substance P Stimulates Neovascularization in Vivo and Proliferation of Cultured Endothelial Cells. Microvasc. Res. 1990, 40, 264–278. [Google Scholar] [CrossRef]
- Greeno, E.W.; Mantyh, P.; Vercellotti, G.M.; Moldow, C.F. Functional Neurokinin 1 Receptors for Substance P Are Expressed by Human Vascular Endothelium. J. Exp. Med. 1993, 177, 1269–1276. [Google Scholar] [CrossRef] [PubMed]
- Kohara, H.; Tajima, S.; Yamamoto, M.; Tabata, Y. Angiogenesis Induced by Controlled Release of Neuropeptide Substance P. Biomaterials 2010, 31, 8617–8625. [Google Scholar] [CrossRef] [PubMed]
- Gálvez, B.G.; Martinez-Perez, C.; Villa-Collar, C.; Alvarez-Peregrina, C.; Sánchez-Tena, M.Á. Influence of Cytokines on Inflammatory Eye Diseases: A Citation Network Study. J. Clin. Med. 2022, 11, 661. [Google Scholar] [CrossRef]
- Wakefield, D.; Lloyd, A. The Role of Cytokines in the Pathogenesis of Inflammatory Eye Disease. Cytokine 1992, 4, 1–5. [Google Scholar] [CrossRef]
- O’Connor, T.M.; O’Connell, J.; O’Brien, D.I.; Goode, T.; Bredin, C.P.; Shanahan, F. The Role of Substance P in Inflammatory Disease. J. Cell. Physiol. 2004, 201, 167–180. [Google Scholar] [CrossRef]
- Mashaghi, A.; Marmalidou, A.; Tehrani, M.; Grace, P.M.; Pothoulakis, C.; Dana, R. Neuropeptide Substance P and the Immune Response. Cell. Mol. Life Sci. CMLS 2016, 73, 4249–4264. [Google Scholar] [CrossRef] [Green Version]
- Li, J.J.; Yi, S.; Wei, L. Ocular Microbiota and Intraocular Inflammation. Front. Immunol. 2020, 11, 609765. [Google Scholar] [CrossRef]
- Kumar, A.; Yu, F.-S.X. Toll-like Receptors and Corneal Innate Immunity. Curr. Mol. Med. 2006, 6, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Akpek, E.K.; Gottsch, J.D. Immune Defense at the Ocular Surface. Eye 2003, 17, 949–956. [Google Scholar] [CrossRef]
- Tran, M.T.; Lausch, R.N.; Oakes, J.E. Substance P Differentially Stimulates IL-8 Synthesis in Human Corneal Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3871–3877. [Google Scholar]
- Hamrah, P.; Huq, S.O.; Liu, Y.; Zhang, Q.; Dana, M.R. Corneal Immunity Is Mediated by Heterogeneous Population of Antigen-Presenting Cells. J. Leukoc. Biol. 2003, 74, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Janelsins, B.M.; Sumpter, T.L.; Tkacheva, O.A.; Rojas-Canales, D.M.; Erdos, G.; Mathers, A.R.; Shufesky, W.J.; Storkus, W.J.; Falo, L.D.; Morelli, A.E.; et al. Neurokinin-1 Receptor Agonists Bias Therapeutic Dendritic Cells to Induce Type 1 Immunity by Licensing Host Dendritic Cells to Produce IL-12. Blood 2013, 121, 2923–2933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taketani, Y.; Marmalidou, A.; Dohlman, T.H.; Singh, R.B.; Amouzegar, A.; Chauhan, S.K.; Chen, Y.; Dana, R. Restoration of Regulatory T-Cell Function in Dry Eye Disease by Antagonizing Substance P/Neurokinin-1 Receptor. Am. J. Pathol. 2020, 190, 1859–1866. [Google Scholar] [CrossRef] [PubMed]
- Cruzat, A.; Qazi, Y.; Hamrah, P. In Vivo Confocal Microscopy of Corneal Nerves in Health and Disease. Ocul. Surf. 2017, 15, 15–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Y.W.; Lee, J.L.; Kang, H.G.; Gu, N.; Byun, H.; Yeo, A.; Noh, H.; Kim, S.; Choi, E.Y.; Song, J.S.; et al. Corneal Lymphangiogenesis Facilitates Ocular Surface Inflammation and Cell Trafficking in Dry Eye Disease. Ocul. Surf. 2018, 16, 306–313. [Google Scholar] [CrossRef]
- Fakih, D.; Zhao, Z.; Nicolle, P.; Reboussin, E.; Joubert, F.; Luzu, J.; Labbé, A.; Rostène, W.; Baudouin, C.; Mélik Parsadaniantz, S.; et al. Chronic Dry Eye Induced Corneal Hypersensitivity, Neuroinflammatory Responses, and Synaptic Plasticity in the Mouse Trigeminal Brainstem. J. Neuroinflamm. 2019, 16, 268. [Google Scholar] [CrossRef] [Green Version]
- Launay, P.-S.; Reboussin, E.; Liang, H.; Kessal, K.; Godefroy, D.; Rostene, W.; Sahel, J.-A.; Baudouin, C.; Melik Parsadaniantz, S.; Reaux Le Goazigo, A. Ocular Inflammation Induces Trigeminal Pain, Peripheral and Central Neuroinflammatory Mechanisms. Neurobiol. Dis. 2016, 88, 16–28. [Google Scholar] [CrossRef] [Green Version]
- Clements, J.L.; Dana, R. Inflammatory Corneal Neovascularization: Etiopathogenesis. Semin. Ophthalmol. 2011, 26, 235–245. [Google Scholar] [CrossRef]
- Lasagni Vitar, R.M.; Triolo, G.; Fonteyne, P.; Acuti Martellucci, C.; Manzoli, L.; Rama, P.; Ferrari, G. Epidemiology of Corneal Neovascularization and Its Impact on Visual Acuity and Sensitivity: A 14-Year Retrospective Study. Front. Med. 2021, 8, 733538. [Google Scholar] [CrossRef]
- Dana, M.R.; Schaumberg, D.A.; Kowal, V.O.; Goren, M.B.; Rapuano, C.J.; Laibson, P.R.; Cohen, E.J. Corneal Neovascularization after Penetrating Keratoplasty. Cornea 1995, 14, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Lucas, K.; Karamichos, D.; Mathew, R.; Zieske, J.D.; Stein-Streilein, J. Retinal Laser Burn-Induced Neuropathy Leads to Substance P-Dependent Loss of Ocular Immune Privilege. J. Immunol. 2012, 189, 1237–1242. [Google Scholar] [CrossRef]
- Paunicka, K.J.; Mellon, J.; Robertson, D.; Petroll, M.; Brown, J.R.; Niederkorn, J.Y. Severing Corneal Nerves in One Eye Induces Sympathetic Loss of Immune Privilege and Promotes Rejection of Future Corneal Allografts Placed in Either Eye: Corneal Nerves and Corneal Graft Rejection. Am. J. Transplant. 2015, 15, 1490–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puri, S.; Kenyon, B.M.; Hamrah, P. Immunomodulatory Role of Neuropeptides in the Cornea. Biomedicines 2022, 10, 1985. [Google Scholar] [CrossRef]
- Neelam, S.; Niederkorn, J.Y. Corneal Nerve Ablation Abolishes Ocular Immune Privilege by Downregulating CD103 on T Regulatory Cells. Investig. Ophthalmol. Vis. Sci. 2020, 61, 25. [Google Scholar] [CrossRef] [Green Version]
- Chui, J.; Di Girolamo, N.; Coroneo, M.T.; Wakefield, D. The Role of Substance P in the Pathogenesis of Pterygia. Investig. Opthalmology Vis. Sci. 2007, 48, 4482–4489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Girolamo, N.; Coroneo, M.; Wakefield, D. Epidermal Growth Factor Receptor Signaling Is Partially Responsible for the Increased Matrix Metalloproteinase-1 Expression in Ocular Epithelial Cells after UVB Radiation. Am. J. Pathol. 2005, 167, 489–503. [Google Scholar] [CrossRef] [Green Version]
- Nowosielski, Y.; Haas, G.; Seifarth, C.; Wohlfarter, W.; Tasan, R.; Verius, M.; Troger, J.; Bechrakis, N. The Involvement of NK1 and Y2 Receptor in the Development of Laser-Induced CNVs in C57Bl/6N Mice. Exp. Eye Res. 2018, 177, 87–95. [Google Scholar] [CrossRef]
- Gheorghe, A.; Mahdi, L.; Musat, O. AGE-RELATED MACULAR DEGENERATION. Rom. J. Ophthalmol. 2015, 59, 74–77. [Google Scholar]
- Hong, H.S.; Kim, S.; Kim, Y.H.; Park, J.H.; Jin, Y.; Son, Y. Substance-P Blocks Degeneration of Retina by Stimulating Migration and Proliferation of Retinal Pigmented Epithelial Cells. Tissue Eng. Regen. Med. 2015, 12, 121–127. [Google Scholar] [CrossRef]
- Hong, H.S.; Kim, S.; Nam, S.; Um, J.; Kim, Y.H.; Son, Y. Effect of Substance P on Recovery from Laser-Induced Retinal Degeneration. Wound Repair Regen. 2015, 23, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Troger, J.; Kremser, B.; Irschick, E.; Göttinger, W.; Kieselbach, G. Substance P in Proliferative Vitreoretinopathy: The Significance of Aqueous Humor Levels for Evolution of the Disease. Graefes Arch. Clin. Exp. Ophthalmol. 1998, 236, 900–903. [Google Scholar] [CrossRef] [PubMed]
- Nagasaki, H.; Shinagawa, K.; Mochizuki, M. Risk Factors for Proliferative Vitreoretinopathy. Prog. Retin. Eye Res. 1998, 17, 77–98. [Google Scholar] [CrossRef]
- Yoo, K.; Son, B.K.; Kim, S.; Son, Y.; Yu, S.-Y.; Hong, H.S. Substance P Prevents Development of Proliferative Vitreoretinopathy in Mice by Modulating TNF-α. Mol. Vis. 2017, 23, 933–943. [Google Scholar]
- Idrees, S.; Sridhar, J.; Kuriyan, A.E. Proliferative Vitreoretinopathy: A Review. Int. Ophthalmol. Clin. 2019, 59, 221–240. [Google Scholar] [CrossRef]
- Muñoz, M.; Rosso, M.; Coveñas, R.; Montero, I.; González-Moles, M.A.; Robles, M.J. Neurokinin-1 Receptors Located in Human Retinoblastoma Cell Lines: Antitumor Action of Its Antagonist, L-732,138. Investig. Opthalmology Vis. Sci. 2007, 48, 2775–2781. [Google Scholar] [CrossRef] [Green Version]
- Dyer, M.A.; Bremner, R. The Search for the Retinoblastoma Cell of Origin. Nat. Rev. Cancer 2005, 5, 91–101. [Google Scholar] [CrossRef]
- Muñoz, M.; Rosso, M.; Coveñas, R. A New Frontier in the Treatment of Cancer: NK-1 Receptor Antagonists. Curr. Med. Chem. 2010, 17, 504–516. [Google Scholar] [CrossRef]
- Muñoz, M.; Rosso, M.; Pérez, A.; Coveñas, R.; Rosso, R.; Zamarriego, C.; Soult, J.A.; Montero, I. Antitumoral Action of the Neurokinin-1-Receptor Antagonist L-733,060 and Mitogenic Action of Substance P on Human Retinoblastoma Cell Lines. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2567–2570. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, M.; Rosso, M. The NK-1 Receptor Antagonist Aprepitant as a Broad Spectrum Antitumor Drug. Investig. New Drugs 2010, 28, 187–193. [Google Scholar] [CrossRef]
- Groneberg, D.A.; Bielory, L.; Fischer, A.; Bonini, S.; Wahn, U. Animal Models of Allergic and Inflammatory Conjunctivitis. Allergy 2003, 58, 1101–1113. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, L.; Blanco, T.; Ge, H.; Xia, Y.; Pang, K.; Chen, Y.; Dana, R. Therapeutic Efficacy of Topical Blockade of Substance P in Experimental Allergic Red Eye. Ocul. Surf. 2022, 26, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Hollows, F.; Moran, D. Cataract--the Ultraviolet Risk Factor. Lancet Lond. Engl. 1981, 2, 1249–1250. [Google Scholar] [CrossRef]
- Gross, J.; Wegener, A.R.; Kronschläger, M.; Schönfeld, C.-L.; Holz, F.G.; Meyer, L.M. UVR-B-Induced NKR-1 Expression in Ocular Tissues Is Blocked by Substance P Receptor Antagonist Fosaprepitant in the Exposed as Well as Unexposed Partner Eye. Ocul. Immunol. Inflamm. 2021, 29, 963–975. [Google Scholar] [CrossRef]
- Bagga, B.; Kate, A.; Joseph, J.; Dave, V.P. Herpes Simplex Infection of the Eye: An Introduction. Community Eye Health 2020, 33, 68–70. [Google Scholar]
- Jerome, A.; Suvas, S. Discrepancy between Neurokinin 1 Receptor Antagonist Treatment and Neurokinin 1 Receptor Knockout Mice in the CD8 T Cell Response to Corneal HSV-1 Infection. J. Immunol. 2018, 200, 126.16. [Google Scholar]
- Poccardi, N.; Rousseau, A.; Haigh, O.; Takissian, J.; Naas, T.; Deback, C.; Trouillaud, L.; Issa, M.; Roubille, S.; Juillard, F.; et al. Herpes Simplex Virus 1 Replication, Ocular Disease, and Reactivations from Latency Are Restricted Unilaterally after Inoculation of Virus into the Lip. J. Virol. 2019, 93, e01586-19. [Google Scholar] [CrossRef]
- McKay, T.B.; Seyed-Razavi, Y.; Ghezzi, C.E.; Dieckmann, G.; Nieland, T.J.F.; Cairns, D.M.; Pollard, R.E.; Hamrah, P.; Kaplan, D.L. Corneal Pain and Experimental Model Development. Prog. Retin. Eye Res. 2019, 71, 88–113. [Google Scholar] [CrossRef]
- Puja, G.; Sonkodi, B.; Bardoni, R. Mechanisms of Peripheral and Central Pain Sensitization: Focus on Ocular Pain. Front. Pharmacol. 2021, 12, 764396. [Google Scholar] [CrossRef]
- Arcourt, A.; Gorham, L.; Dhandapani, R.; Prato, V.; Taberner, F.J.; Wende, H.; Gangadharan, V.; Birchmeier, C.; Heppenstall, P.A.; Lechner, S.G. Touch Receptor-Derived Sensory Information Alleviates Acute Pain Signaling and Fine-Tunes Nociceptive Reflex Coordination. Neuron 2017, 93, 179–193. [Google Scholar] [CrossRef] [Green Version]
- Julius, D.; Basbaum, A.I. Molecular Mechanisms of Nociception. Nature 2001, 413, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Galor, A.; Moein, H.-R.; Lee, C.; Rodriguez, A.; Felix, E.R.; Sarantopoulos, K.D.; Levitt, R.C. Neuropathic Pain and Dry Eye. Ocul. Surf. 2018, 16, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Skaper, S.D.; Facci, L.; Giusti, P. Mast Cells, Glia and Neuroinflammation: Partners in Crime? Immunology 2014, 141, 314–327. [Google Scholar] [CrossRef] [PubMed]
- von Hehn, C.A.; Baron, R.; Woolf, C.J. Deconstructing the Neuropathic Pain Phenotype to Reveal Neural Mechanisms. Neuron 2012, 73, 638–652. [Google Scholar] [CrossRef] [Green Version]
- Toth, M.; Jokić-Begić, N. Psychological Contribution to Understanding the Nature of Dry Eye Disease: A Cross-Sectional Study of Anxiety Sensitivity and Dry Eyes. Health Psychol. Behav. Med. 2020, 8, 202–219. [Google Scholar] [CrossRef]
- Dastjerdi, M.H.; Dana, R. Corneal Nerve Alterations in Dry Eye-Associated Ocular Surface Disease. Int. Ophthalmol. Clin. 2009, 49, 11–20. [Google Scholar] [CrossRef]
- Mehra, D.; Cohen, N.K.; Galor, A. Ocular Surface Pain: A Narrative Review. Ophthalmol. Ther. 2020, 9, 1–21. [Google Scholar] [CrossRef]
- Hagan, S.; Martin, E.; Enríquez-de-Salamanca, A. Tear Fluid Biomarkers in Ocular and Systemic Disease: Potential Use for Predictive, Preventive and Personalised Medicine. EPMA J. 2016, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Galor, A.; Hamrah, P.; Haque, S.; Attal, N.; Labetoulle, M. Understanding Chronic Ocular Surface Pain: An Unmet Need for Targeted Drug Therapy. Ocul. Surf. 2022, 26, 148–156. [Google Scholar] [CrossRef]
- Bignami, F.; Lorusso, A.; Rama, P.; Ferrari, G. Growth Inhibition of Formed Corneal Neovascularization Following Fosaprepitant Treatment. Acta Ophthalmol. 2017, 95, e641–e648. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.B.; Young, A.D.; Marriott, I. The Therapeutic Potential of Targeting Substance P/NK-1R Interactions in Inflammatory CNS Disorders. Front. Cell. Neurosci. 2017, 10, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, J.-P.; Ho, W.-Z.; Yang, J.-H.; Wang, X.; Song, L.; Douglas, S.D. A Non-Peptide Substance P Antagonist down-Regulates SP MRNA Expression in Human Mononuclear Phagocytes. J. Neuroimmunol. 2002, 128, 101–108. [Google Scholar] [CrossRef]
- Sharun, K.; Jambagi, K.; Arya, M.; Aakanksha, C.S.; Patel, P.K.; Dixit, S.K.; Dhama, K. Clinical Applications of Substance P (Neurokinin-1 Receptor) Antagonist in Canine Medicine. Arch. Razi Inst. 2021, 76, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Tang, S.; Li, J.; Wang, W.; Wu, M.; Zhan, X. Neurokinin-1 Receptor Antagonist Can Prevent the Delayed Phase in Patients: A Single Center Retrospect Study. Anticancer Agents Med. Chem. 2021, 21, 2192–2197. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, Y.; Zhang, Z.; Fang, W.; Kang, S.; Luo, Y.; Sheng, J.; Zhan, J.; Hong, S.; Huang, Y.; et al. Neurokinin-1 Receptor Antagonist-Based Triple Regimens in Preventing Chemotherapy-Induced Nausea and Vomiting: A Network Meta-Analysis. J. Natl. Cancer Inst. 2017, 109, djw217. [Google Scholar] [CrossRef] [Green Version]
- Jin, P.; Deng, S.; Sherchan, P.; Cui, Y.; Huang, L.; Li, G.; Lian, L.; Xie, S.; Lenahan, C.; Travis, Z.D.; et al. Neurokinin Receptor 1 (NK1R) Antagonist Aprepitant Enhances Hematoma Clearance by Regulating Microglial Polarization via PKC/P38MAPK/NFκB Pathway After Experimental Intracerebral Hemorrhage in Mice. Neurother. J. Am. Soc. Exp. Neurother. 2021, 18, 1922–1938. [Google Scholar] [CrossRef]
- Wilson, S.E.; Mohan, R.R.; Mohan, R.R.; Ambrósio, R.; Hong, J.; Lee, J. The Corneal Wound Healing Response: Cytokine-Mediated Interaction of the Epithelium, Stroma, and Inflammatory Cells. Prog. Retin. Eye Res. 2001, 20, 625–637. [Google Scholar] [CrossRef]
- Netto, M.V.; Mohan, R.R.; Ambrósio, R., Jr.; Hutcheon, A.E.K.; Zieske, J.D.; Wilson, S.E. Wound Healing in the Cornea: A Review of Refractive Surgery Complications and New Prospects for Therapy. Cornea 2005, 24, 509–522. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demirsoy, I.H.; Ferrari, G. The NK-1 Receptor Signaling: Distribution and Functional Relevance in the Eye. Receptors 2022, 1, 98-111. https://doi.org/10.3390/receptors1010006
Demirsoy IH, Ferrari G. The NK-1 Receptor Signaling: Distribution and Functional Relevance in the Eye. Receptors. 2022; 1(1):98-111. https://doi.org/10.3390/receptors1010006
Chicago/Turabian StyleDemirsoy, Ibrahim Halil, and Giulio Ferrari. 2022. "The NK-1 Receptor Signaling: Distribution and Functional Relevance in the Eye" Receptors 1, no. 1: 98-111. https://doi.org/10.3390/receptors1010006
APA StyleDemirsoy, I. H., & Ferrari, G. (2022). The NK-1 Receptor Signaling: Distribution and Functional Relevance in the Eye. Receptors, 1(1), 98-111. https://doi.org/10.3390/receptors1010006