
GPR56 mRNA Expression Is Modulated by Acute and Chronic Training Variable Manipulations in Resistance-Trained Men


 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
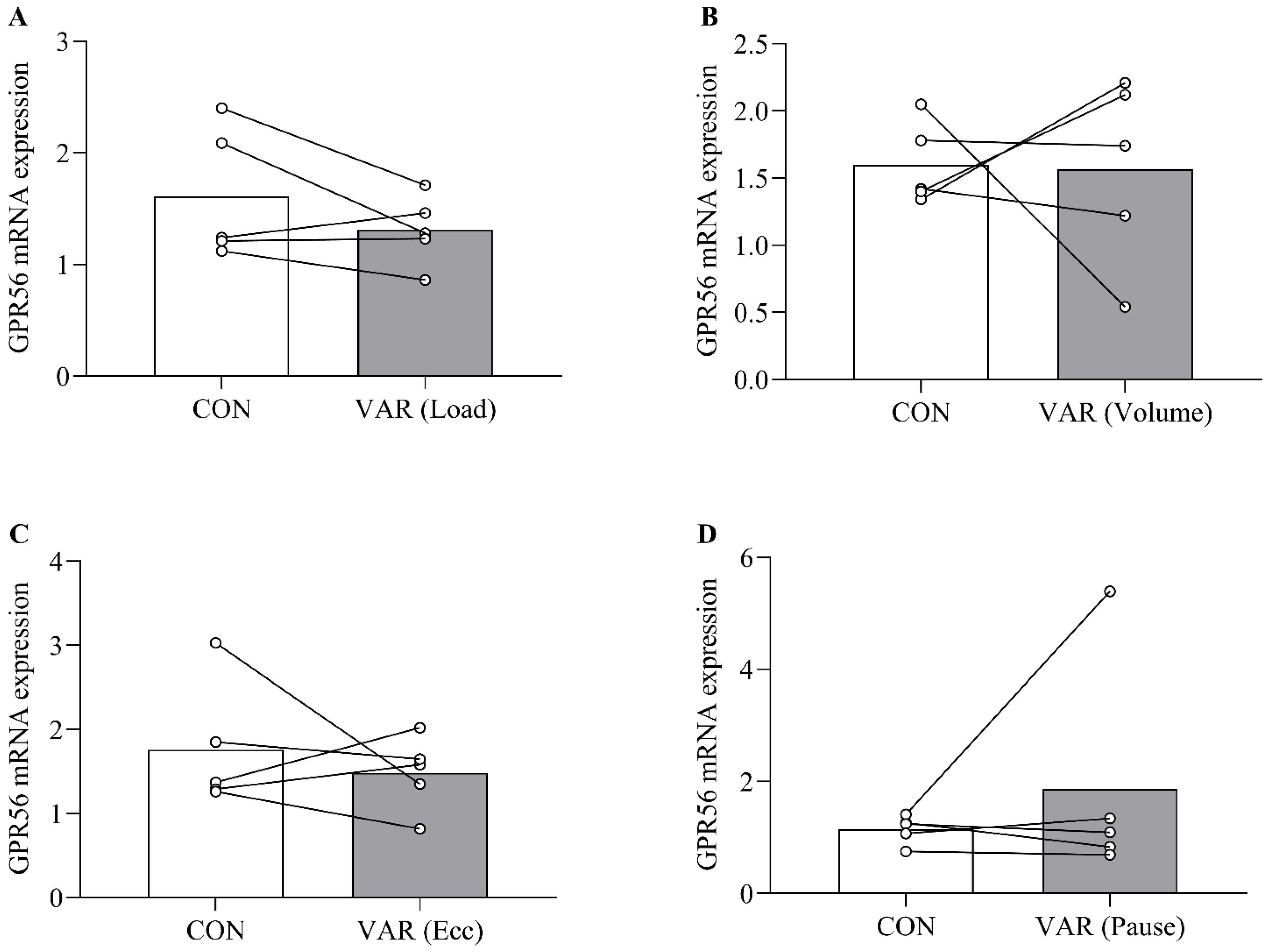
2.1. Acute GPR56 mRNA Expression
2.2. Chronic GPR56 mRNA Expression
2.3. Correlation Analysis
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Experimental Design
4.3. Muscle Biopsy
4.4. Resistance Training
4.5. qRT-PCR
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gordon, P.M.; Liu, D.; Sartor, M.A.; IglayReger, H.B.; Pistilli, E.E.; Gutmann, L.; Nader, G.A.; Hoffman, E. Resistance exercise training influences skeletal muscle immune activation: A microarray analysis. J. Appl. Physiol. 2012, 112, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Mackey, A.L.; Brandstetter, S.; Schjerling, P.; Bojsen-Moller, J.; Qvortrup, K.; Pedersen, M.M.; Doessing, S.; Kjaer, M.; Magnusson, S.P.; Langberg, H. Sequenced response of extracellular matrix deadhesion and fibrotic regulators after muscle damage is involved in protection against future injury in human skeletal muscle. FASEB J. 2011, 25, 1943–1959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahoney, D.J.; Safdar, A.; Parise, G.; Melov, S.; Fu, M.; MacNeil, L.; Kaczor, J.; Payne, E.T.; Tarnopolsky, M.A. Gene expression profiling in human skeletal muscle during recovery from eccentric exercise. Am. J. Physiol. Integr. Comp. Physiol. 2008, 294, R1901–R1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joisten, N.; Kummerhoff, F.; Koliamitra, C.; Schenk, A.; Walzik, D.; Hardt, L.; Knoop, A.; Thevis, M.; Kiesl, D.; Metcalfe, A.J.; et al. Exercise and the Kynurenine pathway: Current state of knowledge and results from a randomized cross-over study comparing acute effects of endurance and resistance training. Exerc. Immunol. Rev. 2020, 26, 24–42. [Google Scholar] [PubMed]
- Schoenfeld, B.J. Potential Mechanisms for a Role of Metabolic Stress in Hypertrophic Adaptations to Resistance Training. Sports Med. 2013, 43, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, V.; Caldow, M.; Massie, V.; Markworth, J.F.; Cameron-Smith, D.; Blazevich, A. Ribosome biogenesis adaptation in resistance training-induced human skeletal muscle hypertrophy. Am. J. Physiol. Metab. 2015, 309, E72–E83. [Google Scholar] [CrossRef] [PubMed]
- Mobley, C.B.; Haun, C.T.; Roberson, P.A.; Mumford, P.; Kephart, W.C.; Romero, M.A.; Osburn, S.C.; Vann, C.G.; Young, K.C.; Beck, D.T.; et al. Biomarkers associated with low, moderate, and high vastus lateralis muscle hypertrophy following 12 weeks of resistance training. PLoS ONE 2018, 13, e0195203. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.D.; Haun, C.T.; Mobley, C.B.; Mumford, P.W.; Romero, M.A.; Roberson, P.A.; Vann, C.G.; McCarthy, J.J. Physiological Differences Between Low Versus High Skeletal Muscle Hypertrophic Responders to Re-sistance Exercise Training: Current Perspectives and Future Research Directions. Front. Physiol. 2018, 9, 834. [Google Scholar] [CrossRef] [PubMed]
- Hornberger, T.A.; Esser, K.A. Mechanotransduction and the regulation of protein synthesis in skeletal muscle. Proc. Nutr. Soc. 2004, 63, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Hornberger, T.A. Mechanotransduction and the regulation of mTORC1 signaling in skeletal muscle. Int. J. Biochem. Cell Biol. 2011, 43, 1267–1276. [Google Scholar] [CrossRef] [Green Version]
- White, J.P.; Wrann, C.D.; Rao, R.R.; Nair, S.K.; Jedrychowski, M.P.; You, J.-S.; Martinez-Redondo, V.; Gygi, S.P.; Ruas, J.; Hornberger, T.; et al. G protein-coupled receptor 56 regulates mechanical overload-induced muscle hypertrophy. Proc. Natl. Acad. Sci. USA 2014, 111, 15756–15761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiaffino, S.; Reggiani, C.; Akimoto, T.; Blaauw, B. Molecular Mechanisms of Skeletal Muscle Hypertrophy. J. Neuromuscul. Dis. 2021, 8, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Kitakaze, T.; Yoshikawa, M.; Kobayashi, Y.; Kimura, N.; Goshima, N.; Ishikawa, T.; Ogata, Y.; Yamashita, Y.; Ashida, H.; Harada, N.; et al. Extracellular transglutaminase 2 induces myotube hypertrophy through G protein-coupled receptor 56. Biochim. Biophys. Acta-Bioenerg. 2020, 1867, 118563. [Google Scholar] [CrossRef] [PubMed]
- Pillon, N.J.; Gabriel, B.M.; Dollet, L.; Smith, J.A.B.; Puig, L.S.; Botella, J.; Bishop, D.J.; Krook, A.; Zierath, J.R. Transcriptomic profiling of skeletal muscle adaptations to exercise and inactivity. Nat. Commun. 2020, 11, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallinson, J.E.; Taylor, T.; Constantin-Teodosiu, D.; Billeter-Clark, R.; Constantin, D.; Franchi, M.V.; Narici, M.V.; Auer, D.; Greenhaff, P.L. Longitudinal hypertrophic and transcriptional responses to high-load eccentric-concentric vs concentric training in males. Scand. J. Med. Sci. Sports 2020, 30, 2101–2115. [Google Scholar] [CrossRef] [PubMed]
- Burd, N.; West, D.W.D.; Staples, A.W.; Atherton, P.J.; Baker, J.M.; Moore, D.; Holwerda, A.; Parise, G.; Rennie, M.J.; Baker, S.K.; et al. Low-Load High Volume Resistance Exercise Stimulates Muscle Protein Synthesis More Than High-Load Low Volume Resistance Exercise in Young Men. PLoS ONE 2010, 5, e12033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilborn, C.D.; Taylor, L.W.; Greenwood, M.; Kreider, R.B.; Willoughby, D.S. Effects of Different Intensities of Resistance Exercise on Regulators of Myogenesis. J. Strength Cond. Res. 2009, 23, 2179–2187. [Google Scholar] [CrossRef] [Green Version]
- Fox, C.D.; Mesquita, P.H.C.; Godwin, J.S.; Angleri, V.; Damas, F.; Ruple, B.A.; Sexton, C.L.; Brown, M.D.; Kavazis, A.N.; Young, K.C.; et al. Frequent Manipulation of Resistance Training Variables Promotes Myofibrillar Spacing Changes in Re-sistance-Trained Individuals. Front. Physiol. 2021, 12, 2239. [Google Scholar]
- Nederveen, J.P.; Joanisse, S.; Snijders, T.; Thomas, A.C.Q.; Kumbhare, D.; Parise, G. The influence of capillarization on satellite cell pool expansion and activation following exercise-induced muscle damage in healthy young men. J. Physiol. 2018, 596, 1063–1078. [Google Scholar] [CrossRef] [Green Version]
- Islam, H.; Edgett, B.; Bonafiglia, J.T.; Shulman, T.; Ma, A.; Quadrilatero, J.; Simpson, C.A.; Gurd, B.J. Repeatability of exercise-induced changes in mRNA expression and technical considerations for qPCR analysis in human skeletal muscle. Exp. Physiol. 2019, 104, 407–420. [Google Scholar] [CrossRef]
- Biazon, T.M.P.C.; Ugrinowitsch, C.; Soligon, S.D.; Oliveira, R.M.; Bergamasco, J.G.; Borghi-Silva, A.; Libardi, C.A. The Association Between Muscle Deoxygenation and Muscle Hypertrophy to Blood Flow Restricted Training Performed at High and Low Loads. Front. Physiol. 2019, 10, 446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Sequence | Reverse Sequence |
|---|---|---|
| RPLP0 | CGGTTTCTGATTGGCTAC | ACGATGTCACTTCCACG |
| RPL13A | GTCTGAAGCCTACAAGAAAG | TGTCAATTTTCTTCTCCACG |
| TFRC | AAGATTCAGGTCAAAGACAG | CTTACTATACGCCACATAACC |
| GAPDH | GATGCTGGTGCTGAGTATGTCG | GTGGTGCAGGATGCATTGCTGA |
| GPR-56 | CTGTAATATGGAAACTGGGGAAA | CCGTAGCTAAACTGAAAACCACC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angleri, V.; Damas, F.; Stotzer, U.S.; Selistre-de-Araujo, H.S.; Santanielo, N.; Soligon, S.D.; Riani Costa, L.A.; Lixandrão, M.E.; Conceição, M.S.; Vechin, F.C.; et al. GPR56 mRNA Expression Is Modulated by Acute and Chronic Training Variable Manipulations in Resistance-Trained Men. Muscles 2022, 1, 16-25. https://doi.org/10.3390/muscles1010002
Angleri V, Damas F, Stotzer US, Selistre-de-Araujo HS, Santanielo N, Soligon SD, Riani Costa LA, Lixandrão ME, Conceição MS, Vechin FC, et al. GPR56 mRNA Expression Is Modulated by Acute and Chronic Training Variable Manipulations in Resistance-Trained Men. Muscles. 2022; 1(1):16-25. https://doi.org/10.3390/muscles1010002
Chicago/Turabian StyleAngleri, Vitor, Felipe Damas, Uliana Sbeguen Stotzer, Heloisa Sobreiro Selistre-de-Araujo, Natalia Santanielo, Samuel Domingos Soligon, Luiz Augusto Riani Costa, Manoel Emílio Lixandrão, Miguel Soares Conceição, Felipe Cassaro Vechin, and et al. 2022. "GPR56 mRNA Expression Is Modulated by Acute and Chronic Training Variable Manipulations in Resistance-Trained Men" Muscles 1, no. 1: 16-25. https://doi.org/10.3390/muscles1010002
APA StyleAngleri, V., Damas, F., Stotzer, U. S., Selistre-de-Araujo, H. S., Santanielo, N., Soligon, S. D., Riani Costa, L. A., Lixandrão, M. E., Conceição, M. S., Vechin, F. C., Roberts, M. D., Ugrinowitsch, C., & Libardi, C. A. (2022). GPR56 mRNA Expression Is Modulated by Acute and Chronic Training Variable Manipulations in Resistance-Trained Men. Muscles, 1(1), 16-25. https://doi.org/10.3390/muscles1010002