
Tree-Related Microhabitats and Multi-Taxon Biodiversity Quantification Exploiting ALS Data

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
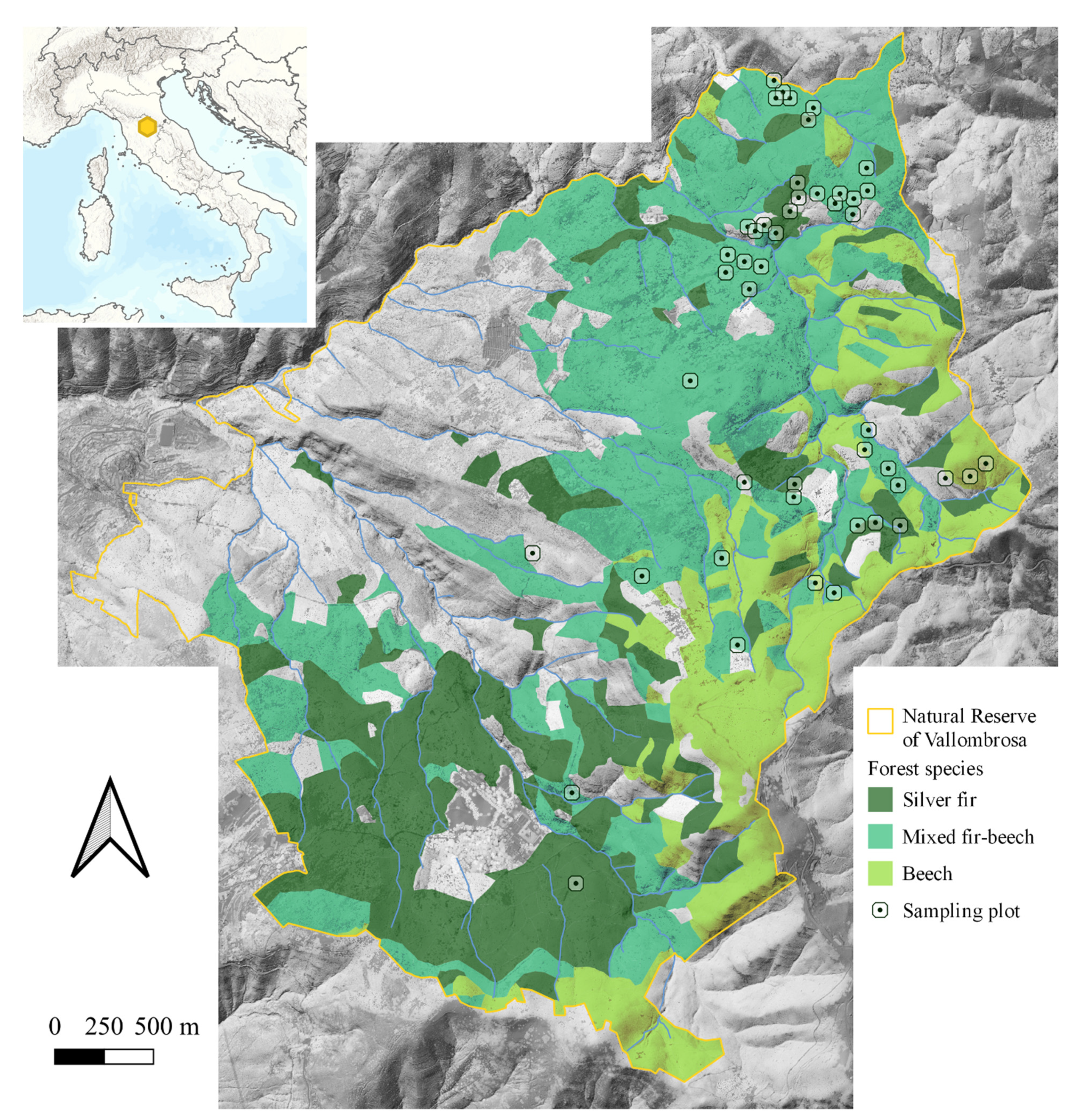
2.1. Study Area

2.2. Reference Data for Biodiversity Indices
2.2.1. Beetle Communities
2.2.2. Birds
2.2.3. Tree-Related Microhabitats
2.3. Biodiversity Indices
2.4. Predictor Variables for Modeling Biodiversity Indices
2.4.1. Airborne Laser Scanning Variables
2.4.2. Auxiliary Variables
2.5. Random Forests Models
2.6. Accuracy Assessment
3. Results
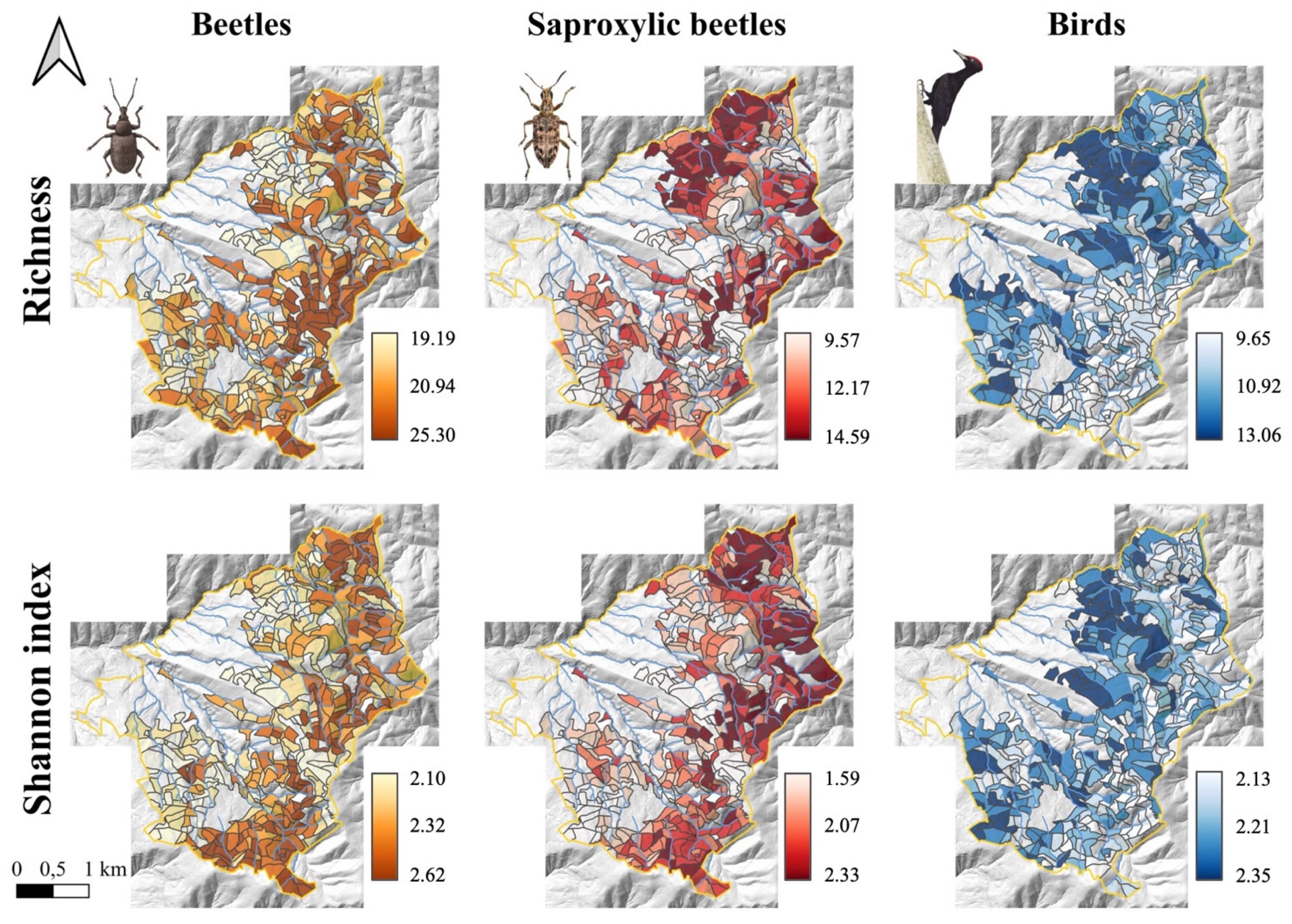
3.1. Saproxylic and Non-Saproxylic Beetles
3.2. Forest-Dwelling Birds
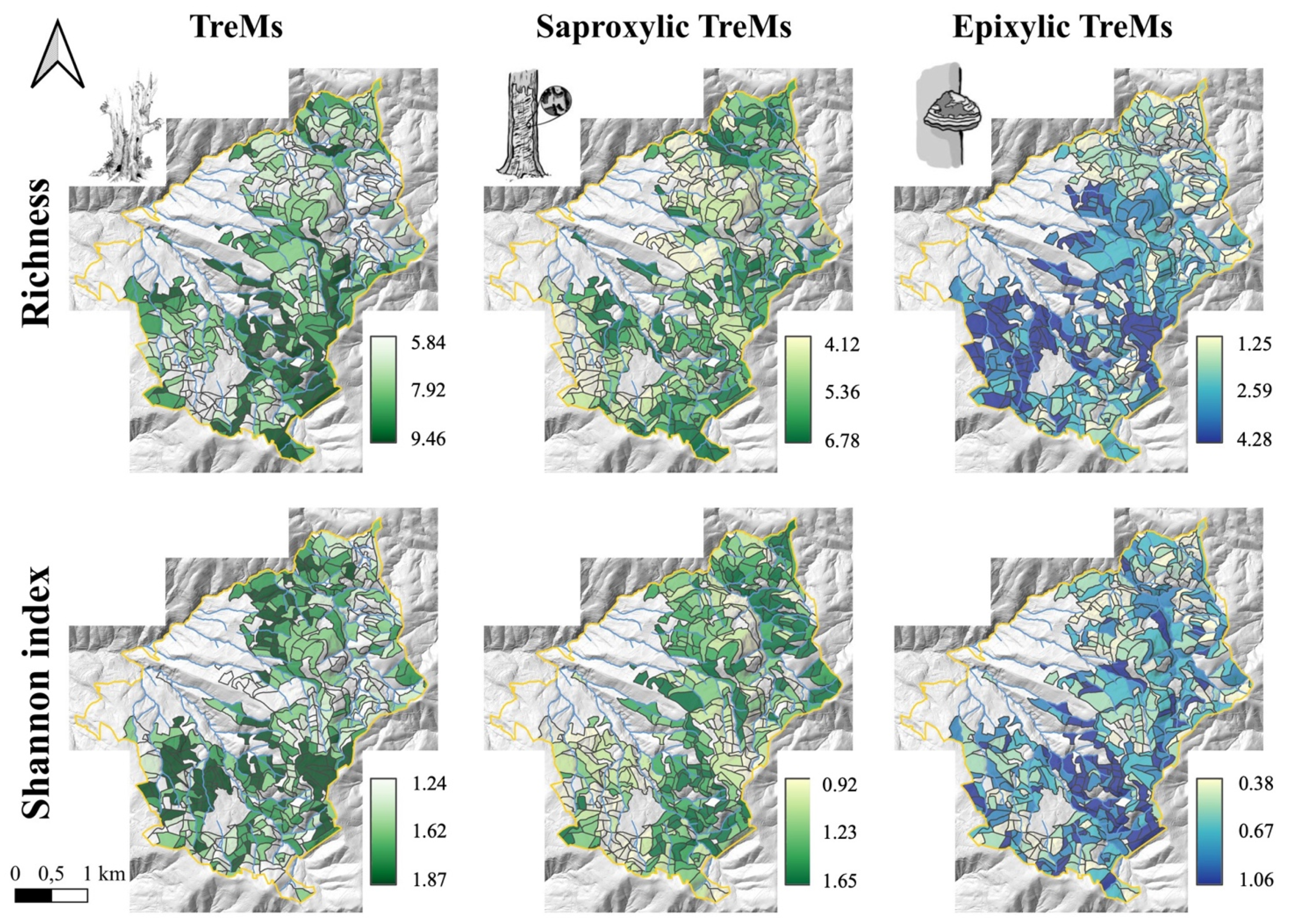
3.3. Tree-Related Microhabitats
3.4. Random Forests Models and Maps of Biodiversity Indices
4. Discussion
4.1. Relationship between ALS Data and Multi-Taxon Biodiversity
4.2. Multi-Taxon Biodiversity and Forest Management
4.3. Biodiversity Conservation
4.4. ALS Data, Limitations, and Opportunities
5. Conclusions
- (1)
- ALS data hold great potential for analyzing the relationship between forest structure and saproxylic and non-saproxylic beetle and bird communities. Forest structure and TreMs were the most important variables in determining the multi-taxon biodiversity in Mediterranean mountain ecosystems. Thus, accurate data on forest structure and microhabitat-type indicators emerged as crucial for forest management and biodiversity conservation.
- (2)
- Remote sensing provides powerful tools to study the diversity and abundance of forest biodiversity indicators (i.e., insects, birds, and TreMs). The large availability of data at different spatial and temporal resolutions allows saproxylic beetle and bird communities to be investigated at the most appropriate scale to discover new ecological, ethological, and conservation information.
- (3)
- Currently, ALS data can capture information on the composition and structure of ecologically suitable habitats for animal species. Habitat resources (trophic niches—TreMs) can be distinguished using the variables obtainable from point clouds. Furthermore, the different biodiversity conditions detected with the ground surveys were mapped at physiologically relevant scales for insects and birds.
- (4)
- In the near future, remote sensing will be increasingly used for specific indicators of forest biodiversity. Although limitations for fully successful implementation are still emerging, technological progress will make it possible to obtain information on threatened species, thus informing nature-based forest management.
- (5)
- In future studies, we suggest including other taxonomic groups related to forest structural traits (e.g., small mammals, spiders, amphibians, lichens, fungi, and bryophytes). This will ensure the comprehensive monitoring of forest ecosystems to identify biodiversity hotspots more effectively.
- (6)
- Multi-taxon biodiversity data permit the definition and strengthening of sustainable management indicators linked to the different functions in forest ecosystems. This is useful for drawing implications for conservation strategies of forest environments and for increasing the resilience of mountain forests threatened by climate change.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. State of the World’s Forests. Food and Agriculture Organization of the United Nations; FAO: Rome, Italy, 2012. [Google Scholar]
- Trentanovi, G.; Campagnaro, T.; Sitzia, T.; Chianucci, F.; Vacchiano, G.; Ammer, C.; Ciach, M.; Nagel, T.A.; del Río, M.; Paillet, Y.; et al. Words apart: Standardizing forestry terms and definitions across European biodiversity studies. For. Ecosyst. 2023, 10, 100128. [Google Scholar] [CrossRef]
- Oettel, J.; Lapin, K. Linking forest management and biodiversity indicators to strengthen sustainable forest management in Europe. Ecol. Indic. 2021, 122, 107275. [Google Scholar] [CrossRef]
- Leitão, P.J.; Toraño Caicoya, A.; Dahlkamp, A.; Guderjan, L.; Griesser, M.; Haverkamp, P.J.; Nordén, J.; Snäll, T.; Schröder, B. Impacts of Forest Management on Forest Bird Occurrence Patterns—A Case Study in Central Europe. Front. For. Glob. Chang. 2022, 5, 786556. [Google Scholar] [CrossRef]
- Kõrkjas, M.; Remm, L.; Lõhmus, P.; Lõhmus, A. From tree-related microhabitats to ecosystem management: A tree-scale investigation in productive forests in Estonia. J. Environ. Manag. 2023, 343, 118245. [Google Scholar] [CrossRef] [PubMed]
- Munro, N.T.; Fischer, J.; Barrett, G.; Wood, J.; Leavesley, A.; Lindenmayer, D.B.B. Bird’s response to revegetation of different structure and floristics e are “restoration plantings” restoring bird communities? Restor. Ecol. 2010, 19, 223–235. [Google Scholar] [CrossRef]
- Bae, S.; Müller, J.; Lee, D.; Vierling, K.T.; Vogeler, J.C.; Vierling, L.A.; Hudak, A.T.; Latifi, H.; Thorn, S. Taxonomic, functional, and phylogenetic diversity of bird assemblages are oppositely associated to productivity and heterogeneity in temperate forests. Remote Sens. Environ. 2018, 215, 145–156. [Google Scholar] [CrossRef]
- Campanaro, A.; Parisi, F. Open datasets wanted for tracking the insect decline: Let’s start from saproxylic beetles. Biodivers. Data J. 2021, 9, e72741. [Google Scholar] [CrossRef]
- Bunce, R.G.H.; Bogers, M.M.B.; Evans, D.; Halada, L.; Jongman, R.H.G.; Mucher, C.A.; Bauch, B.; de Bluste, G.; Parr, T.W.; Olsvig-Whittaker, L. The significance of habitats as indicators of biodiversity and their links to species. Ecol. Indic. 2013, 33, 19–25. [Google Scholar] [CrossRef]
- Lachat, T.; Wermelinger, B.; Gossner, M.M.; Bussler, H.; Isacsson, G.; Müller, J. Saproxylic beetles as indicator species for deadwood amount and temperature in European beech forests. Ecol. Indic. 2012, 23, 323–331. [Google Scholar] [CrossRef]
- Ram, D.; Axelsson, A.L.; Green, M.; Smith, H.G.; Lindström, Å. What drives current population trends in forest birds–forest quantity, quality or climate? A large-scale analysis from northern Europe. For. Ecol. Manag. 2017, 385, 177–188. [Google Scholar] [CrossRef]
- Roberge, J.; Angelstam, P. Indicator species among resident forest birds A cross-regional evaluation in northern Europe. Biol. Conserv. 2006, 130, 134–147. [Google Scholar] [CrossRef]
- Nadkarni, N.M.; McIntosh, A.C.; Cushing, J.B. A framework to categorize forest structure concepts. Forest Ecol. Manag. 2008, 256, 872–882. [Google Scholar] [CrossRef]
- Spina, P.; Parisi, F.; Antonucci, S.; Garfì, V.; Marchetti, M.; Santopuoli, G. Tree-related microhabitat diversity as a proxy for the conservation of beetle communities in managed forests of Fagus sylvatica. For. Int. J. For. Res. 2023, 97, 223–233. [Google Scholar] [CrossRef]
- Vergara, P.M.; Fierro, A.; Carvajal, M.A.; Alaniz, A.J.; Quiroz, M. Multiple environmental drivers for the Patagonian forest-dwelling beetles: Contrasting functional and taxonomic responses across strata and trophic guilds. Sci. Total Environ. 2022, 838, 155906. [Google Scholar] [CrossRef] [PubMed]
- Blondel, J. Origins and dynamics of forest birds of the northern hemisphere. In Ecology and Conservation of Forest Birds; Mikusinski, G., Roberge, J.M., Fuller, R.J., Eds.; Cambridge University Press: Cambridge, UK, 2018; pp. 11–50. [Google Scholar] [CrossRef]
- Kriegel, P.; Vogel, S.; Angeleri, R.; Baldrian, P.; Borken, W.; Bouget, C.; Brin, A.; Bussler, H.; Cocciufa, C.; Feldmann, B.; et al. Ambient and substrate energy influence decomposer diversity differentially across trophic levels. Ecol. Lett. 2023, 26, 1157–1173. [Google Scholar] [CrossRef] [PubMed]
- Gregory, R.D.; Van Strien, A.; Vorisek, P.; Gmelig Meyling, A.W.; Noble, D.G.; Foppen, R.P.; Gibbons, D.W. Developing indicators for European birds. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Storch, F.; Boch, S.; Gossner, M.M.; Feldhaar, H.; Ammer, C.; Schall, P.; Polle, A.; Kroiher, F.; Muller, J.; Bauhus, J. Linking structure and species richness to support forest biodiversity monitoring at large scales. Ann. For. Sci. 2023, 80, 3. [Google Scholar] [CrossRef]
- Hanle, J.; Duguid, M.C.; Ashton, M.S. Legacy forest structure increases bird diversity and abundance in aging young forests. Ecol. Evol. 2020, 10, 1193–1208. [Google Scholar] [CrossRef]
- Carpaneto, G.M.; Baviera, C.; Biscaccianti, A.B.; Brandmayr, P.; Mazzei, A.; Mason, F.; Battistoni, A.; Teofili, C.; Rondinini, C.; Fattorini, S.; et al. A Red List of Italian Saproxylic Beetles: Taxonomic overview, ecological features and conservation issues (Coleoptera). Fragm. Entomol. 2015, 47, 53–126. [Google Scholar] [CrossRef]
- Culbert, P.D.; Radeloff, V.C.; Flather, C.H.; Kellndorfer, J.M.; Rittenhouse, C.D.; Pidgeon, A.M. The influence of vertical and horizontal habitat structure on nationwide patterns of avian biodiversity. Ornithology 2013, 130, 656–665. [Google Scholar] [CrossRef]
- Gustin, M.; Nardelli, R.; Brichetti, P.; Battistoni, A.; Rondinini, C.; Teofili, C. Lista Rossa IUCN degli uccelli nidificanti in Italia. In Comitato Italiano IUCN e Ministero dell’Ambiente e della Tutela del Territorio e del Mare; Ufficio federale dell’ambiente (UFAM): Berna, Switzerland; Stazione ornitologica svizzera: Sempach, Switzerland, 2021. [Google Scholar]
- Lindén, S.F.A.; Lehikoinen, A. Population trends of common breeding forest birds in southern Finland are consistent with trends in forest management and climate change. Ornis Fenn. 2015, 92, 187–203. [Google Scholar]
- Vogeler, J.C.; Hudak, A.T.; Vierling, L.A.; Evans, J.; Green, P.; Vierling, K.T. Terrain and vegetation structural influences on local avian species richness in two mixed-conifer forests. Remote Sens. Environ. 2014, 147, 13–22. [Google Scholar] [CrossRef]
- Clawges, R.; Vierling, K.; Vierling, L.; Rowell, E. The use of airborne lidar to assess avian species diversity, density, and occurrence in a pine/aspen forest. Remote Sens. Environ. 2008, 112, 2064–2073. [Google Scholar] [CrossRef]
- Jacobsen, R.M.; Sverdrup-Thygeson, A.; Birkemoe, T. Scale-specific responses of saproxylic beetles: Combining dead wood surveys with data from satellite imagery. J. Insect Conserv. 2015, 19, 1053–1062. [Google Scholar] [CrossRef]
- Martinuzzi, S.; Vierling, L.A.; Gould, W.A.; Falkowski, M.J.; Evans, J.S.; Hudak, A.T.; Vierling, K.T. Mapping snags and understory shrubs for a LiDAR-based assessment of wildlife habitat suitability. Remote Sens. Environ. 2009, 113, 2533–2546. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, Y.; Pu, R.; Gonzalez-Moreno, P.; Yuan, L.; Wu, K.; Huang, W. Monitoring plant diseases and pests through remote sensing technology: A review. Comput. Electron. Agric. 2019, 165, 104943. [Google Scholar] [CrossRef]
- Filho, F.H.I.; Heldens, W.B.; Kong, Z.; de Lange, E.S. Drones: Innovative technology for use in precision pest management. J. Econ. Entomol. 2020, 113, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Toivonen, J.; Kangas, A.; Maltamo, M.; Kukkonen, M.; Packalen, P. Assessing biodiversity using forest structure indicators based on airborne laser scanning data. For. Ecol. Manag. 2023, 546, 121376. [Google Scholar] [CrossRef]
- Galbraith, S.M.; Vierling, L.A.; Bosque-Perez, N.A. Remote sensing and ecosystem services: Current status and future opportunities for the study of bees and pollination-related services. Curr. For. Rep. 2015, 1, 261–274. [Google Scholar] [CrossRef][Green Version]
- Rada, P.; Padilla, A.; Horák, J.; Micó, E. Public LiDAR data are an important tool for the detection of saproxylic insect hotspots in Mediterranean forests and their connectivity. For. Ecol. Manag. 2022, 520, 120378. [Google Scholar] [CrossRef]
- Kane, V.R.; McGaughey, R.J.; Bakker, J.D.; Gersonde, R.F.; Lutz, J.A.; Franklin, J.F. Comparisons between field- and LiDAR-based measures of stand structural complexity. Can. J. For. Res. 2010, 40, 761–773. [Google Scholar] [CrossRef]
- Müller, J.; Bae, S.; Röder, J.; Chao, A.; Didham, R.K. Airborne LiDAR reveals context dependence in the effects of canopy architecture on arthropod diversity. For. Ecol. Manag. 2014, 312, 129–137. [Google Scholar] [CrossRef]
- Bombi, P.; Gnetti, V.; D’Andrea, E.; De Cinti, B.; Vigna Taglianti, A.; Bologna, M.A.; Matteucci, G. Identifying priority sites for insect conservation in forest ecosystems at high resolution: The potential of LiDAR data. J. Insect Conserv. 2019, 23, 689–698. [Google Scholar] [CrossRef]
- North, M.P.; Kane, J.T.; Kane, V.R.; Asner, G.P.; Berigan, W.; Churchill, D.J.; Conway, S.; Gutiérrez, R.J.; Jeronimo, S.; Keane, J.; et al. Cover of tall trees best predicts California spotted owl habitat. For. Ecol. Manag. 2017, 405, 166–178. [Google Scholar] [CrossRef]
- Bottalico, F.; Travaglini, D.; Fiorentini, S.; Lisa, C.; Nocentini, S. Stand dynamics and natural regeneration in silver fir (Abies alba Mill.) plantations after traditional rotation age. iForest 2014, 7, 313–323. [Google Scholar] [CrossRef]
- Ciancio, O. Riserva Naturale Statale Biogenetica di Vallombrosa. Piano di Gestione e Silvomuseo 2006–2025; Corpo Forestale dello Stato, Ufficio Territoriale per la Biodiversità di Vallombrosa, Reggello (FI): Florence, Italy, 2009; pp. 113–134. ISBN 978-88-87553-17-8. [Google Scholar]
- Nocentini, S.; Ciancio, O.; Portoghesi, L.; Corona, P. Historical roots and the evolving science of forest management under a systemic perspective. Can. J. For. Res. 2021, 51, 163–171. [Google Scholar] [CrossRef]
- QGIS.org. QGIS Geographic Information System. 2024, QGIS Association. Available online: http://www.qgis.org (accessed on 2 January 2024).
- Bouchard, P.; Bousquet, Y.; Davies, A.E.; Alonso-Zarazaga, M.A.; Lawrence, J.F.; Lyal, C.H.C.; Newton, A.F.; Reid, C.A.M.; Schmitt, M.; Slipinski, S.A.; et al. Family-group names in Coleoptera (Insecta). ZooKeys 2011, 88, 1–972. [Google Scholar]
- Audisio, P.; Zarazaga, M.A.; Slipinski, A.; Nilsson, A.; Jelínek, J.; Taglianti, A.V.; Turco, F.; Otero, C.; Canepari, C.; Kral, D.; et al. Fauna Europaea: Coleoptera 2 (excl. series Elateriformia, Scarabaeiformia, Staphyliniformia and superfamily Curculionoidea). Biodivers. Data J. 2015, 3, e4750. [Google Scholar] [CrossRef]
- Bibby, C.J.; Burgess, N.D.; Hillis, D.M.; Hill, D.A.; Mustoe, S. Bird Census Techniques, 2nd ed; Academic Press: London, UK, 2000; ISBN 9780120958313. [Google Scholar]
- Baccetti, N.; Fracasso, G. CISO-COI Check-list of Italian birds-2020. Avocetta 2021, 45, 21–82. [Google Scholar]
- Kraus, D.; Bütler, R.; Krumm, F.; Lachat, T.; Larrieu, L.; Mergner, U.; Paillet, Y.; Rydkvist, T.; Schuck, A.; Winter, S. Catalogue of Tree Microhabitats—Reference Field List Integrate + Technical Paper; European Forest Insitute: Freiburg, Germany, 2006; 16p. [Google Scholar]
- Giannetti, F.; Chirici, G.; Gobakken, T.; Naesset, E.; Travaglini, D.; Puliti, S. A new approach with DTM-independent metrics for forest growing stock prediction using UAV photogrammetric data. Remote Sens. Environ. 2018, 213, 195–205. [Google Scholar] [CrossRef]
- Michałowska, M.; Rapiński, J. A Review of Tree Species Classification Based on Airborne LiDAR Data and Applied Classifiers. Remote Sens. 2021, 13, 353. [Google Scholar] [CrossRef]
- Gschwantner, T.; Alberdi, I.; Bauwens, S.; Bender, S.; Borota, D.; Bosela, M.; Bouriaud, O.; Breidenbach, J.; Donis, J.; Fischer, C.; et al. Growing stock monitoring by European National Forest Inventories: Historical origins, current methods and harmonisation. For. Ecol. Manag. 2022, 505, 119868. [Google Scholar] [CrossRef]
- Laes, D.; Reutebuch, S.E.; McGaughey, R.J.; Mitchell, B. Guidelines to Estimate Forest Inventory Parameters from LiDAR and Field Plot Data; Companion document to the Advanced Lidar Applications; U.S. Department of Agriculture Forest Service: Washington, DC, USA, 2011.
- Næsset, E. Practical large-scale forest stand inventory using a small-footprint airborne scanning laser. Scand. J. For. Res. 2004, 19, 164–179. [Google Scholar] [CrossRef]
- Vangi, E.; D’Amico, G.; Francini, S.; Borghi, C.; Giannetti, F.; Corona, P.; Marchetti, M.; Travaglini, D.; Pellis, G.; Vitullo, M.; et al. Large-scale high-resolution yearly modeling of forest growing stock volume and above-ground carbon pool. Environ. Model. Softw. 2023, 159, 105580. [Google Scholar] [CrossRef]
- Breidenbach, J.; Waser, L.T.; Debella-Gilo, M.; Schumacher, J.; Rahlf, J.; Hauglin, M.; Puliti, S.; Astrup, R. National mapping and estimation of forest area by dominant tree species using Sentinel-2 data. Can. J. For. Res. 2021, 51, 365–379. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Genuer, R.; Poggi, J.M.; Tuleau-Malot, C. VSURF: An R package for variable selection using random forests. R J. 2015, 7, 19–33. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org (accessed on 2 January 2024).
- Wright, M.N.; Ziegler, A. Ranger: A fast implementation of random forests for high dimensional data in C++ and R. J. Stat. Softw. 2017, 77, 1–17. [Google Scholar] [CrossRef]
- Chen, F.; Hou, Z.; Saarela, S.; McRoberts, R.E.; Ståhl, G.; Kangas, A.; Packalen, P.; Li, B.; Xu, Q. Leveraging remotely sensed non-wall-to-wall data for wall-to-wall upscaling in forest inventory. Int. J. Appl. Earth Obs. Geoinf. 2023, 119, 103314. [Google Scholar] [CrossRef]
- Ćosović, M.; Bugalho, M.N.; Thom, D.; Borges, J.G. Stand Structural Characteristics Are the Most Practical Biodiversity Indicators for Forest Management Planning in Europe. Forests 2020, 11, 343. [Google Scholar] [CrossRef]
- Dalponte, M.; Ene, L.T.; Gobakken, T.; Næsset, E.; Gianelle, D. Predicting selected forest stand characteristics with multispectral ALS data. Remote Sens. 2018, 10, 586. [Google Scholar] [CrossRef]
- Herniman, S.; Coops, N.C.; Martin, K.; Thomas, P.; Luther, J.E.; van Lier, O.R. Modelling avian habitat suitability in boreal forest using structural and spectral remote sensing data. Remote Sens. Appl. Soc. Environ. 2020, 19, 100344. [Google Scholar] [CrossRef]
- Rooney, R.C.; Azeria, E.T. The strength of cross-taxon congruence in species composition varies with the size of regional species pools and the intensity of human disturbance. J. Biogeogr. 2015, 42, 439–451. [Google Scholar] [CrossRef]
- Hammond, M.E.; Pokorný, R.; Okae-Anti, D.; Gyedu, A.; Obeng, I.O. The composition and diversity of natural regeneration of tree species in gaps under different intensities of forest disturbance. J. For. Res. 2021, 32, 1843–1853. [Google Scholar] [CrossRef]
- Grove, S.J. Saproxylic insect ecology and the sustainable management of forests. Annu. Rev. Ecol. Syst. 2002, 33, 1–23. [Google Scholar] [CrossRef]
- Martini, I.; Galipò, G.; Foderi, C.; Tocci, R.; Sargentini, C. Ornithical community of Vallombrosa Biogenetic National Nature Reserve (Italy). Eur. Zool. J. 2021, 88, 254–268. [Google Scholar] [CrossRef]
- Lange, M.; Türke, M.; Pašalić, E.; Boch, S.; Hessenmöller, D.; Müller, J.; Prati, D.; Socher, S.A.; Fischer, M.; Weisser, W.W.; et al. Effects of forest management on ground-dwelling beetles (Coleoptera; Carabidae, Staphylinidae) in Central Europe are mainly mediated by changes in forest structure. For. Ecol. Manag. 2014, 329, 166–176. [Google Scholar] [CrossRef]
- Sitzia, T.; Piazzi, C.; Barazzutti, G.; Campagnaro, T. Abandonment of timber harvesting favours European beech over silver fir: Evidence from Val Tovanella Nature Reserve in the southern Dolomites (Northern Italy). J. Prot. Mt. Areas Res. Manag. 2018, 10, 17–27. [Google Scholar] [CrossRef]
- Bütler, R.; Angelstam, P.; Ekelund, P.; Schlaeffer, R. Dead wood threshold values for the three-toed woodpecker presence in boreal and sub-Alpine forest. Biol. Conserv. 2004, 119, 305–318. [Google Scholar] [CrossRef]
- Fahrig, L. When does fragmentation of breeding habitat affect population survival? Ecol. Model. 1998, 105, 273–292. [Google Scholar] [CrossRef]
- García, N.; Numa, C.; Bartolozzi, L.; Brustel, H.; Buse, J.; Norbiato, M.; Recalde, J.I.; Zapata, J.; Dodelin, B.; Alcázar, E.; et al. The Conservation Status and Distribution of Mediterranean Saproxylic Beetles; IUCN: Gland, Switzerland, 2019. [Google Scholar] [CrossRef]
- Tellini Florenzano, G.; Cutini, S.; Campedelli, T.; Londi, G. Ecology and possible evolution of Crested Tit (Lophophanes cristatus) and Black Woodpecker (Dryocopus martius) populations in the Apennines, Italy. In Proceedings of the Bird Numbers 2010 “Monitoring, Indicators and Targets”. Book of Abstracts of the 18th Conference of the European Bird Census Council, Cáceres, Spain, 22–26 March 2010; Bermejo, A., Ed.; SEO/BirdLife: Madrid, Spain, 2010; p. 119. [Google Scholar]
- Batáry, P.; Fronczek, S.; Normann, C.; Scherber, C.; Tscharntke, T. How do edge effect and tree species diversity change bird diversity and avian nest survival in Germany’s largest deciduous forest? For. Ecol. Manag. 2014, 319, 44–50. [Google Scholar] [CrossRef]
- Wesolowski, T.; Fuller, R.J.; Flade, M. Temperate forests. A European perspective on variation and dynamics in bird assemblages. In Ecology and Conservation of Forest Birds; Mikusinski, G., Roberge, J.M., Fuller, R.J., Eds.; Cambridge University Press: Cambridge, UK, 2018. [Google Scholar]
- Wesolowski, T.; Martin, K. Tree holes and hole-nesting birds in European and North-American forests. In Ecology and Conservation of Forest Birds; Mikusinski, G., Roberge, J.M., Fuller, R.J., Eds.; Cambridge University Press: Cambridge, UK, 2018. [Google Scholar]
- Coops, N.C.; Tompalski, P.; Goodbody, T.R.; Queinnec, M.; Luther, J.E.; Bolton, D.K.; White, J.C.; Wulder, M.A.; van Lier, O.R.; Hermosilla, T. Modelling lidar-derived estimates of forest attributes over space and time: A review of approaches and future trends. Remote Sens. Environ. 2021, 260, 112477. [Google Scholar] [CrossRef]
- Sasaki, T.; Imanishi, J.; Fukui, W.; Morimoto, Y. Fine-scale characterization of bird habitat using airborne LiDAR in an urban park in Japan. Urban For. Urban Green. 2016, 17, 16–22. [Google Scholar] [CrossRef]
- Cerrejón, C.; Valeria, O.; Mansuy, N.; Barbé, M.; Fenton, N.J. Predictive mapping of bryophyte richness patterns in boreal forests using species distribution models and remote sensing data. Ecol. Indic. 2020, 119, 106826. [Google Scholar] [CrossRef]
- Dubayah, R.; Blair, J.B.; Goetz, S.; Fatoyinbo, L.; Hansen, M.; Healey, S.; Hofton, M.; Hurtt, G.; Kellner, J.; Luthcke, S. The Global Ecosystem Dynamics Investigation: High-resolution laser ranging of the Earth’s forests and topography. Sci. Remote Sens. 2020, 1, 100002. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Shannon Index | Richness | |||||
|---|---|---|---|---|---|---|
| N° of Selected Variables | R2 | RMSE% | N° of Selected Variables | R2 | RMSE% | |
| Beetles | 4 | 0.12 | 13.7 | 3 | 0.07 | 21.4 |
| Saproxylic beetles | 3 | 0.11 | 13.5 | 3 | 0.11 | 26.4 |
| Birds | 3 | 0.06 | 8.5 | 4 | 0.17 | 17.0 |
| TreMs | 4 | 0.27 | 14.9 | 3 | 0.11 | 26.2 |
| Saproxylic TreMs | 3 | 0.19 | 24.6 | 7 | 0.07 | 32.7 |
| Epixylic TreMs | 3 | 0.24 | 50.2 | 2 | 0.30 | 41.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parisi, F.; D’Amico, G.; Vangi, E.; Chirici, G.; Francini, S.; Cocozza, C.; Giannetti, F.; Londi, G.; Nocentini, S.; Borghi, C.; et al. Tree-Related Microhabitats and Multi-Taxon Biodiversity Quantification Exploiting ALS Data. Forests 2024, 15, 660. https://doi.org/10.3390/f15040660
Parisi F, D’Amico G, Vangi E, Chirici G, Francini S, Cocozza C, Giannetti F, Londi G, Nocentini S, Borghi C, et al. Tree-Related Microhabitats and Multi-Taxon Biodiversity Quantification Exploiting ALS Data. Forests. 2024; 15(4):660. https://doi.org/10.3390/f15040660
Chicago/Turabian StyleParisi, Francesco, Giovanni D’Amico, Elia Vangi, Gherardo Chirici, Saverio Francini, Claudia Cocozza, Francesca Giannetti, Guglielmo Londi, Susanna Nocentini, Costanza Borghi, and et al. 2024. "Tree-Related Microhabitats and Multi-Taxon Biodiversity Quantification Exploiting ALS Data" Forests 15, no. 4: 660. https://doi.org/10.3390/f15040660
APA StyleParisi, F., D’Amico, G., Vangi, E., Chirici, G., Francini, S., Cocozza, C., Giannetti, F., Londi, G., Nocentini, S., Borghi, C., & Travaglini, D. (2024). Tree-Related Microhabitats and Multi-Taxon Biodiversity Quantification Exploiting ALS Data. Forests, 15(4), 660. https://doi.org/10.3390/f15040660