1. Introduction
Since the development of reprogramming technologies, personalized pluripotent stem cell lines (iPSCs) can be generated from any type of somatic cell [
1]. Due to their pluripotency, iPSCs can differentiate into any possible body cell and be triggered to assemble into organ precursors by cell-intrinsic mimicking of developmental morphogenesis or through support by technical means such as bioprinting. The architecture of organoids as three-dimensional, miniaturized and simplified organs resembles their in vivo counterparts and recapitulates at least some functions of the organ [
2]. In the best case, organoids reflect the individual specifics of the donor’s tissue or organ-associated physiology or pathology. Although organoids can also be differentiated from other stem cell types or from tissue biopsies, we will focus here on iPSCs—derived organoids.
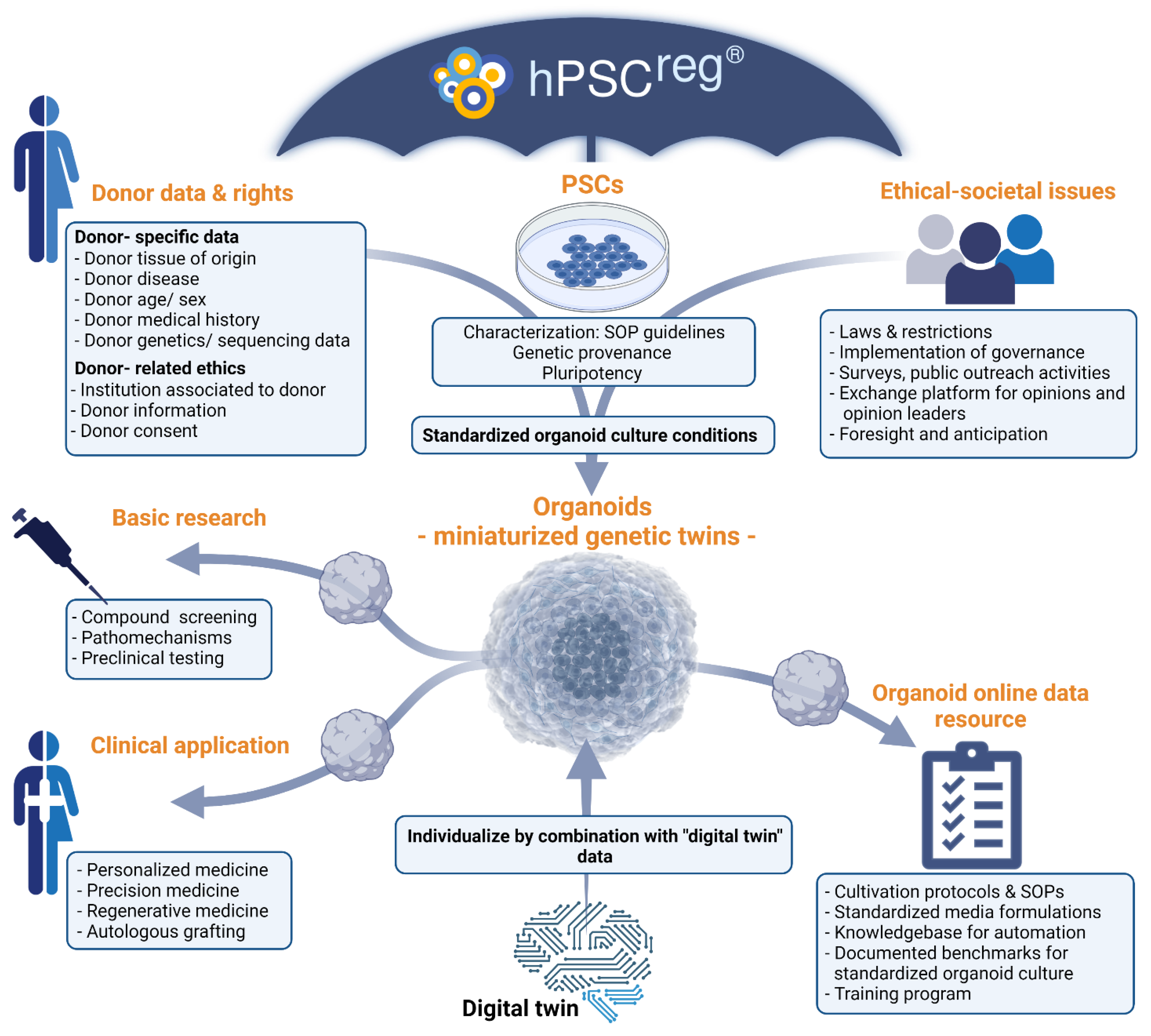
The iPSCs and corresponding organoids derived from them are in principle genetic copies of their donor and thus represent simplified miniature twins. The combination of multiple different organoids, for example, in technically more complex “body on the chip” systems, may iteratively complete this partial twin [
3]. The “twinning” of individuals in cell-based in vitro models is becoming increasingly attractive in the field of personalized medicine in order to individualize drug use, or in cell therapies, where organoids can be used for grafting [
4] (
Figure 1). This “Twinning” is not limited to the field of living in vitro cell cultures but can be extended to the genesis of “digital twins”, where the biological entity is captured digitally by data systems. This digital aspect is not covered here.
Here, we focus on the biological organoid entity and will address challenges related to the growing number of living in vitro organoid-based twins of individual persons and their usage to optimize personalized diagnosis and therapy. This usage requires a thorough understanding of the reproducibility of personalized organoid phenotypes over time and in different laboratories, which must be assessable within these temporal and special distances. Then, we will discuss the ethical aspects related to the inherent immortal property of iPSCs, and thus, their derivatives. We will identify necessary means to manage these challenges transparently using the human pluripotent stem cell registry (hPSCreg) as an exemplary platform. The hPSCreg is a well-established, publicly accessible database, which collects information on human pluripotent stem cells to make the different lines comparable and data findable, accessible, interoperable and reusable (FAIR) [
5,
6,
7]. In addition, hPSCreg is accepted in the field as a sustained knowledge resource, including scientific (biological) data on human PSC lines, as well as information on their ethical provenance, thus offering a complete assessment portfolio, which is utilized by funders for cell line certification. In addition, hPSCreg is providing persistent identifiers for hPSC—allowing for data interoperability. Furthermore, hPSCreg issues cell line certificates based on adherence to high scientific and ethical standards.
In contrast to the patient and donor themselves, their genetically mostly identical stem cells and derived organoid models are modifiable and renewable, making it possible to observe therapeutic or pathogenic effects. Moreover, as human cells they represent a much more comparable model object than animal models. The data obtained from experiments with these miniature twins of a donor can be linked together or even correlated with clinical, lifestyle, environmental and biographical data of the donor. This leads to a large number of diverse data, which can only be managed by technologies allowing data analysis of cells, organoids and whole organisms in the form of increasingly complex digital twins. Technologies including artificial intelligence, machine learning systems, systems biology, quantum computing and the suitable storage of large number of data are being developed for virtual simulation of physiological processes to predict the behaviour of biological system under variable conditions. Similar to the original concept of digital twins from engineering science for the development of prototypes that are optimized on the digital model before real construction, modern computer technology makes it possible for virtually model cells, cell processes, organ functions and interactions, thus creating a virtual physiological human (VPH) or digital twin of a human being or an organ or individual cells [
8,
9,
10,
11,
12].
2. Challenges of Organoid Applications
Organoids can be applied in two important areas: in clinical application, such as in personalized medicine, and in biomedical (basic) research (
Figure 1).
Organoids allow the personalization at the individual level, assuming that the donor’s biological features are actually twinned in the organoid. While the donor’s genetics and those of the organoid are in principle identical, other non-genetic aspects may not be, including those caused by lifestyle, age or systemic environments. Furthermore, even the genetic identity may gradually change upon in vitro cultivation, perhaps affecting the phenotypic relevance of the organoid model. This becomes particularly important when organoids aim to provide models for genetic diseases where environmental factors are known to induce disease progression following the two-hit/second-hit theory [
13]. Co-cultivation conditions and clonal variability in primary donated cells, and in derived iPSCs, may cause further variability in the characteristics of the organoids. Moreover, it is unclear to what extent epigenetic imprints are maintained in iPSCs and derived organoids, and whether these relate to the source cell tissue origin, or donor age [
14,
15,
16]. To what degree genetic and epigenetic variability in general is tolerable for personalized applications is currently unclear. One solution to answer this would be to generate generalized virtual organoid models, which fuse data from a number of standard organoids to achieve a virtual model. This standard organoid, or prototype, might allow us to measure deviations in individual organoids and assess their reproducibility. What such a standard looks like must be determined on target bases by the research community. Alternatively, individual organoid information can be individually assessed for any other organoid and treated individually. In both cases, it is essential to define the data and information required for organoid characterization and comparison (
Table 1).
More detailed and refined parameters for organoid characterization requires a detailed knowledge of the specificities of the different organoid types and would be impossible to list in this review. We propose that stakeholder groups develop these data requirements for thorough characterization, assessment and validation of organoids. These groups may be organized by dedicated organizations such as the Organoid Society (
https://organoids.org/main/main.php, accessed on 6 April 2022).
A basic requirement for such a comparability platform is the development and general agreement on standards for organoids. These standards may vary depending on application; however, basic or mandatory information should form the fundament of a given organoid (
Table 1) [
17,
18]. If such data are deposited in a transparent database, it can be utilized to improve reproducibility of organoids, compare protocols and provide a valuable research resource for organoid-based models. Reliability of organoid usage as a biological twin will be more robust and the risk for errors reduced. The hPSCreg (
https://hpscreg.eu, accessed on 6 April 2022) may act as an established prototype for translation into the organoid field. It registers information on human pluripotent stem cell (hPSC) lines, including data on phenotype, derivation, genetic composition and ethical provenance, which are annotated by publications and project related information. Thereby, registered hPSC lines have to meet agreed, general quality standards of pluripotent cells. The hPSCreg database could, for example, be linking each registered hPSC line to organoids derived from it, thus making this information usable to study penetrance of individual genetic and phenotypic traits along developmental pathways. Furthermore, the generation and application of organoids can be traced, made FAIR and ethical–societal issues adequately addressed by providing ethical provenance information. This includes iPSC-derived organoids representing early embryonic development such as blastoids or gastruloids [
19,
20,
21,
22]. Linkage to the original donors via unique pseudonymized identifiers may further increase utility of such an ecosystem of biological twins comprising individual donors, PSCs and organoid subjects.
3. Organoid Application
The use of personalized organoid models could make it possible to test and optimize drugs, treatments and therapy concepts before their application in patients. This development accelerates towards personalized/precision medicine. Moreover, besides organoid application in pre-clinical testing, direct organoid grafting in patients is an attractive prospect for tissue repair (4; reviewed in [
23]). The clinical data from such trials and personalized drug therapy applications may further feedback into and enrich the individual twin model. A particular advantage of stem cells and their derivatives, such as organoids, is their scaling in endless numbers, allowing for experimental repetitions until robust findings emerge for understanding biological processes. The need for massive numbers of homogenous organoids is anticipated as organoids of various forms are increasingly used in research to understand metabolic and physiological processes as well as disease mechanisms. For example, kidney organoids are used to test drug efficacy in vitro [
24], intestinal organoids are being developed and human inner ear organoid structures have already advanced diagnosis and biomedical knowledge [
25,
26]. Influences of genetic modifications on the growth and development of various organ structures can be studied and tested on these models in a way that would not be possible in humans. This includes basic research on lineage-tracing experiments, e.g., on neural fate determination during development with unprecedented spatiotemporal resolution [
27] and high-throughput microscopical screenings that allow for rapid identification of chemically active compounds [
28]. Unfortunately, these models often encounter weaknesses in uniformity partially due to technical and procedural heterogeneity of the different laboratories, which makes comparability and transparency difficult. In addition, the complex biological environment of organs, as found in the human body (for example, the influence of neighbouring organs, blood vessels and biomechanics such as blood flow, compression, movement and tension as well as functional innervation and vascularization) has a significant influence on the development and functionality of the corresponding organ. Standards on these parameters must at least be transparent and disseminated through training and collaboration. Current efforts and technical advances in automation and controlled manufacturing as well as cryopreservation may target these issues in the near future. These technical advances include perfused culture systems and high-throughput and reproducible bioprinting of organoids [
29,
30].
3.1. Future Organoid Use and Ethical Foresight Requirements
Stem cell application beyond already anticipated future perspectives requires careful foresight to inform donors about possible routes and developments ahead, including explicit information on biological and digital twin aspects. Science is constantly progressing, and today it is difficult to foresee in which specific areas stem cell lines and their derivatives, such as organoids, could one day be used. New areas of application are already emerging today that were unimaginable only a short time ago. A prominent example is the scalable genesis of personalized organoids themselves, and thus, the production of ever-more complex bodies in the laboratory outside the source organism. The formation of animal–human chimeras represents another development. Here, hPSCs are used to generate human organs in animal hosts, for example, to harvest these organs for transplantation [
31]. These chimeric animal models created in a Petri dish could facilitate the study of human development and disease. While the moral status of these “hybrid beings” remains controversial [
32], the relationship between the chimeric being and the human donor would also need attention and could be established by informed consent. Revolutionary developments are taking place in the field of human reproductive medicine. It may soon be possible to induce human germ cells from individual iPSCs. These spermatids and oocytes could then be used for artificial insemination or in vitro fertilization, even in people who can produce only a few or no germ cells, as well as in same-sex partnerships. As molecular mechanisms in humans concerning germ cell development are still poorly understood, the utilization of oogonial or testicular organoids to promote gametogenesis is an attractive possibility [
33,
34]. Similar scenarios are possible for blastoids or gastruloids derived from iPSCs, which are currently used for basic research in early human development, modelling of congenital disorders and testing of drug effects on human development. While it is possible that these organoids may also be used for reproductive purposes, their research use will certainly challenge the currently adapted 14-day rule and trigger new ethical discussions on the definition of developmental thresholds, which would allow for experimentation.
The use of iPSC and derived organoids also raises questions about the commercial profit that companies can or should make from the use of donated material [
35], as well as more anthropological questions about the alienation of the donated material through modifications and combinations with other cells or circumstances from the original donor and its physical and genetic identity. How long can donated living cells be considered part of the donor? The more research progresses, the more societally and ethically challenging situations could arise here [
36]. A look at the dynamics of scientific development shows that the knowledge and research focus at the time of cell donation does not sufficiently correspond to future points in time, nor even the currently often diverse national or regional ethical and legal frameworks. It is thus of relevance when working with organoids as biological twins of their donor that these donors have the option to be informed about and to consent to the application. Similar to current efforts of personal data usage, mechanisms allowing this donor involvement are required. The informed consent process plays a key role in this empowerment.
3.2. Informed Consent Process
An important catalyst for the ethical debate on the consent process in cell or tissue donation was certainly the case of the cancer patient Henrietta Lacks [
37]. In the 1950s, living cancer cells were taken from her in the USA without her knowledge and without her consenting to the use, storage and cultivation of the cells for decades. The genome of these cells was sequenced in 2013, and these data were made public. Only after disputes with Lack’s descendants were these data placed under controlled access. This example illustrates the tremendous importance of the informed consent process, as well as, in view of the immortality of iPSCs, the challenges that will arise for their future use. It is already standard that during the donor consent process, the extent and nature of the use, storage and distribution of the donated material and associated personal data must be explained. Furthermore, the option of non-commercial use or commercial use, transforming the donated material into a commodity, may require consent [
38]. These categories can be treated in different ways in the consent process: either in relation to specific concrete projects or as a “blank authorisation” in the form of a generic or broad consent that allows the use of the cells in (all) possible future areas of application without restrictions [
39]. The weakness of the broad consent process, often applied for biobanks, rejects the notion of donor empowerment with regard to the fate of his cells, especially for future applications hidden in the darkness of uncertainty. Several ethical values must be balanced; the primacy of personal data protection in the sense of preserving the donor’s anonymity must be synchronized with continuous donor contact to enable desired empowerment. However, it is not only donor-related values that need to be upheld, but also socially recognized values, such as scientific freedom and the desire for medical progress. The translation of these conditions requires technical and practical means, which are yet to be developed.
One possible way forward may be the so-called “dynamic consent” where the donor is asked for consent for new projects via a digital tool that protects personal identity. A personalized digital interface would thus be needed to facilitate two-way communication between researchers and donors in order to strengthen the donor’s empowerment and allow for consent adaptation at any time and also to help in deciding on new fields of application [
40]. However, this raises the question of what happens, for example, after the donor’s death: should the cells then be destroyed or released for all research topics, or should it even be possible to transfer decision-making power to authorized third parties, family members or relatives? This raises the question of what status donated cells should have, which is primarily a social, values-related issue. Whether the donation of cells is a genuinely altruistic act, i.e., the handing over of cells from one’s own body for disposal by the general public, or a gift to the general public, or whether they are a product that other people are allowed to modify, sell and distribute needs to be decided and is partially framed by legal documents. Whether the donor has a right to control the donated cells depends mainly on what entities society, or the donor, understands these cells to be: are they still parts of the donors, subject to their dignity and control, or have they become common property?
A first step to maintain a connection between donor and organoid application could be made by providing information about consent content to all users of the iPSC-derived cells. This is implemented in hPSCreg, where informed consent data are collected. These are used to decide whether cells have the relevant ethical provenance to allow certain research also covering the handling of personal data. The implementation of data resources regarding iPSC-derived organoids in hPSCreg and their application would provide for the second part of information needed to manage the duality between user and donor. Only the tools needed for anonymous communication between both parties are missing.
4. Conclusions
An important challenge of using human cells for the production of PSCs and derived organoids, which in principle can be cultivated indefinitely and at any location, is respecting and maintaining consent. To ensure this, central institutions are needed to maintain and manage the ethical provenance of the organoids used. To some extent, this role is already fulfilled by the European database hPSCreg with regard to PSC lines, but this must be adapted for organoids.
The hPSCreg is a well-established, public database where hPSC lines, either embryonic or iPSC lines, and hPSC-associated projects can be registered [Seltmann et al., 2016]. It acts as a central hub for freely accessible information on existing hPSC lines, their quality, characteristics and their derivation to provide a global overview and to monitor the development of the field. hPSCreg contributes to avoiding redundancy in the generation of hPSC lines—so reducing the need to create ever-new lines in using more human tissues and embryos, more than necessary. hPSCreg monitors and analyses the ethical, legal and societal frameworks for stem cell research and application, which have changed a great deal in recent years and decades and can also differ greatly from nation to nation. Using hPSCreg as a basic platform for human PSC-derived organoids could have the advantage of linking the donor data, hPSC data and the derived organoid information together. However, this poses key challenges in terms of monitoring, content and technical implementation. Identification, assessment and follow up of existing organoids per cell line in multiple laboratories appears impossible via a centralized database. One solution may be the provision of basic organoid information via hPSCreg. This basic information would be associated with links to the originators of the organoids to allow the access of more and updated data. This approach may, in addition, be extended to provide open access capabilities for data of organoid researchers. Similarly, organoid protocols, media and assay methods could be either linked to hPSCreg by individual researchers or hPSCreg could provide an accessible database of protocols associated with the hPSC lines and their derived organoid entities. Facilitation of reproducibility by these measures will be complemented by the hPSCreg platform through providing links to publication and other public information on the respective organoids and protocols.
In addition, all consent templates belonging to a stem cell line are kept in the database and are thus available—these consents naturally (containing specific restrictions on use or presenting a broad consent) refer to organoids derived from these PSCs, ideally in unambiguously linked formats. Databases such as hPSCreg are guardians of the respect and preservation of such important documents concerning the donor’s autonomy and will, while preserving his identity through anonymization or pseudonymization processes. If the trend were to move towards “dynamic consent” because of the desire for more donor empowerment, databases such as hPSCreg could play an important management role in improving communication between donors and researchers, whereby the database can, at the same time, be a protective shield to safeguard the identity of the donor.
The disadvantage of globally non-uniform standards in the laboratories for the production of organoids can also be eliminated by such central databases—here, the different standards can be collected and thus made comparable and able to be aligned.
In all three areas of the social–ethical challenge, the need for social discourse is implicit. A database such as hPSCreg could be expanded to a hub to initiate and coordinate this discussion among all concerned stakeholders—patients, donors, researchers, doctors and citizens. The development of such biomedical databases, which also have a view on the ethical provenance of cells, into a hub for the public discussion of the above-mentioned areas of value-related tension would increase the awareness of the population regarding these problems and promote solutions.






{kind=link}