Abstract
Environmentally modified housing [EMH; windowless, insulated sidewalls and ceiling, thermostatically controlled ventilation fans) versus conventional housing [CVH; cross-ventilated, insulated ceiling, ceiling fans) improved broiler performance in the summer. The objective of this investigation was to determine whether social behaviors differed between two population densities (0.06 m2/chick [HD] or 0.07 m2/chick [LD]) in these houses. We used a randomized block statistical design, involving houses, population densities, observation times, and bird age. Behaviors were observed weekly, during the morning and the afternoon. Individual observers focused on the group of broilers in one of three defined 26.76 m2 areas in each of the four pens in each house. Aggressive encounters, tail and back pecking, feather eating, thermoregulatory, preening, and flock mobility were recorded. Feather pecking, eating and aggressive encounters were expressed at greater rates in HD birds in CVH. A salt-deficient diet caused increased feather pecking and aggressive encounters, which decreased after correction of the mistake. Increased heat indices (HIs), HD, and greater light intensity in CVH influenced behaviors and mortality more severely than in EMH. In CVH and EMH, burrowing/thermoregulatory/resting activity increased with increasing HIs. Afternoon preening was elevated significantly in EMH. It was concluded that broilers reared in EMH were more comfortable and experienced improved welfare compared to those reared in CVH.
1. Introduction
In the late 1960s to the mid-1970s, increased attention to alternative housing for poultry production was just becoming an important issue [1]. Earlier observations suggested that windowless housing (EMH) was superior to open conventional curtain sidewall (CVH) poultry houses, primarily in association with reduced production costs, which were lower in windowless housing [1]. Nevertheless, there was no rush to convert to the EMH system because capital investment was increased significantly over CVH.
However, current poultry housing is very different from the CVH in the 1960s and 1970s, and most poultry are now reared in modern EMH [2]. This shift has been due to rising energy costs, increasing feed costs, and lack of performance in CVH with the current strains of broiler chickens [3].
The behavior of chickens has been observed for thousands of years, albeit not so much as a tool for the improvement of poultry performance but for entertainment and amusement. Today, through behavioral studies with poultry, it has been possible to define environmental conditions which are more appropriate for the production of broiler chickens and other varieties of poultry. Recent behavioral studies involving increased environmental complexity and high and low housing densities have revealed that complex housing resulted in increased general activity of broiler chickens [4]. High environmental complexity consisted of a variety of objects such as a PECKstoneTM (a unique enrichment tool designed specifically to prevent and reduce abnormal pecking behavior), a wooden dust bath, perches, hanging bundles of string, metal balls filled with alfalfa hay, dispenser balls filled with whole grain oats, colored balls, Kong balls filled with lettuce, and laser lights engaged twice daily. This positive effect of environmental enrichment was most noticeable in low-density stocking pens, but even in high-complexity, high-density stocking pens there was more frequent activity, but with less foraging, drinking, and eating. During the 7 wk observation period, there were notable changes in activities from 2 wk to 4 wk to 7 wk, with the greatest frequency of activities at 4 wk. Another study [5] evaluated four different stocking densities to evaluate welfare, growth, and blood chemical profiles in broiler chickens. In this study, the investigators observed that each increase in stocking density caused a significant decrease in body weight gain and significant increases in feed conversion ratios, which could have been implied in the high-density broilers involved in the research cited above [4]. Blood profiles among the four stocking densities did not differ, with the exception of reduced serum total proteins and increased total leukocyte counts, both of which are indicators of stress reactions associated with increasing stocking densities. Stocking density appears to be one of the more important focuses of studies on the behaviors of broiler chickens. In another recent publication [6], 18 diverse behaviors were exhibited by broilers at 20 and 31 d of age. Their findings revealed that drinking, resting, head scratching, and undefined behaviors were reduced significantly in broilers in the highest stocking density at 21 d of age, but those particular behaviors did not differ among stocking densities when the chicks aged to 31 d. However, increasing stocking densities negatively impacted performance (body weight, average weight gain, feed intake, and gain:feed) [6], but gait and footpad scores were not impacted by increasing stocking densities [6].
Even though stocking density has recently received much emphasis as a factor attributed to numerous behavioral expressions by broiler chickens, other environmental factors such as light intensity and ambient temperature have not been investigated exhaustively. In studies comparing high (20 lux) and low (5 lux) light intensities on broiler behaviors [7], it was noted that body weight of broilers in high-light-intensity conditions were smaller than the body weights of broilers grown in low-light-intensity conditions, which would be in agreement with the assessment that increased activity in high stocking densities was associated with smaller body weights [4]. It was also noted that high intensity light induced more activity among broilers than in broilers reared in low-light-intensity conditions [7], even though activity in general increased with age in both light intensities [7]. In a recent review [8], light intensity and duration were major components of poultry management practices because light and various wavelengths are very powerful zeitgebers that generally control the expression of all circadian patterns and wellbeing in birds. It was pointed out that visual stimuli drive many normal and even aberrant behaviors in chickens, and broiler social behaviors are definitely regulated by light control [8]. Although light intensity and duration are controlled in most studies dealing with broiler behavior, it appears that these variables are generally in conformation of basic poultry management practices [4,8]. Blue and green light wavelengths enhance the performance of broilers, but the red light wavelength has the potential to increase general activity and especially aggressive behavior, causing decreased performance [8].
Maintenance of environmental quality as a means to mitigate thermal stressors also impacts the behaviors of broiler chickens [9]. Heat-related neural/physiological mediators of behaviors related to body temperature regulation, such as decreased feed intake, increased water intake, panting/gular flutter and enhancing surface areas to increase thermal loss to the atmosphere or to ground temperature, are frequently noted in all broiler flocks exposed to the heat of summer [9]. These aforementioned behaviors along with other social behaviors such as fearfulness can be associated with neurochemical changes in the central nervous system [10].
It has been noted that feed is one of the most important and costly production components [11]. Many studies have assessed feed ingredients and texture relative to feeding behavior [12,13,14]. From these and similar studies, it was learned that broilers and other poultry spend more time at feeders with mash versus pellets and crumbles.
Movement within poultry houses has been investigated showing that space-use patterns are influenced by placement of feeders and waterers [15]. Scientists have studied energetics and behavior in commercial broilers and observed that posture and standing had a significant effect on metabolic rate due to the fact that standing was more energetically expensive than sitting [16]. Thus, it was possible to discern that locomotion would be decreased among current fast-growing, high-yielding broilers than in older commercial lines, especially as broilers age and add increased mass to breast muscles.
Cannibalism, collectively associated with high-frequency feather pecking and the consumption of torn flesh from pen mates, has been observed frequently in broiler flocks. Cannibalism, according to several poultry scientists [17], is a learned behavior that has numerous causes, such as protein, salt, and phosphorus deficiencies, a response to crowding, high-intensity lighting, inadequate access to feeders and waterers, and high temperature exposures, and can be associated with many inadvertent aspects of broiler production management on a global basis.
The current report addresses some social and developmental behaviors of broilers in two different housing systems over an 8 wk growing period. Behaviors such as aggressive encounters, tail and back feather pecking, feather eating, thermoregulatory behavior associated with burrowing into litter to reach the soil surface, preening, and frequency of whole-flock mobility following burrowing into the litter are described. The objective was to illustrate the differences in the expression of the above-named behaviors in broilers grown in CVH versus EMH during a hot–humid summer growing season.
2. Materials and Methods
2.1. Ethical Statement
This study was conducted at North Carolina State University at the North Carolina Agricultural Research Service Chicken Field Educational Unit. The study was approved and monitored by the University Institutional Animal Care and Use Committee (protocol #19-029A).
2.2. Housing
Two scale models of commercial broiler houses (11.0 × 29.25 m [321.75 m2]) on the North Carolina Agricultural Research Service Chicken Field Educational Unit were used from 1 June to 31 July. The control house was a conventional [CVH] (natural cross-ventilated, insulated ceiling, and ceiling fans) with no insulation in the sidewalls. The EMH was windowless with controlled lighting, insulated sidewalls and ceiling, and a thermostatically controlled ventilation fan system. The EMH was enclosed with corrugated aluminum sheet metal siding covering the exterior of the Styrofoam-insulated walls with a minimal R factor of 8. The CVH and EMH had ceiling insulation provided by 7.62 cm thick Styrofoam board insulation with an R Factor of 15. The CVH had ceiling-mounted air-circulating fans to stir the air, but the EMH had two banks of thermostatically controlled fans of fixed and variable speed installed to aid in the maintenance of air flow assisting in control of interior air temperature and flow. The EMH had the fans enclosed with light-dampening baffles to prevent light leakage into the house. Further, the EMH had air inlet baffles along the top of the long walls. In both houses, identical ceiling-mounted incandescent lighting equipment was installed, which was controlled by dimmers in conjunction with timers to maintain low light intensity in the EMH and during the night in the CVH.
2.3. Chick Management
Both houses had dirt floors, which were covered with 12.7 cm of fresh pinewood shavings over used litter before placement of chicks. Before placement of chicks, the houses were divided into 4 large pens (11 × 7.31 m each [80.4 m2]). Each pen had two pancake-type 31,000 BTU LP gas brooders (Hog Slat, Inc., Newton Grove, NC 28366, USA) hung from the ceilings. Chicks were placed into 2 large brooder rings in each pen, and infrared brooder lamps (Winco ESH-72, Hog Slat, Inc., Newton Grove, NC 28366, USA) were used to provide supplemental heat for newly placed chicks in the rings to assure that floor temperature was maintained at 33 °C and air temperature was maintained at 33 °C. Air temperature was reduced gradually to 31.5 °C at the end of week (wk) 1, 28 °C at the end of wk 2, 24 °C at the end of wk 3, and was allowed to maintain ambient temperature thereafter. However, as summer temperatures in mid-June rose, air temperatures and percentage-relative humidity (RH%) in CVH and EMH were allowed to fluctuate daily. It was an experimental goal to maintain the internal air temperature at 24 °C in the EMH, but a small thermal air effect was noted as external temperatures rose. Internal and external air temperatures were determined with the use of a sling psychrometer (Thermo Fisher Scientific, Raleigh, NC 27604, USA) during morning and afternoon observation periods. Based on the difference between dry-bulb and wet-bulb temperatures, RH% was determined using a psychrometric chart. Heat index (HI) was determined using the house internal and external ambient temperatures along with house internal and external RH% values, and then using the U.S. National Oceanic and Atmospheric Administration Heat Index Chart [18] (Table 1). The lighting program for each house provided continuous light for the first 10 days (d) at 20 lux at floor level, but from 10 d to 24 d of age, house lighting was reduced to 5 lux at floor level and maintained at that intensity until the end of the experiment at 8 wk of age. The lighting regimen provided 1 h darkness and 23 h light from placement until 10 d of age. At 10 d of age, in the CVH, the blackout curtain sidewalls were lowered partially to expose the broilers to natural light, which was increasing in photoperiod as the season progressed from late spring to early summer conditions, and nighttime lighting was maintained at 5 lux at floor level to supply 23 h of light and 1 h of darkness. In EMH, light was maintained at 5 lux at floor level from 10 d of age until the end of the experimental period, with 1 h of darkness each day.

Table 1.
Summary of the analyses of variance (main effects and their interactions showing probability > F) used to analyze the number of events associated with the development of social behaviors of broilers in conventional and environmentally modified poultry houses.
2.4. Animal Placement
Broiler breeder eggs (Ross 708) were purchased from a commercial hatchery (Aviagen breeders) and incubated in setters at the North Carolina Agriculture Research Service Poultry Field Laboratory Hatchery. Within 3 h after completion of the hatching process, the chicks were subjected to feather sexing to separate males (short primary wing feathers) from females (long primary wing feathers). Male chicks were placed in pens at densities of either 0.06 m2/chick (1341 chicks/pen; ~16.7 chicks/m2; high density [HD]) or 0.07 m2/chick (1150 chicks/pen; ~14.3 chicks/m2; low density [LD]). There were two large pens of each density per house. Stocking density in this study was less than the maximum recommendation of the National Chicken Council in the United States, which has suggested a range of 32 to 44 kg (18 to 22 chickens) per square meter based on bird weight. Thus, the lower stocking densities involved in this study were selected due to expected body weights at 8 wk of age that would equal or exceed the maximum body weights/m2 stated above.
2.5. Feed
Feeds (starter, grower, and finisher) were manufactured at North Carolina Agriculture Research Service feed mill and met all NRC requirements for broilers [19]. Starter feed (23% crude protein; 12.77 mj/kg [3050 Kcal/kg]) was provided to wk 4; grower feed (21% crude protein; 13.31 mj/kg [3150 Kcal/kg]) was provided to wk 7; finisher/withdrawal feed (19.0% crude protein; 13.40 mj/kg [3200 kcal/kg]) was provided through wk 8. At 5 wk of age, the grower feed was remixed because the original grower mix had been compounded without the inclusion of sodium chloride.
Feed was provided ad libitum via hanging tube-type galvanized metal poultry feeders (Tractor Supply Co., Garner, NC 27529, USA), to which feed was added manually and shaken each day to assure even feed flow to the tray on each feeder. City water was provided ad libitum via Plasson type waterers (Homeplace Market, LLC, Schellsburg, PA 15559, USA). Routine cleaning of waterers and feeders was performed daily.
Anticoccidial (Maxiban [Elanco Animal Health Incorporated, Greenfield, IN 46140, USA] at 60 g/ton fed from d 1 through d 56) medicated starter feed was available from hatching to 4 wk of age, followed by the medicated grower feed from 4 to 7 wk of age, followed by the medicated finisher feed through 8 wk of age. Chicks were vaccinated against Marek’s Disease at the hatchery, and at 4 d of age the chicks were administered vaccines against Newcastle Disease and Infectious Bronchitis via the drinking water.
2.6. Behavioral Observations
Social behaviors were observed by 4 teams of 3 people per team and commenced when chicks were 3 wk old. Before initiation of the observations to be made in the two experimental housing types, the observers were given three daily training opportunities to assist them in making multiple observations of social behaviors of broilers in another conventional house. Training observation times were 30 min during the morning and afternoon sessions. Calibration of the performance of observers was not subject to statistical validation, but each set of training data was compared to that of the PI. Experimental observations were made over 30 min periods for each pen in both CVH and EMH.
Observations were made weekly and recorded during the morning (0800 to 0900 h) and again on the same day in the afternoon (1500 to 1600 h). Teams of observers entered a pen and took positions such that observations could be made in 2 adjacent large pens without gross movements being made by the observers. Each observer took a position from which one-third (26.8 m2) of a larger pen (80.4 m2) could be observed for a 30 min period, and the team would turn and continue to observe the adjacent one-third of the second pen for a 30 min period. This procedure allowed each observer of a team to visualize an arbitrary floor space of 26.8 m2 in each of two adjacent large pens. This method for making observations is inherently a subjective method limited by the ability of the independent observers to make rapid observations and record such behaviors on a mechanical device. However, when visually making observations in real time, there were some missed events, but out of such a seemingly chaotic effort, the occasional loss of an event was not detrimental to the overall quantitative results. Since the observers were observing behaviors from an elevated position, the possibility of lost events was minimized in the event of visual obstruction via a waterer or feeder or due to body overlap.
Before the weekly observations were initiated, 12 birds (four chicks randomly chosen from the center line spanning the arbitrary midline length of each of the three, one third arbitrary divisions of the pens) were marked with either green, blue, or black spray paint on the neck and back region to ascertain whether the male chicks were migrating away from a “home area” within the greater pens. Green-marked chicks were in the arbitrary left-side area of each large pen, the blue-marked chicks were in the center of the arbitrary center area of each large pen, and the black-marked chicks were in the arbitrary right-side area of each large pen. Birds were re-marked as they aged and feather covering improved. Observations did not begin until a minimum of 15 min had elapsed after the observers had entered the pens, which allowed time for the broilers to become accustomed to the presence of the observers and for the resumption of “normal” behaviors. During this period, which ostensibly allowed the broilers to return to base-line behaviors, the observers noted the relative positions of marked chicks in each assigned pen area. Social behaviors were recorded for each pen area using a mechanical counter with 8 channels, with a range of 0–999 events per channel.
The tallied social events per bird density per observation time included aggressive encounters (threats, sparing, fighting, and aggressive pecking to the head or body), back pecking with feather extraction, tail pecking with feather extraction, feather eating (including eating feathers from the floor), preening, temperature regulation (litter burrowing to reach the soil under the litter; not a dust bathing event), and intervals between spontaneous mobility of the whole flock in each pen, followed by a return to litter burrowing. Some litter burrowing was observed at 3 wk of age, and it was noted that between the two houses, there appeared to be a common time for the chicks to remain in a single burrowed location, after which the flock appeared to arise, nearly in unison, move about the home area drinking and eating, and then return to burrow into the litter. The mobility time varied between 5 and 11 min before returning to the litter. To quantify the frequency of these spontaneous mobility events, on a weekly basis, a single observer in each house would record the time intervals of these events over a 2 h observation period in the morning and afternoon of the day before the observations for the social behaviors; this occurred from when the chicks were 3 to 8 wk old.
2.7. Statistical Analysis
The number of behavioral events observed weekly by each observer in each pen were pooled within the morning and afternoon times for the population densities within each housing type for analysis. The experimental design was a randomized block design with two house types, two population densities, and two observation times (morning and afternoon) per each of the six weekly observation periods. Data were assessed within the house and among pens within each observation period in each house. The data were analyzed using the General Linear Models procedure of the Statistical Analysis System (SAS Institute, Inc., Cary, NC 27513, USA) [20]. The main effects and their interactions were considered as being significant if p values were less than or equal to 0.050 (Table 1).
3. Results
The environmental conditions were ambient. From 1 to 8 wk of age, weekly measures of air temperature, relative humidity (RH%), and determination of heat index (HI) as potential measures of comfort were made during the morning and afternoon before the scheduled days for observations (Table 2). There were differences between the two housing environments regarding internal ambient temperature and HIs from the third to the eighth week of the growth period.
Table 2.
Internal and external ambient temperature, relative humidity (% via sling psychrometer), and calculated heat index (HI °C) for conventional (CVH) and environmentally modified housing (EMH) affecting broilers during the experimental period from one to eight weeks of age.
3.1. Observations of Behaviors
Behavioral events over periods of 30 min per pen of broiler chickens from 3 to 8 weeks of age are reported in Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6 and Figure 7. Each recording session consisted of 30 min observation periods in both the morning and the afternoon. The recorded behaviors were associated with broilers in HD and LD pens in the two housing types (CVH vs. EMH). At 3 wk of age, the chicks, which were marked, remained near or within their “home area”, occasionally moving out of that home range. As they aged through the next 2 wk, there were some chicks that moved across the boundaries of their home range for short periods of time, rarely venturing more than one to two meters into those adjoining areas of the greater pen, before they might be approached aggressively, then moving back toward their home areas. However, those that moved the greatest distances from their home range within the large pen also were found to be involved in increased aggressive encounters, especially feather pecking. A few marked birds in each large pen actually wandered the full length and width of the greater pen area. In the CVH, there were commonly one to two wanderers in each pen, and in one large pen in the CVH, there were three wanderers. The EMH had fewer wanderer/roving birds, and there were only five in the whole house, with three in one pen and one each in two pens. Those wandering/roving broilers also tended to make linear-type movements, involving stopping, pecking feathers from one to three chicks involved in litter burrowing, then moving randomly to another area roughly one meter away to commence the feather pecking again. However, the feather pecking was not limited to the wandering/roving chicks, since within the home areas, some assumed that residents within those areas were feather peckers also.
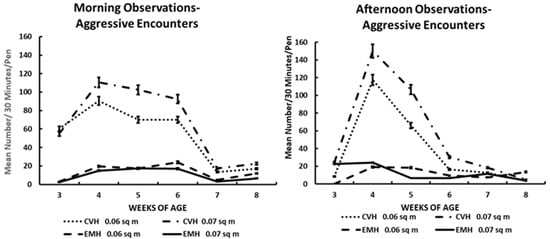
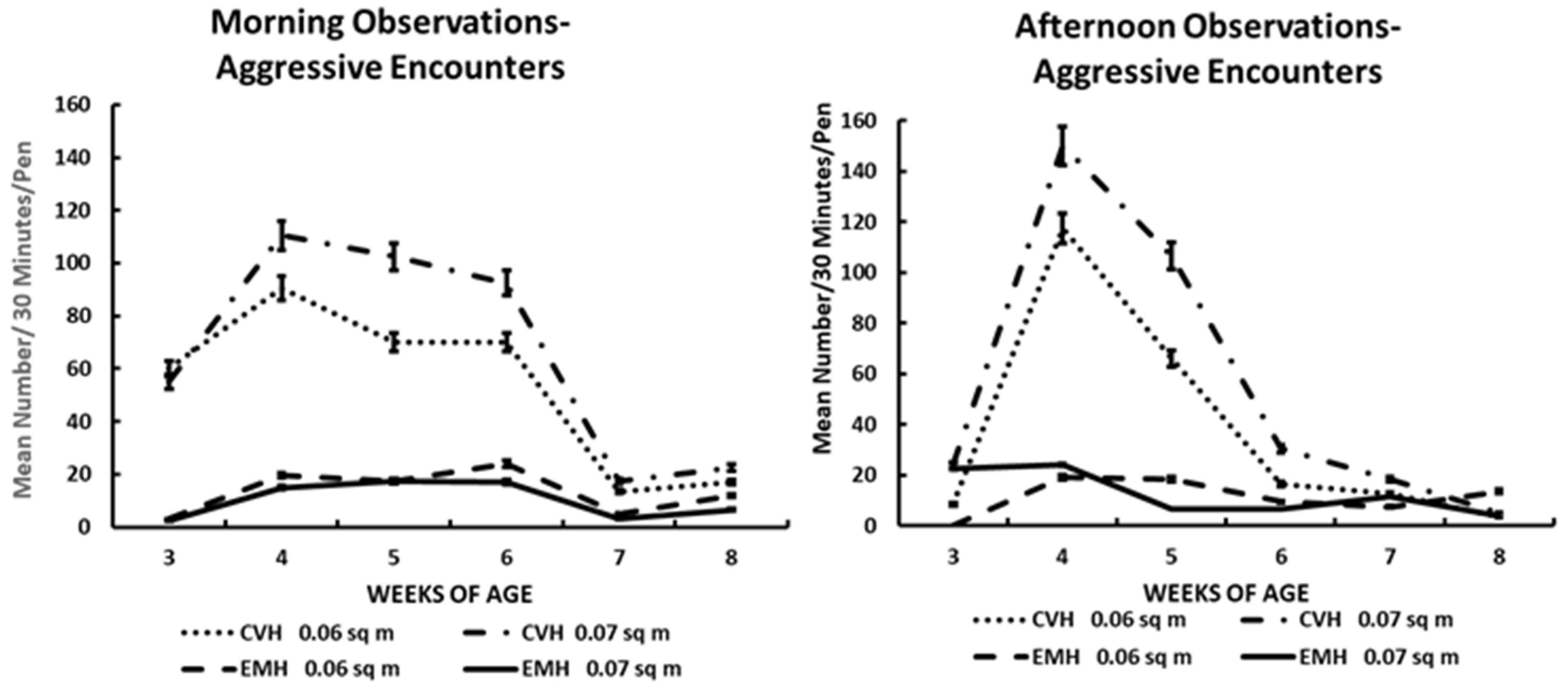
Figure 1.
Aggressive encounters in the morning and afternoon in broiler chickens observed from three to eight weeks of age in conventional (CVH) and environmentally modified housing (EMH). Vertical bars on each data point represent the standard error of the mean.
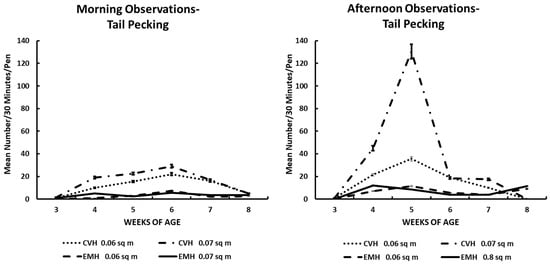
Figure 2.
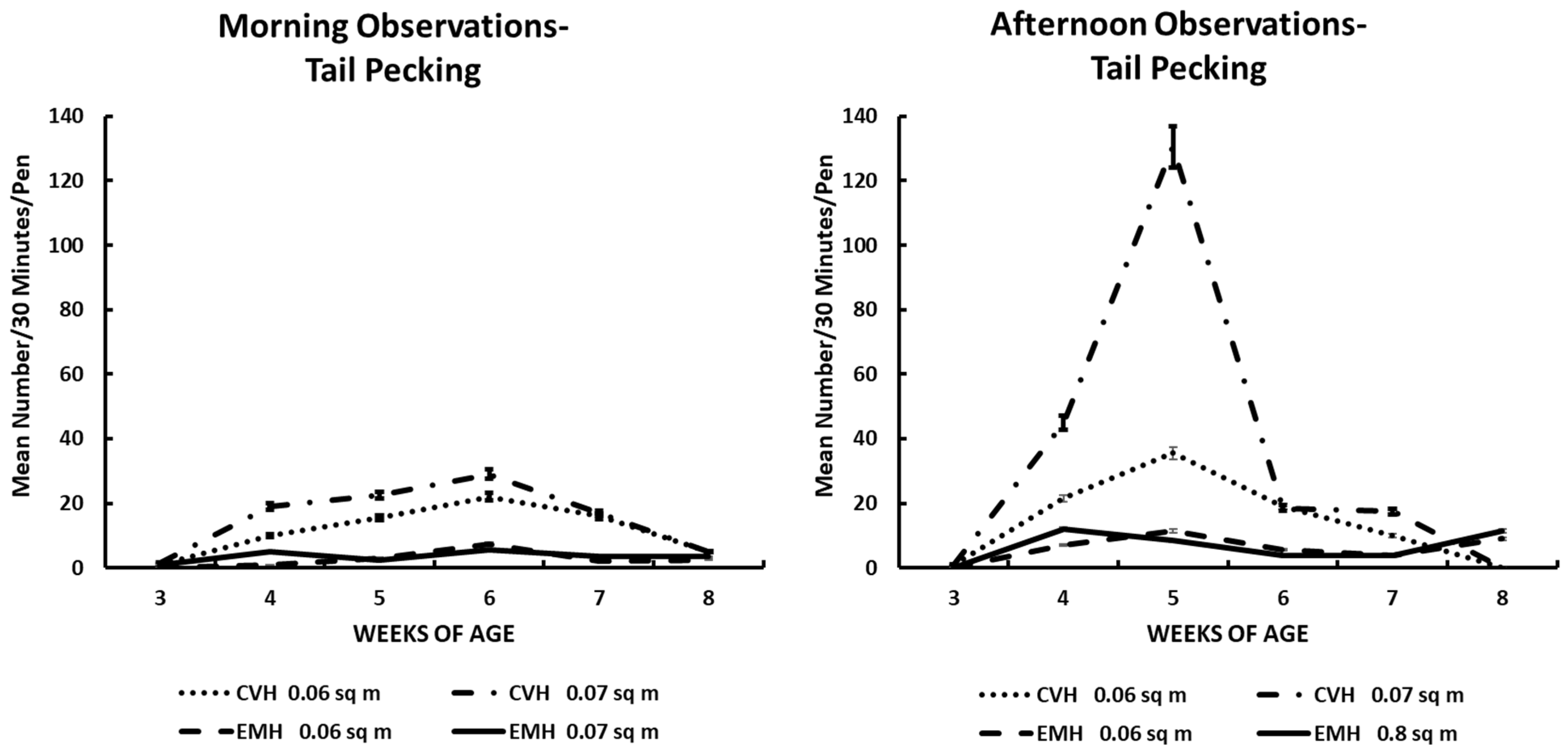
Tail pecking in the morning and afternoon in broiler chickens observed from three to eight weeks of age in conventional (CVH) and environmentally modified housing (EMH). Vertical bars on each data point represent the standard error of the mean.
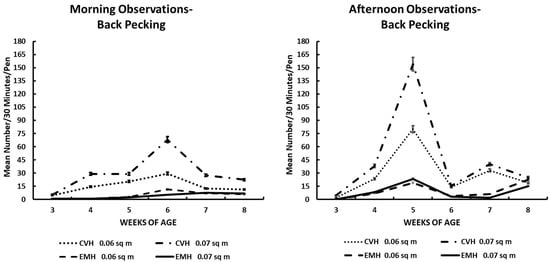
Figure 3.
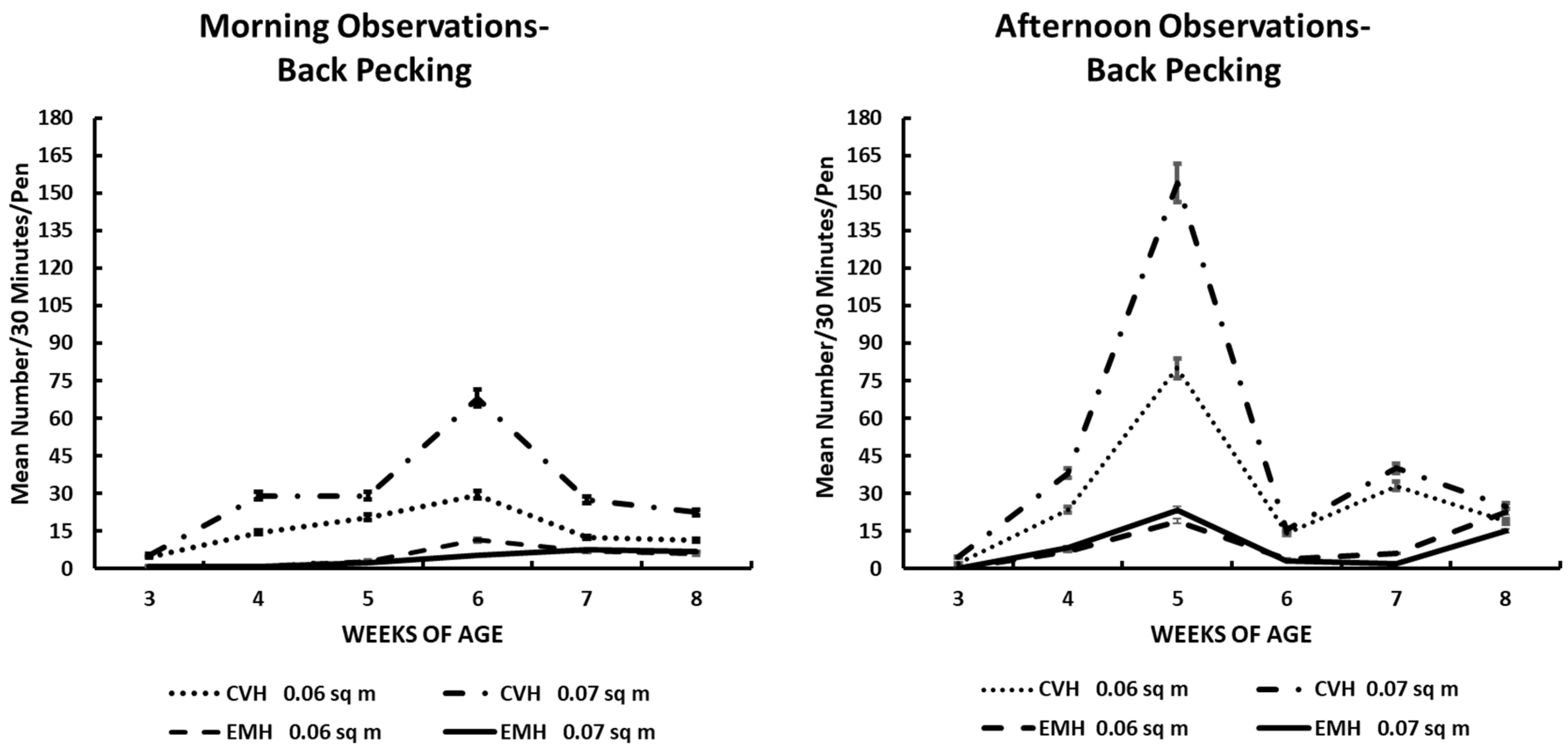
Back pecking activity in the morning and afternoon in broiler chickens observed from three to eight weeks of age in conventional (CVH) and environmentally modified housing (EMH). Vertical bars on each data point represent the standard error of the mean.
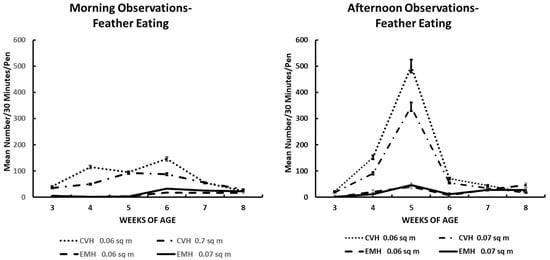
Figure 4.
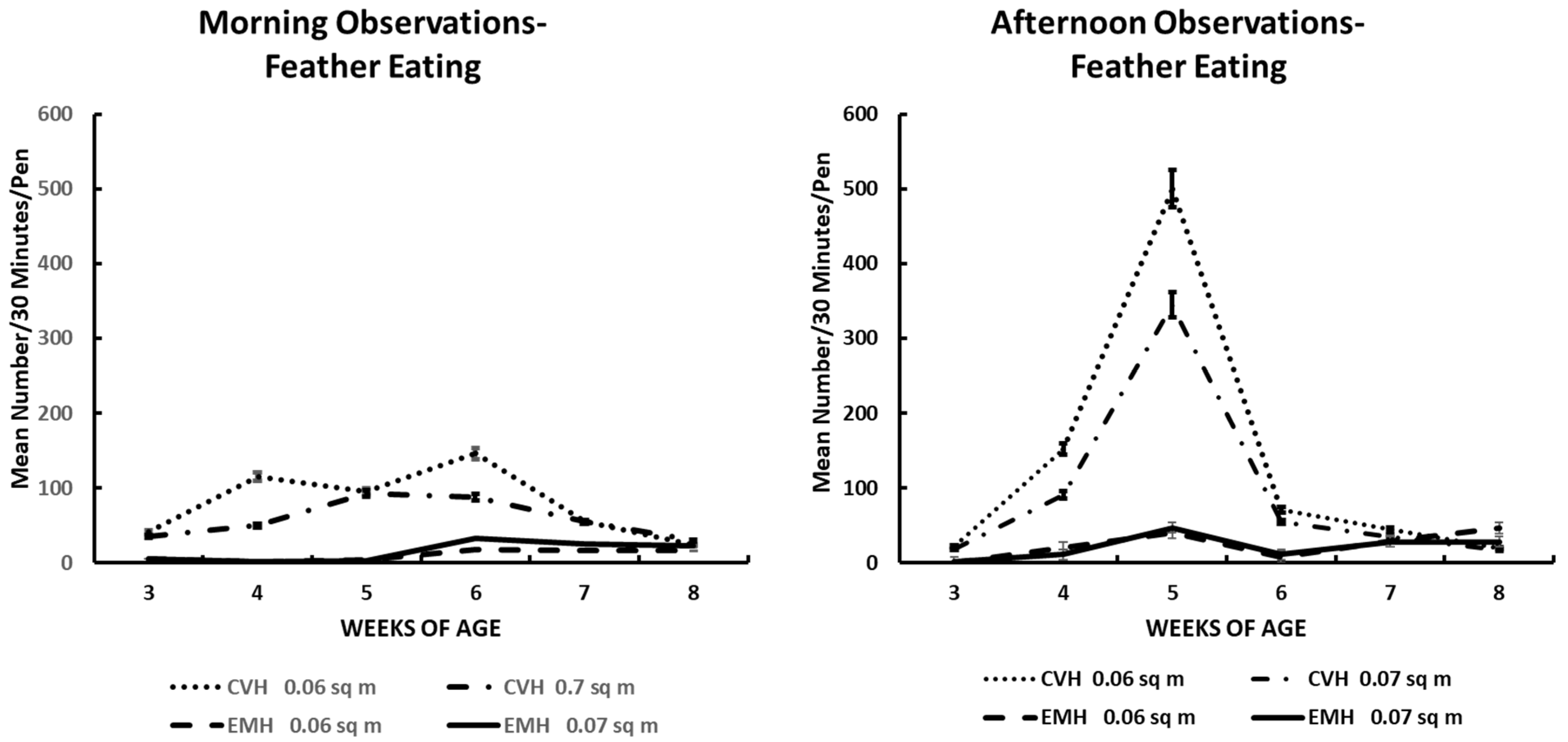
Feather eating activity in the morning and afternoon in broiler chickens observed from three to eight weeks of age in conventional (CVH) and environmentally modified housing (EMH). Vertical bars on each data point represent the standard error of the mean.
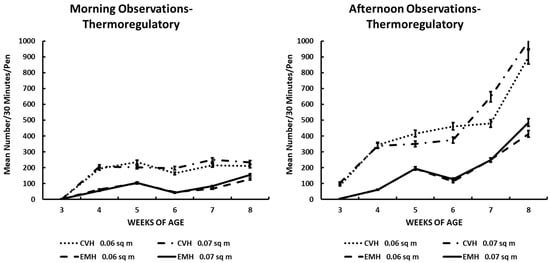
Figure 5.
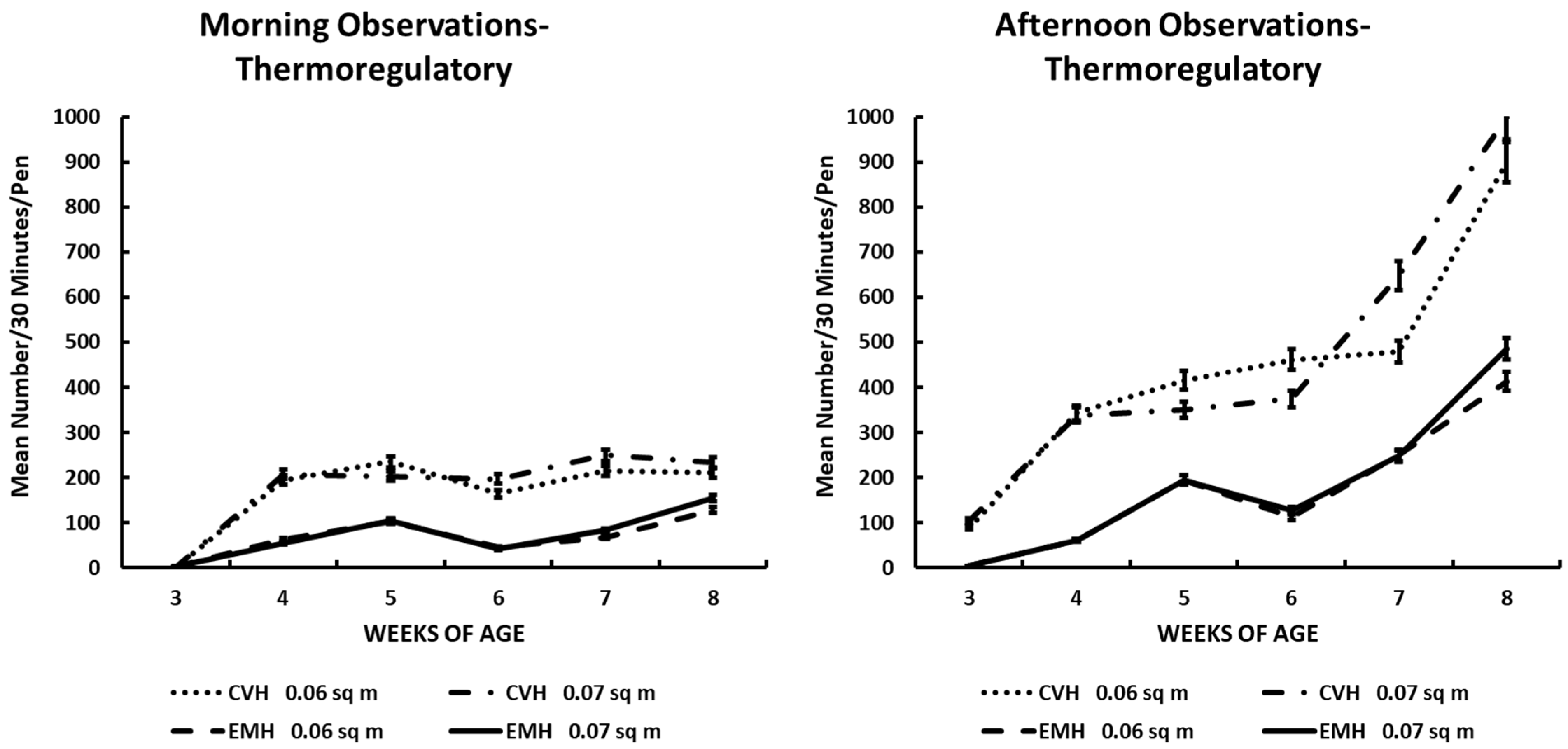
Thermoregulatory activity in the morning and afternoon in broiler chickens observed from three to eight weeks of age in conventional (CVH) and environmentally modified housing (EMH). Vertical bars on each data point represent the standard error of the mean.
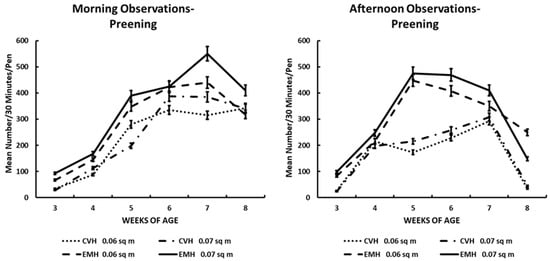
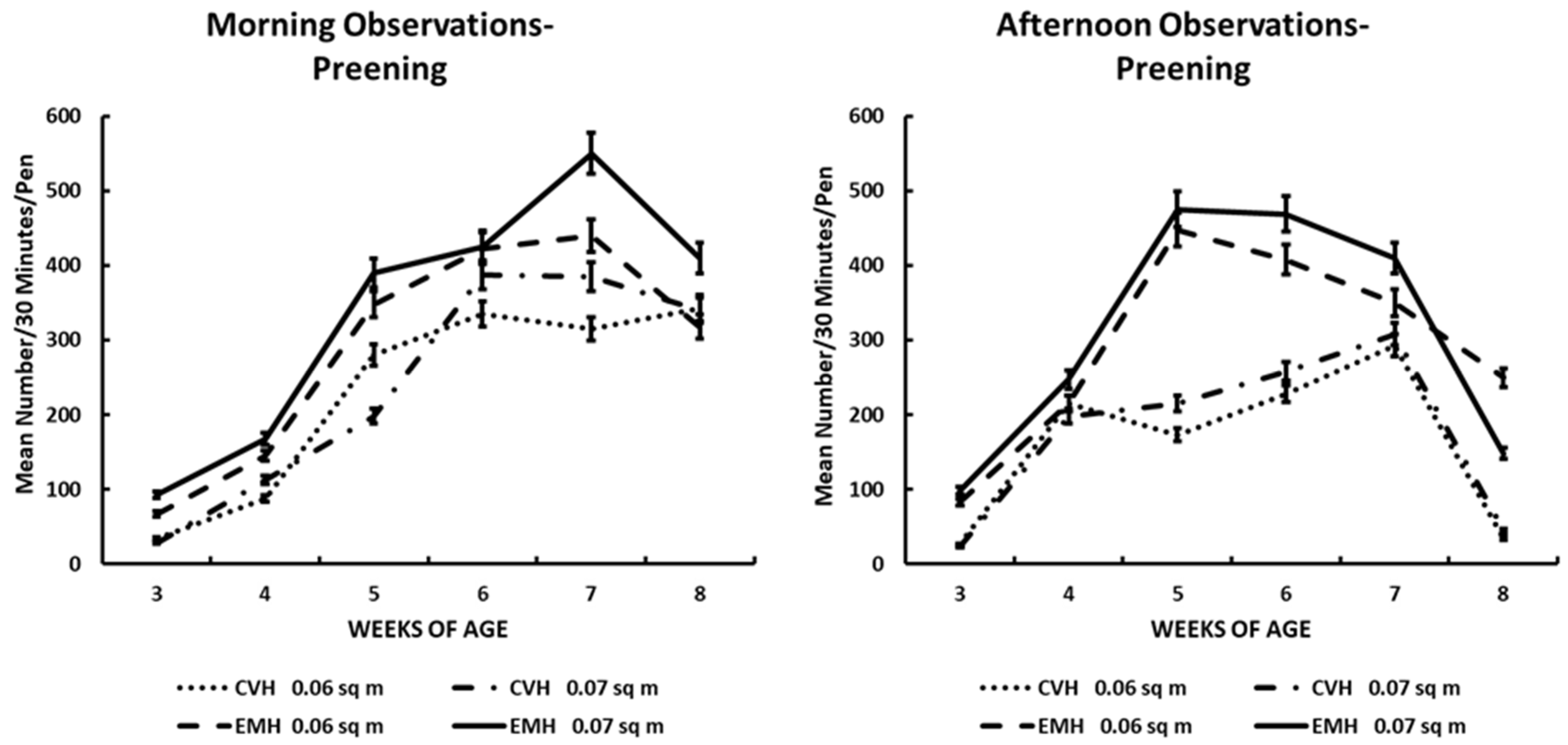
Figure 6.
Preening activity in the morning and afternoon in broiler chickens observed from three to eight weeks of age in conventional (CVN) and environmentally modified housing (EMH). Vertical bars on each data point represent the standard error of the mean.
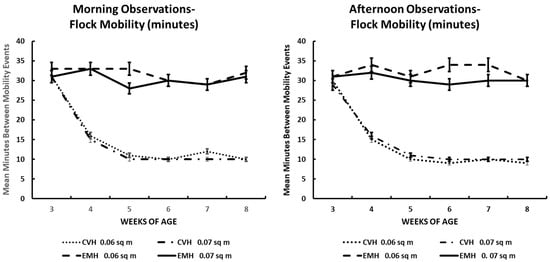
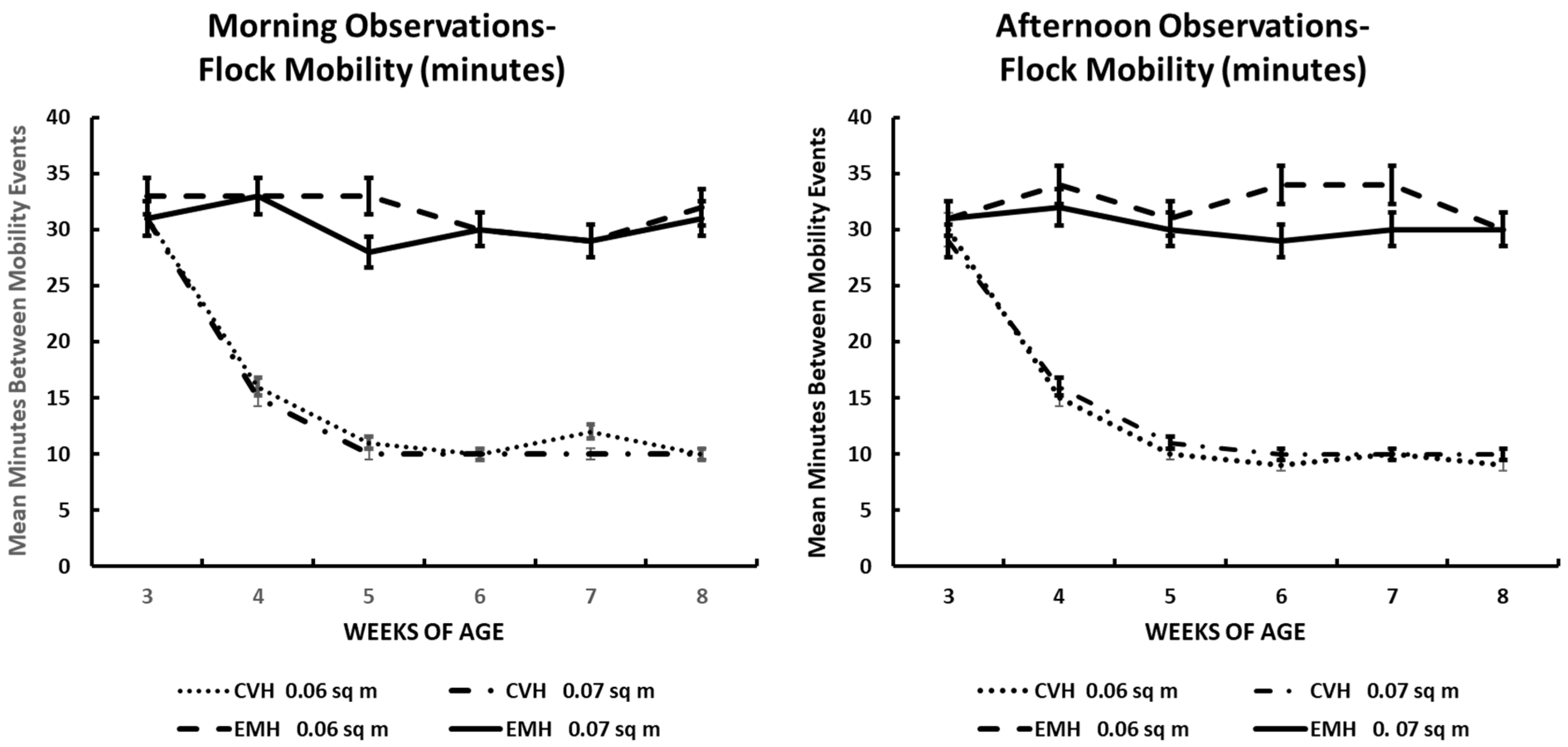
Figure 7.
Mean times (minutes) among whole-flock mobility events following thermoregulatory burrowing in the litter in the morning and afternoon in broiler chickens observed from three to eight weeks of age in conventional (CVH) and environmentally modified housing (EMH). Vertical bars on each data point represent the standard error of the mean.
3.2. Aggressive Behaviors
The total aggressive encounters for morning and afternoon times over the six morning/afternoon observation periods among large pens in the two housing types are shown in Table 1 and Figure 1. The average frequency of aggressive encounters was more numerous in the birds in CVH compared to EMH. Aggressive encounters were greater in the morning in CVH, being elevated significantly as early as 3 wk of age. There was a trend toward increased aggressive encounters due to HD in CVH, which was mirrored in the afternoon in CVH (Table 1; Figure 1). In EMH, aggressive encounters were less than in CVH in both the morning and afternoon, in the respective groups (Figure 1). At 4 and 5 wk of age, during both the morning and afternoon, aggressive encounters spiked in CVH in both HD and LD, and even in EMH, aggressive encounters also increased but not to the level found in CVH. The lower frequency of aggressive encounters in EMH was attributed to a lower light intensity compared to the higher light intensity in CVH. In the time period of 4 to 5 wk of age, when the grower diet was given initially to the chickens, it was discovered that the grower diet lacked sodium chloride. When the sodium chloride was added in a remixed diet, the aggressive encounters began to decline in both houses by 6 wk of age and remained low until 8 wk of age (Table 1; Figure 1).
3.3. Tail and Back Pecking Behavior
Tail pecking appeared to follow the pattern of aggressive encounters (Table 1; Figure 2). There was a clear demarcation of this behavior in the two houses, with greater frequencies found in CVH. Tail feather pecking increased in the birds in CVH by 4 wk of age and remained elevated through 7 wk of age in both population densities in CVH during both the morning and afternoon times (Table 1; Figure 2). In EMH, tail pecking was elevated from wk 4 through wk 5 when the behavioral display decreased rapidly with remixing of the feed (Figure 2). Interactions of the main effects were all significant (Table 1); generally, the interactions associated with time of day, weeks of age, house, and density were significant. The fact that all of these main effects contributed to the many interactions was further demonstrated by the presence of a significant four-way interaction among the four main effects (Table 1).
Back pecking was similar in profile compared to tail pecking in both houses (Table 1; Figure 3). Back pecking behavior was barely evident at 3 wk of age, but at 4 wk through 6 wk, chicks in CVH showed significantly elevated expressions of this behavior both in the morning and the afternoon. After peak expression of this behavioral activity between 5 and 6 wk of age, the expression of this behavior, exhibited primarily in CVH, decreased dramatically (Figure 3) after remixing of the feed. As in the tail pecking behavior, significant interactions among the main effects of house, time of day, and weeks of age were significant, but bird density in pens did not contribute significantly to the interactions of the main effects (Table 1).
3.4. Feather Eating Behavior
Feather eating (Table 1; Figure 4) was not always associated with feather extraction, as seen with tail and back pecking. Many birds per pen could be involved in feather eating from areas of a bird’s body other than the wing or back (behaviors not recorded per location of body feathers eaten). As can be seen in Figure 4, the incidence of feather eating was primarily associated with HD birds in CVH and secondarily with birds in LD groups in CVH (Figure 4). Very little feather eating was observed in EMH over the experimental periods. As associated with other behaviors, the highest rate of feather eating was during the 4 to 6 wk of age during the morning hours and 4 to 5 wk of age during the afternoon hours (Figure 4) in association with low dietary salt concentrations. The rate of feather eating was definitely associated with the main effects of housing, time of day, and weeks of age, and this was associated with significant interaction among these main effects, including a four-factor interaction (Table 1).
3.5. Burrrowing in Litter
Burrowing in the litter was a behavior associated with an attempt to thermoregulate during the extraordinary heat and humidity of the June–July summer period in North Carolina. The expression of burrowing behavior was readily associated with house temperature and outside temperature, along with prevailing RH% in the houses and outside the houses (Table 1 and Table 2; Figure 5). The largest number of birds involved in burrowing behavior was found in CVH (Table 1; Figure 5) in the afternoon. The increase in the number of birds showing burrowing behavior during the afternoons during wk 7 and 8 of the experiment was clearly associated with high ambient temperatures during the last 2 wk of the grow-out (Table 2; Figure 5). Most of the interactions among the main effects noted for burrowing behavior were due to house type, time of day, and weeks of age (Table 1 and Table 2). Pen density had little to contribute to interactions except that in CVH, slightly fewer birds in HD pens were burrowing in the afternoons in CVH during the last week of the study.
3.6. Preening
Preening is a behavior that one associates with feather grooming in birds in general. In this experiment, preening was very low when the chicks were 3 wk old. As the chicks aged through the growing period, the number of birds in each large pen in the two house types increased significantly with weeks of age (Table 1; Figure 6). The increase in preening frequency also was associated significantly with housing type, illustrated by more preening behaviors observed in EMH than in CVH (Table 1; Figure 6). The housing and weeks-of-age effects caused a significant week x house interaction (Table 1), which was influenced significantly by the time of day. When analysis of the time of day was addressed, it revealed that birds in EMH maintained a higher frequency of preening in the afternoon than birds in CVH (Table 1; Figure 6). During the final week of the grow-out period, the hottest temperatures of the season (Table 2) had a significant influence on preening frequency in all birds in both CVH and EMH, which resulted in significant decreases in preening frequency (Table 1; Figure 6). In the hottest time of the grow-out period, preening in CVH decreased to about a total of 50 chicks in large pens in both HD and LD; in EMH, the HD pens held about 175 chicks that were preening, and LD pens held about 230 preening chicks. The deep decreases in preening behavior were in direct response to increased house temperatures at 8 wk of age. Even in the morning, during the final week of the study, it was evident that the increasing house temperature (Table 1 and Table 2) was associated with the decrease in preening behaviors (Table 1; Figure 6).
3.7. Flock Mass Movement
Whole-flock mobility was very well defined. In this instance, nearly all chicks engaged in burrowing behavior in each pen. During these periods, when burrowing was being exhibited, the whole flock in the two houses was involved. During the morning times in both houses, beginning at 3 wk of age, those chicks involved in burrowing would fluff/dig into the litter and rest there for 30–35 min, followed by a drifting wave of chickens that would arise from their burrow and seek feed and water, accompanied by much vocalization. Over a period of less than 2 min, all of the chicks would arise from their burrows, presenting the same behaviors. At 4 wk of age, there was a change in the frequency of exhibition of this behavior in both the morning and afternoon—the time that chicks in CVH spent resting in their burrows decreased to about 15–17 min. Yet, in EMH, residence time in their litter burrows remained about the same in the morning and increased to about 34 min in the afternoon. At 5 wk in CVH, residence time in the litter burrows decreased to about 9–10 min in both the morning and afternoon. Residence in the litter burrows remained around 10 min in CVH in the 6–8 wk periods during both the morning and afternoon (Figure 7). In EMH, residence in litter burrows remained in the 30–33 min range and did not differ significantly between HD and LD pens. Yet, there was a highly significant difference between the two house types throughout the grow-out period (Table 1; Figure 7). Perusal of the ambient temperatures inside each house and ambient temperatures outside the houses on the days of observations revealed that inside temperatures in the CVH reflected an increase along with increasing outside ambient temperatures (Table 2). However, in EMH, inside temperatures were more stable than in CVH. Nevertheless, during the last 3 wk of the grow-out, the increasing outside ambient temperature also induced a 1–3 °C increase inside EMH and a 6–7 °C increase in CVH in the afternoon. This did not cause a decrease in flock burrowing before the induction of mobility in EMH as seen in CVH, but close inspection of residence time in litter burrows showed that a 3 min decrease in resting time was associated with increased internal temperature in EMH during the last 3 wk (Figure 7).
3.8. House Influence on Mortality
Total mortality differed significantly between the two houses. The total mortality in CVH was 6.37% in LD (0.07 m2/bird) population, and total mortality was 6.93% in HD (0.06 m2/bird). In EMH, the lower population density (0.07 m2/bird) had a total mortality of 2.68%, and HD (0.06 m2/bird) had a mortality of 2.94%. The differences in mortality between the houses were significant (p < 0.05), but total mortality differences within houses and between population densities were not different (p > 0.05). It is important to note that the mortality percentage per house type increased during the final 2 wk of the study when ambient temperatures increased (Table 2).
4. Discussion
Observation of poultry behaviors can yield information relative to the comfort and welfare of the birds [11]. During the course of a chicken’s life, one of the most important social behaviors expressed is aggressive behavior. Aggressive behaviors, such as fighting and pecking, are innate behaviors that allow for the establishment of social orders, especially in small groups. Work with small groups of broiler chickens, ranging from 50 to 200 chickens per group, suggested that the establishment of social orders (dominance hierarchies) among commercially grown broilers does exist, but as broilers aged there was a significant reduction in agonistic behaviors, which was attributed to the lack of importance of individual recognition in large groups [21]. However, it was noted in this study that aggressive behaviors peaked around 4 weeks of age, similar to earlier reports [21]. In the current investigation, peak aggressive behaviors also occurred at 4 wk of age, but depending on time of day, salt availability in the grower diet, increased ambient temperature, and housing differences, the aggressive behaviors were still elevated in the CVH until 6 wk of age. In EMH, aggressive behaviors peaked at 4 wk of age but plateaued at 5 wk of age in HD in EMH in the afternoon. These observations, placed in perspective, revealed that aggressive behaviors in a larger flock, simulating conditions in the commercial broiler industry in the Southeastern United States, suggested that the expression of aggressive behaviors in broilers is impacted by dynamic environmental conditions. There appeared to be significant modifying environmental conditions such as bright light intensity in CVH vs. the low light intensity in EMH and increased ambient temperatures with elevated HIs in both housing types.
Feather pecking could easily be a component of aggressive behavior. Indeed, feather pecking can lead to cannibalism, which must be rectified before it erupts into a major condition in which the health and welfare of the flock are beyond correction [17,22]. Tail (Figure 2) and back (Figure 3) feather pecking and feather eating (Figure 7) are persistent behaviors that were most prevalent in the afternoon periods, especially in CVH. Feather pecking has the potential to cause skin lesions that could lead to skin infections, down-grading of carcasses, and increased mortality, which would imply reduced health and welfare in such flocks. Feather pecking can be exacerbated by genetics and environmental factors; among the environmental factors, nutrition and/or the feeding drive might be one of the critical factors involved in the increased frequency of feather pecking and eating [22]. Mixed results have been reported in terms of feather pecking and eating relative to dietary protein sources. When bantam layers were fed either all vegetable protein or all animal protein as dietary supplements, it was noted that there was increased feather pecking and feather coat damage associated with animal protein diets [23]. The same group was fed similar diets to commercial layer stock, resulting in an increased frequency of feather pecking and eating in hens fed an all-vegetable protein diet [24]. In the current study, it was not possible to associate dietary protein sources with feather pecking and eating behaviors because the dietary protein source came from a mixture of soybean and poultry meals. Nevertheless, there was a significant difference between houses for feather pecking and eating with the greatest frequencies found in CVH. The higher light intensity in CVH was associated positively with increased expression of feather pecking and feather eating compared to the expression of these behaviors in EMH. One could offer cavalierly that the birds were better able to see each other in CVH than in EMH. This conclusion might not be far-fetched as it has been reported that broilers reared in low-intensity light appear to develop bupththalmia [25,26,27]. Thus, it is possible that a lower frequency of vision-related behaviors of broilers can be attributed to the lower light intensity in EMH.
The rise in ambient temperature and HI, especially in CVH, cannot be ignored as it was associated with decreased feather pecking activity during the last 3 wk of the grow-out. As the HI increased, there was decreased feather pecking in both the CVH vs. EMH during both morning and afternoon observation periods. These results were somewhat similar to results from a behavioral study on heat stress and feather conditions of broilers fed various levels of Brazilian propolis, a mixture of pollen and beeswax collected by bees foraging certain plants and trees which are rich in flavonoids—a class of antioxidants [28,29]. In a study with propolis, there were no feather pecking differences among the chickens exposed to elevated ambient temperatures [30]. Conclusions drawn from these observations could easily be associated with the broiler’s ability to reduce activity in response to elevated ambient temperature as a means to improve the regulation of body temperature and energy allocation [16].
Thermoregulatory activity as illustrated by the burrowing behavior also implied that general activity was reduced by the increasing daily ambient external temperatures associated with central North Carolina during this exceptionally hot and humid summer season. The burrowing behavior clearly showed that thermal conditions in CVH were somewhat uncomfortable for the broilers. From 4 wk to 8 wk of age, significantly greater numbers of broilers in CVH engaged in this behavior than in EMH, and this was made manifest when one peruses the results depicted in Figure 4. There was at least a two-fold greater number of birds in CVH engaged in this burrowing behavior compared to those birds in EMH. The expression of this thermoregulatory behavior was dramatically greater in CVH during the hottest part of the day during the afternoon observation sessions, especially during the final 3 wk of the trial.
One important conclusion from the burrowing behavior was that there was a negative relationship between the burrowing behavior and general activities of the flocks. Thus, the decrease in the expression of behaviors other than burrowing was related to increasing daily ambient temperatures.
Preening in poultry often is considered a comfort behavior performed in synchrony within a flock [31]. In this investigation, during the morning and afternoon observation periods, there were differences between housing types with preening frequency being greater in EMH compared with CVH. Additionally, there was a time effect showing increased preening frequency with age in each house. However, in the afternoon, there was a clear difference in preening frequency, with a lower frequency being exhibited by broilers in CVH compared to those in EMH. The differences observed between the two housing systems in the afternoon could be addressed in two divergent thought processes. First, from 3 to 5 wk of age, the increased frequency of preening in the EMH could possibly be reflecting a more comfortable temperature environment, and the preening would be considered a comfort behavior, which the birds would perform as if there were no other essential behaviors to perform. From 6 to 8 wk of age, afternoon preening behavior decreased by approximately 80% from a high frequency at 5 wk of age. Decreased preening could be associated with increasing ambient temperature to which the broilers in EMH were not accustomed. On the other hand, in the afternoon, a lower number of broilers in CVH expressed preening behavior. Whether the broilers in CVH had become acclimated to increasing daily temperatures or whether they were in fact responding to chronic cyclical heat distress on a daily basis by expressing displacement preening behavior is not readily apparent [31,32,33,34]. Yet, the possibility exists that the birds in CVH were more uncomfortable due to elevated afternoon HIs likely causing modified displacement preening behavior. Certainly, increasing daily HIs acted as a very potent stressor in chickens [31,35,36]. It has been reported that handling is a stressor in birds and can cause displacement preening in pigeons, which probably induced the activation of the hypothalamic–pituitary–adrenal cortical axis [32]. This hypothesis was tested using the administration of adrenocorticotropic hormone (ACTH) into the forebrain lateral ventricle in handled and non-handled broiler-type chickens [37]. Researchers observed that displacement preening was induced in handled and ACTH-treated chickens compared to saline- and naloxone-treated non-handled chickens. The anterior pituitary secretes ACTH in response to many types of stressors, which leads to increased secretory output of glucocorticoids from the adrenal cortex [38].
In this study, it was noted that the hottest ambient temperatures and HI, during 7 and 8 wk of age, caused a depression in preening behavior in broilers in both CVH and EMH during the afternoon observation times. Thus, in conditions where broilers were exposed to high temperature stressors, displacement preening appears to be diminished after the housing temperature exceeds 30 °C.
Flock mobility, the expression of whole-flock movement from resting in litter burrows, had intervals that were longer in the broilers in EMH vs. CVH. Prolonged intervals in the resting mode in the burrows in EMH might be considered an indication of comfort as opposed to very short intervals of rest in CVH. Displacement behaviors in broiler chickens might not be expressed only as preening behavior, but other types of behaviors might be expressions of displacement, as suggested by the decreased intervals of resting among broilers in CVH. Even though broilers burrowed to the surface of the earth under the litter in each house, the possibility of decreased heat transfer via conduction into the litter and the immediate earth surface appeared to be a driving factor in the expression of decreased resting intervals in broilers in CVH. As early as 3 wk of age, ambient temperature in the morning in CVH through 8 wk of age could range up to more than 2.5 °C greater than in EMH. In the afternoon, internal temperature in CVH could range to more than 12 °C greater than in EMH. The differences between the internal temperatures of CVH versus EMH certainly illustrate how heat distress modifies the expression of common behaviors in flocks of broilers.
However, for a short period of time, beginning at 4 wk of age, the grower diet was deficient in salt (NaCl). The error was discovered and corrected by 5 wk of age. It was during this time that there were peaks in aggressive encounters, back and tail pecking, and feather eating behaviors. After diet re-mixing with the addition of the salt, there were slow declines in the exhibition of the above-named behaviors in CVH. It has been reported that sodium-deficient diets can induce feather pecking in layers that leads to cannibalism [39]. In EMH, only small upward deviations in these behaviors were observed in broilers that were 4 to 6 wk of age. The difference between CVH vs. EMH was significant, but other modifying factors were at play. The modifying agents were light intensity differences between houses and daily increases in ambient temperature within CVH vs. EMH. Each factor appeared to have a differential influence on behaviors observed in the two housing systems. Thus, the housing system does appear to have a major influence on the exhibition of certain behaviors, and it has been documented that the welfare of chickens is dependent on the housing systems that might be used [40,41].
5. Conclusions
This investigation illustrated that broilers reared in EMH were generally more comfortable and experienced improved welfare compared to broilers reared in CVH. The improved welfare in EMH was shown by a greater than two-fold mortality rate in CVH vs. EMH. Resting behavior in litter had longer intervals among broilers in EMH, but resting intervals in CVH were less than 50% of those observed in EMH. Behaviors such as tail and back pecking, feather eating, and aggressive encounters were reduced in EMH vs. CVH. The daily increasing ambient temperatures and HIs impacted the behaviors of broiler chickens more severely in CVH than in EMH. The influence of housing density was not contributory to changes in the behavioral dynamics associated with either house type or age of the broilers. Ostensibly, our housing density, being lower than commercial standards in the United States possibly affected the expression of all social behaviors noted here, but additional study would be required to test this hypothesis. The modification of certain social behaviors among broiler chickens in different housing types in this investigation was believed to have also impacted livability and welfare. Additional research should prove that health and carcass quality might also be positively associated with the production of broilers in EMH.
Author Contributions
Conceptualization, F.W.E. and C.M.O.; methodology, F.W.E. and C.M.O.; software, F.W.E.; validation, F.W.E. and C.M.O.; formal analyses, investigation, data curation, resources, F.W.E.; writing, C.M.O. and F.W.E.; review and editing, F.W.E. and C.M.O.; supervision, F.W.E.; project administration and funding acquisition, F.W.E. All authors have read and agreed to the published version of the manuscript.
Funding
Funding for this student project was made available through support from the North Carolina Agricultural Research Service to the Prestage Department of Poultry Science (No. 418050) and Miscellaneous Gifts to the research program of FEW (662535).
Institutional Review Board Statement
This animal research study was approved by the NC State University Institutional Animal Care and Use Committee (Protocol No. 19-029-A, Approval on 12 September 2019), Box 75114, Raleigh, NC 27695-7514. The studies were conducted in accordance with the local legislation and institutional requirements.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are all included in the article. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
C = Celsius; m = meter; cm = centimeter; CVH = conventional house; EMH = environmentally modified house; HD = high population density; LD = low population density; H = house; D = population density; T = time of day, i.e., morning or afternoon; wk = weeks of age; BTU = British Thermal Units; LP = liquid petroleum; HI = heat index; RH% = percentage-relative humidity; h = hour; d = day; mj = megajoules; kg = kilogram; Kcal = kilocalories, NaCl = dietary table salt.
References
- Parkhurst, C.R.; Baughman, G.R.; Thaxton, J.P.; Garlich, J.D.; Edens, F.W. A comparison of broilers grown in environmentally modified and conventional housing at different population densities. Poult. Sci. 1977, 56, 883–885. [Google Scholar] [CrossRef]
- Liang, Y.; Kidd, M.T.; Watkins, S.E.; Tabler, G.T. Effect of commercial broiler house retrofit: A 4-year study of live performance. J. Appl. Poult. Res. 2013, 22, 211–216. [Google Scholar] [CrossRef]
- Xin, H.; Berry, L.L.; Baxter, T.I.; Tabler, G. Sidewall effects on energy use in broiler houses. J. Appl. Poult. Res. 1993, 2, 176–183. [Google Scholar] [CrossRef]
- Evans, L.; Brooks, G.C.; Anderson, M.G.; Campbell, A.M.; Jacobs, L. Environmental complexity and reduced stocking density promote positive behavioral outcomes in broiler chickens. Animals 2023, 13, 2074. [Google Scholar] [CrossRef]
- Zabir, M.; Miah, M.A.; Alam, M.; Bhulyan, M.E.J.; Haque, M.I.; Sujan, K.M.; Mustari, M. Impacts of stocking density rates on welfare, growth, and hemato-biochemical profile in broiler chickens. J. Adv. Vet. Anim. Res. 2021, 8, 642–649. [Google Scholar] [CrossRef]
- Shynkaruk, T.; Long, K.; LeBlanc, C.; Schwean-Lardner, K. Impact of stocking density on welfare and productivity of broiler chickens reared to 34 d of age. J. Appl. Poult. Res. 2023, 32, 100344. [Google Scholar] [CrossRef]
- Rault, J.-L.; Clark, K.; Groves, P.J.; Cronin, G.M. Light intensity of 2 or 20 lux on broiler behavior, welfare and productivity. Poult. Sci. 2017, 96, 779–787. [Google Scholar] [CrossRef]
- Soliman, F.N.K.; El-Sabrout, K. Light wavelengths/colors: Future prospects for broiler behavior and production. J. Vet. Behav. 2020, 36, 34–39. [Google Scholar] [CrossRef]
- Sumanu, V.O.; Naidoo, V.; Oosthuizen, M.C.; Chamunorwa, J.P. Adverse effects of heat stress during summer on broiler chickens production and antioxidant mitigating effects. Int. J. Biometeorol. 2022, 66, 2379–2393. [Google Scholar] [CrossRef]
- Sumanu, V.O.; Aluwong, T.; Ayo, J.O.; Ogbuagu, N.E. Evaluation of changes in tonic immobility, vigilance, malondialdehyde, and superoxide dismutase in broiler chickens administered fisetin and probiotic (Saccharomyces cerevisiae) and exposed to heat stress. J. Vet. Behav. 2019, 31, 36–42. [Google Scholar] [CrossRef]
- Neves, D.P.; Mehdizadeh, S.A.; Tscharke, M.; Nääs, I.d.A.; Banhazi, T.M. Detection of flock movement and behavior of broiler chickens at different feeders using image analysis. Inform. Process. Agric. 2015, 2, 177–182. [Google Scholar]
- Runnels, T.D.; Malone, G.W.; Klopp, S. The influence of feed texture on broiler performance. Poult. Sci. 1976, 55, 1958–1961. [Google Scholar] [CrossRef]
- Calet, C. The relative value of pellets versus mash and grain in poultry nutrition. World’s Poult. Sci. J. 1965, 21, 23–52. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.S.; Merrill, L.H.; Reedy, C.V.; McGinnis, J. Observations on eating patterns and rate of food passage of birds fed pellets and unpelleted diets. Poult. Sci. 1962, 41, 1414–1419. [Google Scholar] [CrossRef]
- Leone, E.H.; Estevez, I. Space use according to the distribution of resources and level of competition. Poult. Sci. 2008, 87, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Tickle, P.G.; Hutchinson, J.R.; Codd, J.R. Energy allocation and behaviour in the growing broiler chicken. Sci. Rep. 2018, 8, 4562. [Google Scholar] [CrossRef]
- Choct, M.; Hartini, S. Interaction between nutrition and cannibalism in laying hens. Rec. Adv. Anim. Nutr. Aust. 2003, 14, 157–162. [Google Scholar]
- Anderson, G.B.; Bell, M.L.; Peng, R.D. Methods to calculate the heat index as an exposure metric in environmental health research. Environ. Health Perspect. 2013, 121, 1111–1119. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Poultry, 9th ed.; National Academy Press: Washington, DC, USA, 1994; p. 155. [Google Scholar]
- SAS Institute. SAS/STAT Users’ Guide; SAS Institute, Inc.: Cary, NC, USA, 2016. [Google Scholar]
- Estevez, I.; Newberry, R.C.; de Reyna, L.A. Broiler chickens: A tolerant social system? Etologia 1997, 5, 19–29. [Google Scholar]
- Kjaer, J.B.; Bessei, W. The interrelationships of nutrition and feather pecking in the domestic fowl—A review. Archiv Geflügelk. 2013, 77, S.1–S.9. [Google Scholar] [CrossRef]
- Savory, C.J.; Mann, J.S.; Macleod, M.G. Incidence of feather pecking damage in growing bantams in relation to food form, group size, stocking density, dietary tryptophan concentration and dietary protein source. Br. Poult. Sci. 1999, 40, 579–584. [Google Scholar] [CrossRef] [PubMed]
- McKeegan, D.E.F.; Savory, C.J.; Macleod, M.G.; Mitchell, M.A. Developmental pecking damage in layer pullets in relation to dietary protein source. Br. Poult. Sci. 2001, 42, 33–42. [Google Scholar] [CrossRef]
- Blatchford, R.A.; Klasing, K.C.; Shivaprasad, H.L.; Wakenell, P.S.; Archer, G.S.; Mench, J.A. The effect of light intensity on the behavior, eye and leg health, and immune function of broiler chickens. Poult. Sci. 2009, 88, 20–28. [Google Scholar] [CrossRef]
- Deep, A.; Schwean-Lardner, K.; Crown, T.G.; Fancher, B.I.; Clausen, H.L. Effect of light intensity on broiler production, processing characteristics, and welfare. Poult. Sci. 2010, 89, 2326–2333. [Google Scholar] [CrossRef]
- Whitley, R.D.; Albert, R.A.; Brewer, R.N.; McDaniel, G.R.; Pidgeon, G.L.; Mora, E.C. Photoinduced bupthalmic avian eyes: II. Continuous darkness. Poult. Sci. 1985, 64, 1869–1874. [Google Scholar] [CrossRef]
- Havsteen, B.H. The biochemistry and medical significance of the flavonoids. Pharmacol. Therapeut. 2002, 96, 67–202. [Google Scholar]
- Hoşnuter, M.; Gurel, A.; Babuccu, O.; Armutcu, F.; Kargi, E.; Işikdemir, A. The effect of CAPE on lipid peroxidation and nitric oxide levels in the plasma of rats following thermal injury. Burns 2004, 30, 121–125. [Google Scholar] [CrossRef]
- Mahmoud, U.T.; Abdel-Rahman, M.A.M.; Darwish, M.H.A.; Applegate, T.J.; Cheng, H.-W. Behavioral changes and feathering score in heat stressed broiler chickens fed diets containing different levels of propolis. Appl. Anim. Behav. Sci. 2015, 166, 98–105. [Google Scholar] [CrossRef]
- Mench, J.A.; van Tienhoven, A.; Marsh, J.A.; McCormick, C.C.; Cunningham, D.L.; Baker, R.C. Effect of cage and floor pen management on behavior, production and physiological stress response of laying hens. Poult. Sci. 1986, 65, 1058–1069. [Google Scholar] [CrossRef]
- Delius, J.D. Preening and associated comfort behavior in birds. Ann. N. Y. Acad. Sci. 1988, 525, 40–55. [Google Scholar] [CrossRef]
- Duncan, I.J.H.; Wood-Gush, D.G.M. An analysis of displacement preening in the domestic fowl. Anim. Behav. 1972, 20, 68–71. [Google Scholar] [CrossRef]
- Fortomaris, P.; Arsenos, G.; Tserveni-Gousi, A.; Yannakopoulos, A. Performance and behavior of broiler chickens as affected by the housing system. Archiv Geflügelk. 2007, 71, S.97–S.104. [Google Scholar]
- Davis, G.S.; Edens, F.W.; Parkhurst, C.R. Computer-aided heat acclimation in broiler cockerels. Poult. Sci. 1991, 70, 302–306. [Google Scholar] [CrossRef]
- Wang, S.; Edens, F.W. Heat conditioning induces heat shock proteins in broiler chickens and turkey poults. Poult. Sci. 1998, 77, 1636–1645. [Google Scholar] [CrossRef] [PubMed]
- Williams, N.S.; Scampoli, D.L. Handling. ACTH, ACTH1-24, and naloxone effects on preening behavior in domestic chickens. Pharmacol. Biochem. Behav. 1984, 20, 681–682. [Google Scholar] [CrossRef] [PubMed]
- Scanes, C.G. Biology of stress in poultry with emphasis on glucocorticoids and the heterophil to lymphocyte ratio. Poult. Sci. 2016, 95, 2208–2215. [Google Scholar] [CrossRef]
- Cooke, B.C. Cannibalism in laying hens. Vet. Rec. 1992, 131, 495. [Google Scholar] [CrossRef]
- Dawkins, M.S.; Donnelly, C.A.; Jones, T.A. Chicken welfare is influenced more by housing conditions than by stocking density. Nature 2004, 427, 342–344. [Google Scholar] [CrossRef]
- Scheidler, S.E.; Shields, S. Cannibalism by Poultry (G1670); University of Nebraska-Lincoln Extension, Institute of Agriculture and Natural Resources: Lincoln, NE, USA, 2007. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).