
Membrane Engineering for Carotenoid Production in Escherichia coli

Abstract
1. Introduction
2. Results
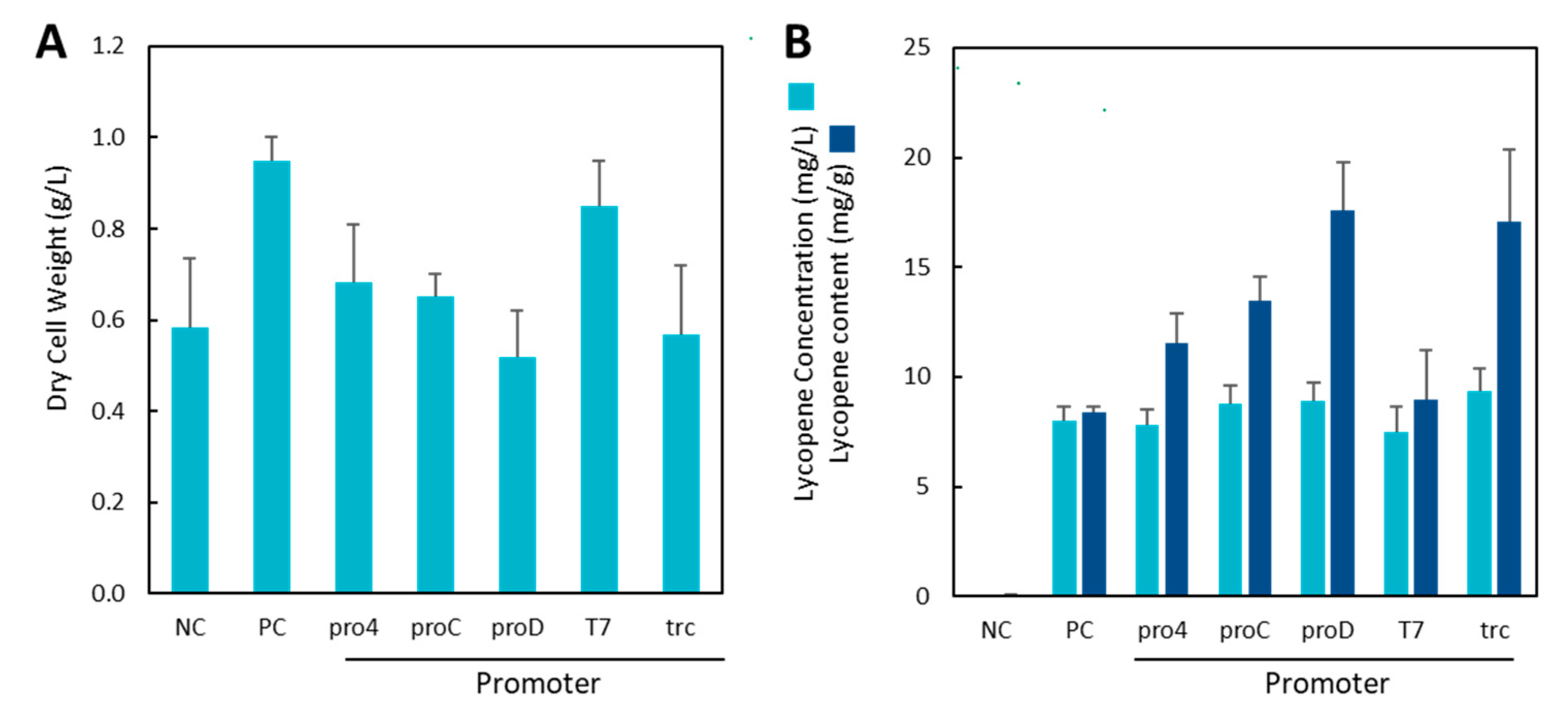
2.1. Construction of IUP Strains
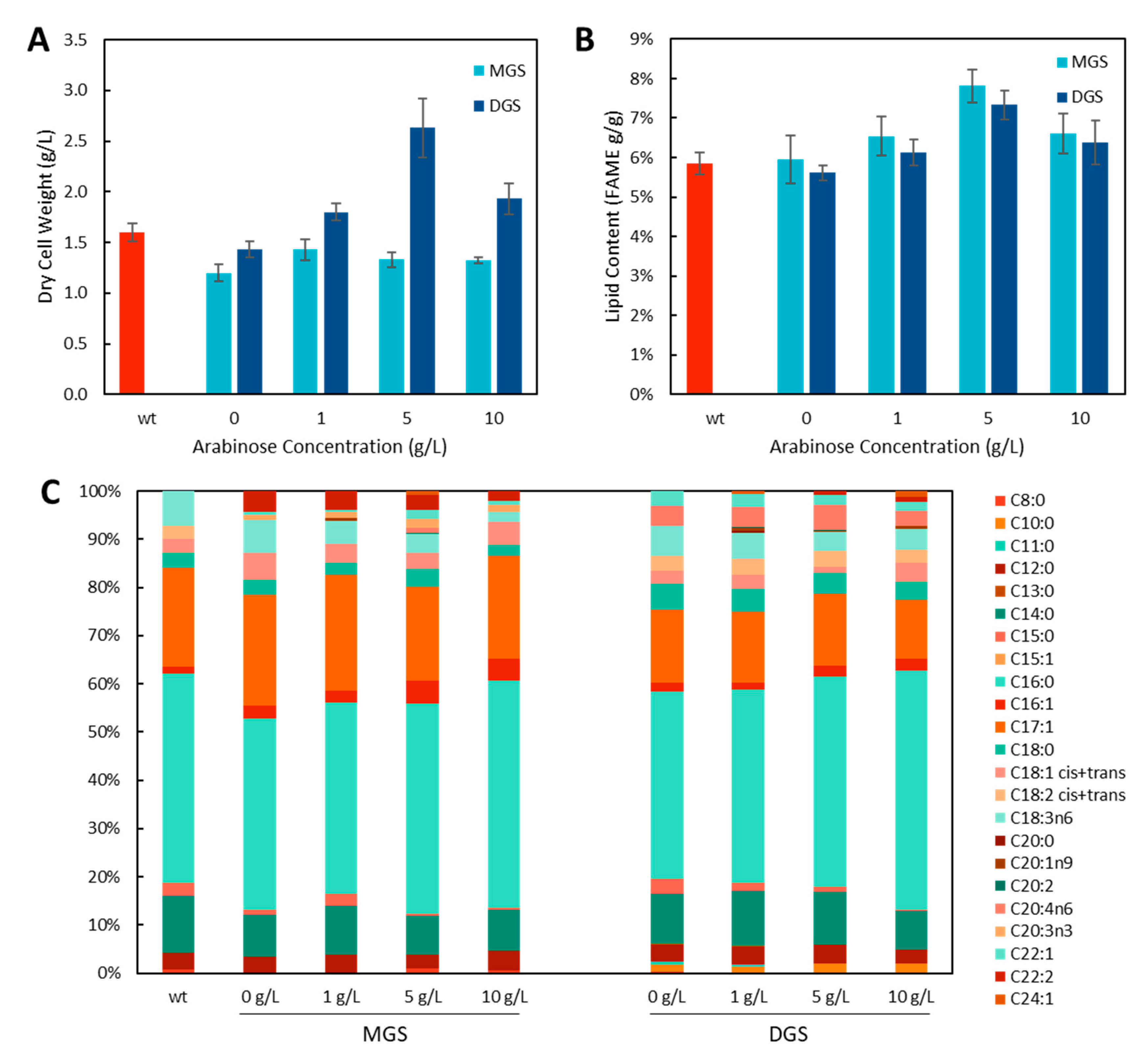
2.2. Increasing Membrane Capacity Using Membrane Modulating Enzymes
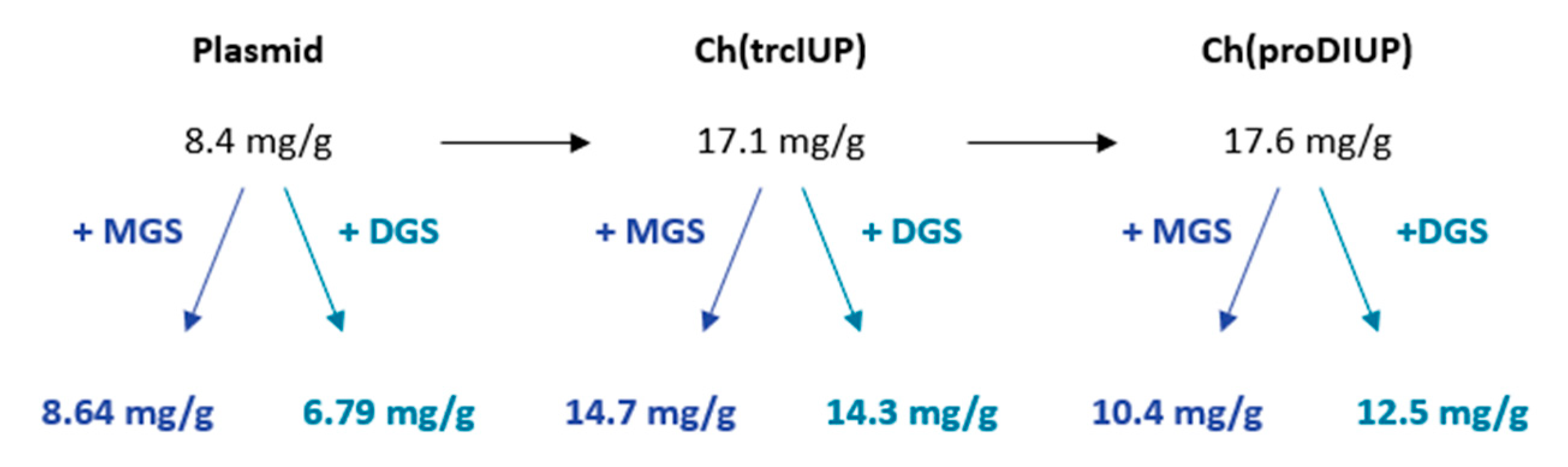
2.3. Lycopene Production in MGS and DGS Strains
2.4. Effect of MGS and DGS on β-Carotene Production
3. Discussion
4. Materials and Methods
4.1. Strains and Routine Cultivation
4.2. Cloning
4.3. Chromosomal Insertion
4.4. Western Blot
4.5. Production of Isoprenoids
4.6. Carotenoid Extraction
4.7. Total Lipid Content by Direct Transesterification to FAME
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A
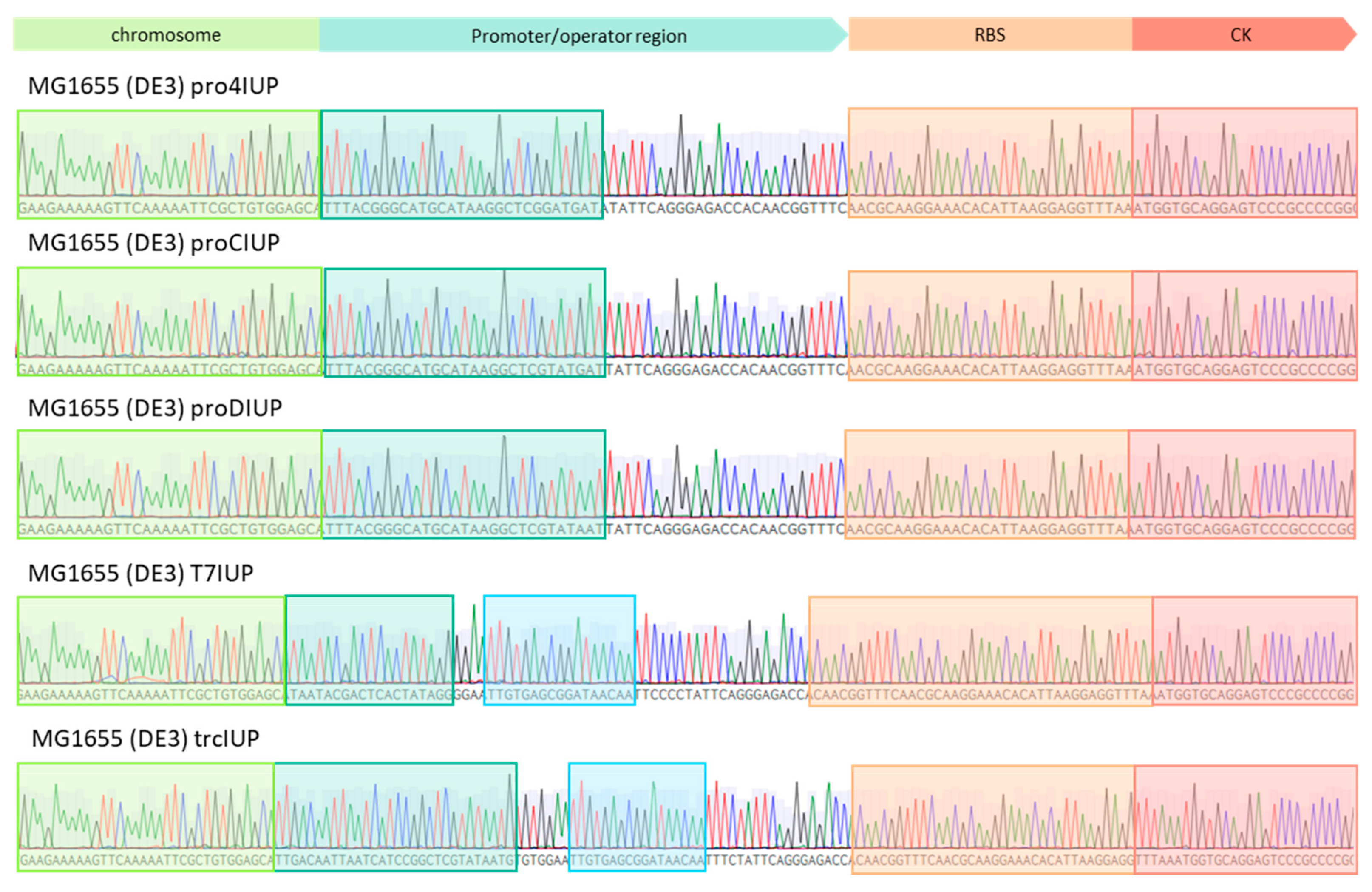

Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cloning Step | Primer Name | Primer Sequence (5′–3′) |
|---|---|---|
| Integration | Int_CP_fwd | gtactcagacgttgcagctg |
| Integration | Int_CP_rev | ggaatacgaccggaccgag |
| pCas-Cm | BB_Cm_fwd | taacggcaaagcagtcagagtagaatag |
| pCas-Cm | BB_Cm_rev | acgcctggtggtcagaccaagtttactc |
| pCas-Cm | CasCmC_2_fwd | gggtctgctatgtggtgcta |
| pCas-Cm | CasCmC_2_rev | ggtgagctggtgatatggga |
| pCas-Cm | CasCmC_fwd | gggtgatgctgccaacttac |
| pCas-Cm | CasCmC_SP1 | gggtgatgctgccaacttac |
| pCas-Cm | Cm_fwd | ttggtctgaccaccaggcgtttaagggc |
| pCas-Cm | Cm_rev | ctctgactgctttgccgttacgcaccac |
| pTargetT step 1 | HMR_fwd | tgaagatccttaacctgggcgtgctacc |
| pTargetT step 1 | HMR_rev | agatcctttaaagcagtatgtggcccgc |
| pTargetT step 1 | Ori_fwd | catactgctttaaaggatctaggtgaagatc |
| pTargetT step 1 | Ori_rev | ccttcctgctccacagcgaatttactagtattatacctaggactg |
| pTargetT step 1 | PT1C_fwd | aggcgagatcaccaaggtag |
| pTargetT step 1 | PT1C_rev | cagcagattacgcgcagaaa |
| pTargetT step 1 | SP1 | gcctgatgcggtattttctcc |
| pTargetT step 1 | Spec_fwd | aaattcgctgtggagcaggaagggttttagagctagaaatagcaag |
| pTargetT step 1 | Spec_rev | gcccaggttaaggatcttcacctagatc |
| pTargetT step 2 | IUP_Pro4_fwd | ctgtggagcatttacgggcatgcataagg |
| pTargetT step 2 | IUP_ProCD_fwd | tattcagggagaccacaacg |
| pTargetT step 2 | IUP_rev | attcccttcccagggttatgcagcggaaaag |
| pTargetT step 2 | IUP_T7_fwd | taatacgactcactataggggaattgtgagcggataacaattcccctattcagggagaccacaacg |
| pTargetT step 2 | IUP_trc_fwd | ttgacaattaatcatccggctcgtataatgtgtggaattgtgagcggataacaatttctattcagggagaccacaacg |
| pTargetT step 2 | PT_BB_fwd | cataaccctgggaagggaattaccgaatg |
| pTargetT step 2 | PT_BB_Pro4_rev | tgcccgtaaatgctccacagcgaatttttg |
| pTargetT step 2 | PT_BB_ProC_rev | cgttgtggtctccctgaataatcatacgagccttatgcatgcccgtaaatgctccacagcgaatttttg |
| pTargetT step 2 | PT_BB_ProD_rev | cgttgtggtctccctgaataattatacgagccttatgcatgcccgtaaatgctccacagcgaatttttg |
| pTargetT step 2 | PT_BB_T7_rev | ggggaattgttatccgctcacaattcccctatagtgagtcgtattatgctccacagcgaatttttg |
| pTargetT step 2 | PT_BB_trc_rev | gaaattgttatccgctcacaattccacacattatacgagccggatgattaattgtcaatgctccacagcgaatttttg |
| pTargetT step 2 | PT2_SP1 | tccggatatgaacaaactgca |
| pTargetT step 2 | PT2C_CP_fwd | tcgccaaccagactgctaat |
| pTargetT step 2 | PT2C_CP_rev | tcgatcccttgttcttggct |
| PCBFD1 backbone | bb_fwd | cgatgagggaatgataagtctgcgatgg |
| PCBFD1 backbone | bb_rev | gggcggattgcttattccagatgcgtgc |
| mgs/dgs fragment | dgsmgs_fwd | ctggaataagcaatccgccctcactacaac |
| mgs/dgs fragment | dgsmgs_fwd | gacttatcattccctcatcgacgccagag |
References
- Vickers, C.E.; Williams, T.C.; Peng, B.; Cherry, J. Recent advances in synthetic biology for engineering isoprenoid production in yeast. Curr. Opin. Chem. Biol. 2017, 40, 47–56. [Google Scholar] [CrossRef]
- Ram, S.; Mitra, M.; Shah, F.; Tirkey, S.R.; Mishra, S. Bacteria as an alternate biofactory for carotenoid production: A review of its applications, opportunities and challenges. J. Funct. Foods 2020, 67, 103867. [Google Scholar] [CrossRef]
- Jing, Y.; Wang, Y.; Zhou, D.; Wang, J.; Li, J.; Sun, J.; Feng, Y.; Xin, F.; Zhang, W. Advances in the synthesis of three typical tetraterpenoids including β-carotene, lycopene and astaxanthin. Biotechnol. Adv. 2022, 61, 108033. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, J.; Yang, R.; Yang, J. Metabolic Engineering Escherichia coli for the Production of Lycopene. Molecules 2020, 25, 3136. [Google Scholar] [CrossRef] [PubMed]
- Ward, V.C.A.; Chatzivasileiou, A.O.; Stephanopoulos, G. Metabolic engineering of Escherichia coli for the production of isoprenoids. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef] [PubMed]
- Chatzivasileiou, A.O.; Ward, V.; Edgar, S.M.; Stephanopoulos, G. Two-step pathway for isoprenoid synthesis. Proc. Natl. Acad. Sci. USA 2018, 116, 506–511. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, H.M.; Wessman, P.; Ge, C.; Edwards, K.; Wieslander, Å. Massive Formation of Intracellular Membrane Vesicles in Escherichia coli by a Monotopic Membrane-bound Lipid Glycosyltransferase. J. Biol. Chem. 2009, 284, 33904–33914. [Google Scholar] [CrossRef] [PubMed]
- Ge, C.; Gómez-Llobregat, J.; Skwark, M.J.; Ruysschaert, J.-M.; Wieslander, Å.; Lindén, M. Membrane remodeling capacity of a vesicle-inducing glycosyltransferase. FEBS J. 2014, 281, 3667–3684. [Google Scholar] [CrossRef] [PubMed]
- Palage, A.M.; Ward, V.C. Strategies for production of hydrophobic compounds. Curr. Opin. Biotechnol. 2022, 75, 102681. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Ye, L.; Zhao, D.; Li, S.; Li, Q.; Zhang, B.; Bi, C. Engineering membrane morphology and manipulating synthesis for increased lycopene accumulation in Escherichia coli cell factories. 3 Biotech 2018, 8, 269. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Low, Z.J.; Ma, X.; Liang, H.; Sinskey, A.J.; Stephanopoulos, G.; Zhou, K. Using biopolymer bodies for encapsulation of hydrophobic products in bacterium. Metab. Eng. 2020, 61, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Beltrán, J.; León-Sampedro, R.; Ramiro-Martínez, P.; de la Vega, C.; Baquero, F.; Levin, B.R.; Millán, S. Translational demand is not a major source of plasmid-associated fitness costs. Philos. Trans. R. Soc. B Biol. Sci. 2021, 377, 20200463. [Google Scholar] [CrossRef] [PubMed]
- Brechun, K.E.; Förschle, M.; Schmidt, M.; Kranz, H. Method for plasmid-based antibiotic-free fermentation. Microb. Cell Factories 2024, 23, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-T.; Lin, M.-S.; Hou, S.-Y. Multiple-Copy-Gene Integration on Chromosome of Escherichia Coli for Beta-Galactosidase Production. K. J. Chem. Eng. 2008, 25, 1082–1087. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, B.; Duan, C.; Sun, B.; Yang, J.; Yang, S. Multigene Editing in the Escherichia coli Genome via the CRISPR-Cas9 System. Appl. Environ. Microbiol. 2015, 81, 2506–2514. [Google Scholar] [CrossRef]
- Schaub, P.; Yu, Q.; Gemmecker, S.; Poussin-Courmontagne, P.; Mailliot, J.; McEwen, A.G.; Ghisla, S.; Al-Babili, S.; Cavarelli, J.; Beyer, P. On the Structure and Function of the Phytoene Desaturase CRTI from Pantoea ananatis, a Membrane-Peripheral and FAD-Dependent Oxidase/Isomerase. PLoS ONE 2012, 7, e39550. [Google Scholar] [CrossRef]
- Vikström, S.; Li, L.; Karlsson, O.P.; Wieslander, Å. Key Role of the Diglucosyldiacylglycerol Synthase for the Nonbilayer−Bilayer Lipid Balance of Acholeplasma laidlawii Membranes. Biochemistry 1999, 38, 5511–5520. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Jing, H.Y.; Li, X.; Zhang, F.; Sun, X.M. Rapid construction of Escherichia coli chassis with genome multi-position integration of isopentenol utilization pathway for efficient and stable terpenoid accumulation. Biotechnol. J. 2023, 18, e2300283. [Google Scholar] [CrossRef]
- Montague, T.G.; Cruz, J.M.; Gagnon, J.A.; Church, G.M.; Valen, E. CHOPCHOP: A CRISPR/Cas9 and TALEN web tool for genome editing. Nucleic Acids Res. 2014, 42, W401–W407. [Google Scholar] [CrossRef] [PubMed]
- Wolin, E.A.; Wolin, M.J.; Wolfe, R. 8 Formation of Methane by Bacterial Extracts*. J. Biol. Chem. 1963, 238, 2882–2886. [Google Scholar] [CrossRef]
- van Wychen, S.; Laurens, L.M.L. Determination of Total Lipids as Fatty Acid Methyl Esters (FAME) by In Situ Transesterification; Laboratory Analytical Procedure (LAP): Golden, CO, USA, 2013. [Google Scholar]




| Host/Strain | Description | Reference |
|---|---|---|
| NEB-5α | fhuA2 Δ(argF-lacZ)U169 phoA glnV44 Φ80 Δ(lacZ)M15 gyrA96 recA1 relA1 endA1 thi-1 hsdR17 | NEB |
| MG1655 (DE3) | ΔendA ΔrecA (λ DE3) | Addgene #37853 |
| pro4IUP | ΔendA ΔrecA (λ DE3), yabP::PPro4 (ck, ipk, idi):: rluA, KanR | This study |
| proCIUP | ΔendA ΔrecA (λ DE3), yabP::PProC (ck, ipk, idi)::rluA, KanR | This study |
| proDIUP | ΔendA ΔrecA (λ DE3), yabP::PProD (ck, ipk, idi)::rluA, KanR | This study |
| T7IUP | ΔendA ΔrecA (λ DE3), yabP::PT7lac (ck, ipk, idi)::rluA, KanR | This study |
| trcIUP | ΔendA ΔrecA (λ DE3), yabP::Ptrclac (ck, ipk, idi)::rluA, KanR | This study |
| PC | MG1655(DE3), pSEVA228pro4IUP | |
| Plasmids | Description | Reference |
| pSEVA228-pro4IUP | RK2, KanR, Ppro4 (ck, ipk, idi) | [6] |
| p5T7-lycipi-ggpps | pSC101, SmR, PT7lacUV (ggpps, ipi, crtI, crtB) | [6] |
| pAC-BETAipi | p15A, CmR, (crtEBIY, idi) | Addgene #53277 |
| p20-ara-mgs | pBR322, AmpR, ParaB (mgs) | This study |
| p20-ara-dgs | pBR322, AmpR, ParaB (dgs) | This study |
| p20-ara-mgs-his | pBR322, AmpR, ParaB (mgs-6xhis) | This study |
| p20-ara-dgs-his | pBR322, AmpR, ParaB (dgs-6xhis) | This study |
| pCBFD1 | pBR322, AmpR, ParaB (cbfd1, hbfd1) | Addgene #53364 |
| pTargetF | pBR322, SmR sgRNA empty scaffold | Addgene #62226 |
| pTarget-yabP | pTargetF with yabP-sgRNA and yabP/rluA homology region | This study |
| pTarget- pro4 | pTarget-yabP with yabP/PPro4 (ck, ipk, idi), KnR /rluA | This study |
| pTarget-proC | pBR322, SmR sgRNA-yabP/PProC (ck, ipk, idi), KnR/rluA | This study |
| pTarget-proD | pBR322, SmR sgRNA-yabP/PProD (ck, ipk, idi), KnR/rluA | This study |
| pTarget-T7 | pBR322, SmR sgRNA-yabP/PT7lac (ck, ipk, idi), KnR/rluA | This study |
| pTarget-trc | pBR322, SmR sgRNA-yabP/Ptrclac (ck, ipk, idi), KnR/rluA | This study |
| pCas | repA101(Ts) KanR Pcas-cas9 ParaB-Red lacIq Ptrc-sgRNA-pBR322 | Addgene #62225 |
| pCas-Cm | repA101(Ts) CmR Pcas-cas9 ParaB-Red lacIq Ptrc-sgRNA-pBR322 | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Bates, E.; Perera, D.S.; Palage, A.M.; Ward, V.C.A. Membrane Engineering for Carotenoid Production in Escherichia coli. SynBio 2024, 2, 349-362. https://doi.org/10.3390/synbio2040022
Li J, Bates E, Perera DS, Palage AM, Ward VCA. Membrane Engineering for Carotenoid Production in Escherichia coli. SynBio. 2024; 2(4):349-362. https://doi.org/10.3390/synbio2040022
Chicago/Turabian StyleLi, Jiaqing, Eileen Bates, Dylan S. Perera, Andreea M. Palage, and Valerie C. A. Ward. 2024. "Membrane Engineering for Carotenoid Production in Escherichia coli" SynBio 2, no. 4: 349-362. https://doi.org/10.3390/synbio2040022
APA StyleLi, J., Bates, E., Perera, D. S., Palage, A. M., & Ward, V. C. A. (2024). Membrane Engineering for Carotenoid Production in Escherichia coli. SynBio, 2(4), 349-362. https://doi.org/10.3390/synbio2040022