
Assessment of the Genetic Diversity of Ulex europaeus in Maui, California, Hawaii and New Zealand by a Method of Microsatellite Markers †



Abstract
1. Introduction
2. Experiments
2.1. Materials and Methods
2.2. Data Analysis
2.3. Estimation of Genetic Distance
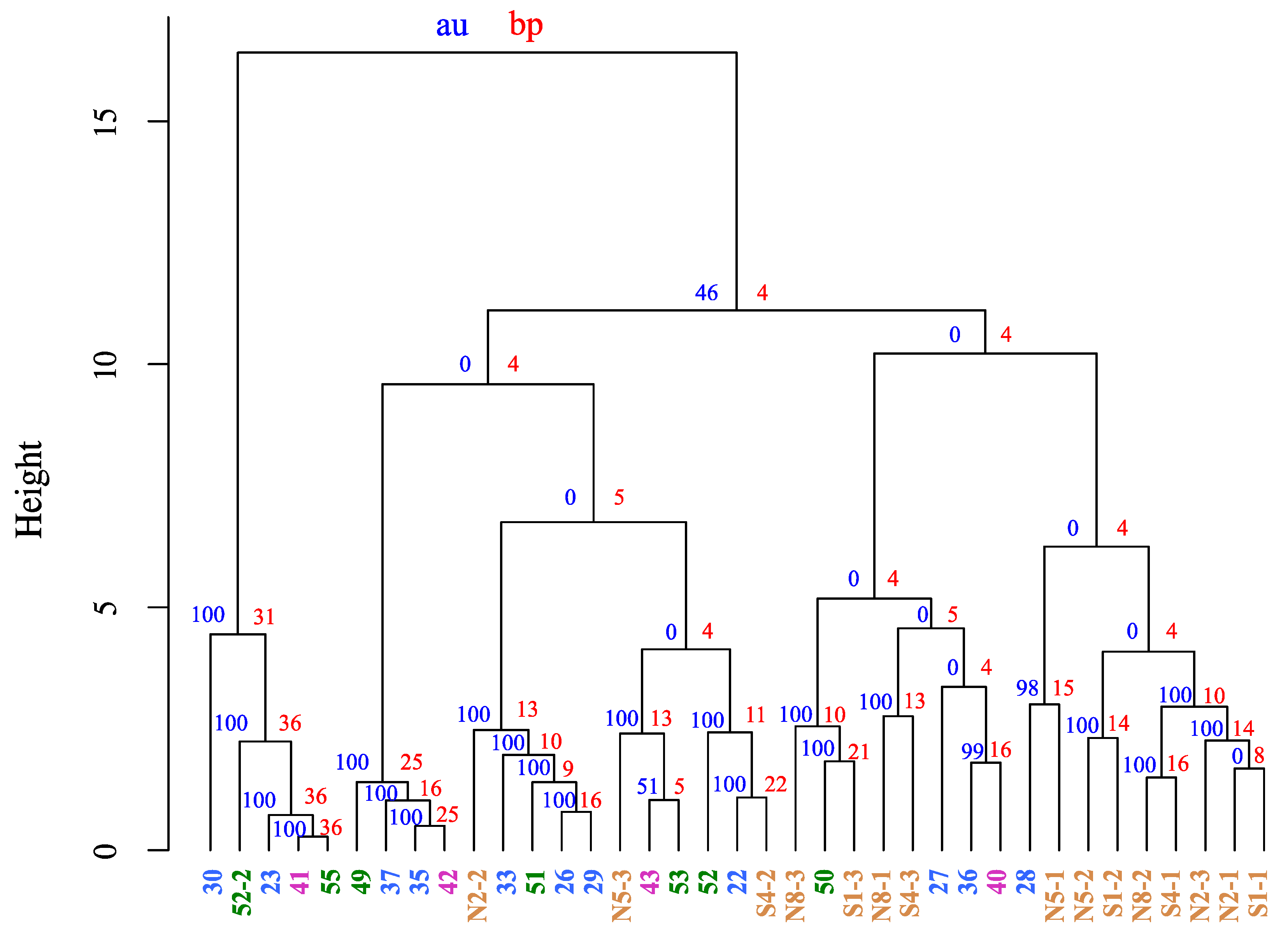
3. Results and Discussion
3.1. Cluster Analysis
3.2. Analysis of Molecular Variance
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Gao, C.; Ren, X.; Mason, A.S.; Li, J.; Wang, W.; Xiao, M.; Fu, D. Revisiting an important component of plant genomes: Microsatellites. Funct. Plant. Biol. 2013, 40, 645–661. [Google Scholar] [CrossRef]
- Ainouche, A.; Bayer, R.; Cubas, P.; Misset, M.T. Phylogenetic relationships within tribe Genisteae (Papilionoideae) with special reference to genus Ulex. In Advances in Lugume Systematics Part 10, Higher Level Systematics; Klitgarrd, B.B., Bruneau, A., Eds.; Royal Botanic Gardens: Kew, UK, 2003; pp. 232–252. [Google Scholar]
- Ainouche, A.; Mahe, F.; Affagard, M.; Ainouche, M.; Misset, M. Molecular evidence for an allopolyploid origin of the invasive European gorse, Ulex europaeus subsp. europaeus (Fabaceae, Genistae). In Proceedings of the International Conference on Polyploidity, Hybridization and Biodiversity, University of Rennes, Rennes, France, 17–20 May 2009; p. 202. [Google Scholar]
- Blischak, P.D.; Kubatko, L.S.; Wolfe, A.D. SNP genotyping and parameter estimation in polyploids using low-coverage sequencing data. Bioinformatics 2017, 34, 407–415. [Google Scholar] [CrossRef]
- Vieira, M.L.C.; Santini, L.; Diniz, A.L.; de Munhoz, C.F. Microsatellite markers: What they mean and why they are so useful. Genet. Mol. Biol. 2016, 39, 312–328. [Google Scholar] [CrossRef] [PubMed]
- Hornoy, B.; Atlan, A.; Roussel, V.; Buckley, Y.M.; Tarayre, M. Two colonisation stages generate two different patterns of genetic diversity within native and invasive ranges of Ulex europaeus. Heredity 2013, 111, 355–363. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2017, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Yáñez-Cuna, J.O.; Arnold, C.D.; Stampfel, G.; Boryń, Ł.M.; Gerlach, D.; Rath, M.; Stark, A. Dissection of thousands of cell type-specific enhancers identifies dinucleotide repeat motifs as general enhancer features. Genome Res. 2014, 24, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, Y.; Asano, R.; Kobayashi, Y.; Fukushima, J.; Takada, K. Comparison of NGS-SSR analyses and fragment analyses at SSR marker genotyping. J. Jpn. For. Sci. 2017, 128, 92. [Google Scholar] [CrossRef]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Location | Mother Tree | Latitude | Longitude | Altitude (m) | Habitat |
|---|---|---|---|---|---|
| Maui | #22 | N20.76 | W156.27 | 1758 | Ranch |
| #23 | N20.76 | W156.27 | 1823 | Ranch | |
| #26 | N20.77 | W156.26 | 1767 | Ranch | |
| #27 | N20.77 | W156.26 | 1763 | Ranch | |
| #28 | N20.77 | W156.26 | 1763 | Ranch | |
| #29 | N20.78 | W156.25 | 1773 | Ranch | |
| #30 | N20.78 | W156.25 | 1768 | Ranch | |
| #33 | N20.79 | W156.25 | 1656 | Ranch | |
| #35 | N20.8 | W156.28 | 1060 | Forest | |
| #36 | N20.8 | W156.28 | 1067 | Forest | |
| #37 | N20.8 | W156.28 | 1046 | Forest | |
| California | #40,41,42,43 | N37.15 | W122.34 | 33, 33, 34, 31 | Fallow land |
| Hawaii | #49 | N19.72 | W155.44 | 2004 | Ranch |
| #50 | N19.72 | W155.43 | 2058 | Ranch | |
| #51 | N19.93 | W155.41 | 2015 | Ranch | |
| #52 | N19.73 | W155.39 | 1952 | Ranch | |
| #52-2 | N19.73 | W155.39 | 1952 | Ranch | |
| #53 | N19.74 | W155.37 | 1929 | Ranch | |
| #55 | N19.77 | W155.36 | 1980 | Ranch | |
| New Zealand | N2-1, 2, 3 | S36.88 | E174.84 | 0 | Fallow land |
| N5-1, 2, 3 | S37.86 | E175.82 | 97 | Roadside of a ranch | |
| N8-1, 2, 3 | S38.99 | E175.76 | 538 | Fallow land with grass | |
| S1-1, 2, 3 | S43.8 | E173 | 623 | Ranch | |
| S4-1, 2, 3 | S43.5 | E172.52 | 38 | Roadside of a ranch |
| Locus | Primers | (Primers) (μM) |
|---|---|---|
| A110 | F: 5′-CTATGGTGAATTTGTGATACAC-3′ | 0.35 |
| R: 5′-ACCTTGTTGCATCTTTACC-3′ | ||
| A125 | F: 5′-GCATATACATACCCGAGGTAAG-3′ | 0.26 |
| R: 5′-AACCTGATGAAATGCACTATTC-3′ | ||
| B4 | F: 5′-GGGCTCTGGCTCTGATAC-3′ | 0.2 |
| R: 5′-TTGGATTAACCAACTTTCCTC-3′ | ||
| B104 | F: 5′-GAACCTTATTCACTGGAATCTG-3′ | 0.3 |
| R: 5′-CCCTTTTCTTTCCTTTCTTAAC-3′ | ||
| B123 | F: 5′-AATTTGCCTGACATTGTTACTC-3′ | 0.22 |
| R: 5′-AGACCGTGTTCATTATGGTTAG-3′ | ||
| Selected Loci | ||||||
|---|---|---|---|---|---|---|
| Sample NO | Population | A110 | A125 | B4 | B104 | B123 |
| 22 | M | 12 | 16 | 0 | 0 | 10 |
| 23 | M | 0 | 0 | 0 | 0 | 23 |
| 26 | M | 10 | 0 | 0 | 8 | 15 |
| 27 | M | 12 | 0 | 0 | 0 | 0 |
| 28 | M | 0 | 0 | 0 | 2 | 0 |
| 29 | M | 17 | 0 | 0 | 6 | 13 |
| 30 | M | 0 | 0 | 0 | 0 | 5 |
| 33 | M | 5 | 0 | 0 | 8 | 26 |
| 35 | M | 12 | 0 | 27 | 0 | 21 |
| 36 | M | 11 | 0 | 0 | 13 | 0 |
| 37 | M | 14 | 0 | 12 | 0 | 19 |
| 40 | C | 12 | 15 | 0 | 14 | 0 |
| 41 | C | 0 | 0 | 0 | 0 | 19 |
| 42 | C | 9 | 0 | 23 | 0 | 18 |
| 43 | C | 11 | 27 | 0 | 14 | 15 |
| 49 | H | 5 | 0 | 36 | 0 | 20 |
| 50 | H | 14 | 0 | 36 | 0 | 0 |
| 51 | H | 17 | 0 | 0 | 2 | 16 |
| 52 | H | 13 | 0 | 0 | 0 | 19 |
| 52-2 | H | 0 | 0 | 0 | 0 | 39 |
| 53 | H | 13 | 48 | 0 | 38 | 15 |
| 55 | H | 0 | 0 | 0 | 0 | 17 |
| N2-1 | N-N | 4 | 7 | 11 | 4 | 17 |
| N2-2 | N-N | 0 | 0 | 0 | 4 | 18 |
| N2-3 | N-N | 0 | 2 | 5 | 6 | 14 |
| N5-1 | N-N | 0 | 22 | 0 | 3 | 0 |
| N5-2 | N-N | 0 | 22 | 3 | 7 | 0 |
| N5-3 | N-N | 0 | 22 | 0 | 10 | 14 |
| N8-1 | N-N | 3 | 23 | 0 | 0 | 0 |
| N8-2 | N-N | 15 | 26 | 43 | 3 | 13 |
| N8-3 | N-N | 11 | 21 | 41 | 0 | 0 |
| S1-1 | N-S | 12 | 20 | 4 | 6 | 16 |
| S1-2 | N-S | 18 | 26 | 7 | 15 | 0 |
| S1-3 | N-S | 13 | 0 | 32 | 20 | 0 |
| S4-1 | N-S | 0 | 24 | 37 | 5 | 10 |
| S4-2 | N-S | 12 | 26 | 0 | 0 | 24 |
| S4-3 | N-S | 21 | 11 | 0 | 0 | 0 |
| Source | df | SS | MS | Est. Var. | % |
|---|---|---|---|---|---|
| Among Pops | 4 | 2960.2 | 740.05 | 27.79 | 5% |
| Within Pops | 32 | 17,276.99 | 539.91 | 539.91 | 95% |
| Total | 36 | 20,237.19 | 567.69 | 100% | |
| Stat | Value | P (rand ≥ data) | |||
| PhiPT | 0.05 | 0.11 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hozawa, M.; Nawata, E. Assessment of the Genetic Diversity of Ulex europaeus in Maui, California, Hawaii and New Zealand by a Method of Microsatellite Markers. Biol. Life Sci. Forum 2021, 4, 5. https://doi.org/10.3390/IECPS2020-08564
Hozawa M, Nawata E. Assessment of the Genetic Diversity of Ulex europaeus in Maui, California, Hawaii and New Zealand by a Method of Microsatellite Markers. Biology and Life Sciences Forum. 2021; 4(1):5. https://doi.org/10.3390/IECPS2020-08564
Chicago/Turabian StyleHozawa, Mika, and Eiji Nawata. 2021. "Assessment of the Genetic Diversity of Ulex europaeus in Maui, California, Hawaii and New Zealand by a Method of Microsatellite Markers" Biology and Life Sciences Forum 4, no. 1: 5. https://doi.org/10.3390/IECPS2020-08564
APA StyleHozawa, M., & Nawata, E. (2021). Assessment of the Genetic Diversity of Ulex europaeus in Maui, California, Hawaii and New Zealand by a Method of Microsatellite Markers. Biology and Life Sciences Forum, 4(1), 5. https://doi.org/10.3390/IECPS2020-08564