
Tomato Fruit Development in Response to Different Irrigation Practices: Developmental Study of Pericarp Cell Layers †


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Experiments
2.1. Plant Material, Growing Conditions, and Soil Water Content
2.2. Fruit Growth Parameter Measurements
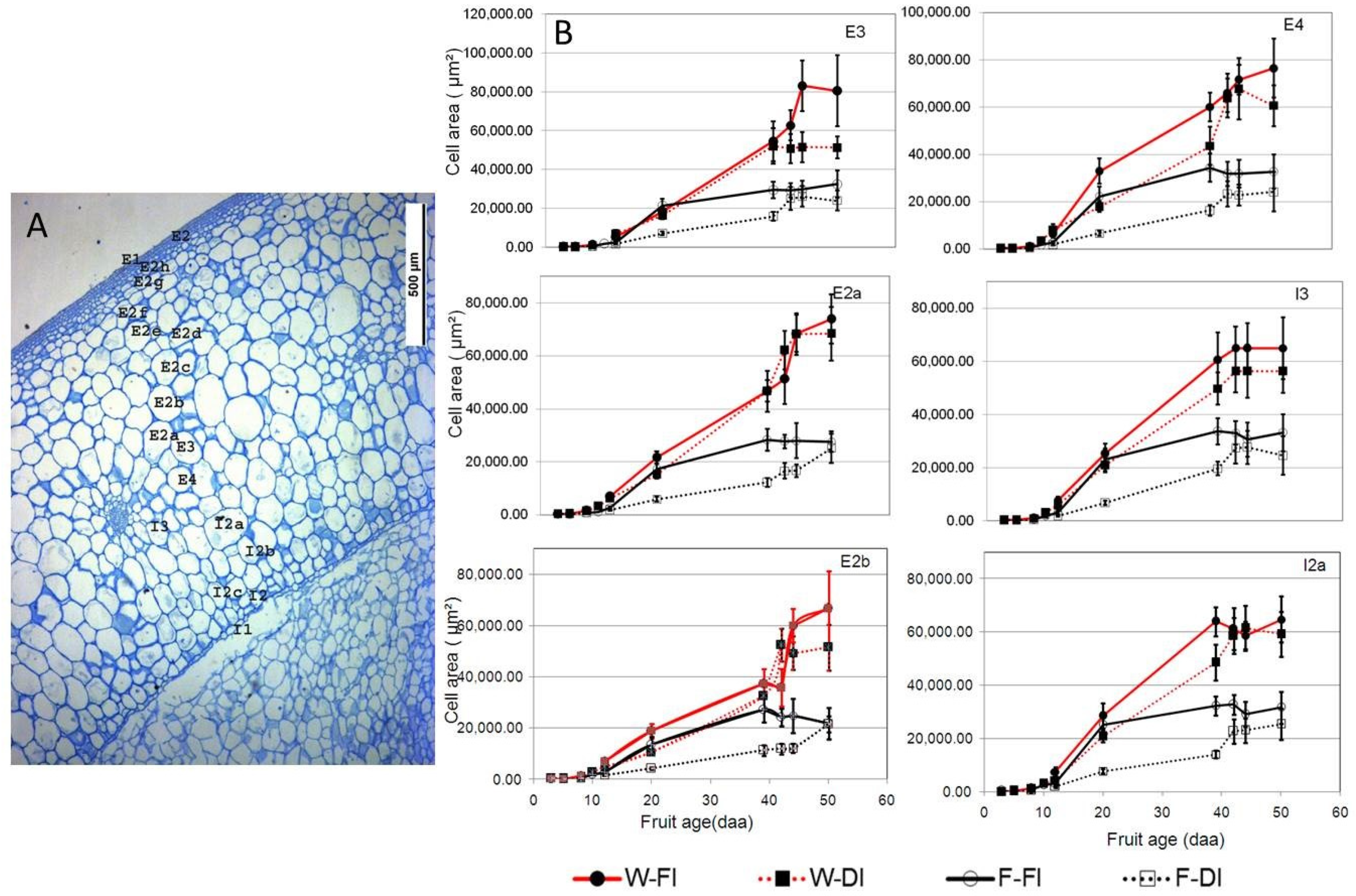
2.3. Histological Analyses
2.4. Statistical Analysis
3. Results
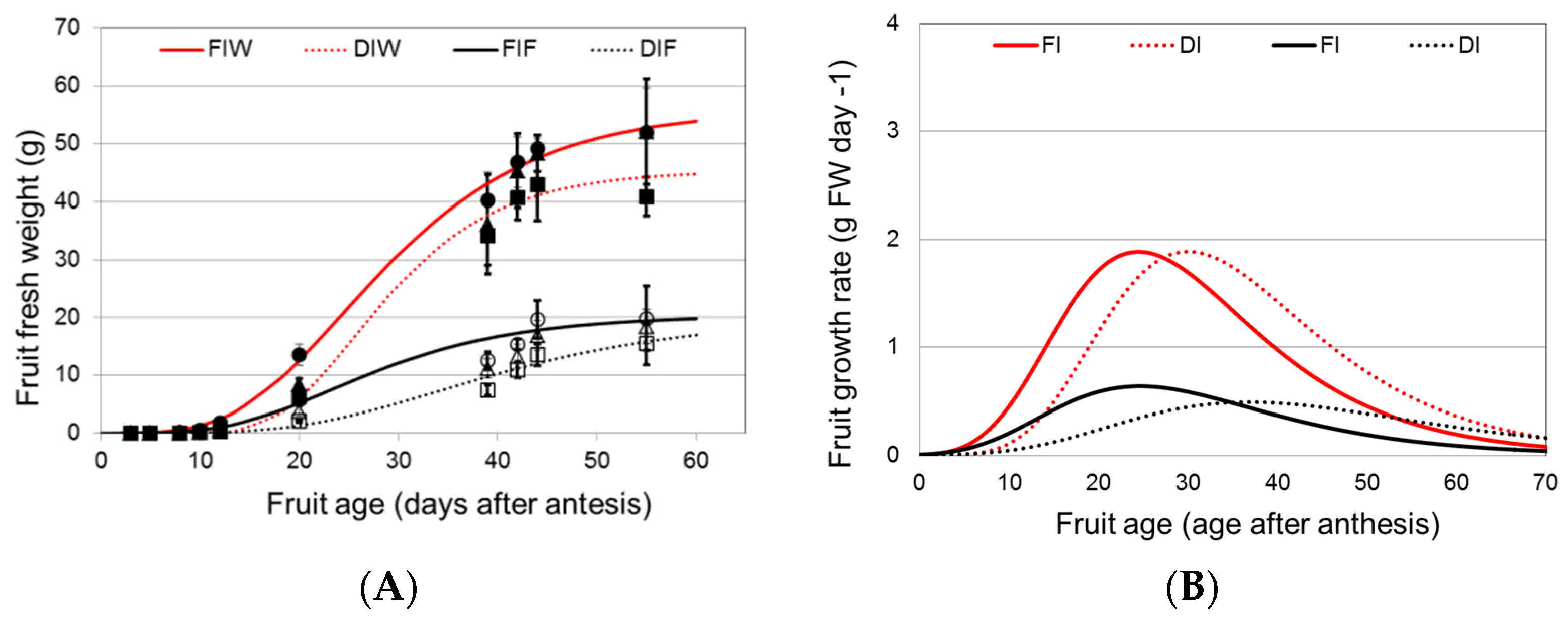
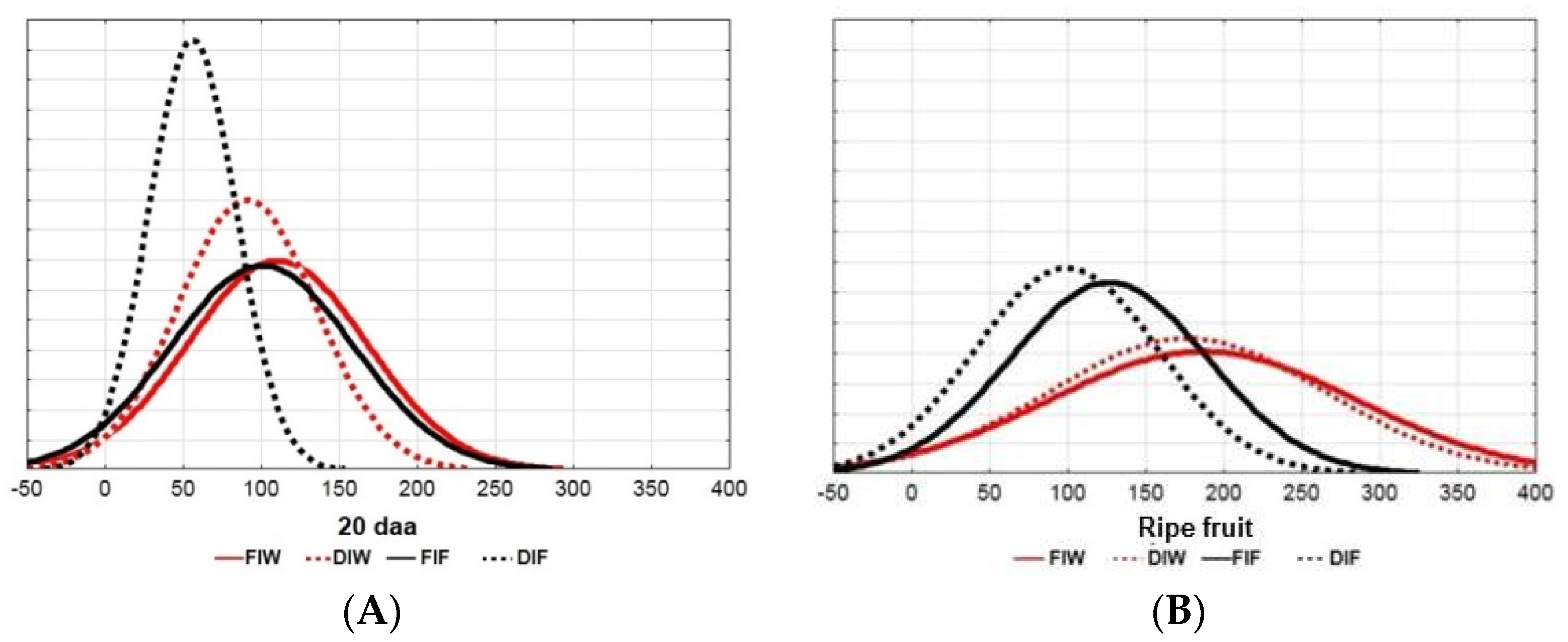
3.1. Effects of DI Treatment on Fruit Growth and Fresh Weight
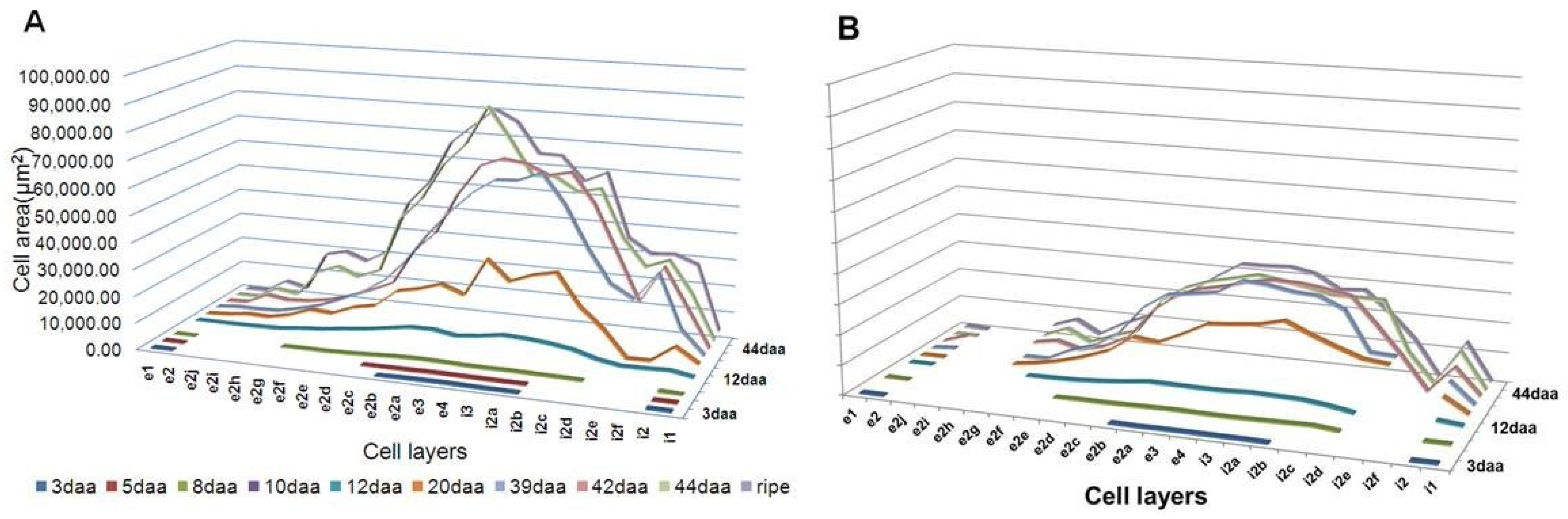
3.2. Effects of DI on the Pericarp Cell Number and Cell Area
3.3. Effects of DI on Cell Growth Kinetics of Different Pericarp Cell Layers
4. Discussion
4.1. Potential Roles of ABA in Fruit Growth in FI
4.2. Implication of Cellular Traits in the Fruit Response to DI
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Nuruddin, M.M.; Madramootoo, C.A.; Dodds, G.T. Effects of water stress at different growth stages on greenhouse tomato yield and quality. HortScience 2003, 38, 1389–1393. [Google Scholar] [CrossRef]
- Marsal, J.; Rapoport, H.F.; Manriqueb, T.; Girona, J. Pear fruit growth under regulated deficit irrigation in container-grown trees. Sci. Hortic. 2000, 85, 243–259. [Google Scholar] [CrossRef]
- Rapoport, H.F.; Costagli, G.; Gucci, R. The Effect of Water Deficit during Early Fruit Development on Olive Fruit Morphogenesis. J. Am. Soc. Hortic. Sci. 2004, 129, 121–127. [Google Scholar] [CrossRef]
- Dorji, K.; Behboudiana, M.H.; Zegbe-Domınguez, J.A. Water relations, growth, yield, and fruit quality of hot pepper under deficit irrigation and partial rootzone drying. Sci. Hortic. 2005, 104, 137–149. [Google Scholar] [CrossRef]
- Ruiz-Sanchez, M.C.; Domingo, R.; Castel, J.R. Deficit irrigation in fruit trees and vines in Spain. Span. J. Agric. Res. 2010, 8, 5–20. [Google Scholar] [CrossRef]
- Nangare, D.D.; Singh, Y.; Kumar, P.S.; Minhas, P.S. Growth, fruit yield and quality of tomato (Lycopersiconesculentum Mill.) as affected by deficit irrigation regulated on phenological basis. Agric. Water Manag. 2016, 171, 73–79. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Z.J.; Andersen, M.N.; Hu, T.T. Tomato yield and water use efficiency—Coupling effects between growth stage specific soil water deficits. Acta Agric. Scand. B Soil Plant Sci. 2015, 65, 460–469. [Google Scholar] [CrossRef]
- Gucci, R.; Lodolini, M.E.; Rapaport, F.H. Water deficit-induced changes in mesocarp cellular processes and the relationship between mesocarp and endocarp during olive fruit development. Tree Physiol. 2009, 29, 1575–1585. [Google Scholar] [CrossRef]
- Rosati, A.; Caporalia, S.; Hammami, S.B.M.; Moreno, A.I.; Paoletti, I.; Rapoport, H.F. Differences in ovary size among olive (Oleaeuropaea L.) cultivars are mainly related to cell number, not to cell size. Sci. Hortic. 2011, 130, 185–190. [Google Scholar] [CrossRef]
- Atta-Aly, M.A.; Riad, G.S.; Lacheene, Z.E.S.; El-Beltagy, A.S. Early ethrel applications extend tomato fruit cell division and increase fruit size and yield with ripening delay. J. Plant. Growth Regul. 1999, 18, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Bertin, N.; Gautier, H.; Roche, C. Number of cells in tomato fruit depending on fruit position and source-sink balance during plant development. J. Plant Growth Regul. 2002, 36, 105–112. [Google Scholar] [CrossRef]
- Ripoll, J.; Urban, L.; Brunel, B.; Bertin, N. Water deficit effects on tomato quality depend on fruit developmental stage and genotype. J. Plant Physiol. 2016, 190, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Bertin, N. Analysis of the Tomato Fruit Growth Response to Temperature and Plant Fruit Load in Relation to Cell Division, Cell Expansion and DNA Endoreduplication. Ann. Bot. 2005, 95, 439–447. [Google Scholar] [CrossRef]
- Cheniclet, C.; Rong, W.Y.; Causse, M.; Frangne, N.; Bolling, L.; Carde, J.P.; Renaudin, J.P. Cell Expansion and Endoreduplication Show a Large Genetic Variability in Pericarp and Contribute Strongly to Tomato Fruit Growth. Plant Physiol. 2005, 139, 1984–1994. [Google Scholar] [CrossRef] [PubMed]
- Fanwoua, J.; de Visser, P.; Heuvelink, E.; Angenent, G.; Yin, X.; Marcelis, L.; Struik, P. Response of Cell Division and Cell Expansion to Local Fruit Heating in Tomato Fruit. J. Am. Soc. Hortic. Sci. 2012, 137, 294–301. [Google Scholar] [CrossRef]
- Chen, G.; Fu, X.; Lips, S.H.; Sagi, M. Control of plant growth resides in the shoot, and not in the root, in reciprocal grafts of flacca and wild-type tomato (Lysopersiconesculentum), in the presence and absence of salinity stress. Plant Soil 2003, 256, 205–215. [Google Scholar] [CrossRef]
- Rancic, D.; Quarrie, S.P.; Pecinar, I. Anatomy of Tomato Fruit and Fruit Pedicel during Fruit Development. In Microscopy: Science, Technology, Applications and Education; Méndez-Vilas, A., Díaz, J., Eds.; Formatex Research Center: Badajoz, Spain, 2010; pp. 851–861. [Google Scholar]
- Tjørve, K.M.; Tjørve, E. The use of Gompertz models in growth analyses, and new Gompertz-model approach: An addition to the Unified-Richards family. PLoS ONE 2017, 12, e0178691. [Google Scholar] [CrossRef]
- Ruzin, S.E. Plant Microtechnique and Microscopy; Oxford University Press: Oxford, UK, 1999; p. 334. [Google Scholar]
- Rasband, W.S. (1997–2009) ImageJ, U.S. National Institutes of Health, Bethesda, Maryland, USA. Available online: http://rsb.info.nih.gov/ij/, (accessed on 14 June 2020).
- Nitsch, L.; Kohlenb, W.; Oplaata, C.; Charnikhova, T.; Cristescu, S.; Michieli, P.; Wolters-Arts, M.; Bouwmeester, H.; Mariani, C.; Vriezen, W.H.; et al. ABA-deficiency results in reduced plant and fruit size in tomato. J. Plant Physiol. 2012, 169, 878–883. [Google Scholar] [CrossRef] [PubMed]
- Kojima, K. Phytohormones in Shoots and Fruits of Tomato; Apoplast solution and seedless fruit. Jpn. Agric. Res. Q. 2005, 39, 77–81. [Google Scholar] [CrossRef][Green Version]
- Neill, S.J.; Horgan, R. Abscisic Acid Production and Water Relations in Wilty Tomato Mutants Subjected to Water Deficiency. J. Exp. Bot. 1985, 36, 1222–1231. [Google Scholar] [CrossRef]
- Davies, W.J.; Bacon, M.A.; Thompson, D.S.; Sobeigh, W.; Rodriguez, L.G. Regulation of leaf and fruit growth in plants in drying soil: Exploitation of the plant’s chemical signaling system and hydraulic architecture to increase the efficiency of water use in agriculture. J. Exp. Bot. 2000, 51, 1617–1626. [Google Scholar] [CrossRef]
- Rančić, D. Morpho-Anatomical Analysis of Tomato in Drought Condition. Ph.D. Thesis, Faculty of Agriculture, University of Belgrade, Beograd, Serbia, 24 July 2011. [Google Scholar]
- Zhang, L.Y.; Peng, Y.B.; Pelleschi-Travier, S.; Fan, Y.; Lu, Y.F.; Lu, Y.M.; Zhang, D.P. Evidence for Apoplasmic Phloem Unloading in Developing Apple Fruit. Plant Physiol. 2004, 135, 574–586. [Google Scholar] [CrossRef] [PubMed]
- Dodd, I.C. Leaf area development of ABA-deficient and wild-type peas at two levels of nitrogen supply. Funct. Plant Bio. 2003, 30, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Ripoll, J.; Urban, L.; Staudt, M.; Lopez-Lauri, F.; Bidel, L.P.R.; Bertin, N. Water shortage and quality of fleshy fruits—Making the most of the unavoidable. Rev. J. Exp. Bot. 2014, 65, 4097–4117. [Google Scholar] [CrossRef] [PubMed]
- Prudent, M.; Bertin, N.; Génard, M.; Munos, S.; Rolland, S.; Garcia, V.; Causse, M. Genotype-dependent response of growing tomato fruit to carbon availability. Plant Cell Environ. 2010, 33, 1186–1204. [Google Scholar] [CrossRef]
- Baldet, P.; Hernould, M.; Laporte, F.; Mounet, F.; Just, D.; Mouras, A.; Rothan, C. Expression of cell proliferation-related genes in early developing flower is affected by fruit load reduction in tomato plants. J. Exp Bot. 2006, 57, 961–970. [Google Scholar] [CrossRef]
- Ojeda, H.; Deloire, A.; Carbonneau, A. Influence of water deficit on grape berry growth. Vitis 2001, 40, 141–145. [Google Scholar] [CrossRef]
- McAtee, P.; Karim, S.; Schaffer, R.J.; David, K. A dynamic interplay between phytohormones is required for fruit development, maturation, and ripening. Front. Plant Sci. 2013, 4, 79. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pećinar, I.; Quarrie, S.P.; Bertin, N.; Rančić, D.; Savić, S.; Jovanović, Z.; Stikić, R. Tomato Fruit Development in Response to Different Irrigation Practices: Developmental Study of Pericarp Cell Layers. Biol. Life Sci. Forum 2021, 4, 105. https://doi.org/10.3390/IECPS2020-08855
Pećinar I, Quarrie SP, Bertin N, Rančić D, Savić S, Jovanović Z, Stikić R. Tomato Fruit Development in Response to Different Irrigation Practices: Developmental Study of Pericarp Cell Layers. Biology and Life Sciences Forum. 2021; 4(1):105. https://doi.org/10.3390/IECPS2020-08855
Chicago/Turabian StylePećinar, Ilinka, Sofija Pekić Quarrie, Nadia Bertin, Dragana Rančić, Slađana Savić, Zorica Jovanović, and Radmila Stikić. 2021. "Tomato Fruit Development in Response to Different Irrigation Practices: Developmental Study of Pericarp Cell Layers" Biology and Life Sciences Forum 4, no. 1: 105. https://doi.org/10.3390/IECPS2020-08855
APA StylePećinar, I., Quarrie, S. P., Bertin, N., Rančić, D., Savić, S., Jovanović, Z., & Stikić, R. (2021). Tomato Fruit Development in Response to Different Irrigation Practices: Developmental Study of Pericarp Cell Layers. Biology and Life Sciences Forum, 4(1), 105. https://doi.org/10.3390/IECPS2020-08855