Abstract
The purpose of this work is to present a three-species harvesting food web model that takes into account the interactions of susceptible prey, infected prey, and predator species. Prey species are assumed to expand logistically in the absence of predator species. The Crowley–Martin and Beddington–DeAngelis functional responses are used by predators to consume both susceptible and infected prey. Additionally, susceptible prey is consumed by infected prey in the formation of a Holling type II response. Both prey species are considered when prey harvesting is taken into account. Boundedness, positivity, and positive invariance are considered in this study. The investigation covers all the equilibrium points that are biologically feasible. Local stability is evaluated by analyzing the distribution of eigen values, while global stability is evaluated using suitable Lyapunov functions. Also, Hopf bifurcation is analyzed at the harvesting rate . At the end, we evaluate the numerical solutions based on our findings.
1. Introduction
In the natural environment, a variety of diseases may arise and spread among species when they interact with other organisms. Mathematical models have evolved into important tools for evaluating disease propagation and control. An eco-epidemiological model of diseased three-species food webs includes infectious prey, susceptible prey, and predators. At the beginning of the 20th century, several strategies were established in mathematical ecology to predict the presence of organisms and species of growth. The first significant attempt in this field was the well-known traditional Lotka–Volterra model [1] in 1927.
The investigation of predator–prey relationships is a crucial field of ecological research. The mathematical modeling of epidemics has become a prominent field of research. In this field, a substantial quantity of research has been performed [2,3,4]. Furthermore, mining and harvesting are practiced on a large number of the species found in the natural environment. Harvesting of the species is required for coexistence, and hence, the researchers were quite interested in the proposed ecological models. Different methods of harvesting have been proposed and explored, including constant harvesting, density-dependent proportional harvesting, and nonlinear harvesting [5,6]. By considering the above, in this work, we propose and study an eco-epidemiological prey–predator model involving different functional responses of harvesting. The majority of functional responses, like Holling types, are classified as “prey-dependent” because they depend on either the predator or the prey [7]. Both the prey and the predator are taken into account in Crowley–Martin reactions. In the Beddington–DeAngelis form, handling prey and hunting prey are viewed as two separate and independent actions. The response function of Beddington–DeAngelis, Holling type II, and Crowley–Martin forms are considered in this work. The main goal of this study is to analyze how disease and prey harvesting affect the predator–prey relationship. To the best of our knowledge, no studies have looked at an eco-epidemiological model of the three-species food web of harvesting with varying functional responses.
Section 2 addresses the mathematical expression. Some preliminary observations are presented in Section 3. The boundary equilibrium points and stability are shown in Section 4. In Section 5.1, the coexistence condition of the interior equilibrium point is determined by examining its local stability. The global stability analysis for is verified in Section 5.2. Furthermore, Section 6, investigates the Hopf bifurcation based on the harvesting rate . The MATLAB software tool (https://www.mathworks.com/products/matlab.html accessed on 1 March 2024) is used quantitatively to validate all the key results in Section 7. The conclusion of this research, as well as the environmental impacts of our results, are shown in Section 8, which ends our research.
2. Formation and Flowchart of the Equation
Prey harvesting is incorporated into the models for a predator–prey system.
by the positive conditions , and . The detailed environmental illustrations of the parameters are given in Table 1.

Table 1.
Ecological description of the model.
To minimize the parameters in model (1), we modify the variables as follows: The transformations can be utilized to formulate the Equation (1) in a dimensionless form (). Figure 1 displays the the model’s flowchart with the harvesting of various functional responses.
where . Now, the model’s conditions are , and .
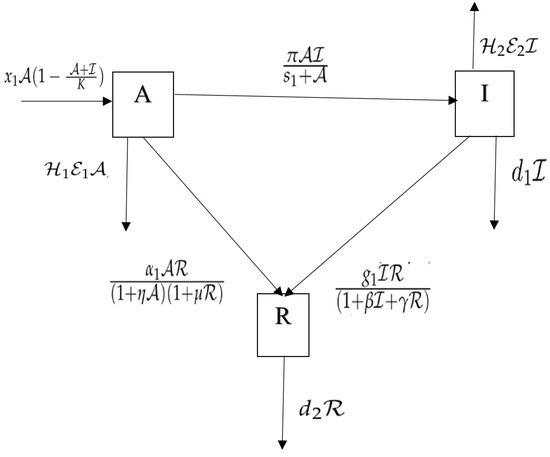
Figure 1.
Flowchart of the model with different functional responses.
3. Positivity and Boundedness
Let and , where
The Equation can be denoted as where with Thus, for . is continuous and a Lipschitzian function on . It contains non-negative conditions. So, the region is under an invariant condition.
Theorem 1.
The model’s (2) potential responses are bounded, and it is in .
4. Presence of Boundary Equilibrium Points
- is the equilibria of a trivial point. Here, (0, 0, 0) exists.
- is no infection and predator-free equilibria; exists for <
- is the equilibria of without a predator; where = and= . exists for and .
- is the no diseases of the equilibria; where = and= . exists for and .
- is the equilibria of the coexistent state; . It exists for , , , , where=, =, and=.
5. Stability Analysis
5.1. Local Analysis
The matrix of Jacobian equations is used to investigate the local stability at a point in , which is
Theorem 2.
The following are the points to verify the stability condition of model (2). They are as follows:
- 1.
- The trivial point of equilibria is LAS if .
- 2.
- The infectious and predator-free points are LAS if , , and .
- 3.
- The equilibria with no predator is LAS if , , and .
Proof.
- The trivial point of equilibria of the eigen values are , , and −δ. Hence, it is LAS when ; if not, it is unstable.
- The eigen values of are , , and . Hence, it is LAS if , , and ; if not, it is unstable.
- The matrix in its Jacobian form is
The characteristic form of is , where and . As a result, one of the eigenvalues of the equation is , i.e., negative. Hence, the other two must likewise be negative. So, is LAS if , , and . □
Theorem 3.
The infectious-free point of equilibrium is LAS if , , and .
(This demonstration is equivalent to Theorem 2, condition (3).)
Theorem 4.
The equilibrium point is LAS if , , and .
Proof.
, , and . The root of the characteristic equation is negative real parts if and only if , , and . According to Routh–Hurwitz, is LAS. □
5.2. Global Analysis
Theorem 5.
The point is GAS in and or and .
Proof.
A suitable Lyapunov function is expressed as
,
where are positive constant.
Differentiating with regard to t,
The region area and is negative:
and it shows that is a suitable Lyapunov function for all the solutions in W. □
6. Analysis of the Hopf Bifurcation
Theorem 6.
If the bifurcating parameter exceeds a substantial value, then Hopf bifurcation occurs in the system (2). The presence of the Hopf bifurcation requirements listed below is =
- 1.
- 2.
- where γ represents the positive value of the equilibrium point and is the zero of the characteristic equation.
Proof.
For , let (3) denote
(i.e) and are the roots of the Equation (6). To establish that the Hopf bifurcation exists at the point, we must fulfill the transversality requirement. . For all , the roots of the form and Now, we check the condition Let = in (6), and we obtain where
, and . To complete the Equation (6), we need and and then we differentiate and with respect to . Because
and at on and . So, ,
(i.e) and .
Thus, the condition . It has been confirmed that the transversality criteria apply to system (2), and the Hopf bifurcation occurs at □
7. Numerical Calculations of the Model
To verify the theoretical conclusions, this part performs a calculation on system (2). Here, the harvesting rate is employed as an adjustable element. The simulation is accomplished by utilizing MATLAB software tools for the fixed parameter. Here, x = 0.2, = 0.1, d = 0.2, = 0.21, = (variable), = 0.13, = 0.3, and = 0.11. If is 0.21, when bifurcation occurs, the model (2) for the non-negative equilibrium is LAS (0.52764, 0.0916818, and 0.203662) and the rest of the adjustable elements have identical values. The model’s (2) stability is lost by increasing the bifurcation adjustable element to = 0.47, leading to being LAU at (0.53824, 0.0917748, and 0.320178). Model (2) is able to pass the transversality conditions for . Hence, Figure 2 displays how the model’s behavior changes at a harvesting rate of = 0.47.
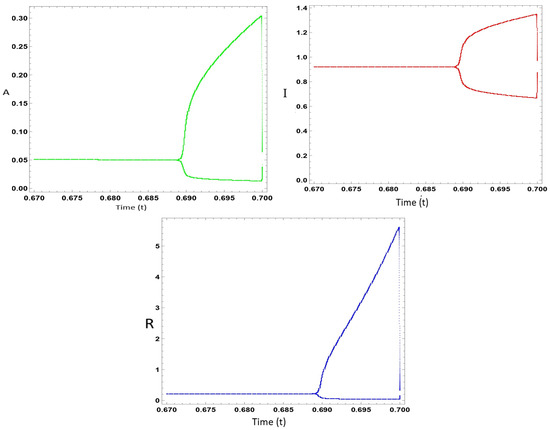
Figure 2.
Dynamical changes in Model (2) at harvesting rate = 0.47.
8. Conclusions and Discussion
Our investigation involved examining an eco-epidemiological model where sick prey are harvested from the prey species, and the predator eats both sick and healthy prey. The developed system (2) has been shown to be biologically well behaved by the boundedness and positivity results. In the event that if the growing rate of uninfected prey is lower than the harvest rate, then the population tends to be extinct. It has been demonstrated that both the local stability at every ecologically possible point and the coexistence (2) are stable. The analytical and numerical outcomes of the Hopf bifurcation for the harvesting rate have been analyzed and evaluated in the above. The dynamic of prey harvesting is powerful due to the complex behavior demonstrated in this study. Thus, we believe that ordinary differential equations will be utilized to solve many future technological equations.
Author Contributions
Conceptualization, T.M. and T.N.G.; methodology, M.S.P.; software, T.M.; validation, A.Y., M.S.P. and T.N.G.; formal analysis, M.S.P.; investigation, T.M.; resources, T.N.G.; data curation, T.M.; writing—original draft preparation, T.M.; writing—review and editing, T.M.; visualization, M.S.P.; supervision, T.N.G.; project administration, A.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data in this article are available from the corresponding author upon request.
Acknowledgments
Authors extend their appreciation to Sri Ramakrishna Mission Vidyalaya College of Arts and Science for providing all research facilities. Atlast, the authors would like to thank anonymous reviewers, organizers and sponsors of the conference.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| LAS | Locally Asymptotically Stable |
| GAS | Globally Asymptotically Stable |
| LAU | Locally Asymptotically Unstable |
References
- May, R.M. Stability and Complexity in Model Ecosystems; Princeton University Press: Princeton, NJ, USA, 2019; Volume 1. [Google Scholar]
- Kant, S.; Kumar, V. Dynamics of a Prey-Predator System with Infection in Prey; Department of Mathematics, Texas State University: San Marcos, TX, USA, 2017. [Google Scholar]
- Pradeep, M.S.; Gopal, T.N.; Magudeeswaran, S.; Deepak, N.; Muthukumar, S. Stability analysis of diseased preadator–prey model with holling type II functional response. AIP Conf. Proc. 2023, 2901, 030017. [Google Scholar]
- Thangavel, M.; Thangaraj, N.G.; Manickasundaram, S.P.; Arunachalam, Y. An Eco–Epidemiological Model Involving Prey Refuge and Prey Harvesting with Beddington–DeAngelis, Crowley–Martin and Holling Type II Functional Responses. Eng. Proc. 2023, 56, 325. [Google Scholar] [CrossRef]
- Abdulghafour, A.S.; Naji, R.K. The impact of refuge and harvesting on the dynamics of prey-predator system. Sci. Int. 2018, 30, 315–323. [Google Scholar]
- Agnihotri, K.B.; Gakkhar, S. The dynamics of disease transmission in a prey predator system with harvesting of prey. Int. J. Adv. Res. Comput. Eng. Technol. 2012, 1, 1–17. [Google Scholar]
- Magudeeswaran, S.; Vinoth, S.; Sathiyanathan, K.; Sivabalan, M. Impact of fear on delayed three species food-web model with Holling type-II functional response. Int. J. Biomath. 2022, 15, 2250014. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).