
Gene Expression Profile of Neutrophil Extracellular Traps (NETs) Stimulated by L-Amino Acid Oxidase from Calloselasma rhodostoma Venom †

 , ,
, , 

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Chemicals and Reagents
2.2. Isolation and Biochemical Characterization of Cr-LAAO
2.3. Isolation and Activation of Neutrophils
2.4. Gene Expression Profile
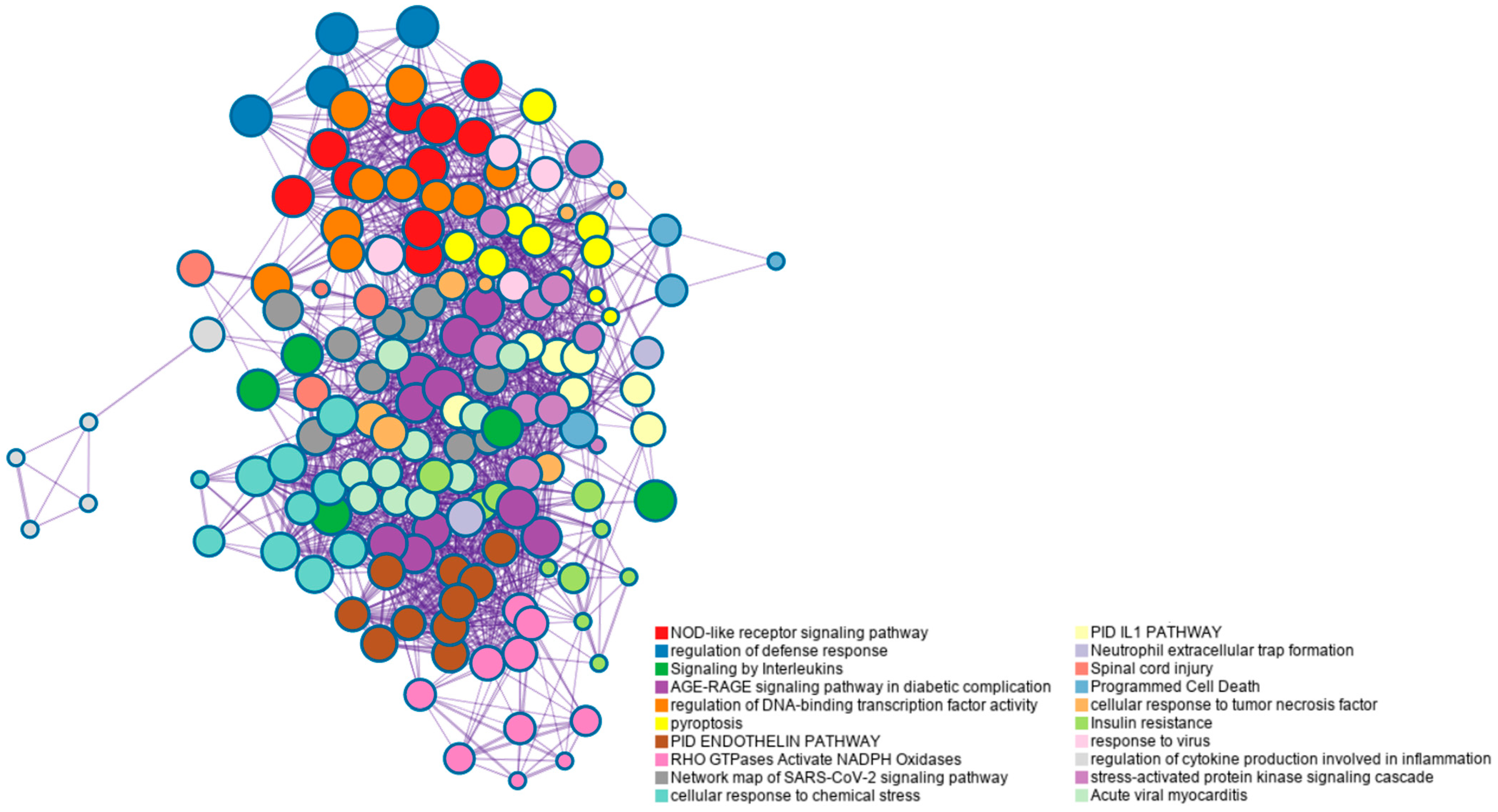
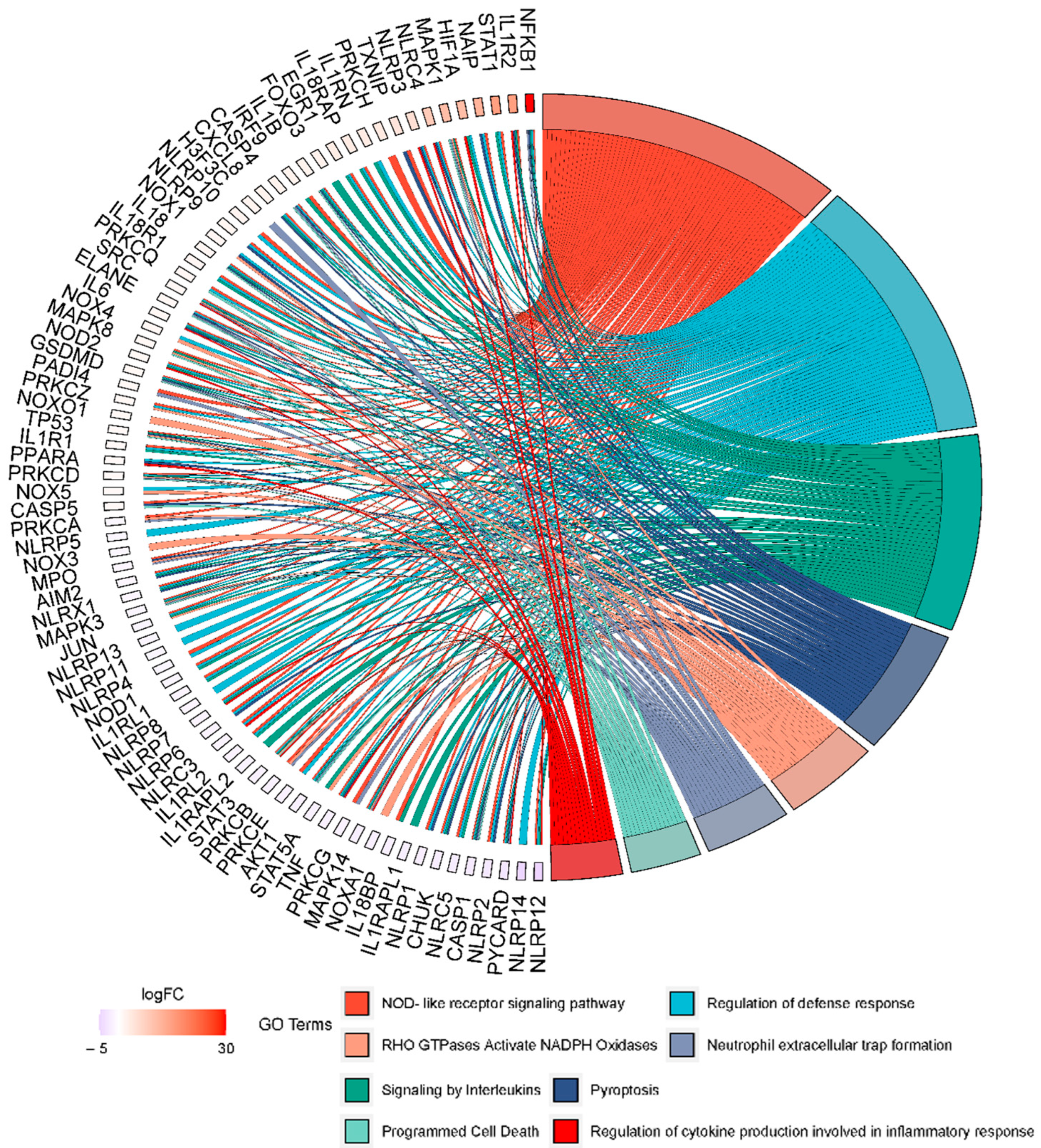
3. Results and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Du, X.-Y.; Clemetson, K.J. Snake Venom L-Amino Acid Oxidases. Toxicon 2002, 40, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Izidoro, L.F.M.; Sobrinho, J.C.; Mendes, M.M.; Costa, T.R.; Grabner, A.N.; Rodrigues, V.M.; da Silva, S.L.; Zanchi, F.B.; Zuliani, J.P.; Fernandes, C.F.C.; et al. Snake Venom L-Amino Acid Oxidases: Trends in Pharmacology and Biochemistry. Biomed Res. Int. 2014, 2014, 196754. [Google Scholar] [CrossRef]
- Paloschi, M.V.; Pontes, A.S.; Soares, A.M.; Zuliani, J.P. An Update on Potential Molecular Mechanisms Underlying the Actions of Snake Venom L-Amino Acid Oxidases (LAAOs). Curr. Med. Chem. 2018, 25, 2520–2530. [Google Scholar] [CrossRef] [PubMed]
- Zuliani, J.P.; Paloschi, M.V.; Pontes, A.S.; Boeno, C.N.; Lopes, J.A.; Setubal, S.S.; Zanchi, F.B.; Soares, A.M. Reptile Venom L-Amino Acid Oxidases—Structure and Function. Handb. Venoms Toxins Reptil. 2021, 2, 413–430. [Google Scholar] [CrossRef]
- Rodrigues, R.S.; da Silva, J.F.; Boldrini França, J.; Fonseca, F.P.P.; Otaviano, A.R.; Henrique Silva, F.; Hamaguchi, A.; Magro, A.J.; Braz, A.S.K.; dos Santos, J.I.; et al. Structural and Functional Properties of Bp-LAAO, a New L-Amino Acid Oxidase Isolated from Bothrops Pauloensis Snake Venom. Biochimie 2009, 91, 490–501. [Google Scholar] [CrossRef]
- Pontes, A.S.; Setúbal, S.d.S.; Xavier, C.V.; Lacouth-Silva, F.; Kayano, A.M.; Pires, W.L.; Nery, N.M.; Boeri de Castro, O.; da Silva, S.D.; Calderon, L.A.; et al. Effect of L-Amino Acid Oxidase from Calloselasma Rhodosthoma Snake Venom on Human Neutrophils. Toxicon 2014, 80, 27–37. [Google Scholar] [CrossRef]
- Pontes, A.S.; Setúbal, S.d.S.; Nery, N.M.; da Silva, F.S.; da Silva, S.D.; Fernandes, C.F.C.; Stábeli, R.G.; Soares, A.M.; Zuliani, J.P. P38 MAPK Is Involved in Human Neutrophil Chemotaxis Induced by L-Amino Acid Oxidase from Calloselasma Rhodosthoma. Toxicon 2016, 119, 106–116. [Google Scholar] [CrossRef]
- Paloschi, M.V.; Boeno, C.N.; Lopes, J.A.; Eduardo dos Santos da Rosa, A.; Pires, W.L.; Pontes, A.S.; da Silva Setúbal, S.; Soares, A.M.; Zuliani, J.P. Role of L-Amino Acid Oxidase Isolated from Calloselasma Rhodostoma Venom on Neutrophil NADPH Oxidase Complex Activation. Toxicon 2018, 145, 48–55. [Google Scholar] [CrossRef]
- Mortaz, E.; Alipoor, S.D.; Adcock, I.M.; Mumby, S.; Koenderman, L. Update on Neutrophil Function in Severe Inflammation. Front. Immunol. 2018, 9, 2171. [Google Scholar] [CrossRef]
- Singh, P.; Kumar, N.; Singh, M.; Kaur, M.; Singh, G.; Narang, A.; Kanwal, A.; Sharma, K.; Singh, B.; Napoli, M.D.; et al. Neutrophil Extracellular Traps and NLRP3 Inflammasome: A Disturbing Duo in Atherosclerosis, Inflammation and Atherothrombosis. Vaccines 2023, 11, 261. [Google Scholar] [CrossRef]
- Pires, R.H.; Felix, S.B.; Delcea, M. The Architecture of Neutrophil Extracellular Traps Investigated by Atomic Force Microscopy. Nanoscale 2016, 8, 14193–14202. [Google Scholar] [CrossRef] [PubMed]
- Thiam, H.R.; Wong, S.L.; Wagner, D.D.; Waterman, C.M. Cellular Mechanisms of NETosis. Annu. Rev. Cell Dev. Biol. 2020, 36, 191–218. [Google Scholar] [CrossRef] [PubMed]
- Paloschi, M.V.; Lopes, J.A.; Boeno, C.N.; Silva, M.D.S.; Evangelista, J.R.; Pontes, A.S.; da Silva Setúbal, S.; Rego, C.M.A.; Néry, N.M.; Ferreira e Ferreira, A.A.; et al. Cytosolic Phospholipase A2-α Participates in Lipid Body Formation and PGE2 Release in Human Neutrophils Stimulated with an l-Amino Acid Oxidase from Calloselasma Rhodostoma Venom. Sci. Rep. 2020, 10, 10976. [Google Scholar] [CrossRef] [PubMed]
- Paloschi, M.V.; Boeno, C.N.; Lopes, J.A.; Rego, C.M.A.; Silva, M.D.S.; Santana, H.M.; Serrath, S.N.; Ikenohuchi, Y.J.; Farias, B.J.C.; Felipin, K.P.; et al. Reactive Oxygen Species-Dependent-NLRP3 Inflammasome Activation in Human Neutrophils Induced by l-Amino Acid Oxidase Derived from Calloselasma Rhodostoma Venom. Life Sci. 2022, 308, 120962. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape Provides a Biologist-Oriented Resource for the Analysis of Systems-Level Datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice Hall: Hoboken, NJ, USA, 1999. [Google Scholar]
- Hochberg, Y.; Benjamini, Y. More Powerful Procedures for Multiple Significance Testing. Stat. Med. 1990, 9, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J. A Coefficient of Agreement for Nominal Scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- Luo, W.; Friedman, M.S.; Shedden, K.; Hankenson, K.D.; Woolf, P.J. GAGE: Generally Applicable Gene Set Enrichment for Pathway Analysis. BMC Bioinform. 2009, 10, 161. [Google Scholar] [CrossRef]
- Luo, W.; Brouwer, C. Pathview: An R/Bioconductor Package for Pathway-Based Data Integration and Visualization. Bioinformatics 2013, 29, 1830–1831. [Google Scholar] [CrossRef]
- Luo, W.; Pant, G.; Bhavnasi, Y.K.; Blanchard, S.G.; Brouwer, C. Pathview Web: User Friendly Pathway Visualization and Data Integration. Nucleic Acids Res. 2017, 45, W501–W508. [Google Scholar] [CrossRef]
- Lamkanfi, M.; Dixit, V.M. Mechanisms and Functions of Inflammasomes. Cell 2014, 157, 1013–1022. [Google Scholar] [CrossRef]
- Ouyang, W.; Rutz, S.; Crellin, N.K.; Valdez, P.A.; Hymowitz, S.G. Regulation and Functions of the IL-10 Family of Cytokines in Inflammation and Disease. Annu. Rev. Immunol. 2011, 29, 71–109. [Google Scholar] [CrossRef]
- Bedard, K.; Krause, K.-H. The NOX Family of ROS-Generating NADPH Oxidases: Physiology and Pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef] [PubMed]
- Kenny, E.F.; Herzig, A.; Krüger, R.; Muth, A.; Mondal, S.; Thompson, P.R.; Brinkmann, V.; von Bernuth, H.; Zychlinsky, A. Diverse Stimuli Engage Different Neutrophil Extracellular Trap Pathways. Elife 2017, 6, e24437. [Google Scholar] [CrossRef]
- Pilsczek, F.H.; Salina, D.; Poon, K.K.H.; Fahey, C.; Yipp, B.G.; Sibley, C.D.; Robbins, S.M.; Green, F.H.Y.; Surette, M.G.; Sugai, M.; et al. A Novel Mechanism of Rapid Nuclear Neutrophil Extracellular Trap Formation in Response to Staphylococcus Aureus. J. Immunol. 2010, 185, 7413–7425. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paloschi, M.V.; Serrath, S.N.; Silva, M.D.S.; Cardozo, D.G.; Ugarte, A.V.E.; Boeno, C.N.; Lopes, J.A.; Rego, C.M.A.; Santana, H.M.; Ikenohuchi, Y.J.; et al. Gene Expression Profile of Neutrophil Extracellular Traps (NETs) Stimulated by L-Amino Acid Oxidase from Calloselasma rhodostoma Venom. Biol. Life Sci. Forum 2023, 24, 10. https://doi.org/10.3390/IECT2023-14811
Paloschi MV, Serrath SN, Silva MDS, Cardozo DG, Ugarte AVE, Boeno CN, Lopes JA, Rego CMA, Santana HM, Ikenohuchi YJ, et al. Gene Expression Profile of Neutrophil Extracellular Traps (NETs) Stimulated by L-Amino Acid Oxidase from Calloselasma rhodostoma Venom. Biology and Life Sciences Forum. 2023; 24(1):10. https://doi.org/10.3390/IECT2023-14811
Chicago/Turabian StylePaloschi, Mauro Valentino, Suzanne Nery Serrath, Milena Daniela Souza Silva, Daniel Gomes Cardozo, Andrey Victor Echeverria Ugarte, Charles Nunes Boeno, Jéssica Amaral Lopes, Cristina Matiele Alves Rego, Hallison Mota Santana, Yoda Janaina Ikenohuchi, and et al. 2023. "Gene Expression Profile of Neutrophil Extracellular Traps (NETs) Stimulated by L-Amino Acid Oxidase from Calloselasma rhodostoma Venom" Biology and Life Sciences Forum 24, no. 1: 10. https://doi.org/10.3390/IECT2023-14811
APA StylePaloschi, M. V., Serrath, S. N., Silva, M. D. S., Cardozo, D. G., Ugarte, A. V. E., Boeno, C. N., Lopes, J. A., Rego, C. M. A., Santana, H. M., Ikenohuchi, Y. J., da Silva, C. P., Farias, B. J. C., Felipin, K. P., Eulálio, M. d. M. C., Cruz, L. F., Magalhães, J. G. d. S., Setúbal, S. d. S., Soares, A. M., & Zuliani, J. P. (2023). Gene Expression Profile of Neutrophil Extracellular Traps (NETs) Stimulated by L-Amino Acid Oxidase from Calloselasma rhodostoma Venom. Biology and Life Sciences Forum, 24(1), 10. https://doi.org/10.3390/IECT2023-14811