
Inoculum Production of Monascus purpureus with Chenopodium quinoa in Submerged Culture †

 ,
,  , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain of Interest
2.2. Evaluation of the Parameters rpm, pH and Sodium Chloride Concentration
2.3. Quantification of N-Acetylglucosamine
2.4. Statistical Analysis
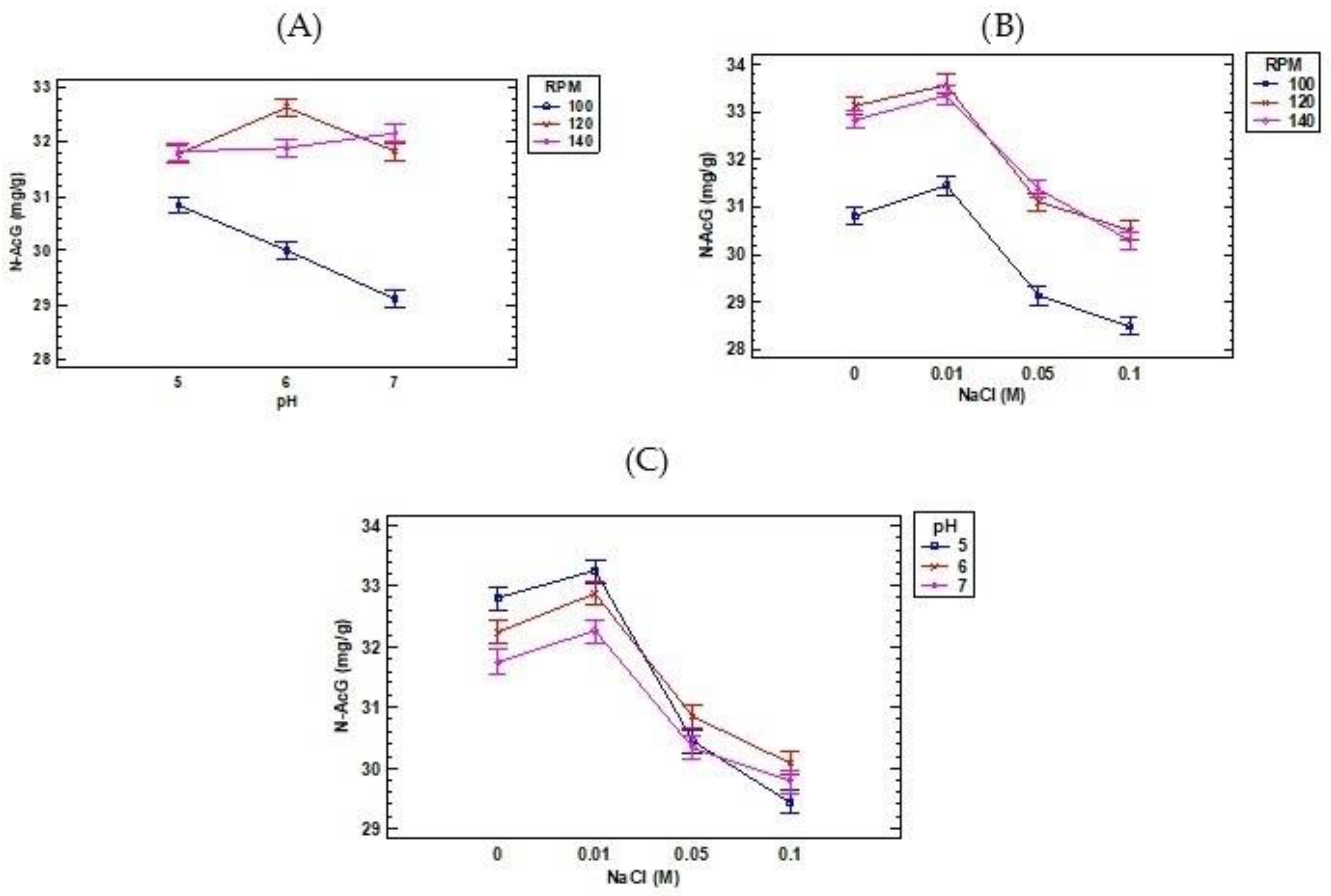
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davoudi, H.M.; Shahidi, F.; Tabatabaei, F.Y.; Sarabi, M.J.; Eshaghi, Z. Biological detoxification of Monascus purpureus pigments by heat-treated Saccharomyces cerevisiae. J. Sci. Food Agric. 2019, 99, 4439–4444. [Google Scholar] [CrossRef] [PubMed]
- Saithong, P.; Chitisankul, W.T.; Nitipan, S. Comparative study of red yeast rice with high monacolin K, low citrinin concentration and pigments in white rice and brown rice. Czech J. Food Sci. 2019, 37, 75–80. [Google Scholar] [CrossRef]
- Zhen, Z.; Xiong, X.; Liu, Y.; Zhang, J.; Wang, S.; Li, L.; Gao, M. NaCl inhibits citrinin and stimulates Monascus pigments and monacolin K production. Toxins. 2019, 11, 118. [Google Scholar] [CrossRef] [PubMed]
- Arzapalo, D.Q.; Huamán, C.K.; Quispe, S.M.; Espinoza, S.C. Extraction and characterization of the starch of three varieties of quinoa (Chenopodium quinoa Willd) black collana, pasakalla red and white junin. J. Chem. Soc. Peru 2015, 81, 44–54. [Google Scholar]
- Pellegrini, M.; Lucas, G.R.; Ricci, A.; Fontecha, J.; Fernández, L.J.; Pérez, J.A.; Viuda, M.M. Chemical, fatty acid, polyphenolic profile, techno-functional and antioxidant properties of flours obtained from quinoa (Chenopodium quinoa Willd) seeds. Ind. Crops Prod. 2018, 111, 38–46. [Google Scholar] [CrossRef]
- Ma, L.; Lin, Y.Q.; Yang, C.; Ying, Z.H.; Jiang, X.L. Production of liquid seed of an edible fungus, Sparassis latifolia by submerged fermentation and mycelium growth in pine wood sawdust. Sci. Hortic. 2016, 209, 22–30. [Google Scholar] [CrossRef]


{kind=link}
| Source | Sum of Squares | Df | Mean Square | F-Ratio | p-Value |
|---|---|---|---|---|---|
| main effects | |||||
| A:rpm | 100.82 | 2 | 50.41 | 855.15 | 0.0000 |
| B:pH | 5.11 | 2 | 2.56 | 43.42 | 0.0000 |
| C:NaCl (M) | 164.28 | 3 | 54.76 | 928.97 | 0.0000 |
| interactions | |||||
| AB | 19.52 | 4 | 4.88 | 82.82 | 0.0000 |
| AC | 1.12 | 6 | 0.186 | 3.17 | 0.0081 |
| BC | 7.45 | 6 | 1.240 | 21.05 | 0.0000 |
| ABC | 12.03 | 12 | 1.003 | 17.02 | 0.0000 |
| residual | 4.24 | 72 | 0.059 | ||
| total (corrected) | 314.60 | 107 |
| RPM | Count | LS Mean | LS Sigma | Homogeneous Groups |
|---|---|---|---|---|
| 100 | 36 | 29.9755 | 0.0404651 | X |
| 140 | 36 | 31.964 | 0.0404651 | X |
| 120 | 36 | 32.081 | 0.0404651 | X |
| pH | Count | LS Mean | LS Sigma | Homogeneous Groups |
|---|---|---|---|---|
| 7 | 36 | 31.0329 | 0.0404651 | X |
| 5 | 36 | 31.4769 | 0.0404651 | X |
| 6 | 36 | 31.5107 | 0.0404651 | X |
| NaCl (M) | Count | LS Mean | LS Sigma | Homogeneous Groups |
|---|---|---|---|---|
| 0.10 | 27 | 29.7666 | 0.046725 | X |
| 0.05 | 27 | 30.5383 | 0.046725 | X |
| 0.0 | 27 | 32.2596 | 0.046725 | X |
| 0.01 | 27 | 32.7961 | 0.046725 | X |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tucta-Huillca, F.; Quispe-Rivera, E.; Cadavez, V.; Gonzales-Barron, U.; Silva-Jaimes, M.; Morales, J.J. Inoculum Production of Monascus purpureus with Chenopodium quinoa in Submerged Culture. Biol. Life Sci. Forum 2022, 18, 62. https://doi.org/10.3390/Foods2022-12995
Tucta-Huillca F, Quispe-Rivera E, Cadavez V, Gonzales-Barron U, Silva-Jaimes M, Morales JJ. Inoculum Production of Monascus purpureus with Chenopodium quinoa in Submerged Culture. Biology and Life Sciences Forum. 2022; 18(1):62. https://doi.org/10.3390/Foods2022-12995
Chicago/Turabian StyleTucta-Huillca, Franz, Evelyn Quispe-Rivera, Vasco Cadavez, Ursula Gonzales-Barron, Marcial Silva-Jaimes, and Juan Juscamaita Morales. 2022. "Inoculum Production of Monascus purpureus with Chenopodium quinoa in Submerged Culture" Biology and Life Sciences Forum 18, no. 1: 62. https://doi.org/10.3390/Foods2022-12995
APA StyleTucta-Huillca, F., Quispe-Rivera, E., Cadavez, V., Gonzales-Barron, U., Silva-Jaimes, M., & Morales, J. J. (2022). Inoculum Production of Monascus purpureus with Chenopodium quinoa in Submerged Culture. Biology and Life Sciences Forum, 18(1), 62. https://doi.org/10.3390/Foods2022-12995