
A Co-Infection Model for Onchocerciasis and Lassa Fever with Optimal Control Analysis



Abstract
1. Introduction
2. OLF Co-Infection Model
- Every person is born with the ability to catch Lassa fever and onchocerciasis, implying that humans are at risk of contracting the diseases.
- Once susceptible people become sick, they transform into exposed people with immunity but are not yet contagious.
- Only individuals who are exposed to the virus become contagious.
- Infectious individuals can die naturally or as a consequence of the disease, and if they do otherwise, they can recover as a result of treatment.
- That person could simultaneously contract both Lassa fever and onchocerciasis.
- All rodents including black flies have vulnerability during birth.
- Each type of rodent species may perish spontaneously or as a result of hunting and the application of pesticides.
- Infected sensitive rodents are exposed but not contagious rodents.
- Only exposed rodents and black flies become infected.
- Infected rodents become infected when they consume or consume fluids from ill rodents.
- Afflicted rodents and black flies are infectious for life, implying that there is no recovered class for rodent and black fly populations.
Basic Model Features Model (1) without Controls
3. Analysis of Model (1) with Non-Controls
3.1. Disease-Free Equilibrium
3.2. Basic Reproduction Number, and
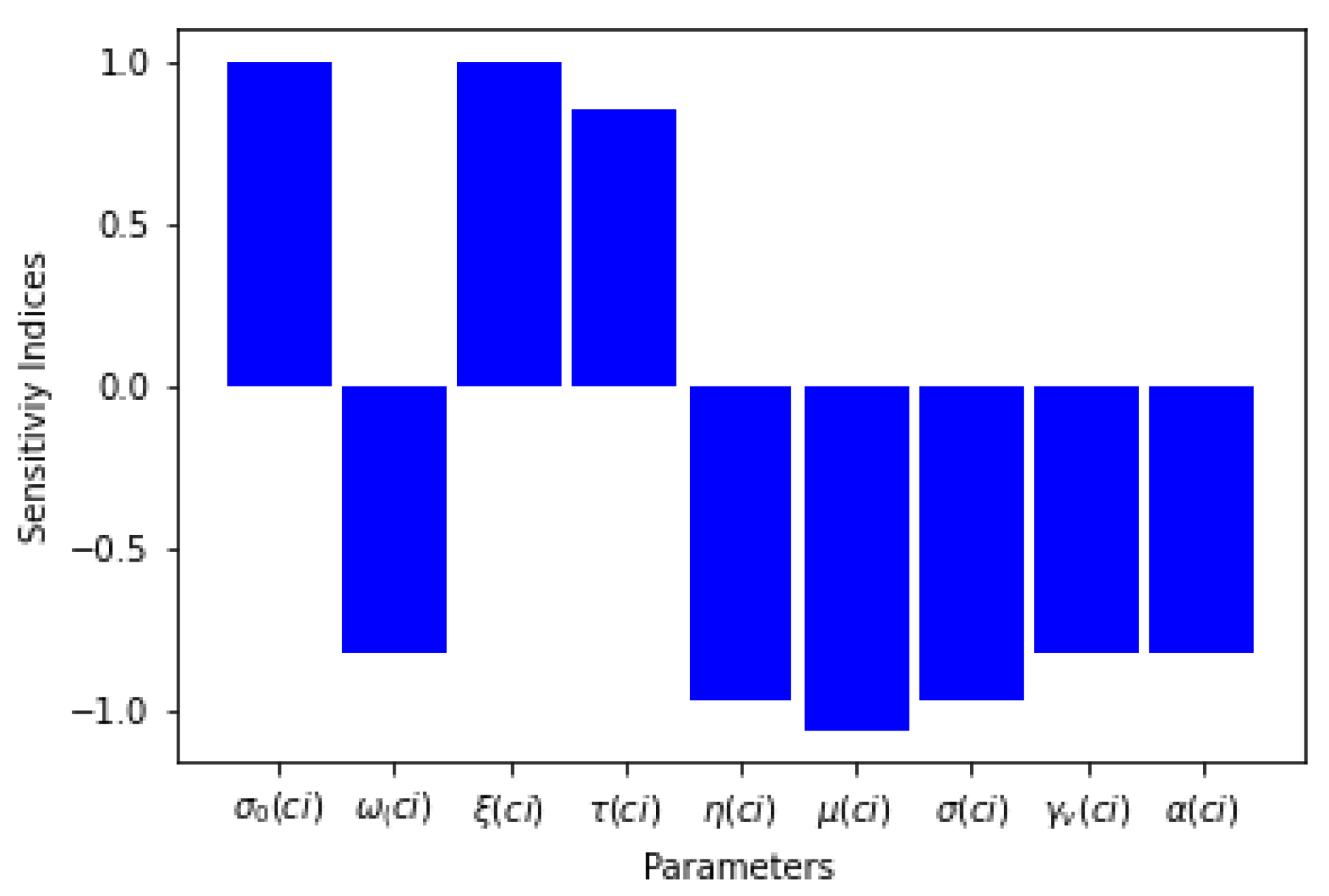
3.2.1. Analysis of the ’s Sensitivity,
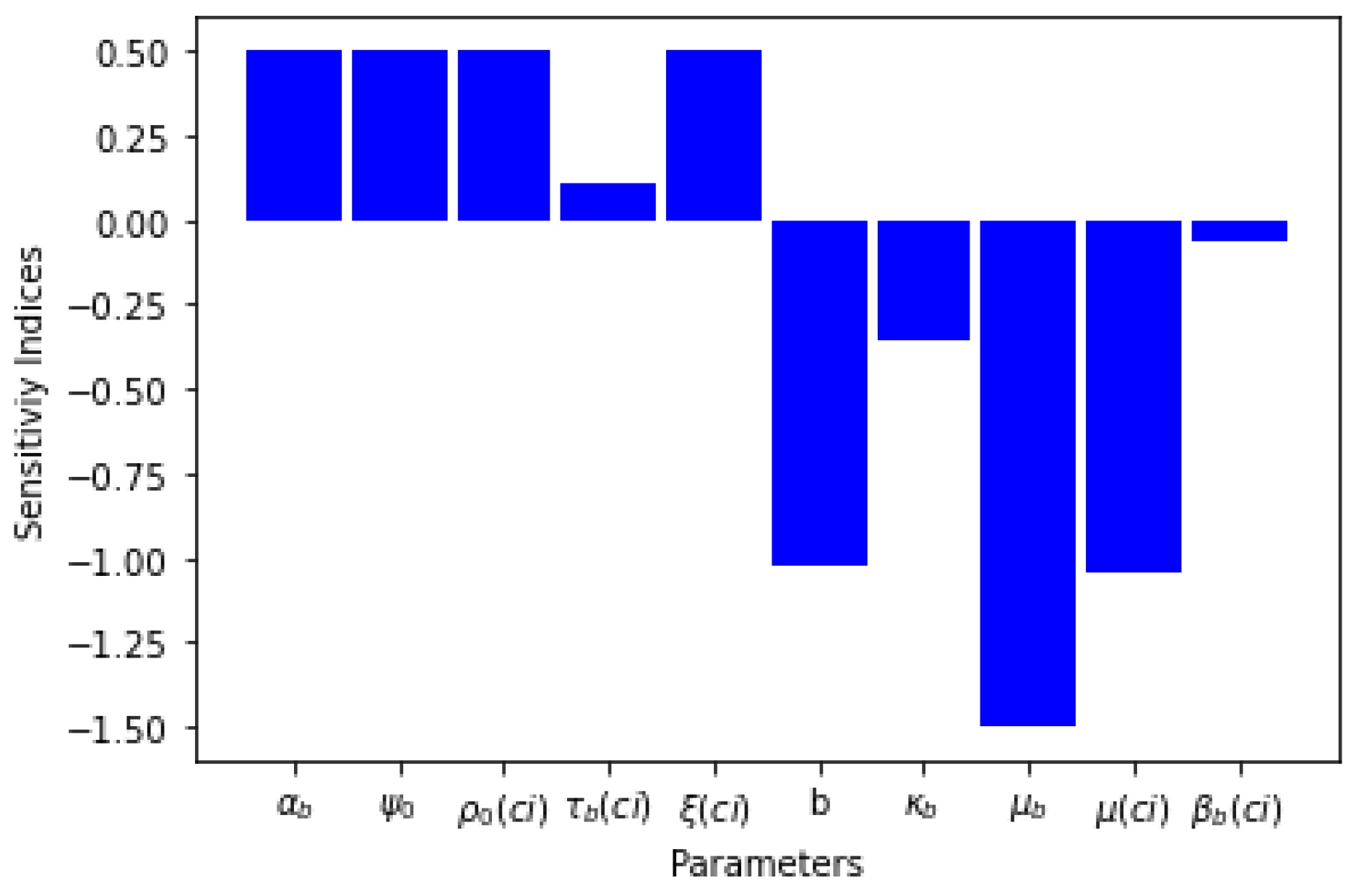
3.2.2. Sensitivity Analysis of Basic Reproduction Number,
3.3. Existence of Endemic Equilibrium
4. OLF Co-Infection Model Optimal Control
4.1. Global Stability Analysis
4.2. Invasion and Co-Existence
4.3. Analysis of Optimal Control
4.4. Existence of an Optimal Control
- (i)
- The set of controls and the corresponding state variables are non-empty;
- (ii)
- The control set is convex and closed;
- (iii)
- The right-hand side of the state system is bounded by a linear function in the state and control;
- (iv)
- The integrand of the functional is convex on and is bounded below by where and .
4.5. The Optimal System
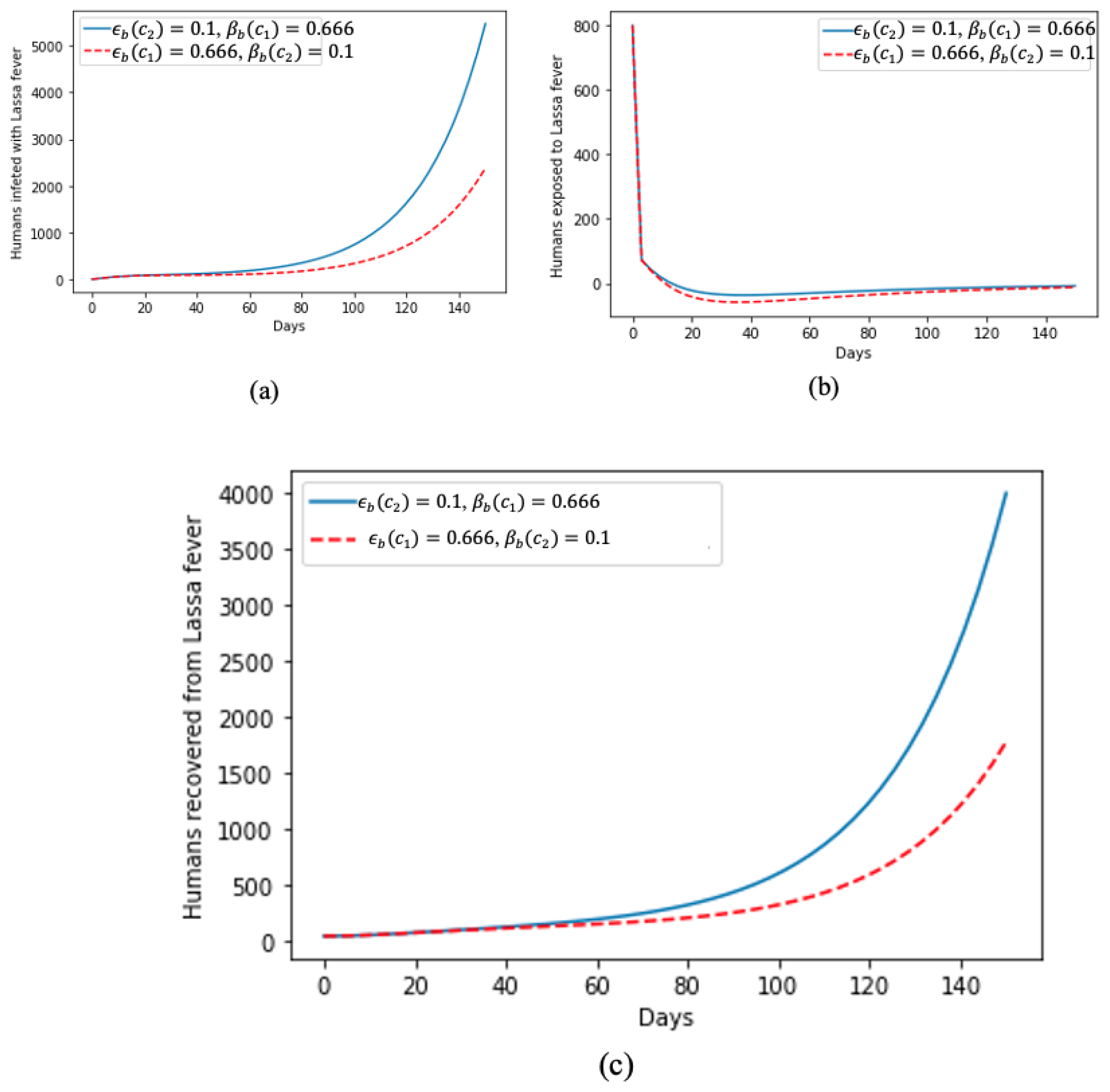
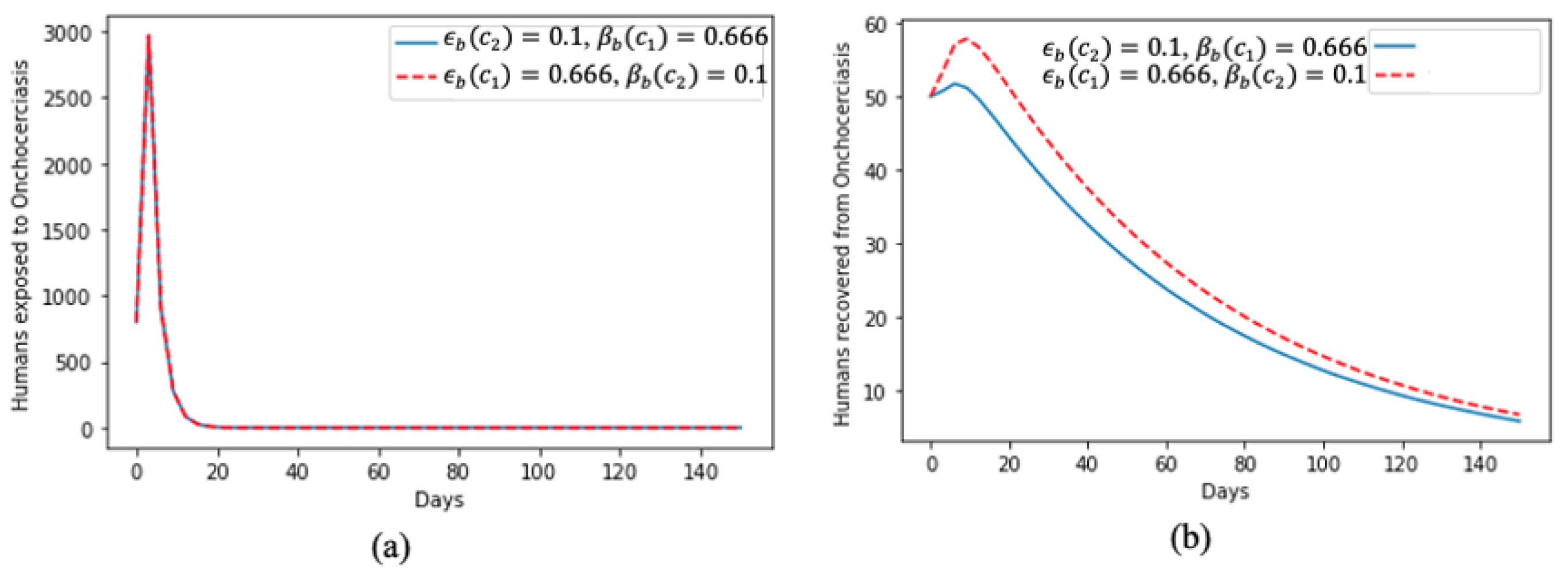
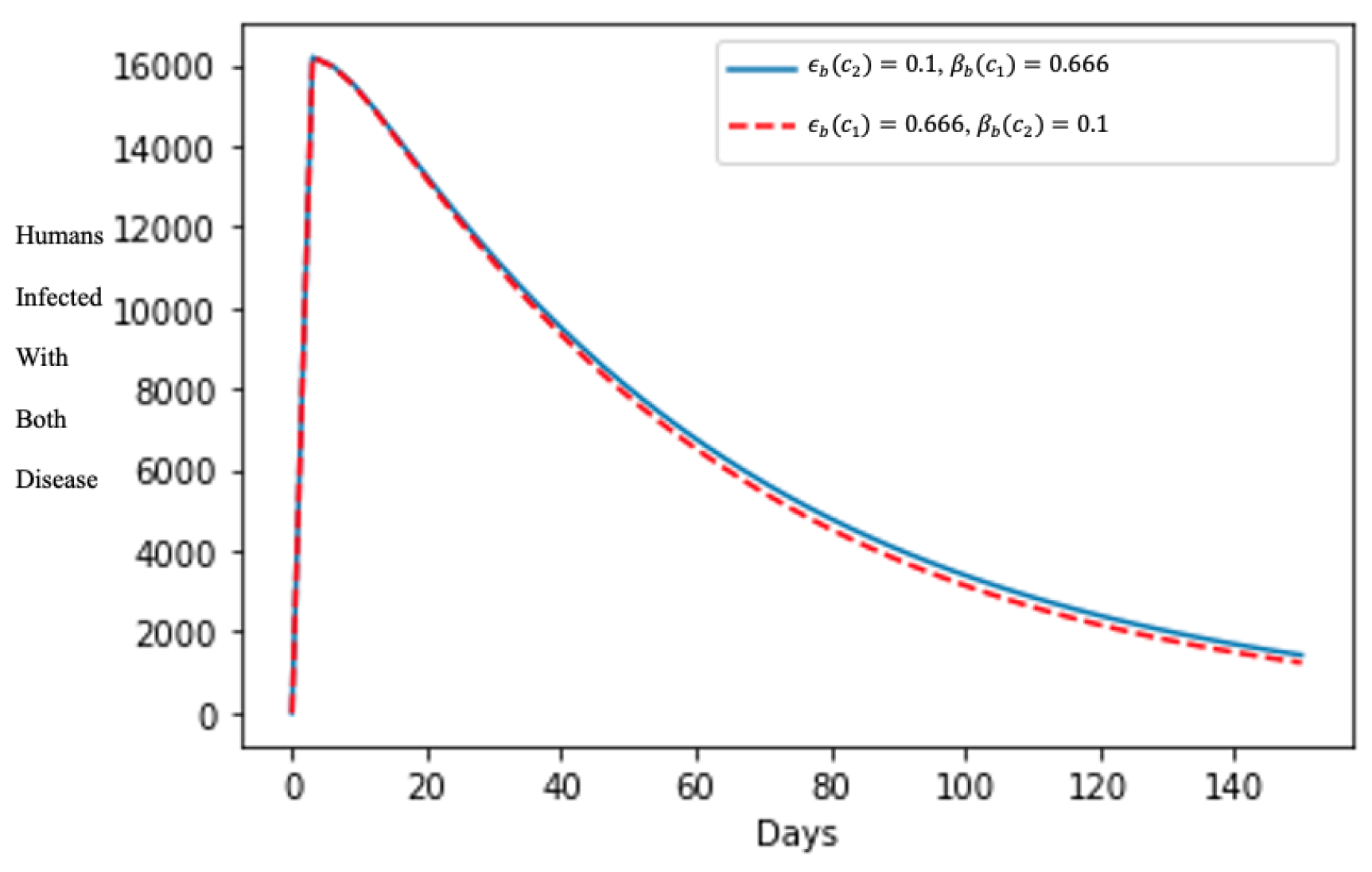
5. Numerical Simulation of the Model and Discussion of Results
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amazigo, U.; Noma, M.; Bump, J.; Bentin, B.; Liese, B.; Yameogo, L.; Zouré, H.; Seketeli, A. Chapter 15. In Onchocerciasis Disease and Mortality in Sub Saharan Africa; World Bank: Washington, DC, USA, 2006. [Google Scholar]
- Oguoma, I.C.; Acho, T.M. Mathematical modelling of the spread and control of onchocerciasis in tropical countries: Case study in Nigeria. Hindawi 2014, 2014, 631658. [Google Scholar] [CrossRef]
- Hassan, A.; Shaban, N. Onchocerciasis dynamics: Modelling the effects of treatment, education and vector control. J. Biol. Dyn. 2020, 14, 245–268. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. African programme for onchocerciasis control: Meeting of national onchocerciasis task forces, September 2012. Wkly. Epidemiol. Rec. 2012, 87, 494–502. [Google Scholar]
- Abioye, A.I.; Peter, O.J.; Ogunseye, H.A.; Oguntolu, F.A.; Oshinubi, K.; Ibrahim, A.A.; Khan, I. Mathematical model of COVID-19 in Nigeria with optimal control. Results Phys. 2021, 28, 104598. [Google Scholar] [CrossRef] [PubMed]
- Oshinubi, K.; Peter, O.J.; Addai, E.; Mwizerwa, E.; Babasola, O.; Nwabufo, I.V.; Sane, I.; Adam, U.M.; Adeniji, A.; Agbaje, J.O. Mathematical Modelling of Tuberculosis Outbreak in an East African Country Incorporating Vaccination and Treatment. Computation 2023, 11, 143. [Google Scholar] [CrossRef]
- Mopecha, J.P.; Thieme, H.R. Competitive Dynamics in a Model for Onchocerciasis with Cross-Immunity. Can. Appl. Math. Q. 2003, 11, 339–376. [Google Scholar]
- Peter, O.J.; Ibrahim, M.O.; Edogbanya, H.O.; Oguntolu, F.A.; Oshinubi, K.; Ibrahim, A.A.; Ayoola, T.A.; Lawal, J.O. Direct and indirect transmission of typhoid fever model with optimal control. Results Phys. 2021, 27, 104463. [Google Scholar] [CrossRef]
- Basáñez, M.-G.; Ricárdez-Esquinca, J. Models for the population biology and control of human onchocerciasis. Trends Parasitol. 2001, 17, 430–438. [Google Scholar] [CrossRef]
- Ngungu, M.; Addai, E.; Adeniji, A.; Adam, U.M.; Oshinubi, K. Mathematical epidemiological modeling and analysis of monkeypox dynamism with non-pharmaceutical intervention using real data from United Kingdom. Front. Public Health 2023, 11, 1101436. [Google Scholar] [CrossRef]
- Niger, A.M.; Gumel, A.B. Mathematical analysis of the role of repeated exposure on malaria transmission dynamics. Differ. Equ. Dyn. Syst. 2008, 16, 251–287. [Google Scholar] [CrossRef]
- Pontryagin, L.S.; Boltyanskii, V.T.; Gamkrelidze, R.V.; Mischeuko, E.F. The mathematical Theory of Optimal Processes; Selected Works IV; Gordon and Breach Science Publishers: New York, NY, USA, 1986. [Google Scholar]
- Plaisier, A.P.; Alley, E.S.; van Oortmarssen, G.J.; Boatin, B.A.; Habbema, J.D.F. Required duration of combined annual ivermectin treatment and vector control program in west Africa. Bull. World Health Organ. 1997, 75, 237. [Google Scholar] [PubMed]
- Remme, J.; Sole, G.D.; van Oortmarssen, G.J. The predicted and observed decline in onchocerciasis infection during 14 years of successful control of black flies in West Africa. Bull. World Health Organ. 1990, 68, 331–339. [Google Scholar]
- Bawa, M.; Abdulrahman, S.; Jimoh, O.R.; Adabara, N.U. Stability analysis of disease free equilibrium state for Lassa fever disease. J. Sci. Technol. Math. Educ. 2012, 9, 115–123. [Google Scholar]
- Tomori, O.; Fabiyi, A.; Sorungbe, A.; Smith, A.; Cormick, J.B. Viral hemorrhagic fever antibodies in Nigeria populations. Am. J. Trop. Med. Hyg. 1998, 38, 407–410. [Google Scholar] [CrossRef]
- den Driessche, P.V.; Watmough, J. Reproduction numbers and subthreshold endemic equilibria for compartmental models of disease transmission. Math. Biosci. 2002, 180, 29–48. [Google Scholar] [CrossRef] [PubMed]
- James, T.O.; Abdulrahman, S.; Akinyemi, S.; Akinwande, N.I. Dynamics transmission of Lassa fever. Int. J. Innov. Res. Educ. Sci. 2015, 2, 2349–5219. [Google Scholar]
- Onifade, A.A. Subharmonic Bifurcation in Malaria-Lassa Fever Co-Infection Epidemic Model with Optimal Control Application. Ph.D. Thesis, University of Ibadan, Ibadan, Nigeria, 2021. [Google Scholar]
- McKendrick, J.Q.; Tennant, W.S.; Tildesley, M.J. Modelling seasonality of Lassa fever incidences and vector dynamics in Nigeria. PLoS Neglected Trop. Dis. 2023, 17, e0011543. [Google Scholar] [CrossRef]
- Cheke, R.A.; Sowah, S.A.; Avissey, H.S.; Fiasorgbor, G.K.; Garms, R. Seasonal variation in onchocerciasis transmission by Simulium squamosum at perennial breeding sites in Togo. Trans. R. Soc. Trop. Med. Hyg. 1992, 86, 67–71. [Google Scholar] [CrossRef]
- Blayneh, K.W.; Cao, Y.; Kwon, H.-D. Optimal control of vector-borne diseases: Treatment and prevention. Discret. Contin. Dyn. Syst. B 2009, 11, 587–611. [Google Scholar] [CrossRef]
- Okuonghae, D.; Okuonghae, R.A. Mathematical model for Lassa fever. J. Niger. Assoc. Math. Phys. 2006, 10, 457–464. [Google Scholar] [CrossRef]
- Ogabi, C.O.; Olusa, T.V.; Madufor, M.O. Controlling Lassa Fever Transmission in Northern Part of Edo State, Nigeria Using Sir Model. N. Y. Sci. J. 2012, 5, 190–197. [Google Scholar]
- Adeniji, A.A.; Noufe, H.A.; Mkolesia, A.C.; Shatalov, M.Y. An Approximate Solution to Predator-prey Models Using The Differential Transform Method and Multi-step Differential Transform Method, in Comparison with Results of The Classical Runge-kutta Method. Math. Stat. 2021, 9, 799–805. [Google Scholar]
- Smith, M.E.; Bilal, S.; Lakwo, T.L.; Habomugisha, P.; Tukahebwa, E.; Byamukama, E.; Katabarwa, M.N.; Richards, F.O.; Cupp, E.W.; Unnasch, T.R.; et al. Accelerating river blindness elimination by supplementing MDA with a vegetation “slash and clear” vector control strategy: A data-driven modeling analysis. Sci. Rep. 2019, 9, 15274. [Google Scholar] [CrossRef] [PubMed]
- Alley, W.S.; van Oortmarssen, G.J.; Boatin, B.A.; Nagelkerke, N.J.; Plaisier, A.P.; Remme, J.H.; Lazdins, J.; Borsboom, G.J.; Habbema, J.D.F. Macrofilaricides and onchocerciasis control, mathematical modelling of the prospects for elimination. BMC Public Health 2001, 1, 12. [Google Scholar] [CrossRef] [PubMed]
- Hamley, J.I.D.; Milton, P.; Walker, M.; Basáñez, M.G. Modelling exposure heterogeneity and density dependence in onchocerciasis using a novel individual-based transmission model, EPIONCHO-IBM: Implications for elimination and data needs. PLoS Neglected Trop. Dis. 2019, 13, e0007557. [Google Scholar] [CrossRef]
- Bako, D.U.; Akinwande, N.I.; Enagi, A.I.; Kuta, F.A.; Abdulrahman, S. Stability Analysis of a Mathematical Model for Onchocerciaisis Disease Dynamics. J. Appl. Sci. Environ. Manag. 2017, 21, 663–671. [Google Scholar] [CrossRef]
- Hamley, J.I.D.; Walker, M.; Coffeng, L.E.; Milton, P.; de Vlas, S.J.; Stolk, W.A.; Basáñez, M. Structural Uncertainty in Onchocerciasis Transmission Models Influences the Estimation of Elimination Thresholds and Selection of Age Groups for Seromonitoring. J. Infect. Dis. 2020, 221 (Suppl. S5), S510–S518. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Definition | Symbols |
|---|---|
| Susceptible to human recruitment | |
| Susceptible to black fly recruitment | |
| Susceptible term for rodent recruitment | |
| Rate of black fly biting | b |
| Rodent interaction rate | |
| Human interaction rate | |
| Onchocerciasis transmission rate in humans | |
| Onchocerciasis transmission rate in black flies | |
| Lassa fever transmission rate in individuals by infectious individuals | |
| Lassa fever transmission rate in individuals by infectious rodents | |
| Lassa fever transmission rate in rodents by infected rodents | |
| Human mortality rate per capita | |
| Black fly mortality rate per capita | |
| Rodent death rate per capita | |
| Rodent mortality rate due to hunting | |
| Black fly seasonal variation | |
| Rodent seasonal fluctuation | |
| Lassa fever progression rate in an exposed individual | |
| Onchocerciasis progression rate in the exposed human host | |
| Lassa fever progression rate in exposed rodents | |
| Onchocerciasis progression rate in exposed black flies | |
| The proportion of effective human onchocerciasis treatment | |
| Infectious human onchocerciasis treatment rate | |
| Proportion of successful Lassa fever treatment for exposed humans | |
| Lassa fever diagnostic tests for humans who have been exposed | |
| Proportion of successful Lassa fever treatment for infected humans | |
| Infectious human Lassa fever treatment rate | |
| Force of infection for Lassa fever in humans by infectious humans | |
| Force of infection for Lassa fever in humans by infectious rodents | |
| Force of infection for Lassa fever in rodent population by infectious rodents | |
| Onchocerciasis infection rate | |
| Rate of recovery from Lassa fever | |
| Rate of recovery from onchocerciasis | |
| Rate of loss of immunity to onchocerciasis |
| Symbols | Value | Source |
|---|---|---|
| 0.000212 | Estimated | |
| 0.065 | [11] | |
| 0.0054 | [16] | |
| b | 0.8 | Fixed |
| 0.75 | Assumed | |
| 0.55 | Assumed | |
| 0.099 | [22] | |
| 0.089 | [22] | |
| 0.00814 | [22] | |
| 0.055 | [16] | |
| 0.073 | Assumed | |
| 0.0000545 | Estimated | |
| 0.0665 | Estimated | |
| 0.058 | [23] | |
| 0.295 | Assumed | |
| 0.5 | Assumed | |
| 0.00021 | Assumed | |
| 0.082 | [24] | |
| 0.0585 | Assumed | |
| 0.83 | Assumed | |
| 0.0554 | Assumed | |
| 0–0.1 | Assumed to vary | |
| 0–0.1 | Assumed to vary | |
| 0.09 | Assumed | |
| 0.049 | Assumed | |
| 0–0.189 | Assumed to vary | |
| 0.43 | Assumed | |
| 0.0013692 | Assumed | |
| 0–0.2 | Assumed to vary | |
| 0–0.3 | Assumed to vary | |
| 0–0.2 | Assumed to vary | |
| 0–0.3 | Assumed to vary | |
| 0–0.5 | Assumed to vary | |
| 0–0.4 | Assumed to vary |
| Parameters | Sensitivity Index |
|---|---|
| 1 | |
| 1 | |
| Parameters | Sensitivity Index |
|---|---|
| b | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adeyemo, K.M.; Oshinubi, K.; Adam, U.M.; Adeniji, A. A Co-Infection Model for Onchocerciasis and Lassa Fever with Optimal Control Analysis. AppliedMath 2024, 4, 89-119. https://doi.org/10.3390/appliedmath4010006
Adeyemo KM, Oshinubi K, Adam UM, Adeniji A. A Co-Infection Model for Onchocerciasis and Lassa Fever with Optimal Control Analysis. AppliedMath. 2024; 4(1):89-119. https://doi.org/10.3390/appliedmath4010006
Chicago/Turabian StyleAdeyemo, Kabiru Michael, Kayode Oshinubi, Umar Muhammad Adam, and Adejimi Adeniji. 2024. "A Co-Infection Model for Onchocerciasis and Lassa Fever with Optimal Control Analysis" AppliedMath 4, no. 1: 89-119. https://doi.org/10.3390/appliedmath4010006
APA StyleAdeyemo, K. M., Oshinubi, K., Adam, U. M., & Adeniji, A. (2024). A Co-Infection Model for Onchocerciasis and Lassa Fever with Optimal Control Analysis. AppliedMath, 4(1), 89-119. https://doi.org/10.3390/appliedmath4010006