
Safety Toxicology Study of Reassortant Mopeia–Lassa Vaccine in Guinea Pigs

 ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Study Design
3.2. Clinical Observations
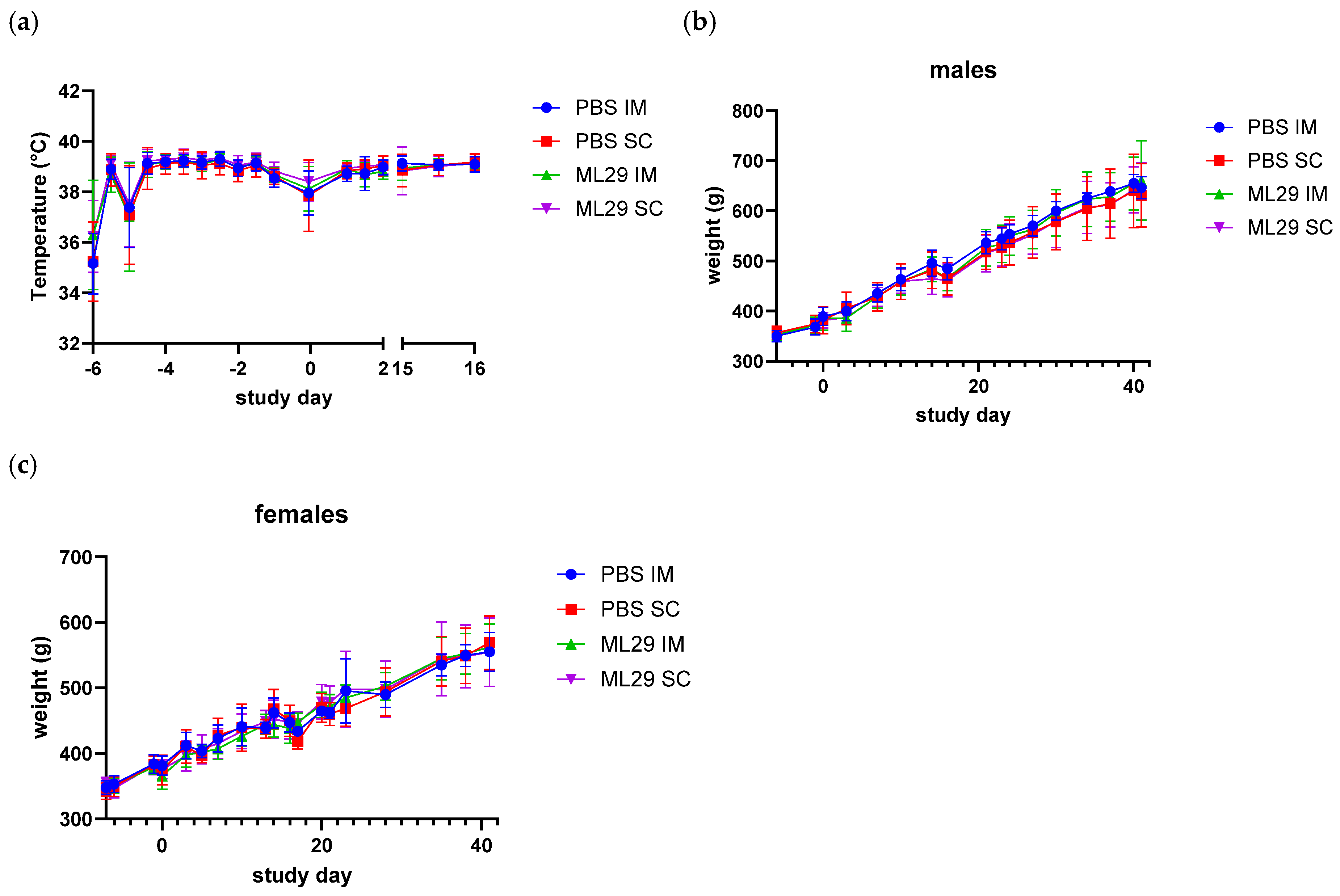
3.3. Body Temperature, Weight, and Food Consumption
3.4. Clinical Chemistry, Hematology, and Pathology
3.5. Biodistribution and Viral Shedding
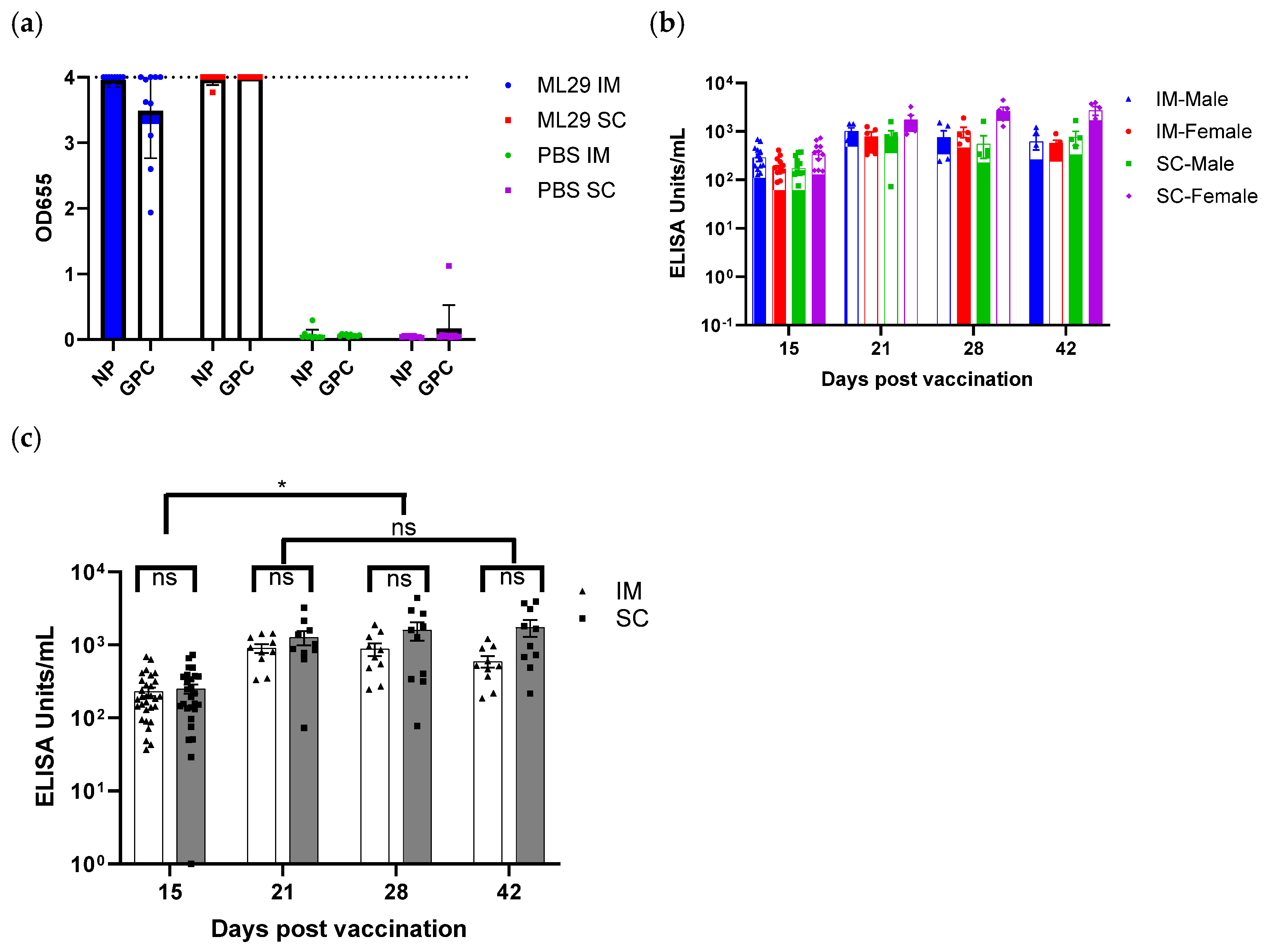
3.6. Immunogenicity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CFR | Code of Federal Regulations |
| CRO | Contract Research Organization |
| CDMO | Contract Development and Manufacturing Organization |
| ELISA | Enzyme-Linked Immunosorbent Assay |
| EMA | European Medicines Agency |
| FDA | Food and Drug Administration |
| HRP | Horseradish Peroxidase |
| IM | Intramuscular |
| GLP | Good Laboratory Practices |
| GMP | Good Manufacturing Practices |
| GP | Glycoprotein |
| GPC | Glycoprotein Precursor |
| LASV | Lassa Virus |
| LCMV | Lymphocytic Choriomeningitis Virus |
| LF | Lassa Fever |
| MOPV | Mopeia Virus |
| NHP | Non-Human Primates |
| NP | Nucleoprotein |
| PCR | Polymerase Chain Reaction |
| qPCR | Quantitative Polymerase Chain Reaction |
| qRT-PCR | Quantitative Reverse Transcriptase-PCR |
| RdRp | RNA-dependent RNA polymerase |
| SC | Subcutaneous |
| SSP | Stable Signal Peptide |
| TPP | Target Product Profile |
| USDA | United States Department of Agriculture |
| WHO | World Health Organization |
References
- Besson, M.E.; Pepin, M.; Metral, P.A. Lassa Fever: Critical Review and Prospects for Control. Trop. Med. Infect. Dis. 2024, 9, 178. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Falzarano, D.; Feldmann, H. Vaccines for viral hemorrhagic fevers–progress and shortcomings. Curr. Opin. Virol. 2013, 3, 343–351. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Warner, B.M.; Safronetz, D.; Stein, D.R. Current perspectives on vaccines and therapeutics for Lassa Fever. Virol. J. 2024, 21, 320. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Garry, R.F. Lassa fever-the road ahead. Nat. Rev. Microbiol. 2023, 21, 87–96. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Happi, A.N.; Happi, C.T.; Schoepp, R.J. Lassa fever diagnostics: Past, present, and future. Curr. Opin. Virol. 2019, 37, 132–138. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Siddle, K.J.; Eromon, P.; Barnes, K.G.; Mehta, S.; Oguzie, J.U.; Odia, I.; Schaffner, S.F.; Winnicki, S.M.; Shah, R.R.; Qu, J.; et al. Genomic Analysis of Lassa Virus during an Increase in Cases in Nigeria in 2018. N. Engl. J. Med. 2018, 379, 1745–1753. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Andersen, K.G.; Shapiro, B.J.; Matranga, C.B.; Sealfon, R.; Lin, A.E.; Moses, L.M.; Folarin, O.A.; Goba, A.; Odia, I.; Ehiane, P.E.; et al. Clinical Sequencing Uncovers Origins and Evolution of Lassa Virus. Cell 2015, 162, 738–750. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Manning, J.T.; Forrester, N.; Paessler, S. Lassa virus isolates from Mali and the Ivory Coast represent an emerging fifth lineage. Front. Microbiol. 2015, 6, 1037. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Monath, T.P.; Newhouse, V.F.; Kemp, G.E.; Setzer, H.W.; Cacciapuoti, A. Lassa virus isolation from Mastomys natalensis rodents during an epidemic in Sierra Leone. Science 1974, 185, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Olayemi, A.; Cadar, D.; Magassouba, N.; Obadare, A.; Kourouma, F.; Oyeyiola, A.; Fasogbon, S.; Igbokwe, J.; Rieger, T.; Bockholt, S.; et al. New Hosts of The Lassa Virus. Sci. Rep. 2016, 6, 25280. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ogunro, B.N.; Olugasa, B.O.; Kayode, A.; Ishola, O.O.; Kolawole, O.N.; Odigie, E.A.; Happi, C. Detection of Antibody and Antigen for Lassa Virus Nucleoprotein in Monkeys from Southern Nigeria. J. Epidemiol. Glob. Health 2019, 9, 125–127. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Happi, A.N.; Ogunsanya, O.A.; Ayinla, A.O.; Sijuwola, A.E.; Saibu, F.M.; Akano, K.; Nwofoke, C.; Elias, O.T.; Achonduh-Atijegbe, O.; Daodu, R.O.; et al. Lassa virus in novel hosts: Insights into the epidemiology of lassa virus infections in southern Nigeria. Emerg. Microbes Infect. 2024, 13, 2294859. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Dan-Nwafor, C.C.; Ipadeola, O.; Smout, E.; Ilori, E.; Adeyemo, A.; Umeokonkwo, C.; Nwidi, D.; Nwachukwu, W.; Ukponu, W.; Omabe, E.; et al. A cluster of nosocomial Lassa fever cases in a tertiary health facility in Nigeria: Description and lessons learned, 2018. Int. J. Infect. Dis. 2019, 83, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Shieh, W.J.; Demby, A.; Jones, T.; Goldsmith, C.S.; Rollin, P.E.; Ksiazek, T.G.; Peters, C.J.; Zaki, S.R. Pathology and Pathogenesis of Lassa Fever: Novel Immunohistochemical Findings in Fatal Cases and Clinico-pathologic Correlation. Clin. Infect. Dis. 2022, 74, 1821–1830. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Yun, N.E.; Walker, D.H. Pathogenesis of Lassa fever. Viruses 2012, 4, 2031–2048. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ficenec, S.C.; Percak, J.; Arguello, S.; Bays, A.; Goba, A.; Gbakie, M.; Shaffer, J.G.; Emmett, S.D.; Schieffelin, J.S.; Bausch, D. Lassa Fever Induced Hearing Loss: The Neglected Disability of Hemorrhagic Fever. Int. J. Infect. Dis. 2020, 100, 82–87. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mateer, E.J.; Huang, C.; Shehu, N.Y.; Paessler, S. Lassa fever-induced sensorineural hearing loss: A neglected public health and social burden. PLoS Negl. Trop. Dis. 2018, 12, e0006187. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- McCormick, J.B.; Fisher-Hoch, S.P. Lassa fever. Curr. Top. Microbiol. Immunol. 2002, 262, 75–109. [Google Scholar] [CrossRef] [PubMed]
- Simons, D. Lassa fever cases suffer from severe underreporting based on reported fatalities. Int. Health 2023, 15, 608–610. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Johnson, K.M.; McCormick, J.B.; Webb, P.A.; Smith, E.S.; Elliott, L.H.; King, I.J. Clinical virology of Lassa fever in hospitalized patients. J. Infect. Dis. 1987, 155, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Radoshitzky, S.R.; Buchmeier, M.J.; Charrel, R.N.; Gonzalez, J.J.; Gunther, S.; Hepojoki, J.; Kuhn, J.H.; Lukashevich, I.S.; Romanowski, V.; Salvato, M.S.; et al. ICTV Virus Taxonomy Profile: Arenaviridae 2023. J Gen Virol. 2023, 104, 001891. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Xing, J.; Ly, H.; Liang, Y. The Z proteins of pathogenic but not nonpathogenic arenaviruses inhibit RIG-I-like receptor-dependent interferon production. J. Virol. 2015, 89, 2944–2955. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Cohen, J. Unfilled Vials. Science 2016, 351, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.H.; Johnson, K.M.; Lange, J.V.; Gardner, J.J.; Kiley, M.P.; McCormick, J.B. Experimental infection of rhesus monkeys with Lassa virus and a closely related arenavirus, Mozambique virus. J. Infect. Dis. 1982, 146, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Moshkoff, D.A.; Salvato, M.S.; Lukashevich, I.S. Molecular characterization of a reassortant virus derived from Lassa and Mopeia viruses. Virus Genes 2007, 34, 169–176. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lukashevich, I.S.; Patterson, J.; Carrion, R.; Moshkoff, D.; Ticer, A.; Zapata, J.; Brasky, K.; Geiger, R.; Hubbard, G.B.; Bryant, J.; et al. A live attenuated vaccine for Lassa fever made by reassortment of Lassa and Mopeia viruses. J. Virol. 2005, 79, 13934–13942. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Johnson, D.M.; Cubitt, B.; Pfeffer, T.L.; de la Torre, J.C.; Lukashevich, I.S. Lassa Virus Vaccine Candidate ML29 Generates Truncated Viral RNAs Which Contribute to Interfering Activity and Attenuation. Viruses. 2021, 13, 214. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Carrion, R., Jr.; Patterson, J.L.; Johnson, C.; Gonzales, M.; Moreira, C.R.; Ticer, A.; Brasky, K.; Hubbard, G.B.; Moshkoff, D.; Zapata, J.; et al. A ML29 reassortant virus protects guinea pigs against a distantly related Nigerian strain of Lassa virus and can provide sterilizing immunity. Vaccine 2007, 25, 4093–4102. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lukashevich, I.S.; Carrion, R., Jr.; Salvato, M.S.; Mansfield, K.; Brasky, K.; Zapata, J.; Cairo, C.; Goicochea, M.; Hoosien, G.E.; Ticer, A.; et al. Safety, immunogenicity, and efficacy of the ML29 reassortant vaccine for Lassa fever in small non-human primates. Vaccine 2008, 26, 5246–5254. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zapata, J.C.; Poonia, B.; Bryant, J.; Davis, H.; Ateh, E.; George, L.; Crasta, O.; Zhang, Y.; Slezak, T.; Jaing, C.; et al. An attenuated Lassa vaccine in SIV-infected rhesus macaques does not persist or cause arenavirus disease but does elicit Lassa virus-specific immunity. Virol. J. 2013, 10, 52. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lukashevich, I.S.; Pushko, P. Vaccine platforms to control Lassa fever. Expert. Rev. Vaccines 2016, 15, 1135–1150. [Google Scholar] [CrossRef] [PubMed]
- Lukashevich, I.S.; Paessler, S.; de la Torre, J.C. Lassa virus diversity and feasibility for universal prophylactic vaccine. F1000Research 2019, 8, 134. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Verdier, F. Non-clinical vaccine safety assessment. Toxicology 2002, 174, 37–43. [Google Scholar] [CrossRef] [PubMed]
- FDA. Guidance for Industry. M3(R2) Nonclinical Safety Studies for the Conduct of Human Clinical Trials and Marketing Authorization for Pharmaceuticals 2010 [Cited 2025 15 May]. Available online: https://www.fda.gov/media/71542/download (accessed on 23 May 2025).
- Draize, J.H. Methods for the study of irritation and toxicity of substances applied topically to the skin and the mucous membranes. J. Pharmacol. Exp. Ther. 1944, 82, 377–390. [Google Scholar] [CrossRef]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 7th ed.; Iowa State University Press: Ames, IA, USA, 1980. [Google Scholar]
- Dunnett, C.W.; Crisafio, R. The operating characteristics of some official weight variation tests for tablets. J. Pharm. Pharmacol. 1955, 7, 314–327. [Google Scholar] [CrossRef] [PubMed]
- McCormick, J.B. Clinical, epidemiologic, and therapeutic aspects of Lassa fever. Med Microbiol Immunol. 1986, 175, 153–155. [Google Scholar] [CrossRef] [PubMed]
- Baize, S.; Marianneau, P.; Loth, P.; Reynard, S.; Journeaux, A.; Chevallier, M.; Tordo, N.; Deubel, V.; Contamin, H. Early and strong immune responses are associated with control of viral replication and recovery in lassa virus-infected cynomolgus monkeys. J. Virol. 2009, 83, 5890–5903. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hensley, L.E.; Smith, M.A.; Geisbert, J.B.; Fritz, E.A.; Daddario-DiCaprio, K.M.; Larsen, T.; Geisbert, T.W. Pathogenesis of Lassa fever in cynomolgus macaques. Virol. J. 2011, 8, 205. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Baillet, N.; Reynard, S.; Perthame, E.; Hortion, J.; Journeaux, A.; Mateo, M.; Carnec, X.; Schaeffer, J.; Picard, C.; Barrot, L.; et al. Systemic viral spreading and defective host responses are associated with fatal Lassa fever in macaques. Commun. Biol. 2021, 4, 27. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Cashman, K.A.; Smith, M.A.; Twenhafel, N.A.; Larson, R.A.; Jones, K.F.; Allen, R.D., 3rd; Dai, D.; Chinsangaram, J.; Bolken, T.C.; Hruby, D.E.; et al. Evaluation of Lassa antiviral compound ST-193 in a guinea pig model. Antiviral Res. 2011, 90, 70–79. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bell, T.M.; Shaia, C.I.; Bearss, J.J.; Mattix, M.E.; Koistinen, K.A.; Honnold, S.P.; Zeng, X.; Blancett, C.D.; Donnelly, G.C.; Shamblin, J.D.; et al. Temporal Progression of Lesions in Guinea Pigs Infected With Lassa Virus. Vet. Pathol. 2017, 54, 549–562. [Google Scholar] [CrossRef] [PubMed]
- Deschambault, Y.; Soule, G.; Klassen, L.; Sloan, A.; Audet, J.; Azaransky, K.; Musa, A.S.; Ahmad, A.; Akinpelu, A.M.; Mba, N.; et al. An Outbred Guinea Pig Disease Model for Lassa Fever Using a Host-Adapted Clade III Nigerian Lassa Virus. Viruses. 2023, 15, 769. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zapata, J.C.; Pauza, C.D.; Djavani, M.M.; Rodas, J.D.; Moshkoff, D.; Bryant, J.; Ateh, E.; Garcia, C.; Lukashevich, I.S.; Salvato, M.S. Lymphocytic choriomeningitis virus (LCMV) infection of macaques: A model for Lassa fever. Antivir. Res. 2011, 92, 125–138. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Okokhere, P.; Colubri, A.; Azubike, C.; Iruolagbe, C.; Osazuwa, O.; Tabrizi, S.; Chin, E.; Asad, S.; Ediale, E.; Rafiu, M.; et al. Clinical and laboratory predictors of Lassa fever outcome in a dedicated treatment facility in Nigeria: A retrospective, observational cohort study. Lancet Infect. Dis. 2018, 18, 684–695. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Rossi, G.; Liu, K.F.; Kershaw, H.; Riddell, D.; Hyndman, T.H.; Monks, D.; Musk, G.C. Biological Variation in Biochemistry Analytes in Laboratory Guinea Pigs (Cavia porcellus). Vet Sci. 2023, 10, 621. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gary, J.M.; Welch, S.R.; Ritter, J.M.; Coleman-McCray, J.; Huynh, T.; Kainulainen, M.H.; Bollweg, B.C.; Parihar, V.; Nichol, S.T.; Zaki, S.R.; et al. Lassa Virus Targeting of Anterior Uvea and Endothelium of Cornea and Conjunctiva in Eye of Guinea Pig Model. Emerg. Infect. Dis. 2019, 25, 865–874. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hastie, K.M.; Saphire, E.O. Lassa virus glycoprotein: Stopping a moving target. Curr. Opin. Virol. 2018, 31, 52–58. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sama, D.J.; Haider, N.; Guitian, J.; Osman, A.Y.; Ntoumi, F.; Zumla, A.; Kock, R.; Ansumana, R. Identifying risk factors for clinical Lassa fever in Sierra Leone, 2019–2021. Epidemiol. Infect. 2024, 152, e177. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Baric, T.J.; Reneer, Z.B. Animal Models, Therapeutics, and Vaccine Approaches to Emerging and Re-Emerging Flaviviruses. Viruses 2024, 17, 1. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lauring, A.S.; Jones, J.O.; Andino, R. Rationalizing the development of live attenuated virus vaccines. Nat. Biotechnol. 2010, 28, 573–579. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ambrosio, A.; Saavedra, M.; Mariani, M.; Gamboa, G.; Maiza, A. Argentine hemorrhagic fever vaccines. Hum. Vaccin. 2011, 7, 694–700. [Google Scholar] [CrossRef] [PubMed]
- Nunberg, J.H.; Westover, J.B.; York, J.; Jung, K.H.; Bailey, K.W.; Boardman, K.M.; Li, M.; Furnell, R.S.; Wasson, S.R.; Murray, J.S.; et al. Restoration of virulence in the attenuated Candid#1 vaccine virus requires reversion at both positions 168 and 427 in the envelope glycoprotein GPC. J. Virol. 2024, 98, e0011224. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Riviere, Y.; Ahmed, R.; Southern, P.J.; Buchmeier, M.J.; Oldstone, M.B. Genetic mapping of lymphocytic choriomeningitis virus pathogenicity: Virulence in guinea pigs is associated with the L RNA segment. J. Virol. 1985, 55, 704–709. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ahmed, R.; Simon, R.S.; Matloubian, M.; Kolhekar, S.R.; Southern, P.J.; Freedman, D.M. Genetic analysis of in vivo-selected viral variants causing chronic infection: Importance of mutation in the L RNA segment of lymphocytic choriomeningitis virus. J. Virol. 1988, 62, 3301–3308. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Taniguchi, S.; Saito, T.; Paroha, R.; Huang, C.; Paessler, S.; Maruyama, J. Unraveling factors responsible for pathogenic differences in Lassa virus strains. bioRxiv 2024. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kiley, M.P.; Lange, J.V.; Johnson, K.M. Protection of rhesus monkeys from Lassa virus by immunisation with closely related Arenavirus. Lancet 1979, 2, 738. [Google Scholar] [CrossRef] [PubMed]
- Darbre, S.; Johnson, S.; Kallert, S.; Lambert, P.H.; Siegrist, C.A.; Pinschewer, D.D. The Nucleoprotein Is Required for Lymphocytic Choriomeningitis Virus-Based Vaccine Vector Immunogenicity. J. Virol. 2015, 89, 11734–11738. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mateo, M.; Reynard, S.; Carnec, X.; Journeaux, A.; Baillet, N.; Schaeffer, J.; Picard, C.; Legras-Lachuer, C.; Allan, R.; Perthame, E.; et al. Vaccines inducing immunity to Lassa virus glycoprotein and nucleoprotein protect macaques after a single shot. Sci. Transl. Med. 2019, 11, eaaw3163. [Google Scholar] [CrossRef] [PubMed]
- Fisher-Hoch, S.P.; McCormick, J.B. Lassa fever vaccine. Expert. Rev. Vaccines 2004, 3, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Prescott, J.B.; Marzi, A.; Safronetz, D.; Robertson, S.J.; Feldmann, H.; Best, S.M. Immunobiology of Ebola and Lassa virus infections. Nat. Rev. Immunol. 2017, 17, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Murphy, H.; Ly, H. Understanding Immune Responses to Lassa Virus Infection and to Its Candidate Vaccines. Vaccines 2022, 10, 1668. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- WHO. WHO Target Product Profile for Lassa Virus Vaccine; WHO: Geneva, Switzerland, 2017. Available online: https://www.who.int/publications/m/item/who-target-product-profile-for-lassa-virus-vaccine (accessed on 20 May 2025).
- Reed, N.S.; Brewer, C.C.; Akintunde, G.; Blackie, F.F.; Charles, L.; Fast, P.; Lambert, P.H.; Okogbenin, S.; Paessler, S.; Pinschewer, D.D.; et al. Report of a SPEAC webinar 22 september 2023: Sensorineural hearing loss, lassa virus disease and vaccines. Vaccine 2025, 43 Pt. 1, 126525. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Flaxman, A.; Sebastian, S.; Appelberg, S.; Cha, K.M.; Ulaszewska, M.; Purushotham, J.; Gilbride, C.; Sharpe, H.; Spencer, A.J.; Bibi, S.; et al. Potent immunogenicity and protective efficacy of a multi-pathogen vaccination targeting Ebola, Sudan, Marburg and Lassa viruse. PLoS Pathog. 2024, 20, e1012262. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.L.; Woolsey, C.; Borisevich, V.; Agans, K.N.; Prasad, A.N.; Deer, D.J.; Geisbert, J.B.; Dobias, N.S.; Fenton, K.A.; Cross, R.W.; et al. A recombinant VSV-vectored vaccine rapidly protects nonhuman primates against lethal Nipah virus disease. Proc. Natl. Acad. Sci. USA 2022, 119, e2200065119. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tschismarov, R.; Van Damme, P.; Germain, C.; De Coster, I.; Mateo, M.; Reynard, S.; Journeaux, A.; Tomberger, Y.; Withanage, K.; Haslwanter, D.; et al. Immunogenicity, safety, and tolerability of a recombinant measles-vectored Lassa fever vaccine: A randomised, placebo-controlled, first-in-human trial. Lancet 2023, 401, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Carnec, X.; Mateo, M.; Page, A.; Reynard, S.; Hortion, J.; Picard, C.; Yekwa, E.; Barrot, L.; Barron, S.; Vallve, A.; et al. A Vaccine Platform against Arenaviruses Based on a Recombinant Hyperattenuated Mopeia Virus Expressing Heterologous Glycoproteins. J. Virol. 2018, 92, e02230-17. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Saito, T.; Reyna, R.A.; Taniguchi, S.; Littlefield, K.; Paessler, S.; Maruyama, J. Vaccine Candidates against Arenavirus Infections. Vaccines 2023, 11, 635. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kurup, D.; Fisher, C.R.; Scher, G.; Yankowski, C.; Testa, A.; Keshwara, R.; Abreu-Mota, T.; Lambert, R.; Ferguson, M.; Rinaldi, W.; et al. Tetravalent Rabies-Vectored Filovirus and Lassa Fever Vaccine Induces Long-term Immunity in Nonhuman Primates. J. Infect. Dis. 2021, 224, 995–1004. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Carey, B.D.; Yu, S.; Geiger, J.; Ye, C.; Huzella, L.M.; Reeder, R.J.; Mehta, M.; Hirsch, S.; Bernbaum, R.; Cubitt, B.; et al. A Lassa virus live attenuated vaccine candidate that is safe and efficacious in guinea pigs. NPJ Vaccines 2024, 9, 220. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fischer, R.J.; Purushotham, J.N.; van Doremalen, N.; Sebastian, S.; Meade-White, K.; Cordova, K.; Letko, M.; Jeremiah Matson, M.; Feldmann, F.; Haddock, E.; et al. ChAdOx1-vectored Lassa fever vaccine elicits a robust cellular and humoral immune response and protects guinea pigs against lethal Lassa virus challenge. NPJ Vaccines 2021, 6, 32. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Andrade, V.M.; Cashman, K.; Rosenke, K.; Wilkinson, E.; Josleyn, N.; Lynn, G.; Steffens, J.; Vantongeren, S.; Wells, J.; Schmaljohn, C.; et al. The DNA-based Lassa vaccine INO-4500 confers durable protective efficacy in cynomolgus macaques against lethal Lassa fever. Commun. Med. 2024, 4, 253. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ronk, A.J.; Lloyd, N.M.; Zhang, M.; Atyeo, C.; Perrett, H.R.; Mire, C.E.; Hastie, K.M.; Sanders, R.W.; Brouwer, P.J.M.; Saphire, E.O.; et al. A Lassa virus mRNA vaccine confers protection but does not require neutralizing antibody in a guinea pig model of infection. Nat. Commun. 2023, 14, 5603. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Travieso, T.; Li, J.; Mahesh, S.; Mello, J.; Blasi, M. The use of viral vectors in vaccine development. NPJ Vaccines 2022, 7, 75. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]


{kind=link}
{kind=link}
{kind=link}
| Group Number | Test Article | Target Dose (PFU) 1 | Route | Injection Day(s) | Animal Designation and Number | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Toxicology 2 | Recovery 3 | Biodistribution | ||||||||
| Male | Female | Male | Female | Male | Female | |||||
| 1 | Control (PBS) | 0 | IM | 1 and 15 | 10 | 10 | 5 | 5 | 15 | 15 |
| 2 | Control (PBS) | 0 | SC | 1 and 15 | 10 | 10 | 5 | 5 | -- | -- |
| 3 | ML29-SF LASV Vaccine | 1 × 104 | IM | 1 and 15 | 10 | 10 | 5 | 5 | 15 | 15 |
| 4 | ML29-SF LASV Vaccine | 1 × 104 | SC | 1 and 15 | 10 | 10 | 5 | 5 | 15 | 15 |
| Hematology | Clinical Chemistry |
|---|---|
| cell morphology 1 | albumin (Alb) |
| total leukocyte count (WBC) | A/G ratio (A/G) |
| neutrophils (NEU) | aspartate aminotransferase (AST) |
| lymphocytes (LYM) | total protein (T-Prot; TPRO) |
| monocytes (MONO) | gamma glutamyl transpeptidase (GGT) |
| basophils (BAS) | glucose (Gluc, GLU) |
| eosinophils (EOS) | triglyceride (Trig) |
| % neutrophils (NEU%) | creatinine (Creat, CREA) |
| % lymphocytes (LYM%) | potassium (K) |
| % monocytes (MON%) | calcium (Ca) |
| % basophils (BAS%) | creatine kinase (CK, CPK) |
| % eosinophils (EOS%) | globulin (Glob) |
| erythrocyte count (RBC-Red Blood Cell) | alanine aminotransferase (ALT) |
| hemoglobin concentration (HGB, HB) | alkaline phosphatase (ALP) |
| hematocrit (HCT) | total bilirubin (T-Bili, TBIL) |
| mean corpuscular volume (MCV) | direct bilirubin (D-Bili, DBIL) |
| mean corpuscular hemoglobin (MCH) | cholesterol (Chol) |
| mean corpuscular hemoglobin concentration (MCHC) | urea nitrogen (UN) |
| platelet count (PLT) | sodium (Na) |
| mean platelet volume (MPV) | chloride (Cl) |
| red blood cell distribution width (RDW%) | phosphorus (Phos, P) |
| reticulocytes (% and ABS, RET and RET%) |
| Forward Primer (5’ to 3’) | TCCAACATATTGCCACCATC |
| Probe (5’ to 3’) | FAM-TGCCTTCACAGCTGCACCCA-MGB |
| Reverse Primer (5’ to 3’) | GCTGACTCAAAGTCATCCCA |
| Synthetic RNA (5’ to 3)’ | GGGUUGAUUGUCUCCAACAUAUUGCCACCAUCCAGCAU GCAAGCUCCUGCCUUCACAGCUGCACCCAAGCUAAAAU UAUAACCUGAGAUAUUCAAAGAGCUUUUCUUGGUGUCA AUCAUAUUUAGGAUGGGAUGACUUUGAGUCAGCCUGUCUAAG |
| Group Number | Treatment, Route | Mortality/Morbidity | General Health | Ophthal- mology | Food Consumption | Organ Weights | Hematology | Clinical Chemistry | Shedding * |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Control PBS, IM | None | Healthy | No Change | Normal | Normal | No Change | No Change | ND |
| 2 | Control PBS, SC | None | Healthy | No Change | Normal | Normal | No Change | No Change | ND |
| 3 | ML29-SF LASV Vaccine, IM | None | Healthy | No Change | Normal | Normal | No Change | No Change | ND |
| 4 | ML29-SF LASV Vaccine, SC | None | Healthy | No Change | Normal | Normal | No Change | No Change | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wahle, B.S.; Pushko, P.; Albanese, K.; Johnson, D.M.; Tretyakova, I.; Lukashevich, I.S.; Rudge, T. Safety Toxicology Study of Reassortant Mopeia–Lassa Vaccine in Guinea Pigs. Future Pharmacol. 2025, 5, 26. https://doi.org/10.3390/futurepharmacol5020026
Wahle BS, Pushko P, Albanese K, Johnson DM, Tretyakova I, Lukashevich IS, Rudge T. Safety Toxicology Study of Reassortant Mopeia–Lassa Vaccine in Guinea Pigs. Future Pharmacology. 2025; 5(2):26. https://doi.org/10.3390/futurepharmacol5020026
Chicago/Turabian StyleWahle, Bradley S., Peter Pushko, Katie Albanese, Dylan M. Johnson, Irina Tretyakova, Igor S. Lukashevich, and Thomas Rudge. 2025. "Safety Toxicology Study of Reassortant Mopeia–Lassa Vaccine in Guinea Pigs" Future Pharmacology 5, no. 2: 26. https://doi.org/10.3390/futurepharmacol5020026
APA StyleWahle, B. S., Pushko, P., Albanese, K., Johnson, D. M., Tretyakova, I., Lukashevich, I. S., & Rudge, T. (2025). Safety Toxicology Study of Reassortant Mopeia–Lassa Vaccine in Guinea Pigs. Future Pharmacology, 5(2), 26. https://doi.org/10.3390/futurepharmacol5020026