

Metabolomics Approach to Reveal the Effects of Ocean Acidification on the Toxicity of Harmful Microalgae: A Review of the Literature

Abstract

1. Introduction
2. Algal Toxins
2.1. Paralytic Shellfish Poisoning
2.2. Diarrhetic Shellfish Poisoning
2.3. Amnesic Shellfish Poisoning
2.4. Neurotoxic Shellfish Poisoning
2.5. Azaspiracid Shellfish Poisoning
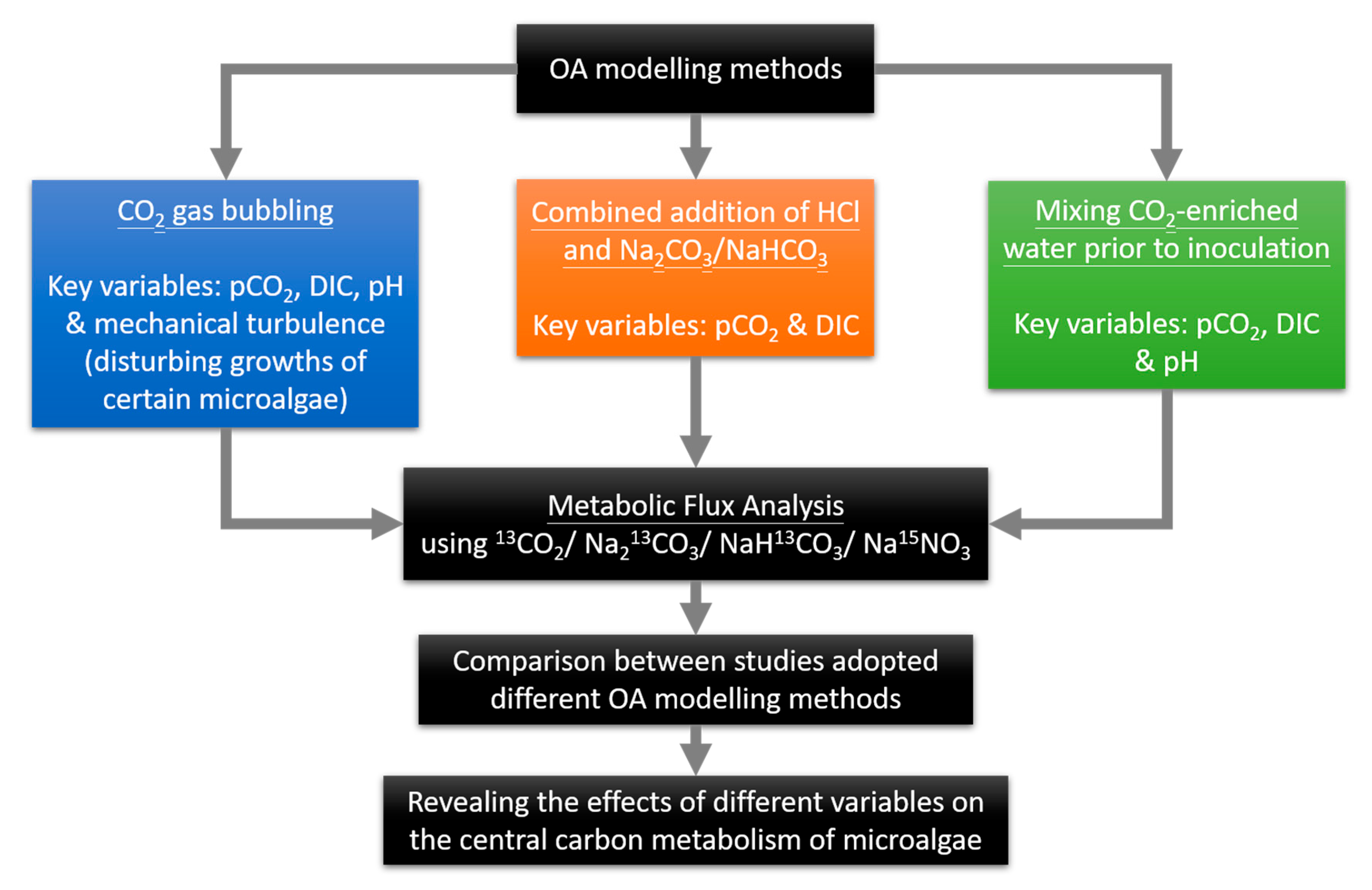
3. Ocean Acidification
3.1. Effects of OA on the Toxicity of Microalgae
3.1.1. STXs-Producing Microalgae
3.1.2. DA-Producing Microalgae
3.1.3. PbTx-Producing Microalgae
3.2. OA May Affect the Central Carbon Metabolism of Toxic Microalgae
4. Future Research Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- IPCC. IPCC Special Report on the Ocean and Cryosphere in a Changing Climate; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2019. [Google Scholar]
- Lopez, C.; Jewett, E.; Dortch, Q.; Walton, B.; Hudnell, H. Scientific Assessment of Freshwater Harmful Algal Blooms; Interagency Working Group on Harmful Algal Blooms, Hypoxia, and Human Health of the Joint Subcommittee on Ocean Science and Technology: Washington, DA, USA, 2008.
- Gerssen, A.; Pol-Hofstad, I.E.; Poelman, M.; Mulder, P.P.; van den Top, H.J.; de Boer, J. Marine toxins: Chemistry, toxicity, occurrence and detection, with special reference to the Dutch situation. Toxins 2010, 2, 878–904. [Google Scholar] [CrossRef] [PubMed]
- Van Dolah, F.M. Marine algal toxins: Origins, health effects, and their increased occurrence. Environ. Health Perspect. 2000, 108 (Suppl. S1), 133–141. [Google Scholar] [CrossRef]
- Wang, D.Z. Neurotoxins from marine dinoflagellates: A brief review. Mar. Drugs 2008, 6, 349–371. [Google Scholar] [CrossRef] [PubMed]
- Kouakou, C.R.C.; Poder, T.G. Economic impact of harmful algal blooms on human health: A systematic review. J. Water Health 2019, 17, 499–516. [Google Scholar] [CrossRef] [PubMed]
- Park, T.G.; Lim, W.A.; Park, Y.T.; Lee, C.K.; Jeong, H.J. Economic impact, management and mitigation of red tides in Korea. Harmful Algae 2013, 30, S131–S143. [Google Scholar] [CrossRef]
- Larkin, S.L.; Adams, C.M.; Ballyram, D.M.; Hodges, A. Red tides and coastal businesses: Measuring economic consequences in Florida. Soc. Nat. Resour. 2007, 20, 2007. [Google Scholar] [CrossRef]
- Sanseverino, I.; Conduto, D.; Pozzoli, L.; Dobricic, S.; Lettieri, T. Algal bloom and its economic impact; Publications Office of the European Union: Luxembourg, 2016. [Google Scholar] [CrossRef]
- Gobler, C.J. Climate Change and Harmful Algal Blooms: Insights and perspective. Harmful Algae 2020, 91, 101731. [Google Scholar] [CrossRef]
- Moore, S.K.; Trainer, V.L.; Mantua, N.J.; Parker, M.S.; Laws, E.A.; Backer, L.C.; Fleming, L.E. Impacts of climate variability and future climate change on harmful algal blooms and human health. In Proceedings of the Centers for Oceans and Human Health Investigators Meeting, Woods Hole, MA, USA, 24–27 April 2007; p. S4. [Google Scholar]
- Wells, M.L.; Karlson, B.; Wulff, A.; Kudela, R.; Trick, C.; Asnaghi, V.; Berdalet, E.; Cochlan, W.; Davidson, K.; De Rijcke, M.; et al. Future HAB science: Directions and challenges in a changing climate. Harmful Algae 2020, 91, 101632. [Google Scholar] [CrossRef]
- Wells, M.L.; Trainer, V.L.; Smayda, T.J.; Karlson, B.S.; Trick, C.G.; Kudela, R.M.; Ishikawa, A.; Bernard, S.; Wulff, A.; Anderson, D.M.; et al. Harmful algal blooms and climate change: Learning from the past and present to forecast the future. Harmful Algae 2015, 49, 68–93. [Google Scholar] [CrossRef]
- Griffith, A.W.; Gobler, C.J. Harmful algal blooms: A climate change co-stressor in marine and freshwater ecosystems. Harmful Algae 2020, 91, 101590. [Google Scholar] [CrossRef]
- Kazmi, S.S.U.H.; Yapa, N.; Karunarathna, S.C.; Suwannarach, N. Perceived Intensification in Harmful Algal Blooms Is a Wave of Cumulative Threat to the Aquatic Ecosystems. Biology 2022, 11, 852. [Google Scholar] [CrossRef]
- Hallegraeff, G.; Enevoldsen, H.; Zingone, A.; Schweibold, L.; De Rijcke, M. GLOBAL Harmful Algal Bloom Status Report 2021; UNESCO: Paris, France, 2021. [Google Scholar]
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef] [PubMed]
- Schantz, E.J.; Ghazarossian, V.E.; Schnoes, H.K.; Strong, F.M.; Springer, J.P.; Pezzanite, J.O.; Clardy, J. Letter: The structure of saxitoxin. J. Am. Chem. Soc. 1975, 97, 1238. [Google Scholar] [CrossRef] [PubMed]
- Oshima, Y.; Blackburn, S.; Hallegraeff, G. Comparative study on paralytic shellfish toxin profiles of the dinoflagellate Gymnodinium catenatum from three different countries. Mar. Biol. 1993, 116, 471–476. [Google Scholar] [CrossRef]
- Usup, G.; Kulis, D.M.; Anderson, D.M. Growth and toxin production of the toxic dinoflagellate Pyrodinium bahamense var. compressum in laboratory cultures. Nat. Toxins 1994, 2, 254–262. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Evans, W.R.; Yin, Q.Q.; Bell, P.; Moczydlowski, E. Evidence for paralytic shellfish poisons in the freshwater cyanobacterium Lyngbya wollei (Farlow ex Gomont) comb. nov. Appl. Environ. Microbiol. 1997, 63, 3104–3110. [Google Scholar] [CrossRef]
- Hoeger, S.J.; Shaw, G.; Hitzfeld, B.C.; Dietrich, D.R. Occurrence and elimination of cyanobacterial toxins in two Australian drinking water treatment plants. Toxicon 2004, 43, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, W.; Li, D.; Shen, Y.; Li, G.; Liu, Y. First report of aphantoxins in China--waterblooms of toxigenic Aphanizomenon flos-aquae in Lake Dianchi. Ecotoxicol. Environ. Saf. 2006, 65, 84–92. [Google Scholar] [CrossRef]
- Pomati, F.; Sacchi, S.; Rossetti, C.; Giovannardi, S.; Onodera, H.; Oshima, Y.; Neilan, B.A. The Freshwater Cyanobacterium Planktothrix Sp. Fp1: Molecular Identification and Detection of Paralytic Shellfish Poisoning Toxins. J. Phycol. 2000, 36, 553–562. [Google Scholar] [CrossRef]
- Cestele, S.; Catterall, W.A. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie 2000, 82, 883–892. [Google Scholar] [CrossRef]
- Llewellyn, L.E. Saxitoxin, a toxic marine natural product that targets a multitude of receptors. Nat. Prod. Rep. 2006, 23, 200–222. [Google Scholar] [CrossRef] [PubMed]
- Deeds, J.R.; Landsberg, J.H.; Etheridge, S.M.; Pitcher, G.C.; Longan, S.W. Non-traditional vectors for paralytic shellfish poisoning. Mar. Drugs 2008, 6, 308–348. [Google Scholar] [CrossRef] [PubMed]
- Klemm, K.; Cembella, A.; Clarke, D.; Cusack, C.; Arneborg, L.; Karlson, B.; Liu, Y.; Naustvoll, L.; Siano, R.; Gran-Stadniczenko, S.; et al. Apparent biogeographical trends in Alexandrium blooms for northern Europe: Identifying links to climate change and effective adaptive actions. Harmful Algae 2022, 119, 102335. [Google Scholar] [CrossRef] [PubMed]
- Schweibold, L. Global Status of Harmful Algal Blooms: First Steps of a Metadata Approach; ResearchGate: Berlin, Germany, 2017. [Google Scholar] [CrossRef]
- Liu, M.; Zheng, J.; Krock, B.; Ding, G.; MacKenzie, L.; Smith, K.F.; Gu, H. Dynamics of the Toxic Dinoflagellate Alexandrium pacificum in the Taiwan Strait and Its Linkages to Surrounding Populations. Water 2021, 13, 2681. [Google Scholar] [CrossRef]
- Selina, M.; Konovalova, G.; Morozova, T.; Orlova, T.Y. Genus Alexandrium Halim, 1960 (Dinophyta) from the Pacific coast of Russia: Species composition, distribution, and dynamics. Russ. J. Mar. Biol. 2006, 32, 321–332. [Google Scholar] [CrossRef]
- Anderson, D.M.; Fachon, E.; Pickart, R.S.; Lin, P.; Fischer, A.D.; Richlen, M.L.; Uva, V.; Brosnahan, M.L.; McRaven, L.; Bahr, F.; et al. Evidence for massive and recurrent toxic blooms of Alexandrium catenella in the Alaskan Arctic. Proc. Natl. Acad. Sci. USA 2021, 118, e2107387118. [Google Scholar] [CrossRef] [PubMed]
- Egmond, H.P.; Van Apeldoorn, M. Marine Biotoxins; Food & Agriculture Organization: Rome, Italy, 2004. [Google Scholar]
- Hamano, Y.; Kinoshita, Y.; Yasumoto, T. Enteropathogenicity of Diarrhetic Shellfish Toxins in Intestinal Models Studies on Diarrhetic Shellfish Toxins. I. Food Hyg. Saf. Sci. (Shokuhin Eiseigaku Zasshi) 1986, 27, 375–379_1. [Google Scholar] [CrossRef]
- FAO. Toxicity Equivalence Factors for Marine Biotoxins Associated with Bivalve Molluscs; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- WHO. Aquatic (Marine and Freshwater) Biotoxins-Environmental Health Criteria 37; WHO: Geneva, Switzerland, 1984. [Google Scholar]
- Reboreda, A.; Lago, J.; Chapela, M.J.; Vieites, J.M.; Botana, L.M.; Alfonso, A.; Cabado, A.G. Decrease of marine toxin content in bivalves by industrial processes. Toxicon 2010, 55, 235–243. [Google Scholar] [CrossRef]
- Dounay, A.B.; Forsyth, C.J. Okadaic acid: The archetypal serine/threonine protein phosphatase inhibitor. Curr. Med. Chem. 2002, 9, 1939–1980. [Google Scholar] [CrossRef]
- Takai, A.; Murata, M.; Torigoe, K.; Isobe, M.; Mieskes, G.; Yasumoto, T. Inhibitory effect of okadaic acid derivatives on protein phosphatases. A study on structure-affinity relationship. Biochem. J. 1992, 284 Pt 2, 539–544. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.Y.; Lin, L.; Gao, Y.; Hong, H.S.; Wang, D.Z. Quantitative proteomic analysis of okadaic acid treated mouse small intestines reveals differentially expressed proteins involved in diarrhetic shellfish poisoning. J. Proteomics 2012, 75, 2038–2052. [Google Scholar] [CrossRef] [PubMed]
- Falconer, I.R. Algal toxins in seafood and drinking water; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Heil, C.A.; Glibert, P.M.; Fan, C. Prorocentrum minimum (Pavillard) Schiller. Harmful Algae 2005, 4, 449–470. [Google Scholar] [CrossRef]
- Edwards, M.; Johns, D.; Leterme, S.; Svendsen, E.; Richardson, A. Regional climate change and harmful algal blooms in the northeast Atlantic. Limnol. Oceanogr. 2006, 51, 820–829. [Google Scholar] [CrossRef]
- Bates, S.S.; Hubbard, K.A.; Lundholm, N.; Montresor, M.; Leaw, C.P. Pseudo-nitzschia, Nitzschia, and domoic acid: New research since 2011. Harmful Algae 2018, 79, 3–43. [Google Scholar] [CrossRef] [PubMed]
- Steele, T.S.; Brunson, J.K.; Maeno, Y.; Terada, R.; Allen, A.E.; Yotsu-Yamashita, M.; Chekan, J.R.; Moore, B.S. Domoic acid biosynthesis in the red alga Chondria armata suggests a complex evolutionary history for toxin production. Proc. Natl. Acad. Sci. USA 2022, 119, e2117407119. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.; Boyd, R.; Freitas, A.d.; Falk, M.; Foxall, R.; Jamieson, W.; Laycock, M.; McCulloch, A.; McInnes, A.; Odense, P. Identification of domoic acid, a neuroexcitatory amino acid, in toxic mussels from eastern Prince Edward Island. Can. J. Chem. 1989, 67, 481–490. [Google Scholar] [CrossRef]
- de la Iglesia, P.; Gimenez, G.; Diogene, J. Determination of dissolved domoic acid in seawater with reversed-phase extraction disks and rapid resolution liquid chromatography tandem mass spectrometry with head-column trapping. J. Chromatogr. A 2008, 1215, 116–124. [Google Scholar] [CrossRef]
- Berman, F.W.; Murray, T.F. Domoic acid neurotoxicity in cultured cerebellar granule neurons is mediated predominantly by NMDA receptors that are activated as a consequence of excitatory amino acid release. J. Neurochem. 1997, 69, 693–703. [Google Scholar] [CrossRef]
- Larm, J.A.; Beart, P.M.; Cheung, N.S. Neurotoxin domoic acid produces cytotoxicity via kainate- and AMPA-sensitive receptors in cultured cortical neurones. Neurochem. Int. 1997, 31, 677–682. [Google Scholar] [CrossRef]
- Pinheiro, P.; Mulle, C. Kainate receptors. Cell Tissue Res. 2006, 326, 457–482. [Google Scholar] [CrossRef]
- Paoletti, P.; Neyton, J. NMDA receptor subunits: Function and pharmacology. Curr. Opin. Pharmacol. 2007, 7, 39–47. [Google Scholar] [CrossRef]
- Carcache, L.M.; Rodriguez, J.; Rein, K.S. The structural basis for kainoid selectivity at AMPA receptors revealed by low-mode docking calculations. Bioorganic Med. Chem. 2003, 11, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Hampson, D.R.; Manalo, J.L. The activation of glutamate receptors by kainic acid and domoic acid. Nat. Toxins 1998, 6, 153–158. [Google Scholar] [CrossRef]
- Colman, J.R.; Nowocin, K.J.; Switzer, R.C.; Trusk, T.C.; Ramsdell, J.S. Mapping and reconstruction of domoic acid-induced neurodegeneration in the mouse brain. Neurotoxicol. Teratol. 2005, 27, 753–767. [Google Scholar] [CrossRef]
- Giordano, G.; Klintworth, H.M.; Kavanagh, T.J.; Costa, L.G. Apoptosis induced by domoic acid in mouse cerebellar granule neurons involves activation of p38 and JNK MAP kinases. Neurochem. Int. 2008, 52, 1100–1105. [Google Scholar] [CrossRef]
- Pulido, O.M. Domoic acid toxicologic pathology: A review. Mar. Drugs 2008, 6, 180–219. [Google Scholar] [CrossRef] [PubMed]
- Perl, T.M.; Bedard, L.; Kosatsky, T.; Hockin, J.C.; Todd, E.C.; Remis, R.S. An outbreak of toxic encephalopathy caused by eating mussels contaminated with domoic acid. N. Engl. J. Med 1990, 322, 1775–1780. [Google Scholar] [CrossRef] [PubMed]
- Munday, R.; Holland, P.T.; McNabb, P.; Selwood, A.I.; Rhodes, L.L. Comparative toxicity to mice of domoic acid and isodomoic acids A, B and C. Toxicon 2008, 52, 954–956. [Google Scholar] [CrossRef] [PubMed]
- Sawant, P.M.; Tyndall, J.D.; Holland, P.T.; Peake, B.M.; Mountfort, D.O.; Kerr, D.S. In vivo seizure induction and affinity studies of domoic acid and isodomoic acids-D, -E and -F. Neuropharmacology 2010, 59, 129–138. [Google Scholar] [CrossRef]
- Sawant, P.M.; Weare, B.A.; Holland, P.T.; Selwood, A.I.; King, K.L.; Mikulski, C.M.; Doucette, G.J.; Mountfort, D.O.; Kerr, D.S. Isodomoic acids A and C exhibit low KA receptor affinity and reduced in vitro potency relative to domoic acid in region CA1 of rat hippocampus. Toxicon 2007, 50, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Silver, M.W.; Bargu, S.; Coale, S.L.; Benitez-Nelson, C.R.; Garcia, A.C.; Roberts, K.J.; Sekula-Wood, E.; Bruland, K.W.; Coale, K.H. Toxic diatoms and domoic acid in natural and iron enriched waters of the oceanic Pacific. Proc. Natl. Acad. Sci. USA 2010, 107, 20762–20767. [Google Scholar] [CrossRef]
- Landsberg, J.H.; Flewelling, L.J.; Naar, J. Karenia brevis red tides, brevetoxins in the food web, and impacts on natural resources: Decadal advancements. Harmful Algae 2009, 8, 598–607. [Google Scholar] [CrossRef]
- Bourdelais, A.J.; Tomas, C.R.; Naar, J.; Kubanek, J.; Baden, D.G. New fish-killing alga in coastal Delaware produces neurotoxins. Environ. Health Perspect. 2002, 110, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Baden, D.G.; Bourdelais, A.J.; Jacocks, H.; Michelliza, S.; Naar, J. Natural and derivative brevetoxins: Historical background, multiplicity, and effects. Environ. Health Perspect. 2005, 113, 621–625. [Google Scholar] [CrossRef]
- Ramsdell, J. The Molecular and Integrative Basis to Brevetoxin Toxicity. In Seafood and Freshwater Toxins; Food Science and Technology; Taylor & Francis Group: Boca Raton, FL, USA, 2008; pp. 519–550. [Google Scholar]
- Konoki, K.; Baden, D.G.; Scheuer, T.; Catterall, W.A. Molecular Determinants of Brevetoxin Binding to Voltage-Gated Sodium Channels. Toxins 2019, 11, 513. [Google Scholar] [CrossRef]
- Morris, P.D.; Campbell, D.S.; Taylor, T.J.; Freeman, J.I. Clinical and epidemiological features of neurotoxic shellfish poisoning in North Carolina. Am. J. Public Health 1991, 81, 471–474. [Google Scholar] [CrossRef]
- Steidinger, K.A. Historical perspective on Karenia brevis red tide research in the Gulf of Mexico. Harmful Algae 2009, 8, 549–561. [Google Scholar] [CrossRef]
- Watkins, S.M.; Reich, A.; Fleming, L.E.; Hammond, R. Neurotoxic shellfish poisoning. Mar. Drugs 2008, 6, 431–455. [Google Scholar] [CrossRef]
- Tillmann, U. Amphidomataceae. In Harmful Algae Blooms, a Compendium Desk Reference; Wiley-Blackwell Publishing: Chichester, West Sussex, 2018; pp. 575–582. [Google Scholar]
- Satake, M.; Ofuji, K.; Naoki, H.; James, K.J.; Furey, A.; McMahon, T.; Silke, J.; Yasumoto, T. Azaspiracid, a new marine toxin having unique spiro ring assemblies, isolated from Irish mussels, Mytilus edulis. J. Am. Chem. Soc. 1998, 120, 9967–9968. [Google Scholar] [CrossRef]
- Krock, B.; Tillmann, U.; Tebben, J.; Trefault, N.; Gu, H. Two novel azaspiracids from Azadinium poporum, and a comprehensive compilation of azaspiracids produced by Amphidomataceae, (Dinophyceae). Harmful Algae 2019, 82, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Twiner, M.J.; Rehmann, N.; Hess, P.; Doucette, G.J. Azaspiracid shellfish poisoning: A review on the chemistry, ecology, and toxicology with an emphasis on human health impacts. Mar. Drugs 2008, 6, 39–72. [Google Scholar] [CrossRef] [PubMed]
- Furey, A.; O’Doherty, S.; O’Callaghan, K.; Lehane, M.; James, K.J. Azaspiracid poisoning (AZP) toxins in shellfish: Toxicological and health considerations. Toxicon 2010, 56, 173–190. [Google Scholar] [CrossRef] [PubMed]
- Twiner, M.J.; Doucette, G.J.; Rasky, A.; Huang, X.P.; Roth, B.L.; Sanguinetti, M.C. Marine algal toxin azaspiracid is an open-state blocker of hERG potassium channels. Chem. Res. Toxicol. 2012, 25, 1975–1984. [Google Scholar] [CrossRef]
- Tillmann, U.; Edvardsen, B.; Krock, B.; Smith, K.F.; Paterson, R.F.; Voss, D. Diversity, distribution, and azaspiracids of Amphidomataceae (Dinophyceae) along the Norwegian coast. Harmful Algae 2018, 80, 15–34. [Google Scholar] [CrossRef]
- McGirr, S.; Clarke, D.; Kilcoyne, J.; Silke, J.; Touzet, N. Co-localisation of Azaspiracid Analogs with the Dinoflagellate Species Azadinium spinosum and Amphidoma languida in the Southwest of Ireland. Microb. Ecol. 2022, 83, 635–646. [Google Scholar] [CrossRef]
- Akselman, R.; Negri, R.M. Blooms of Azadinium cf. spinosum Elbrächter et Tillmann (Dinophyceae) in northern shelf waters of Argentina, Southwestern Atlantic. Harmful Algae 2012, 19, 30–38. [Google Scholar] [CrossRef]
- Percopo, I.; Siano, R.; Rossi, R.; Soprano, V.; Sarno, D.; Zingone, A. A new potentially toxic Azadinium species (Dinophyceae) from the Mediterranean Sea, A. dexteroporum sp. nov. J. Phycol. 2013, 49, 950–966. [Google Scholar] [CrossRef]
- Gu, H.; Luo, Z.; Krock, B.; Witt, M.; Tillmann, U. Morphology, phylogeny and azaspiracid profile of Azadinium poporum (Dinophyceae) from the China Sea. Harmful Algae 2013, 21–22, 64–75. [Google Scholar] [CrossRef]
- Kim, J.H.; Tillmann, U.; Adams, N.G.; Krock, B.; Stutts, W.L.; Deeds, J.R.; Han, M.S.; Trainer, V.L. Identification of Azadinium species and a new azaspiracid from Azadinium poporum in Puget Sound, Washington State, USA. Harmful Algae 2017, 68, 152–167. [Google Scholar] [CrossRef]
- Tillmann, U.; Trefault, N.; Krock, B.; Parada-Pozo, G.; De la Iglesia, R.; Vásquez, M. Identification ofAzadinium poporum(Dinophyceae) in the Southeast Pacific: Morphology, molecular phylogeny, and azaspiracid profile characterization. J. Plankton Res. 2017, 39, 350–367. [Google Scholar] [CrossRef]
- Tillmann, U.; Jaen, D.; Fernandez, L.; Gottschling, M.; Witt, M.; Blanco, J.; Krock, B. Amphidoma languida (Amphidomatacea, Dinophyceae) with a novel azaspiracid toxin profile identified as the cause of molluscan contamination at the Atlantic coast of southern Spain. Harmful Algae 2017, 62, 113–126. [Google Scholar] [CrossRef]
- Wietkamp, S.; Tillmann, U.; Clarke, D.; Toebe, K. Molecular detection and quantification of the azaspiracid-producing dinoflagellate Amphidoma languida (Amphidomataceae, Dinophyceae). J. Plankton Res. 2019, 41, 101–113. [Google Scholar] [CrossRef]
- Keeling, C.D.; Piper, S.C.; Bacastow, R.B.; Wahlen, M.; Whorf, T.P.; Heimann, M.; Meijer, H.A. Exchanges of Atmospheric CO2 and 13CO2 with the Terrestrial Biosphere and Oceans from 1978 to 2000. I. Global Aspects; Springer: New York, NY, USA, 2001. [Google Scholar]
- IPCC. Climate Change 2014: Synthesis Report; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Sabine, C.L.; Feely, R.A.; Gruber, N.; Key, R.M.; Lee, K.; Bullister, J.L.; Wanninkhof, R.; Wong, C.S.; Wallace, D.W.; Tilbrook, B.; et al. The oceanic sink for anthropogenic CO2. Science 2004, 305, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Kroeker, K.J.; Kordas, R.L.; Crim, R.N.; Singh, G.G. Meta-analysis reveals negative yet variable effects of ocean acidification on marine organisms. Ecol. Lett. 2010, 13, 1419–1434. [Google Scholar] [CrossRef]
- Al Jabri, H.; Taleb, A.; Touchard, R.; Saadaoui, I.; Goetz, V.; Pruvost, J. Cultivating Microalgae in Desert Conditions: Evaluation of the Effect of Light-Temperature Summer Conditions on the Growth and Metabolism of Nannochloropsis QU130. Appl. Sci. 2021, 11, 3799. [Google Scholar] [CrossRef]
- Procházková, G.; Brányiková, I.; Zachleder, V.; Brányik, T. Effect of nutrient supply status on biomass composition of eukaryotic green microalgae. J. Appl. Phycol. 2013, 26, 1359–1377. [Google Scholar] [CrossRef]
- Brand, L.E.; Campbell, L.; Bresnan, E. Karenia: The biology and ecology of a toxic genus. Harmful Algae 2012, 14, 156–178. [Google Scholar] [CrossRef]
- Shearn-Bochsler, V.; Lance, E.W.; Corcoran, R.; Piatt, J.; Bodenstein, B.; Frame, E.; Lawonn, J. Fatal paralytic shellfish poisoning in Kittlitz’s Murrelet (Brachyramphus brevirostris) nestlings, Alaska, USA. J. Wildl. Dis. 2014, 50, 933–937. [Google Scholar] [CrossRef]
- Garcia, C.; Perez, F.; Contreras, C.; Figueroa, D.; Barriga, A.; Lopez-Rivera, A.; Araneda, O.F.; Contreras, H.R. Saxitoxins and okadaic acid group: Accumulation and distribution in invertebrate marine vectors from Southern Chile. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess 2015, 32, 984–1002. [Google Scholar] [CrossRef] [PubMed]
- Bejarano, A.C.; VanDola, F.M.; Gulland, F.M.; Rowles, T.K.; Schwacke, L.H. Production and toxicity of the marine biotoxin domoic acid and its effects on wildlife: A review. Hum. Ecol. Risk Assess. 2008, 14, 544–567. [Google Scholar] [CrossRef]
- Burkholder, J.M.; Shumway, S.E.; Glibert, P.M. Food web and ecosystem impacts of harmful algae. Harmful Algal Bloom. A Compend. Desk Ref. 2018, 243–336. [Google Scholar]
- Roggatz, C.C.; Fletcher, N.; Benoit, D.M.; Algar, A.C.; Doroff, A.; Wright, B.; Wollenberg Valero, K.C.; Hardege, J.D. Saxitoxin and tetrodotoxin bioavailability increases in future oceans. Nat. Clim. Change 2019, 9, 840–844. [Google Scholar] [CrossRef]
- Hattenrath-Lehmann, T.K.; Smith, J.L.; Wallace, R.B.; Merlo, L.; Koch, F.; Mittelsdorf, H.; Goleski, J.A.; Anderson, D.M.; Gobler, C.J. The effects of elevated CO(2) on the growth and toxicity of field populations and cultures of the saxitoxin-producing dinoflagellate, Alexandrium fundyense. Limnol Oceanogr 2015, 60, 198–214. [Google Scholar] [CrossRef] [PubMed]
- Lian, Z.; Li, F.; He, X.; Chen, J.; Yu, R.-C. Rising CO2 will increase toxicity of marine dinoflagellate Alexandrium minutum. J. Hazard. Mater. 2022, 431, 128627. [Google Scholar] [CrossRef]
- Tatters, A.O.; Flewelling, L.J.; Fu, F.; Granholm, A.A.; Hutchins, D.A. High CO2 promotes the production of paralytic shellfish poisoning toxins by Alexandrium catenella from Southern California waters. Harmful Algae 2013, 30, 37–43. [Google Scholar] [CrossRef]
- Brandenburg, K.M.; Krock, B.; Klip, H.C.L.; Sluijs, A.; Garbeva, P.; Van de Waal, D.B. Intraspecific variation in multiple trait responses of Alexandrium ostenfeldii towards elevated pCO(2). Harmful Algae 2021, 101, 101970. [Google Scholar] [CrossRef] [PubMed]
- Van de Waal, D.B.; Eberlein, T.; John, U.; Wohlrab, S.; Rost, B. Impact of elevated pCO(2) on paralytic shellfish poisoning toxin content and composition in Alexandrium tamarense. Toxicon 2014, 78, 58–67. [Google Scholar] [CrossRef]
- Sun, J.; Hutchins, D.A.; Feng, Y.; Seubert, E.L.; Caron, D.A.; Fu, F.-X. Effects of changing pCO2 and phosphate availability on domoic acid production and physiology of the marine harmful bloom diatom Pseudo-nitzschia multiseries. Limnol. Oceanogr. 2011, 56, 829–840. [Google Scholar] [CrossRef]
- Tatters, A.O.; Fu, F.X.; Hutchins, D.A. High CO2 and silicate limitation synergistically increase the toxicity of Pseudo-nitzschia fraudulenta. PLoS ONE 2012, 7, e32116. [Google Scholar] [CrossRef]
- Wingert, C.J.; Cochlan, W.P. Effects of ocean acidification on the growth, photosynthetic performance, and domoic acid production of the diatom Pseudo-nitzschia australis from the California Current System. Harmful Algae 2021, 107, 102030. [Google Scholar] [CrossRef]
- Wohlrab, S.; John, U.; Klemm, K.; Eberlein, T.; Forsberg Grivogiannis, A.M.; Krock, B.; Frickenhaus, S.; Bach, L.T.; Rost, B.; Riebesell, U.; et al. Ocean acidification increases domoic acid contents during a spring to summer succession of coastal phytoplankton. Harmful Algae 2020, 92, 101697. [Google Scholar] [CrossRef] [PubMed]
- Bercel, T.L.; Kranz, S.A. Insights into carbon acquisition and photosynthesis in Karenia brevis under a range of CO2 concentrations. Prog. Oceanogr. 2019, 172, 65–76. [Google Scholar] [CrossRef]
- Errera, R.M.; Yvon-Lewis, S.; Kessler, J.D.; Campbell, L. Reponses of the dinoflagellate Karenia brevis to climate change: pCO2 and sea surface temperatures. Harmful Algae 2014, 37, 110–116. [Google Scholar] [CrossRef]
- Raven, J.A.; Cockell, C.S.; De La Rocha, C.L. The evolution of inorganic carbon concentrating mechanisms in photosynthesis. Philos. Trans. R. Soc. Lond B. Biol. Sci. 2008, 363, 2641–2650. [Google Scholar] [CrossRef] [PubMed]
- Tortell, P.D. Evolutionary and ecological perspectives on carbon acquisition in phytoplankton. Limnol. Oceanogr. 2000, 45, 744–750. [Google Scholar] [CrossRef]
- Kellmann, R.; Mihali, T.K.; Jeon, Y.J.; Pickford, R.; Pomati, F.; Neilan, B.A. Biosynthetic intermediate analysis and functional homology reveal a saxitoxin gene cluster in cyanobacteria. Appl. Environ. Microbiol. 2008, 74, 4044–4053. [Google Scholar] [CrossRef]
- Taroncher-Oldenburg, G.; Anderson, D.M. Identification and characterization of three differentially expressed genes, encoding S-adenosylhomocysteine hydrolase, methionine aminopeptidase, and a histone-like protein, in the toxic dinoflagellate Alexandrium fundyense. Appl. Environ. Microbiol. 2000, 66, 2105–2112. [Google Scholar] [CrossRef]
- Pang, M.; Xu, J.; Qu, P.; Mao, X.; Wu, Z.; Xin, M.; Sun, P.; Wang, Z.; Zhang, X.; Chen, H. Effect of CO(2) on growth and toxicity of Alexandrium tamarense from the East China Sea, a major producer of paralytic shellfish toxins. Harmful Algae 2017, 68, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Hackett, J.D.; Wisecaver, J.H.; Brosnahan, M.L.; Kulis, D.M.; Anderson, D.M.; Bhattacharya, D.; Plumley, F.G.; Erdner, D.L. Evolution of saxitoxin synthesis in cyanobacteria and dinoflagellates. Mol. Biol. Evol. 2013, 30, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, A.; Loram, J.E.; Hackett, J.D.; Anderson, D.M.; Plumley, F.G.; Bhattacharya, D. Origin of saxitoxin biosynthetic genes in cyanobacteria. PLoS ONE 2009, 4, e5758. [Google Scholar] [CrossRef] [PubMed]
- Sako, Y.; Yoshida, T.; Uchida, A.; Arakawa, O.; Noguchi, T.; Ishida, Y. Purification and characterization of a sulfotransferase specific to N-21 of saxitoxin and gonyautoxin 2+ 3 from the toxic dinoflagellate Gymnodinium catenatum (Dinophyceae). J. Phycol. 2001, 37, 1044–1051. [Google Scholar] [CrossRef]
- Soto-Liebe, K.; Murillo, A.A.; Krock, B.; Stucken, K.; Fuentes-Valdes, J.J.; Trefault, N.; Cembella, A.; Vasquez, M. Reassessment of the toxin profile of Cylindrospermopsis raciborskii T3 and function of putative sulfotransferases in synthesis of sulfated and sulfonated PSP toxins. Toxicon 2010, 56, 1350–1361. [Google Scholar] [CrossRef]
- Giordano, M.; Norici, A.; Hell, R. Sulfur and phytoplankton: Acquisition, metabolism and impact on the environment. New Phytol. 2005, 166, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Shibagaki, N.; Grossman, A. The state of sulfur metabolism in algae: From ecology to genomics. In Sulfur Metabolism in Phototrophic Organisms; Springer: Berlin/Heidelberg, Germany, 2008; pp. 231–267. [Google Scholar]
- Lundholm, N.; Hansen, P.J.; Kotaki, Y. Effect of pH on growth and domoic acid production by potentially toxic diatoms of the genera Pseudo-nitzschia and Nitzschia. Mar. Ecol. Prog. Ser. 2004, 273, 1–15. [Google Scholar] [CrossRef]
- Shi, D.; Hong, H.; Su, X.; Liao, L.; Chang, S.; Lin, W. The physiological response of marine diatoms to ocean acidification: Differential roles of seawater pCO(2) and pH. J. Phycol. 2019, 55, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Trimborn, S.; Lundholm, N.; Thoms, S.; Richter, K.U.; Krock, B.; Hansen, P.J.; Rost, B. Inorganic carbon acquisition in potentially toxic and non-toxic diatoms: The effect of pH-induced changes in seawater carbonate chemistry. Physiol. Plant. 2008, 133, 92–105. [Google Scholar] [CrossRef]
- Brunson, J.K.; McKinnie, S.M.K.; Chekan, J.R.; McCrow, J.P.; Miles, Z.D.; Bertrand, E.M.; Bielinski, V.A.; Luhavaya, H.; Obornik, M.; Smith, G.J.; et al. Biosynthesis of the neurotoxin domoic acid in a bloom-forming diatom. Science 2018, 361, 1356–1358. [Google Scholar] [CrossRef]
- MillerO, F.J.; Woosley, R.; DiTrolio, B.; Waters, J. Effect of ocean acidification on the speciation of metals in seawater. Oceanography 2009, 22, 72–85. [Google Scholar] [CrossRef]
- Perez, P.; Beiras, R.; Fernandez, E. Monitoring copper toxicity in natural phytoplankton assemblages: Application of Fast Repetition Rate fluorometry. Ecotoxicol. Environ. Saf. 2010, 73, 1292–1303. [Google Scholar] [CrossRef]
- Rue, E.; Bruland, K. Domoic acid binds iron and copper: A possible role for the toxin produced by the marine diatom Pseudo-nitzschia. Mar. Chem. 2001, 76, 127–134. [Google Scholar] [CrossRef]
- Lelong, A.; Jolley, D.F.; Soudant, P.; Hegaret, H. Impact of copper exposure on Pseudo-nitzschia spp. physiology and domoic acid production. Aquat. Toxicol. 2012, 118–119, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Bates, S.S.; Léger, C.; Satchwell, M.; Boyer, G.L. The effects of iron on domoic acid production by Pseudo-nitzschia multiseries. In Proceedings of the 9th International Conference on Harmful Algal Blooms, Hobart, Tasmania, 7–11 February 2000; pp. 320–323. [Google Scholar]
- Kobayashi, K.; Takata, Y.; Kodama, M. Direct contact between Pseudo-nitzschia multiseries and bacteria is necessary for the diatom to produce a high level of domoic acid. Fish. Sci. 2009, 75, 771–776. [Google Scholar] [CrossRef]
- Bates, S.S.; Douglas, D.J.; Doucette, G.J.; Leger, C. Enhancement of domoic acid production by reintroducing bacteria to axenic cultures of the diatom Pseudo-nitzschia multiseries. Nat. Toxins 1995, 3, 428–435. [Google Scholar] [CrossRef]
- Bates, S.S.; Gaudet, J.; Kaczmarska, I.; Ehrman, J.M. Interaction between bacteria and the domoic-acid-producing diatom Pseudo-nitzschia multiseries (Hasle) Hasle; can bacteria produce domoic acid autonomously? Harmful Algae 2004, 3, 11–20. [Google Scholar] [CrossRef]
- Maas, E.W.; Law, C.S.; Hall, J.A.; Pickmere, S.; Currie, K.I.; Chang, F.; Voyles, K.M.; Caird, D. Effect of ocean acidification on bacterial abundance, activity and diversity in the Ross Sea, Antarctica. Aquat. Microb. Ecol. 2013, 70, 1–15. [Google Scholar] [CrossRef]
- Schada von Borzyskowski, L.; Bernhardsgrutter, I.; Erb, T.J. Biochemical unity revisited: Microbial central carbon metabolism holds new discoveries, multi-tasking pathways, and redundancies with a reason. Biol. Chem. 2020, 401, 1429–1441. [Google Scholar] [CrossRef]
- Dickson, A.G. The carbon dioxide system in seawater: Equilibrium chemistry and measurements. Guide Best Pract. Ocean. Acidif. Res. Data Report. 2010, 1, 17–40. [Google Scholar]
- Gattuso, J.-P.; Lavigne, H. Approaches and software tools to investigate the impact of ocean acidification. Biogeosciences 2009, 6, 2121–2133. [Google Scholar] [CrossRef]
- Gattuso, J.-P.; Lee, K.; Rost, B.; Schulz, K. Approaches and Tools to Manipulate the Carbonate Chemistry. Publications Office of the European Union: Luxembourg, 2010. [Google Scholar]
- Llaveria, G.; Figueroa, R.I.; Garces, E.; Berdalet, E. Cell Cycle and Cell Mortality of Alexandrium Minutum (Dinophyceae) under Small-Scale Turbulence Conditions 1. J. Phycol. 2009, 45, 1106–1115. [Google Scholar] [CrossRef]
- Shi, D.; Xu, Y.; Morel, F. Effects of the pH/pCO 2 control method on medium chemistry and phytoplankton growth. Biogeosciences 2009, 6, 1199–1207. [Google Scholar] [CrossRef]
- Hurd, C.L.; Hepburn, C.D.; Currie, K.I.; Raven, J.A.; Hunter, K.A. Testing the effects of ocean acidification on algal metabolism: Considerations for experimental designs1. J. Phycol. 2009, 45, 1236–1251. [Google Scholar] [CrossRef]
- Rost, B.; Zondervan, I.; Wolf-Gladrow, D. Sensitivity of phytoplankton to future changes in ocean carbonate chemistry: Current knowledge, contradictions and research directions. Mar. Ecol. Prog. Ser. 2008, 373, 227–237. [Google Scholar] [CrossRef]
- Cornwall, C.E.; Hurd, C.L. Experimental design in ocean acidification research: Problems and solutions. ICES J. Mar. Sci. 2016, 73, 572–581. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics—The link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Tsuchiya, S.; Omura, T.; Koike, K.; Oikawa, H.; Konoki, K.; Oshima, Y.; Yotsu-Yamashita, M. Metabolomic study of saxitoxin analogues and biosynthetic intermediates in dinoflagellates using (15)N-labelled sodium nitrate as a nitrogen source. Sci. Rep. 2019, 9, 3460. [Google Scholar] [CrossRef]
- Brown, E.R.; Moore, S.G.; Gaul, D.A.; Kubanek, J. Predator cues target signaling pathways in toxic algal metabolome. Limnol. Oceanogr. 2022, 67, 1227–1237. [Google Scholar] [CrossRef]
- Jang, C.; Chen, L.; Rabinowitz, J.D. Metabolomics and Isotope Tracing. Cell 2018, 173, 822–837. [Google Scholar] [CrossRef]
- Rahim, M.; Ragavan, M.; Deja, S.; Merritt, M.E.; Burgess, S.C.; Young, J.D. INCA 2.0: A tool for integrated, dynamic modeling of NMR- and MS-based isotopomer measurements and rigorous metabolic flux analysis. Metab. Eng. 2022, 69, 275–285. [Google Scholar] [CrossRef]
- Wang, Y.; Wondisford, F.E.; Song, C.; Zhang, T.; Su, X. Metabolic Flux Analysis-Linking Isotope Labeling and Metabolic Fluxes. Metabolites 2020, 10, 447. [Google Scholar] [CrossRef]
- Treves, H.; Kuken, A.; Arrivault, S.; Ishihara, H.; Hoppe, I.; Erban, A.; Hohne, M.; Moraes, T.A.; Kopka, J.; Szymanski, J.; et al. Carbon flux through photosynthesis and central carbon metabolism show distinct patterns between algae, C(3) and C(4) plants. Nat. Plants 2022, 8, 78–91. [Google Scholar] [CrossRef]
- Gimenez, I.; Waldbusser, G.G.; Langdon, C.J.; Hales, B.R. The dynamic ocean acidification manipulation experimental system: Separating carbonate variables and simulating natural variability in laboratory flow-through experiments. Limnol. Oceanogr. Methods 2019, 17, 343–361. [Google Scholar] [CrossRef]
- West, J.J.; Järnberg, L.; Berdalet, E.; Cusack, C. Understanding and Managing Harmful Algal Bloom Risks in a Changing Climate: Lessons From the European CoCliME Project. Front. Clim. 2021, 3. [Google Scholar] [CrossRef]
- Kremp, A.; Godhe, A.; Egardt, J.; Dupont, S.; Suikkanen, S.; Casabianca, S.; Penna, A. Intraspecific variability in the response of bloom-forming marine microalgae to changed climate conditions. Ecol. Evol. 2012, 2, 1195–1207. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.M.; Andersen, P.; Bricelj, V.M.; Cullen, J.J.; Rensel, J.J. Monitoring and Management Strategies for Harmful Algal Blooms in Coastal Waters; UNESCO: Paris, France, 2001. [Google Scholar]
- Anderson, C.R.; Kudela, R.M.; Kahru, M.; Chao, Y.; Rosenfeld, L.K.; Bahr, F.L.; Anderson, D.M.; Norris, T.A. Initial skill assessment of the California Harmful Algae Risk Mapping (C-HARM) system. Harmful Algae 2016, 59, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Davidson, K.; Anderson, D.M.; Mateus, M.; Reguera, B.; Silke, J.; Sourisseau, M.; Maguire, J. Forecasting the risk of harmful algal blooms. Harmful Algae 2016, 53, 1–7. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Algal Species (Strains) | OA Modeling Method | Experimental Parameters a | Significant Outcomes b | Reference | |
|---|---|---|---|---|---|
| Control | High pCO2 Treatment | ||||
| STXs-Producing Microalgae | |||||
| Alexandrium fundyense (NBP8) isolated from Northport Bay | Laboratory culture CO2 gas bubbling | Experiment No. 1 pH: 8.085 ± 0.009 AT: 1824 ± 87 μmol L−1 Calculated pCO2: 41 ± 2 Pa DIC: 1600 ± 80 μmol L−1 Length of experiment: 15 days | Experiment No. 1 pH: 7.629 ± 0.031 AT: 1678 ± 123 μmol L−1 Calculated pCO2: 122 ± 6 Pa DIC: 1606 ± 116 μmol L−1 Length of experiment: 15 days | Experiment No. 1 Growth rate increased * | [97] |
| Experiment No. 2 pH: 8.068 ± 0.013 AT: 1858 ± 104 μmol L−1 Calculated pCO2: 44 ± 4 Pa DIC: 1639 ± 100 μmol L−1 Length of experiment: 27 days | Experiment No. 2 pH: 7.741 ± 0.007 AT: 1975 ± 128 μmol L−1 Calculated pCO2: 110 ± 7 Pa DIC: 1868 ± 125 μmol L−1 Length of experiment: 27 days | Experiment No. 2 Total cellular toxicity increased * GTX1/4 increased * | [97] | ||
| Experiment No. 3 pH: 8.075 ± 0.057 AT: 1994 ± 355 μmol L−1 Calculated pCO2: 46 ± 2 Pa DIC: 1790 ± 329 μmol L−1 Length of experiment: 15 days | Experiment No. 3 pH: 7.545 ± 0.034 AT: 1966 ± 231 μmol L−1 Calculated pCO2: 177 ± 9 Pa DIC: 1941 ± 243 μmol L−1 Length of experiment: 15 days | Experiment No. 3 Growth rate increased * | [97] | ||
| Experiment No. 4 pH: 8.096 ± 0.032 AT: 2122 ± 123 μmol L−1 Calculated pCO2: 47 ± 2 Pa DIC: 1885 ± 102 μmol L−1 Length of experiment: 12 days | Experiment No. 4 pH: 7.592 ± 0.022 AT: 1977 ± 240 μmol L−1 Calculated pCO2: 162 ± 12 Pa DIC: 1977 ± 240 μmol L−1 Length of experiment: 12 days | Experiment No. 4 Growth rate increased * Total cellular toxicity increased * GTX1/4 increased * | [97] | ||
| Alexandrium fundyense (CCMP2304) isolated from the Bay of Fundy | Experiment No. 5 pH: 8.118 ± 0.008 AT: 2167 ± 53 μmol L−1 Calculated pCO2: 45 ± 1 Pa DIC: 1912 ± 44 μmol L−1 Length of experiment: 24 days | Experiment No. 5 pH: 7.873 ± 0.033 AT: 2186 ± 91 μmol L−1 Calculated pCO2: 87 ± 4 Pa DIC: 2033 ± 71 μmol L−1 Length of experiment: 24 days | Experiment No. 5 Growth rate: increased * | [97] | |
| Experiment No. 6 pH: 8.041 ± 0.012 AT: 1729 ± 56 μmol L−1 Calculated pCO2: 44 ± 1 Pa DIC: 1539 ± 49 μmol L−1 Length of experiment: 12 days | Experiment No. 6 pH: 7.547 ± 0.028 AT: 1889 ± 90 μmol L−1 Calculated pCO2: 169 ± 4 Pa DIC: 1845 ± 84 μmol L−1 Length of experiment: 12 days | Experiment No. 6 Growth rate increased * | [97] | ||
| Experiment No. 7 pH: 8.086 ± 0.048 AT: 2121 ± 213 μmol L−1 Calculated pCO2: 48 ± 3 Pa DIC: 1888 ± 183 μmol L−1 Length of experiment: 12 days | Experiment No. 7 pH: 7.556 ± 0.038 AT: 2184 ± 167 μmol L−1 Calculated pCO2: 191 ± 4 Pa DIC: 2138 ± 157 μmol L−1 Length of experiment: 12 days | Experiment No. 7 Growth rate increased * | [97] | ||
| Alexandrium minutum (AM-1) isolated from the South China Sea | Laboratory culture CO2 gas bubbling | 400 ppm CO2 treatment Length of experiment: 37 days | 800 ppm CO2 treatment Length of experiment: 37 days | 400 vs. 800 ppm CO2 Total cellular toxicity increased ** | [98] |
| 400 ppm CO2 treatment Length of experiment: 37 days | 1200 ppm CO2 treatment Length of experiment: 37 days | 400 vs. 1200 ppm CO2 Total cellular toxicity increased ** | [98] | ||
| Alexandrium catenella (A-11c) isolated from Jalama Beach | Laboratory culture CO2 gas bubbling | 380 µatm CO2 treatment pH: 8.169 Calculated pCO2: 285 µatm DIC: 1957 μmol L−1 Length of experiment: 7 days | 750 µatm CO2 treatment pH: 7.925 Calculated pCO2: 571 µatm DIC: 2103 μmol L−1 Length of experiment: 7 days | 380 vs. 750 µatm CO2 Growth rate increased *** Total cellular toxicity increased *** | [99] |
| Alexandrium ostenfeldii (AON13) isolated from Ouwerkerkse Kreek | Laboratory culture CO2 gas bubbling prior to cell inoculation c | 400 µatm CO2 treatment pH: 8.23 ± 0.02 Calculated pCO2: 213 ± 11 µatm DIC: 1188 ± 41 μmol L−1 Length of experiment: 14 days | 1000 µatm CO2 treatment pH: 7.82 ± 0.01 Calculated pCO2: 527 ± 30 µatm DIC: 1168 ± 56 μmol L−1 Length of experiment: 14 days | 400 vs. 1000 µatm CO2 STX increased * | [100] |
| Alexandrium ostenfeldii (AON15) isolated from Ouwerkerkse Kreek | 400 µatm CO2 treatment pH: 8.07 ± 0.07 Calculated pCO2: 357 ± 36 µatm DIC: 1120 ± 21 μmol L−1 Length of experiment: 14 days | 1000 µatm CO2 treatment pH: 7.75 ± 0.08 Calculated pCO2: 676 ± 55 µatm DIC: 1117 ± 10 μmol L−1 Length of experiment: 14 days | 400 vs. 1000 µatm CO2 Growth rate increased ** Total cellular toxicity decreased *** GTX increased * | [100] | |
| Alexandrium ostenfeldii (AON5.26) isolated from Ouwerkerkse Kreek | 400 µatm CO2 treatment pH: 8.17 ± 0.05 Calculated pCO2: 286 ± 13 µatm DIC: 1085 ± 15 μmol L−1 Length of experiment: 14 days | 1000 µatm CO2 treatment pH: 7.86 ± 0.03 Calculated pCO2: 479 ± 18 µatm DIC: 1126 ± 2 μmol L−1 Length of experiment: 14 days | Growth rate increased *** GTX increased *** STX and C1/2 decreased *** | [100] | |
| Alexandrium tamarense (Alex5) | Laboratory culture CO2 gas bubbling | 380 µatm CO2 treatment pH: 8.27 ± 0.07 AT: 2439 ± 1 μmol L−1 Calculated pCO2: 315 ± 57 µatm DIC: 2117 ± 41 μmol L−1 Length of experiment: 8 days | 800 µatm CO2 treatment pH: 7.97 ± 0.10 AT: 2434 ± 2 μmol L−1 Calculated pCO2: 706 ± 154 µatm DIC: 2245 ± 37 μmol L−1 Length of experiment: 8 days | 380 vs. 800 µatm CO2 Total cellular toxicity decreased * Non-sulfated STX analogues decreased * Di-sulfated STX analogues increased * | [101] |
| 380 µatm CO2 treatment pH: 8.27 ± 0.07 AT: 2439 ± 1 μmol L−1 Calculated pCO2: 315 ± 57 µatm DIC: 2117 ± 41 μmol L−1 Length of experiment: 8 days | 1200 µatm CO2 treatment pH: 7.83 ± 0.12 AT: 2418 ± 1 μmol L−1 Calculated pCO2: 995 ± 248 µatm DIC: 2283 ± 34 μmol L−1 Length of experiment: 8 days | 380 vs. 1200 µatm CO2 Total cellular toxicity decreased * Non-sulfated STX analogues decreased * Mono-sulfated STX analogues increased * Di-sulfated STX analogues increased * | [101] | ||
| Alexandrium tamarense (Alex2) | 380 µatm CO2 treatment pH: 8.27 ± 0.02 AT: 2384 ± 4 μmol L−1 Calculated pCO2: 305 ± 20 µatm DIC: 2111 ± 14 μmol L−1 Length of experiment: 8 days | 800 µatm CO2 treatment pH: 7.90 ± 0.03 AT: 2390 ± 6 μmol L−1 Calculated pCO2: 810 ± 60 µatm DIC: 2229 ± 33 μmol L−1 Length of experiment: 8 days | 380 vs. 800 µatm CO2 No significant changes in growth rate, total cellular toxicity, and toxin profile | [101] | |
| 380 µatm CO2 treatment pH: 8.27 ± 0.02 AT: 2384 ± 4 μmol L−1 Calculated pCO2: 305 ± 20 µatm DIC: 2111 ± 14 μmol L−1 Length of experiment: 8 days | 1200 µatm CO2 treatment pH: 7.75 ± 0.04 AT: 2386 ± 9 μmol L−1 Calculated pCO2: 1167 ± 112 µatm DIC: 2279 ± 14 μmol L−1 Length of experiment: 8 days | 380 vs. 1200 µatm CO2 No significant changes in growth rate, total cellular toxicity, and toxin profile | [101] | ||
| DA-producing microalgae | |||||
| Pseudo-nitzschia multiseries (CCMP2708) isolated from Eastern Canada | Laboratory culture CO2 gas bubbling | 22 Pa CO2 treatment pH: 8.40 ± 0.03 Calculated pCO2: 22 ± 2 Pa DIC: 1970 ± 4 μmol L−1 Length of experiment: around 4 to 6 weeks | 41 Pa CO2 treatment pH: 8.19 ± 0.02 Calculated pCO2: 40 ± 3 Pa DIC: 2066 ± 11 μmol L−1 Length of experiment: around 4 to 6 weeks | 22 vs. 41 Pa CO2 Growth rate increased *** Carbon fixation rate increased ** | [102] |
| 22 Pa CO2 treatment pH: 8.40 ± 0.03 Calculated pCO2: 22 ± 2 Pa DIC: 1970 ± 4 μmol L−1 Length of experiment: around 4 to 6 weeks | 74 Pa CO2 treatment pH: 7.96 ± 0.01 Calculated pCO2: 73 ± 1 Pa DIC: 2177 ± 6 μmol L−1 Length of experiment: around 4 to 6 weeks | 22 vs. 74 Pa CO2 Growth rate increased *** Carbon fixation rate increased *** Cellular DA content increased *** | [102] | ||
| 41 Pa CO2 treatment pH: 8.19 ± 0.02 Calculated pCO2: 40 ± 3 Pa DIC: 2066 ± 11 μmol L−1 Length of experiment: around 4 to 6 weeks | 74 Pa CO2 treatment pH: 7.96 ± 0.01 Calculated pCO2: 73 ± 1 Pa DIC: 2177 ± 6 μmol L−1 Length of experiment: around 4 to 6 weeks | 41 vs. 74 Pa CO2 Growth rate increased *** Cellular DA content increased ** | [102] | ||
| Pseudo-nitzschia fraudulenta isolated from Ventura County | Laboratory culture CO2 gas bubbling | 200 ppm CO2 treatment pH: 8.43 Calculated pCO2: 198 ppm DIC: 1965 μmol L−1 | 360 ppm CO2 treatment pH: 8.23 Calculated pCO2: 357 ppm DIC: 2107 μmol L−1 | 200 vs. 360 ppm CO2 Growth rate increased *** Carbon fixation rate increased | [103] |
| 200 ppm CO2 treatment pH: 8.43 Calculated pCO2: 198 ppm DIC: 1965 μmol L−1 | 765 ppm CO2 treatment pH: 7.95 Calculated pCO2: 764 ppm DIC: 2249 μmol L−1 | 200 vs. 765 ppm CO2 Growth rate increased *** Carbon fixation rate increased | [103] | ||
| Pseudo-nitzschia australis (HAB 200) isolated from northern Monterey Bay | Laboratory culture CO2 gas bubbling | pH 8.1 treatment Length of experiment: 7 days Exponential growth phase pH: 8.14 ± 0.01 AT: 2232 ± 9 μmol L−1 Calculated pCO2: 406 ± 8 µatm DIC: 2032 ± 11 μmol L−1 Stationary growth phase pH: 8.14 ± 0.01 AT: 2239 ± 25 μmol L−1 Calculated pCO2: 410 ± 2 µatm DIC: 2052 ± 21 μmol L−1 | pH 8.0 treatment Length of experiment: 8 days Exponential growth phase pH: 8.03 ± 0.01 AT: 2248 ± 25 μmol L−1 Calculated pCO2: 550 ± 8 µatm DIC: 2093 ± 21 μmol L−1 Stationary growth phase pH: 8.04 ± 0.01 AT: 2254 ± 14 μmol L−1 Calculated pCO2: 537 ± 19 µatm DIC: 2107 ± 17 μmol L−1 | pH 8.1 vs. pH 8.0 No significant response | [104] |
| pH 8.1 treatment Length of experiment: 7 days Exponential growth phase pH: 8.14 ± 0.01 AT: 2232 ± 9 μmol L−1 Calculated pCO2: 406 ± 8 µatm DIC: 2032 ± 11 μmol L−1 Stationary growth phase pH: 8.14 ± 0.01 AT: 2239 ± 25 μmol L−1 Calculated pCO2: 410 ± 2 µatm DIC: 2052 ± 21 μmol L−1 | pH 7.9 treatment Length of experiment: 8 days Exponential growth phase pH: 7.93 ± 0.02 AT: 2255 ± 20 μmol L−1 Calculated pCO2: 719 ± 34 µatm DIC: 2136 ± 25 μmol L−1 Stationary growth phase pH: 7.93 ± 0.02 AT: 2279 ± 14 μmol L−1 Calculated pCO2: 732 ± 28 µatm DIC: 2175 ± 13 μmol L−1 | pH 8.1 vs. pH 7.9 Maximum carbon fixation rate increased ** | [104] | ||
| pH 8.0 treatment Length of experiment: 8 days Exponential growth phase pH: 8.03 ± 0.01 AT: 2248 ± 25 μmol L−1 Calculated pCO2: 550 ± 8 µatm DIC: 2093 ± 21 μmol L−1 Stationary growth phase pH: 8.04 ± 0.01 AT: 2254 ± 14 μmol L−1 Calculated pCO2: 537 ± 19 µatm DIC: 2107 ± 17 μmol L−1 | pH 7.9 treatment Length of experiment: 8 days Exponential growth phase pH: 7.93 ± 0.02 AT: 2255 ± 20 μmol L−1 Calculated pCO2: 719 ± 34 µatm DIC: 2136 ± 25 μmol L−1 Stationary growth phase pH: 7.93 ± 0.02 AT: 2279 ± 14 μmol L−1 Calculated pCO2: 732 ± 28 µatm DIC: 2175 ± 13 μmol L−1 | pH 8.0 vs. pH 7.9 Maximum carbon fixation rate increased * | [104] | ||
| pH 8.1 treatment Length of experiment: 7 days Exponential growth phase pH: 8.14 ± 0.01 AT: 2232 ± 9 μmol L−1 Calculated pCO2: 406 ± 8 µatm DIC: 2032 ± 11 μmol L−1 Stationary growth phase pH: 8.14 ± 0.01 AT: 2239 ± 25 μmol L−1 Calculated pCO2: 410 ± 2 µatm DIC: 2052 ± 21 μmol L−1 | pH 7.8 treatment Length of experiment: 9 days Exponential growth phase pH: 7.81 ± 0.03 AT: 2292 ± 3 μmol L−1 Calculated pCO2: 980 ± 64 µatm DIC: 2213 ± 11 μmol L−1 Stationary growth phase pH: 7.84 ± 0.01 AT: 2348 ± 17 μmol L−1 Calculated pCO2: 929 ± 12 µatm DIC: 2271 ± 16 μmol L−1 | pH 8.1 vs. pH 7.8 Growth rate decreased **** Maximum carbon fixation rate increased * | [104] | ||
| pH 8.0 treatment Length of experiment: 8 days Exponential growth phase pH: 8.03 ± 0.01 AT: 2248 ± 25 μmol L−1 Calculated pCO2: 550 ± 8 µatm DIC: 2093 ± 21 μmol L−1 Stationary growth phase pH: 8.04 ± 0.01 AT: 2254 ± 14 μmol L−1 Calculated pCO2: 537 ± 19 µatm DIC: 2107 ± 17 μmol L−1 | pH 7.8 treatment Length of experiment: 9 days Exponential growth phase pH: 7.81 ± 0.03 AT: 2292 ± 3 μmol L−1 Calculated pCO2: 980 ± 64 µatm DIC: 2213 ± 11 μmol L−1 Stationary growth phase pH: 7.84 ± 0.01 AT: 2348 ± 17 μmol L−1 Calculated pCO2: 929 ± 12 µatm DIC: 2271 ± 16 μmol L−1 | pH 8.0 vs. pH 7.8 Growth rate decreased **** Maximum carbon fixation rate increased * | [104] | ||
| pH 7.9 treatment Length of experiment: 8 days Exponential growth phase pH: 7.93 ± 0.02 AT: 2255 ± 20 μmol L−1 Calculated pCO2: 719 ± 34 µatm DIC: 2136 ± 25 μmol L−1 Stationary growth phase pH: 7.93 ± 0.02 AT: 2279 ± 14 μmol L−1 Calculated pCO2: 732 ± 28 µatm DIC: 2175 ± 13 μmol L−1 | pH 7.8 treatment Length of experiment: 9 days Exponential growth phase pH: 7.81 ± 0.03 AT: 2292 ± 3 μmol L−1 Calculated pCO2: 980 ± 64 µatm DIC: 2213 ± 11 μmol L−1 Stationary growth phase pH: 7.84 ± 0.01 AT: 2348 ± 17 μmol L−1 Calculated pCO2: 929 ± 12 µatm DIC: 2271 ± 16 μmol L−1 | pH 7.9 vs. pH 7.8 Growth rate Decreased **** | [104] | ||
| Pseudo-nitzschia spp. in Gullmar Fjord | Mesocosm, addition of CO2-saturated seawater | 380 µatm CO2 treatment Length of experiment: 111 days | 1000 µatm CO2 treatment Calculated pCO2: 760 ± 175 µatm Length of experiment: 111 days | 380 vs. 1000 µatm CO2 Cellular DA content increased * | [105] |
| PbTx-producing microalgae | |||||
| Karenia brevis (CCFWC-126) isolated from the Gulf of Mexico | Laboratory culture CO2 gas bubbling | 150 µatm CO2 treatment Before inoculation pH: 8.51 ± 0.04 AT: 2320 ± 3 μmol L−1 Calculated pCO2: 118.9 ± 0.1 µatm DIC: 1679 ± 2 μmol L−1 Mid exponential phase pH: 8.63 ± 0.05 AT: 2428 ± 64 μmol L−1 Calculated pCO2: 102.9 ± 16.8 µatm DIC: 1714 ± 4 μmol L−1 | 400 µatm CO2 treatment Before inoculation pH: 8.12 ± 0.02 AT: 2348 ± 18 μmol L−1 Calculated pCO2: 446.6 ± 31.0 µatm DIC: 2008 μmol L−1 Mid exponential phase pH: 8.22 ± 0.03 AT: 2376 ± 14 μmol L−1 Calculated pCO2: 355.6 ± 30.4 µatm DIC: 1982 ± 26 μmol L−1 | 150 vs. 400 µatm CO2 No significant response | [106] |
| 150 µatm CO2 treatment Before inoculation pH: 8.51 ± 0.04 AT: 2320 ± 3 μmol L−1 Calculated pCO2: 118.9 ± 0.1 µatm DIC: 1679 ± 2 μmol L−1 Mid exponential phase pH: 8.63 ± 0.05 AT: 2428 ± 64 μmol L−1 Calculated pCO2: 102.9 ± 16.8 µatm DIC: 1714 ± 4 μmol L−1 | 780 µatm CO2 treatment Before inoculation pH: 7.89 ± 0.01 AT: 2309 ± 1 μmol L−1 Calculated pCO2: 812.7 ± 0.5 µatm DIC: 2089 ± 1 μmol L−1 Mid exponential phase pH: 7.95 ± 0.04 AT: 2342 ± 19 μmol L−1 Calculated pCO2: 753.4 ± 78.8 µatm DIC: 2103 ± 4 μmol L−1 | 150 vs. 780 µatm CO2 No significant response | [106] | ||
| 400 µatm CO2 treatment Before inoculation pH: 8.12 ± 0.02 AT: 2348 ± 18 μmol L−1 Calculated pCO2: 446.6 ± 31.0 µatm DIC: 2008 μmol L−1 Mid exponential phase pH: 8.22 ± 0.03 AT: 2376 ± 14 μmol L−1 Calculated pCO2: 355.6 ± 30.4 µatm DIC: 1982 ± 26 μmol L−1 | 780 µatm CO2 treatment Before inoculation pH: 7.89 ± 0.01 AT: 2309 ± 1 μmol L−1 Calculated pCO2: 812.7 ± 0.5 µatm DIC: 2089 ± 1 μmol L−1 Mid exponential phase pH: 7.95 ± 0.04 AT: 2342 ± 19 μmol L−1 Calculated pCO2: 753.4 ± 78.8 µatm DIC: 2103 ± 4 μmol L−1 | 400 vs. 780 µatm CO2 No significant response | [106] | ||
| Karenia brevis (Wilson) isolated from John’s Pass | Laboratory culture CO2 gas bubbling prior to cell inoculation2 | 250 ppm CO2 treatment Length of experiment: 9 days Before inoculation Calculated pCO2: 241.2 µatm DIC: 1727.3 ± 33 μmol L−1 At the beginning of the experiment AT: 2082.8 ± 34.3 μmol L−1 Calculated pCO2: 134.2 µatm At the end of the experiment AT: 2198.6 ± 62.0 μmol L−1 Calculated pCO2: 80.5 µatm DIC: 1562.6 ± 14.5 μmol L−1 | 350 ppm CO2 treatment Length of experiment: 9 days Before inoculation Calculated pCO2: 318.4 µatm DIC: 1733.9 ± 163.7 μmol L−1 At the beginning of the experiment AT: 2021.3 ± 33.6 μmol L−1 Calculated pCO2: 115.6 µatm At the end of the experiment AT: 2250.5 ± 0.6 μmol L−1 Calculated pCO2: 55.1 µatm DIC: 1509.9 ± 259.7 μmol L−1 | No significant response | [107] |
| 250 ppm CO2 treatment Length of experiment: 9 days Before inoculation Calculated pCO2: 241.2 µatm DIC: 1727.3 ± 33 μmol L−1 At the beginning of the experiment AT: 2082.8 ± 34.3 μmol L−1 Calculated pCO2: 134.2 µatm At the end of the experiment AT: 2198.6 ± 62.0 μmol L−1 Calculated pCO2: 80.5 µatm DIC: 1562.6 ± 14.5 μmol L−1 | 1000 ppm CO2 treatment Length of experiment: 9 days Before inoculation Calculated pCO2: 1131.9 µatm DIC: 1986 ± 17.2 μmol L−1 At the beginning of the experiment AT: 2076.6 ± 6 μmol L−1 Calculated pCO2: 438.9 µatm At the end of the experiment AT: 2224.7 ± 10.7 μmol L−1 Calculated pCO2: 166.1 µatm DIC: 1750.8 ± 21.1 μmol L−1 | 250 vs. 1000 ppm CO2 Growth rate increased *** | [107] | ||
| 350 ppm CO2 treatment Length of experiment: 9 days Before inoculation Calculated pCO2: 318.4 µatm DIC: 1733.9 ± 163.7 μmol L−1 At the beginning of the experiment AT: 2021.3 ± 33.6 μmol L−1 Calculated pCO2: 115.6 µatm At the end of the experiment AT: 2250.5 ± 0.6 μmol L−1 Calculated pCO2: 55.1 µatm DIC: 1509.9 ± 259.7 μmol L−1 | 1000 ppm CO2 treatment Length of experiment: 9 days Before inoculation Calculated pCO2: 1131.9 µatm DIC: 1986 ± 17.2 μmol L−1 At the beginning of the experiment AT: 2076.6 ± 6 μmol L−1 Calculated pCO2: 438.9 µatm At the end of the experiment AT: 2224.7 ± 10.7 μmol L−1 Calculated pCO2: 166.1 µatm DIC: 1750.8 ± 21.1 μmol L−1 | 350 vs. 1000 ppm CO2 Growth rate increased *** | [107] | ||
| Karenia brevis (SP1) isolated from South Padre Island | 300 ppm CO2 treatment Length of experiment: 9 days Before inoculation Calculated pCO2: 299 ± 157.8 µatm At the beginning of the experiment Calculated pCO2: 211.8 ± 32 µatm At the end of the experiment Calculated pCO2: 99 ± 32.7 µatm | 1000 ppm CO2 treatment Length of experiment: 9 days Before inoculation Calculated pCO2: 1084 ± 14.1 µatm At the beginning of the experiment Calculated pCO2: 398.6 ± 3.4 µatm At the end of the experiment Calculated pCO2: 219 ± 11.1 µatm | 300 vs. 1000 ppm CO2 Growth rate increased | [107] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsui, T.-K.V.; Kong, H.-K. Metabolomics Approach to Reveal the Effects of Ocean Acidification on the Toxicity of Harmful Microalgae: A Review of the Literature. AppliedChem 2023, 3, 169-195. https://doi.org/10.3390/appliedchem3010012
Tsui T-KV, Kong H-K. Metabolomics Approach to Reveal the Effects of Ocean Acidification on the Toxicity of Harmful Microalgae: A Review of the Literature. AppliedChem. 2023; 3(1):169-195. https://doi.org/10.3390/appliedchem3010012
Chicago/Turabian StyleTsui, Tsz-Ki Victoria, and Hang-Kin Kong. 2023. "Metabolomics Approach to Reveal the Effects of Ocean Acidification on the Toxicity of Harmful Microalgae: A Review of the Literature" AppliedChem 3, no. 1: 169-195. https://doi.org/10.3390/appliedchem3010012
APA StyleTsui, T.-K. V., & Kong, H.-K. (2023). Metabolomics Approach to Reveal the Effects of Ocean Acidification on the Toxicity of Harmful Microalgae: A Review of the Literature. AppliedChem, 3(1), 169-195. https://doi.org/10.3390/appliedchem3010012