

Morphological Diversity of Different Male Morphotypes of Giant Freshwater Prawn Macrobrachium rosenbergii (De Man, 1879)
 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Specimens and Sampling
2.2. Identification of Morphotypes
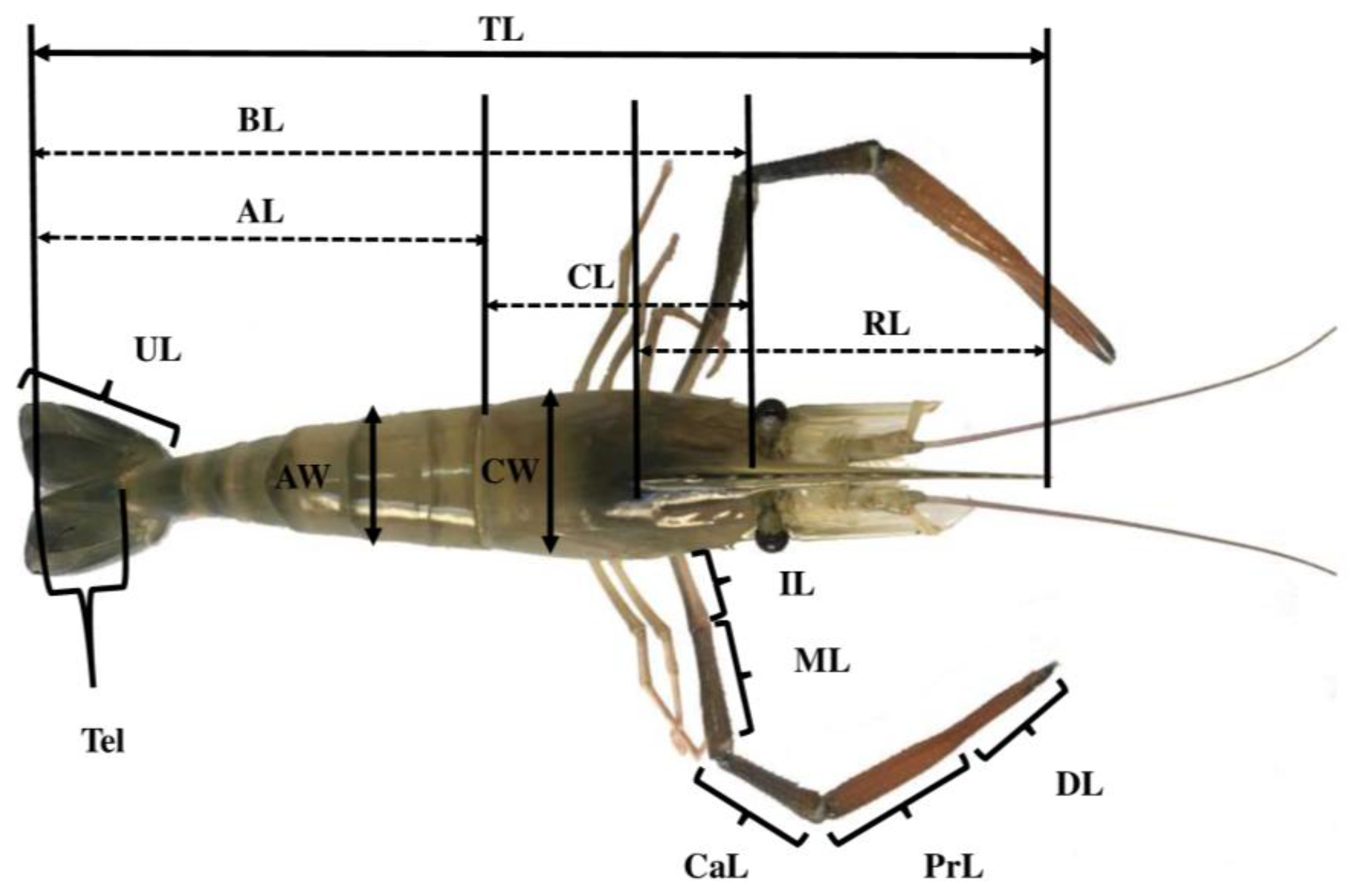
2.3. Morphometric Study
2.4. Statistical Analyses
3. Results
3.1. The Variance of Morphometric Traits
3.2. Relative Growth Analysis
3.3. Description of Major Cheliped in Different Male Morphotypes
4. Discussions
4.1. Morphotype Diversity
4.2. Relative Growth Patterns
4.3. Morphological Diversity of Chelipeds
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jasmine, S.; Molina, M.; Hossain, M.Y.; Jewel, M.A.S.; Ahamed, F.; Fulanda, B. Potential and economic viability of freshwater prawn Macrobrachium rosenbergii (de Man, 1879) polyculture with Indian major carps in Northwestern Bangladesh. Our Nat. 2011, 9, 61–72. [Google Scholar] [CrossRef]
- Thanh, N.M.; Nguyen, N.H.; Ponzoni, R.W.; Vu, N.T.; Barnes, A.C.; Mather, P.B. Estimates of strain additive and non-additive genetic effects for growth traits in a diallel cross of three strains of giant freshwater prawn (Macrobrachium rosenbergii) in Vietnam. Aquaculture 2010, 299, 30–36. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization of the United Nations). Yearbook of Fishery Statistics: Summary Tables; FAO: Rome, Italy, 2021; Available online: http://www.fao.org (accessed on 21 April 2023).
- Yu, L.; Zhu, X.; Liang, J.; Fan, J.; Chen, C. Analysis of genetic structure of wild and cultured Giant Freshwater Prawn (Macrobrachium rosenbergii) using newly developed microsatellite. Front. Mar. Sci. 2019, 6, 323. [Google Scholar] [CrossRef]
- Wortham, J.L.; Van Maurik, L.N. Morphology and morphotypes of the Hawaiian river shrimp, Macrobrachium grandimanus. J. Crustac. Biol. 2012, 32, 545–556. [Google Scholar] [CrossRef]
- Karplus, I.; Malecha, S.R.; Sagi, A. The biology and management of size variation. In Freshwater Prawn Culture, The Farming of Macrobrachium rosenbergii; New, M.B., Valenti, W.C., Eds.; Blackwell: Oxford, UK, 2000; pp. 259–289. [Google Scholar]
- Moraes-Riodades, P.M.; Valenti, W.C. Morphotypes in male Amazon River prawns, Macrobrachium amazonicum. Aquaculture 2004, 236, 297–307. [Google Scholar] [CrossRef]
- Akintola, S.L.; Kumolu-Johnson, C.A.; Whenu, O.O.; Anetekhai, M.A.; Ajibade, M.A. Three morphotypes of Macrobrachium macrobrachion (Decapoda, Palaemonidae): Preliminary results of analysis of morphological characters, with particular emphasis on the spines and rostrum. Zoology 2008, 6, 45–54. [Google Scholar] [CrossRef]
- Soundarapandian, P.; Dinakaran, G.K.; Varadharajan, D. Alternative mating strategies in male morphotypes of the prawn Macrobrachium idellaidella (Hilgendorf, 1898). J. Aquac. Res. Dev. 2013, 5, 1–10. [Google Scholar]
- Nogueira, C.S.; Pantaleão, J.A.F.; Almeida, A.C.; Costa, R.C. Male morphotypes of the freshwater prawn Macrobrachium brasiliense (Decapoda: Caridea: Palaemonidae). Invertebr. Biol. 2020, 139, e12279. [Google Scholar] [CrossRef]
- Rojas, R.; Morales, M.C.; Rivadeneira, M.M.; Thiel, M. Male morphotypes in the Andrean River shrimp Cryphiops caementarius (Decapoda: Caridea): Morphology, coloration and injuries. J. Zool. 2012, 288, 21–32. [Google Scholar] [CrossRef]
- Tang, Q.; Xia, Z.; Cai, M.; Du, H.; Yang, J.; Xu, Y.; Zhang, H.; Li, J.; Wu, Y.; Xie, J.; et al. Identification of differentially expressed genes and signalling pathways to elucidate molecular mechanisms underlying growth differences among the male morphotypes of Macrobrachium rosenbergii. Aquac. Res. 2020, 51, 5040–5049. [Google Scholar] [CrossRef]
- Karplus, I.; Barki, A. Male morphotypes and alternative mating tactics in freshwater prawns of the genus Macrobrachium: A review. Rev. Aquac. 2019, 11, 925–940. [Google Scholar] [CrossRef]
- Banu, M.R.; Christianus, A.; Ikhsan, N.F.M.; Rajaee, A.H. Effect of cold shock and hormone on growth and male morphotypes of freshwater prawn, Macrobrachium rosenbergii (de Man). Aquac. Res. 2016, 47, 3740–3746. [Google Scholar] [CrossRef]
- Ranjeet, K.; Kurup, B.M. Heterogeneous individual growth of Macrobrachium rosenbergii male morphotypes. Naga ICLARM Q. 2002, 25, 13–18. [Google Scholar]
- Dinh, H.; Nguyen, N.H. Genetic inheritance of female and male morphotypes in giant freshwater prawn Macrobrachium rosenbergii. PLoS ONE 2014, 9, 90–142. [Google Scholar] [CrossRef]
- Kuris, A.M.; Ra’anan, Z.; Sagi, A.; Cohen, D. Morphotypic differentiation of male Malaysian giant prawns, Macrobrachium rosenbergii. J. Crustac. Biol. 1987, 7, 219–237. [Google Scholar] [CrossRef]
- Pantaleão, J.A.F.; Hirose, G.L.; Costa, R.C. Occurrence of male morphotypes of Macrobrachium amazonicum (Caridea, Palaemonidae) in a population with an entirely freshwater life cycle. Braz. J. Biol. 2014, 74, S223–S232. [Google Scholar] [CrossRef]
- Knox, E.M.; Ng, R.T. Algorithms for mining distance based outliers in large datasets. In Proceedings of the International Conference on Very Large Data Bases, New York, NY, USA, 24–27 August 1998; Morgan Kaufmann Publishers Inc.: San Francisco, CA, USA, 1998; pp. 392–403. [Google Scholar]
- Sun, M.M.; Huang, J.H.; Jiang, S.G.; Yang, Q.B.; Zhou, F.L.; Zhu, C.Y.; Su, T.F. Morphometric analysis of four different populations of Penaeus monodon (Crustacea, Decapoda, Penaeidae). Aquac. Res. 2013, 45, 113–123. [Google Scholar] [CrossRef]
- Hawkins, S.J.; Hartnoll, R.G. The influence of barnacle cover on the numbers, growth and behaviour of Patella vulgata on a verticalpier. J. Mar. Biol. Assoc. UK 1982, 62, 855–867. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis; Prentice Hall: Upper Saddle River, NJ, USA, 2010. [Google Scholar]
- Ra’anan, Z.; Cohen, D. Ontogeny of social structure and population dynamics in the giant freshwater prawn, Macrobrachium rosenbergii (de Man) In Crustacean Issues. 2. Crustacean growth; Wenner, A., Schram, F.R., Eds.; A. A. Balkema Publishers: Amsterdam, The Netherlands, 1985. [Google Scholar]
- Ahammad, A.S.; Rahman, M.S.; Ahmed, M.B.U.; Rabbi, M.F.; Wahab, M.A. Morphogenetic characterization of morphotypes of the giant freshwater prawn, Macrobrachium rosenbergii. Bangladesh J. Fish. 2019, 31, 31–40. [Google Scholar]
- Flexa, C.E.; Silva, K.C.A.; Cintra, I.H.A. Morfometria do camarão-canela, Macrobrachium amazonicum (Heller, 1862), no município de Cametá-Pará. Bol. Técnico.-Científico. Do Cepnor. 2005, 5, 41–54. [Google Scholar] [CrossRef]
- Freire, J.L.; Bentes, B.; Fontes, V.B.; Silva EM, D. Morphometric discrimination among three stocks of Macrobrachium amazonicum in the Brazilian Amazon. Limnologica 2017, 64, 1–10. [Google Scholar] [CrossRef]
- Freire, J.L.; Marques, C.B.; Silva, B.B. Population structure and reproductive biology of Macrobrachium amazonicum (Heller, 1862) (Decapoda: Palaeomonidae) in an estuary in northeast Pará, Brazil. Braz. J. Aquat. Sci. Technol. 2012, 16, 65–76. (In Portuguese) [Google Scholar] [CrossRef]
- Santos, J.A.; Sampaio, C.M.; Soares Filho, A.A. Male population structure of the Amazon river prawn (Macrobrachium amazonicum) in a natural environment. Nauplius 2006, 14, 55–63. [Google Scholar]
- Silva-Oliveira, G.C.; Ready, J.S.; Iketani, G.; Bastos, S.; Gomes, G.; Sampaio, I.; Maciel, C. The invasive status of Macrobrachium rosenbergii (De Man, 1879) in Northern Brazil, with an estimation of areas at risk globally. Aquat. Invasions 2011, 6, 319–328. [Google Scholar] [CrossRef]
- Paschoal, L.R.P.; Zara, F.J. The androgenic gland in male morphotypes of the Amazon River prawn Macrobrachium amazonicum (Heller, 1862). Gen. Comp. Endocrinol. 2019, 275, 6–14. [Google Scholar] [CrossRef]
- Vergamini, F.G.; Pileggi, L.G.; Mantelatto, F.L. Genetic variability of the Amazon River prawn Macrobrachium amazonicum (Decapoda, Caridea, Palaemonidae). Contrib. Zool. 2011, 80, 67–83. [Google Scholar] [CrossRef]
- Pantaleão, J.A.F.; Hirose, G.L.; Costa, R.C. Relative growth, morphological sexual maturity, and size of Macrobrachium amazonicum (Heller 1862) (Crustacea, Decapoda, Palaemonidae) in a population with an entirely freshwater life cycle. Invertebr. Reprod. Dev. 2012, 56, 180–190. [Google Scholar] [CrossRef]
- Taddei, F.G.; Herrera, D.R.; Davanso, T.M.; Silva, T.E.D.; Costa, R.C.; Fransozo, A. Length/weight relationship and condition factor of Macrobrachium jelskii (Miers, 1877) and M. brasiliense (Heller, 1862) (Decapoda, Palaemonidae) in two locations in the state of São Paulo. Nauplius 2017, 25, e2017022. [Google Scholar] [CrossRef]
- Karplus, I.; Sagi, A. The biology and management of size variation. In Freshwater Prawns: Biology and Farming; New, M.B., Valenti, W.C., Tidwell, J.H., D’Abramo, L.R., Kutty, M.N., Eds.; Wiley-Blackwell: Chichester, UK, 2010; pp. 316–345. [Google Scholar]
- Rocha, S.S.D.; Barbosa, R.D.J. Population biology of Macrobrachium jelskii (Miers, 1877) (Decapoda, Palaemonidae) from an artificial pond in Bahia, Brazil. Nauplius 2017, 25, e2017023. [Google Scholar] [CrossRef]
- Banu, M.R.; Siraj, S.S.; Christianus, A.; Ikhsan, N.F.M.; Rajaee, A.H. Genetic variation among different morphotypes of the male freshwater prawn Macrobrachium rosenbergii (De Man). Aquac. Rep. 2015, 1, 15–19. [Google Scholar] [CrossRef]
- Maciel, C.R.; Valenti, W.C. Biology, fisheries, and aquaculture of the Amazon River prawn Macrobrachium amazonicum: A review. Nauplius 2009, 17, 61–79. [Google Scholar]
- Aziz, D.; Nguyen, V.T.; Rahi, M.L.; Hurwood, D.A.; Mather, P.B. Identification of genes that potentially affect social dominance hierarchy in adult male giant freshwater prawns (Macrobrachium rosenbergii). Aquaculture 2017, 476, 168–184. [Google Scholar] [CrossRef]
- Xue, H.B.; Wu, X.J.; Li, Z.H.; Liu, Y.; Yin, X.L.; Wang, W.N. Correlation and causation between the intestinal microbiome and male morphotypes in the giant freshwater prawn Macrobrachium rosenbergii. Aquaculture 2021, 531, 735–936. [Google Scholar] [CrossRef]
- Garcia-Perez, A.; Alston, D.E. Comparisons of male and female morphotypes distribution of freshwater prawn, Macrobrachium rosenbergii, in monoculture versus polyculture with Nile tilapia, Oreochromis niloticus. Caribb. J. Sci. 2000, 36, 340–342. [Google Scholar]
- Pillai, B.R.; Lalrinsanga, P.L.; Ponzoni, R.W.; Khaw, H.L.; Mahapatra, K.D.; Mohanty, S.; Patra, G.; Naik, N.; Pradhan, H.; Jayasankar, P. Phenotypic and genetic parameters for body traits in the giant freshwater prawn (Macrobrachium rosenbergii) in India. Aquac. Res. 2017, 48, 5741–5750. [Google Scholar] [CrossRef]
- Rios, P.; Pantaleão, A.F.; Hirose, L. Occurrence of male morphotypes in the freshwater prawn Macrobrachium acanthurus Wiegmann, 1836 (Decapoda, Palaemonidae). Invertebr. Reprod. Dev. 2021, 65, 268–278. [Google Scholar] [CrossRef]
- Karplus, I. Social control of growth in Macrobrachium rosenbergii (De Man): A review and prospects for future research. Aquac. Res. 2005, 36, 238–254. [Google Scholar] [CrossRef]
- Ranjeet, K.; Kurup, B.M. Primary intrinsic factors governing the heterogenous individual growth in male morphotypes of Macrobrachium rosenbergii. In Proceedings of the International Conference on Fisheries, Aquaculture and Environment in the NW Indian Ocean, Muscat, Oman, 8–10 January 2001; Sultan Qaboos University, Sultanate of Oman: Sultan Qaboos, Oman, 2001. [Google Scholar]
- Correa, C.; Baeza, J.A.; Dupré, E.; Hinojosa, I.A.; Thiel, M. Mating behavior and fertilization success of three ontogenetic stages of male rock shrimp Rhynchocinetes typus (Decapoda: Caridea). J. Crustac. Biol. 2000, 20, 628–640. [Google Scholar] [CrossRef][Green Version]
- Thiel, M.; Chak, S.T.C.; Dumont, C.P. Male morphotypes and mating behavior of the dancing shrimp Rhynchocinetes brucei (Decapoda: Caridea). J. Crustac. Biol. 2010, 30, 580–588. [Google Scholar] [CrossRef]
- Mariappan, P.; Balasundaram, C. Studies on the morphometry of Macrobrachium nobilii (Decapoda, Palaemonidae). Braz. Arch. Biol. Technol. 2004, 47, 441–449. [Google Scholar] [CrossRef]
- Silva, G.M.F.; Ferreira, M.A.P.; Ledebur, E.I.C.F.V.; Rocha RM, D. Gonadal structure analysis of Macrobrachium amazonicum (Heller, 1862) from a wild population: A new insight into the morphotype characterization. Aquac. Res. 2009, 40, 798–803. [Google Scholar] [CrossRef]
- Hoshan, I.; Yesmin, A.; Ray, M.; Ahmed, F.F.; Mahfuj, S. Multivariate morphometric differentiation of Macrobrachium species (Crustacea: Palaemonidae) along the northern rivers of Bangladesh. Bangladesh J. Fish. 2022, 34, 27–39. [Google Scholar] [CrossRef]
- Ismael, D.; New, M.B. Biology. In Freshwater Prawn Culture: The Farming of Macrobrachium rosenbergii; New, M.B., Valenti, W.C., Eds.; Blackwell: Oxford, UK, 2000; pp. 18–40. [Google Scholar]
- Rossi, N.; Pantaleão, J.A.F.; Mantelatto, F.L.M. Integrated morphometric and molecular analyses indicate three male morphotypes in the freshwater prawn Macrobrachium olfersii (Decapoda, Palaemonidae) along the Brazilian neotropical region. Acta Zool. 2022, 1–14. [Google Scholar] [CrossRef]
- Short, J.W. A revision of Australian river prawns, Macrobrachium (Crustacea: Decapoda: Palaemonidae). Hydrobiologia 2004, 525, 1–100. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morphometric Variables | Morphotypes | |||||||
|---|---|---|---|---|---|---|---|---|
| SM (n = 62) | BC (n = 40) | OC (n = 62) | OBC (n = 51) | |||||
| Mean ± SD | Range | Mean ± SD | Range | Mean ± SD | Range | Mean ± SD | Range | |
| Total length | 89.87 ± 10.62 d | 72.68–116.68 | 161.36 ± 11.47 b | 140.00–186.00 | 170.16 ± 10.78 a | 131.00–183.00 | 143.07 ± 16.47 c | 117.45–225.95 |
| Body length | 67.32 ± 9.42 d | 52.64–108.51 | 123.74 ±9.16 b | 107.02–143.91 | 129.48 ± 8.28 a | 96.71–139.30 | 110.15 ± 11.55 c | 90.16–157.65 |
| Rostrum length | 33.15 ± 4.74 d | 23.17–44.25 | 59.42 ± 5.19 b | 43.56–70.23 | 61.66 ± 4.69 a | 43.48–69.21 | 52.96 ± 8.14 c | 26.78–95.30 |
| Abdominal length | 45.03 ± 5.04 c | 36.45–58.61 | 79.73 ± 5.92 a | 65.65- 96.42 | 81.02 ± 5.26 a | 63.47- 90.10 | 69.91 ± 8.29 b | 58.22- 110.24 |
| Abdominal width | 9.80 ± 1.36 c | 7.93–13.74 | 20.08 ± 2.30 ab | 7.98–54.03 | 21.23 ± 4.84 a | 15.18–50.61 | 18.33 ± 5.24 b | 14.14–52.9 |
| Abdominal depth | 12.41 ± 1.28 c | 9.98–15.17 | 23.61 ± 2.78 ab | 12.56–60.88 | 24.87 ± 5.04 a | 18.58–57.16 | 21.54 ± 5.14 b | 17.74–55.31 |
| Carapace length | 20.71 ± 2.8 c | 15.61–27.68 | 43.50 ± 4.35 a | 29.59–55.3 | 45.82 ± 3.65 a | 33.22–52.9 | 40.17 ± 6.40 b | 32.65–78.02 |
| Carapace width | 12.18 ± 1.9 c | 9.21–18.58 | 26.73 ± 3.02 a | 14.63–33.56 | 28.33 ± 2.53 a | 19.26–30.99 | 24.77 ± 5.54 b | 19.57–59.7 |
| Carapace depth | 13.95 ± 1.99 c | 10.37–18.5 | 32.29 ± 3.02 a | 21.25–41.23 | 32.62 ± 2.90 ab | 23.73–39.85 | 30.68 ± 7.47 b | 24.49–67.78 |
| Major cheliped length | 50.79 ± 8.94 d | 36.40–76.89 | 140.46 ± 21.86 c | 105.37–275.20 | 159.58 ± 28.64 b | 85.00–196.00 | 193.32 ± 30.09 a | 151.49–260.00 |
| Propodus length | 10.23 ± 1.96 d | 6.71–15.46 | 32.07 ± 7.05 c | 16.23–68.20 | 36.61 ± 7.92 b | 17.62–50.48 | 48.13 ± 7.73 a | 33.20–67.62 |
| Propodus width | 1.90 ± 0.47 c | 1.15–3.01 | 6.41 ± 1.18 b | 3.67–10.57 | 7.39 ± 1.34 a | 3.49–8.37 | 7.05 ± 0.97 a | 5.18–9.64 |
| Carpus length | 11.81 ± 2.25 c | 8.21–17.38 | 29.28 ± 6.76 b | 20.81–66.92 | 34.30 ± 6.77 b | 19.10–45.72 | 46.49 ± 10.70 a | 32.35–86.77 |
| Carpus width | 1.69 ± 0.45 c | 0.97–2.67 | 6.14 ± 1.06 b | 4.31–9.72 | 6.65 ± 1.19 ab | 3.24–8.41 | 7.06 ± 1.08 a | 5.11–9.57 |
| Telson length | 11.18 ± 1.35 d | 8.64–14.49 | 20.05 ± 1.45 b | 17.28–56.04 | 21.80 ± 4.85 a | 16.52–56.54 | 18.33 ± 1.77 c | 14.96–23.78 |
| Uropod length | 15.26 ± 1.85 d | 11.24–19.60 | 26.48 ± 3.14 b | 16.96–65.94 | 28.39 ± 5.37 a | 20.88–31.86 | 23.77 ± 2.45 c | 18.27–29.53 |
| Major cheliped weight | 0.14 ± 0.17 d | 0.01–0.70 | 5.14 ± 2.40 c | 2.02–30.71 | 8.98 ± 4.53 b | 1.40–11.25 | 11.22 ± 5.49 a | 3.18–27.37 |
| Carapace weight | 2.91 ± 1.24 c | 1.21–6.67 | 28.29 ± 5.71 a | 19.86–41.12 | 29.55 ± 6.03 a | 11.12–38.13 | 20.51 ± 6.02 b | 11.57–35.67 |
| Abdominal weight | 3.74 ± 1.36 d | 1.73–7.88 | 22.47 ± 4.24 b | 13.40–35.92 | 25.25 ± 4.03 a | 11.42–33.97 | 14.66 ± 4.35 c | 8.31–24.83 |
| Wet weight | 7.25 ± 2.86 d | 3.20–16.20 | 58.83 ± 10.85 b | 40.59–99.60 | 68.52 ± 14.19 a | 25.00–79.35 | 48.34 ± 15.14 c | 25.70–88.36 |
| Variables | Components | |
|---|---|---|
| PC1 | PC2 | |
| Body weight (Bw) | 0.141 | 0.398 |
| Major cheliped weight (MCw) | 0.778 | 0.503 |
| Major cheliped length (MCL) | 0.883 | 0.429 |
| Propodus length (PrL) | 0.844 | 0.491 |
| Propodus width (PrW) | 0.834 | 0.22 |
| Carpus length (CaL) | 0.782 | 0.544 |
| Carpus width (CaW) | 0.936 | 0.256 |
| Total length (TL) | 0.965 | −0.174 |
| Rostrum length (RL) | 0.947 | −0.142 |
| Body length (BL) | 0.967 | −0.149 |
| Carapace length (CL) | 0.978 | −0.098 |
| Carapace width (CW) | 0.969 | −0.109 |
| Carapace depth (CD) | 0.943 | −0.041 |
| Abdominal length (AL) | 0.964 | −0.199 |
| Abdominal width (AW) | 0.892 | −0.227 |
| Abdominal depth (AD) | 0.909 | −0.225 |
| Telson length (TeL) | 0.871 | −0.236 |
| Uropod length (UL) | 0.893 | −0.253 |
| Carapace weight (Cw) | 0.941 | −0.149 |
| Abdominal weight (Aw) | 0.908 | −0.256 |
| Cumulative variance explained | 78.302 | 8.46 |
| Eigenvalues | 15.66 | 1.69 |
| Relationship | Morphotypes | n | a | b | r2 | p-Value | Allometry |
|---|---|---|---|---|---|---|---|
| TL vs. CL | SM | 62 | 2.01 | 0.82 | 0.88 | 0.000 | − |
| BC | 40 | 3.20 | 0.51 | 0.58 | 0.001 | − | |
| OC | 62 | 2.50 | 0.68 | 0.72 | 0.000 | − | |
| OBC | 51 | 1.78 | 0.86 | 0.87 | 0.0047 | − | |
| AL vs. CL | SM | 62 | 1.53 | 0.75 | 0.84 | 0.000 | − |
| BC | 40 | 2.44 | 0.51 | 0.52 | 0.000 | − | |
| OC | 62 | 1.95 | 0.64 | 0.63 | 0.000 | − | |
| OBC | 51 | 1.38 | 0.77 | 0.88 | 0.000 | − | |
| MCL vs. CL | SM | 62 | 3.28 | 0.45 | 0.56 | 0.000 | − |
| BC | 40 | 1.08 | 0.93 | 0.56 | 0.000 | = | |
| OC | 62 | 1.46 | 1.03 | 0.36 | 0.000 | = | |
| OBC | 51 | −0.89 | 1.55 | 0.52 | 0.075 | + | |
| CaL vs. CL | SM | 62 | −0.23 | 0.88 | 0.53 | 0.000 | − |
| BC | 40 | 1.17 | 0.97 | 0.61 | 0.193 | = | |
| OC | 62 | 0.03 | 1.02 | 0.41 | 0.000 | = | |
| OBC | 51 | −2.12 | 1.47 | 0.55 | 0.007 | + | |
| PrL vs. CL | SM | 62 | 0.10 | 0.33 | 0.99 | 0.000 | − |
| BC | 40 | 0.31 | 0.97 | 0.99 | 0.000 | = | |
| OC | 62 | 0.32 | 0.97 | 0.46 | 0.000 | = | |
| OBC | 51 | 0.48 | 1.42 | 0.35 | 0.000 | + | |
| RL vs. CL | SM | 62 | 1.70 | 0.82 | 0.91 | 0.000 | − |
| BC | 40 | 2.9 | 0.50 | 0.53 | 0.000 | − | |
| OC | 62 | 1.23 | 0.76 | 0.59 | 0.000 | − | |
| OBC | 51 | 1.42 | 0.69 | 0.60 | 0.000 | − |
| Morphotype | Segment | Spine Height | Spine Angle | Spination | Color | ||||
|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Range | Mean | SD | Range | ||||
| dactylus | 0.52 | 0.09 | 0.36–0.65 | 60.25 | 6.05 | 51.52–73.8 | + | orange | |
| ischium | 0.43 | 0.05 | 0.36–0.50 | 64.31 | 3.67 | 52–67.4 | ++ | beige blue | |
| OC | merus | 0.74 | 0.18 | 0.5–1.04 | 61.59 | 3.92 | 53.13–68.4 | + + | pale blue |
| carpus | 0.81 | 0.07 | 0.68–0.94 | 59.37 | 5.42 | 42.3–67 | + + | blue | |
| propodus | 0.61 | 0.09 | 0.51–0.90 | 61.75 | 6.05 | 53–75.3 | + + | orange | |
| dactylus | 0.62 | 0.11 | 0.30–0.72 | 67.48 | 4.56 | 38.78–78.91 | + | deep blue | |
| ischium | 0.51 | 0.09 | 0.28–0.55 | 63.12 | 9.16 | 45–73.6 | + + | pale | |
| BC | merus | 0.86 | 0.22 | 0.57–1.22 | 76.38 | 5.68 | 62.5–83.4 | + + | beige blue |
| carpus | 1.07 | 0.2 | 0.73–1.41 | 83.00 | 7.64 | 64.8–92.5 | + + + | blue | |
| propodus | 0.67 | 0.11 | 0.45–0.97 | 68.99 | 4.56 | 58.7–77.5 | + + + | blue | |
| dactylus | 0.71 | 0.11 | 0.56–1.00 | 60.70 | 10.05 | 37.2–77.41 | + | deep blue | |
| ischium | 0.48 | 0.08 | 0.3–0.61 | 54.66 | 7.99 | 40.6–70.1 | + + | light blue | |
| OBC | merus | 0.86 | 0.17 | 0.55–1.13 | 57.37 | 5.25 | 50–68.03 | + + + | deep blue |
| carpus | 1.16 | 0.19 | 0.87–1.44 | 76.00 | 7.05 | 66.13–96.1 | + + + | deep blue | |
| propodus | 0.86 | 0.11 | 0.72–1.15 | 62.20 | 10.06 | 38.8–79 | + + + | deep blue | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, S.; Zhong, Z.; Lan, X.; Luo, J.; Tang, Q.; Xia, Z.; Yi, S.; Yang, G. Morphological Diversity of Different Male Morphotypes of Giant Freshwater Prawn Macrobrachium rosenbergii (De Man, 1879). Aquac. J. 2023, 3, 133-148. https://doi.org/10.3390/aquacj3020012
Ibrahim S, Zhong Z, Lan X, Luo J, Tang Q, Xia Z, Yi S, Yang G. Morphological Diversity of Different Male Morphotypes of Giant Freshwater Prawn Macrobrachium rosenbergii (De Man, 1879). Aquaculture Journal. 2023; 3(2):133-148. https://doi.org/10.3390/aquacj3020012
Chicago/Turabian StyleIbrahim, Salifu, Zhenxiao Zhong, Xuan Lan, Jinping Luo, Qiongying Tang, Zhenglong Xia, Shaokui Yi, and Guoliang Yang. 2023. "Morphological Diversity of Different Male Morphotypes of Giant Freshwater Prawn Macrobrachium rosenbergii (De Man, 1879)" Aquaculture Journal 3, no. 2: 133-148. https://doi.org/10.3390/aquacj3020012
APA StyleIbrahim, S., Zhong, Z., Lan, X., Luo, J., Tang, Q., Xia, Z., Yi, S., & Yang, G. (2023). Morphological Diversity of Different Male Morphotypes of Giant Freshwater Prawn Macrobrachium rosenbergii (De Man, 1879). Aquaculture Journal, 3(2), 133-148. https://doi.org/10.3390/aquacj3020012