
Fatty Acid Profiles of Selected Microalgae Used as Live Feeds for Shrimp Postlarvae in Vietnam

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lipid Class Composition
2.2. Methylation and Analysis of Fatty Acid Methyl Esters (FAME)
3. Results
3.1. The Lipid and Fatty Acid Profiles of 10 Microalgal Species
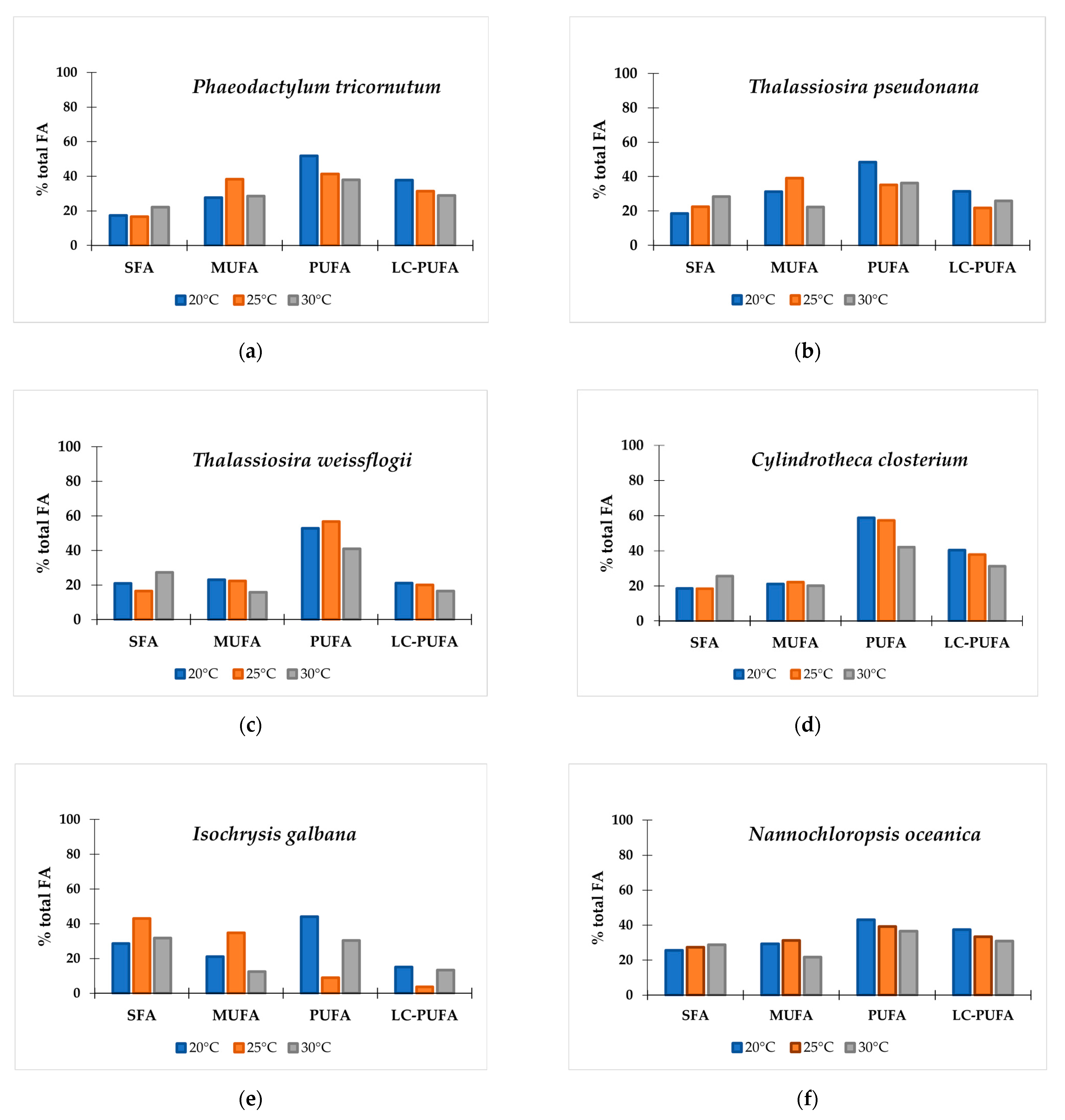
3.2. The Effect of Temperature on Fatty Acid Profiles in 10 Microalgal Species
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Lembi, C.A.; Waaland, J.R. Algae and Human Affairs; Cambridge University Press: Cambridge, UK, 1988; pp. 5–22. [Google Scholar]
- Metting, F. Biodiversity and application of microalgae. J. Ind. Microbiol. 1996, 17, 477–489. [Google Scholar] [CrossRef]
- Apt, K.E.; Behrens, P.W. Commercial developments in microalgal biotechnology. J. Phycol. 1999, 35, 215–226. [Google Scholar] [CrossRef]
- Ariede, M.B.; Candido, T.M.; Jacome, A.L.M.; Velasco, M.V.R.; de Carvalho, J.C.M.; Baby, A.R. Cosmetic attributes of algae-A review. Algal Res. 2017, 25, 483–487. [Google Scholar] [CrossRef]
- Priyadarshani, I.; Rath, B. Commercial and industrial applications of micro algae–A review. J. Algal Biomass Util. 2012, 3, 89–100. [Google Scholar]
- Mehariya, S.; Goswami, R.K.; Verma, P.; Lavecchia, R.; Zuorro, A. Integrated approach for wastewater treatment and biofuel production in microalgae biorefineries. Energies 2021, 14, 2282. [Google Scholar] [CrossRef]
- Zullaikah, S.; Utomo, A.T.; Yasmin, M.; Ong, L.K.; Ju, Y.H. Ecofuel conversion technology of inedible lipid feedstocks to renewable fuel. In Advances in Eco-Fuels for a Sustainable Environment; Elsevier: Amsterdam, The Netherlands, 2019; pp. 237–276. [Google Scholar]
- Salama, E.-S.; Kurade, M.B.; Abou-Shanab, R.A.; El-Dalatony, M.M.; Yang, I.-S.; Min, B.; Jeon, B.-H. Recent progress in microalgal biomass production coupled with wastewater treatment for biofuel generation. Renew. Sustain. Energy Rev. 2017, 79, 1189–1211. [Google Scholar] [CrossRef]
- D’Alessandro, E.B.; Antoniosi Filho, N.R. Concepts and studies on lipid and pigments of microalgae: A review. Renew. Sustain. Energy Rev. 2016, 58, 832–841. [Google Scholar] [CrossRef]
- Demirbas, A. Use of algae as biofuel sources. Energy Convers. Manag. 2010, 51, 2738–2749. [Google Scholar] [CrossRef]
- Tam, L.T.; Van Cong, N.; Ha, N.C.; Hang, N.T.M.; Van Minh, C.; Hong, D.D. Cultivation and biomass production of the diatom Thalassiosira weissflogii as a live feed for white-leg shrimp in hatcheries and commercial farms in Vietnam. J. Appl. Phycol. 2021, 33, 1559–1577. [Google Scholar] [CrossRef]
- Deshmukh, S.; Kumar, R.; Bala, K. Microalgae biodiesel: A review on oil extraction, fatty acid composition, properties and effect on engine performance and emissions. Fuel Process. Technol. 2019, 191, 232–247. [Google Scholar] [CrossRef]
- Kandathil Radhakrishnan, D.; AkbarAli, I.; Schmidt, B.V.; John, E.M.; Sivanpillai, S.; Thazhakot Vasunambesan, S. Improvement of nutritional quality of live feed for aquaculture: An overview. Aquac. Res. 2020, 51, 1–17. [Google Scholar] [CrossRef]
- Glencross, B.D. Exploring the nutritional demand for essential fatty acids by aquaculture species. Rev. Aquac. 2009, 1, 71–124. [Google Scholar] [CrossRef]
- Figueiredo, J.; Lin, J.; Anto, J.; Narciso, L. The consumption of DHA during embryogenesis as an indicative of the need to supply DHA during early larval development: A review. J. Aquac. Res. Dev. 2012, 3, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Anger, K. Patterns of growth and chemical composition in decapod crustacean larvae. Invertebr. Reprod. Dev. 1998, 33, 159–176. [Google Scholar] [CrossRef]
- Harrison, K.E. The role of nutrition in maturation, reproduction and embryonic development of decapod crustacean: A review. J. Shellfish Res. 1990, 9, 1–28. [Google Scholar]
- Hubbs, C.; Blaxter, J. Ninth larval fish conference: Development of sense organs and behaviour of Teleost larvae with special reference to feeding and predator avoidance. Trans. Am. Fish. Soc. 1986, 115, 98–114. [Google Scholar] [CrossRef]
- Griffiths, M.J.; van Hille, R.P.; Harrison, S.T. Lipid productivity, settling potential and fatty acid profile of 11 microalgal species grown under nitrogen replete and limited conditions. J. Appl. Phycol. 2012, 24, 989–1001. [Google Scholar] [CrossRef]
- Volkman, J. Sterols in microorganisms. Appl. Microbiol. Biotechnol. 2003, 60, 495–506. [Google Scholar] [CrossRef]
- Huu, N. Personal Communication; Directorate of Fisheries: Hanoi, Vietnam, 2019. [Google Scholar]
- Ngoc, Q.T.K.; Xuan, B.B.; Sandorf, E.D.; Phong, T.N.; Trung, L.C.; Hien, T.T. Willingness to adopt improved shrimp aquaculture practices in Vietnam. Aquac. Econ. Manag. 2021, 1–24. [Google Scholar] [CrossRef]
- Hai, T.N.; Duc, P.M.; Son, V.N.; Minh, T.H.; Phuong, N.T. Innovation in seed production and farming of marine shrimp in Vietnam. World Aquac. 2015, 46, 32–37. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Bakes, M.J.; Nichols, P.D. Lipid, fatty acid and squalene composition of liver oil from six species of deep-sea sharks collected in southern Australian waters. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1995, 110, 267–275. [Google Scholar] [CrossRef]
- Chang, K.J.L.; Mansour, M.P.; Dunstan, G.A.; Blackburn, S.I.; Koutoulis, A.; Nichols, P.D. Odd-chain polyunsaturated fatty acids in thraustochytrids. Phytochemistry 2011, 72, 1460–1465. [Google Scholar] [CrossRef]
- Huerlimann, R.; De Nys, R.; Heimann, K. Growth, lipid content, productivity, and fatty acid composition of tropical microalgae for scale-up production. Biotechnol. Bioeng. 2010, 107, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Fernández, E.; Acosta-Salmón, H.; Southgate, P.C. The nutritional value of seven species of tropical microalgae for black-lip pearl oyster (Pinctada margaritifera, L.) larvae. Aquaculture 2006, 257, 491–503. [Google Scholar] [CrossRef]
- Sargent, J.; McEvoy, L.; Bell, J. Requirements, presentation and sources of polyunsaturated fatty acids in marine fish larval feeds. Aquaculture 1997, 155, 117–127. [Google Scholar] [CrossRef]
- Patil, V.; Källqvist, T.; Olsen, E.; Vogt, G.; Gislerød, H.R. Fatty acid composition of 12 microalgae for possible use in aquaculture feed. Aquac. Int. 2007, 15, 1–9. [Google Scholar] [CrossRef]
- Lin, Y.-H.; Chang, F.-L.; Tsao, C.-Y.; Leu, J.-Y. Influence of growth phase and nutrient source on fatty acid composition of Isochrysis galbana CCMP 1324 in a batch photoreactor. Biochem. Eng. J. 2007, 37, 166–176. [Google Scholar] [CrossRef]
- Fidalgo, J.; Cid, A.; Torres, E.; Sukenik, A.; Herrero, C. Effects of nitrogen source and growth phase on proximate biochemical composition, lipid classes and fatty acid profile of the marine microalga Isochrysis galbana. Aquaculture 1998, 166, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Savvidou, M.G.; Boli, E.; Logothetis, D.; Lymperopoulou, T.; Ferraro, A.; Louli, V.; Mamma, D.; Kekos, D.; Magoulas, K.; Kolisis, F.N. A study on the effect of macro-and micro-nutrients on Nannochloropsis oceanica growth, fatty acid composition and magnetic harvesting efficiency. Plants 2020, 9, 660. [Google Scholar] [CrossRef]
- Sirisuk, P.; Sunwoo, I.; Kim, S.H.; Awah, C.C.; Ra, C.H.; Kim, J.-M.; Jeong, G.-T.; Kim, S.-K. Enhancement of biomass, lipids, and polyunsaturated fatty acid (PUFA) production in Nannochloropsis oceanica with a combination of single wavelength light emitting diodes (LEDs) and low temperature in a three-phase culture system. Bioresour. Technol. 2018, 270, 504–511. [Google Scholar] [CrossRef]
- Mitani, E.; Nakayama, F.; Matsuwaki, I.; Ichi, I.; Kawabata, A.; Kawachi, M.; Kato, M. Fatty acid composition profiles of 235 strains of three microalgal divisions within the NIES Microbial Culture Collection. Microb. Resour. Syst 2017, 33, 1929. [Google Scholar]
- Volkman, J.K.; Brown, M.R.; Dunstan, G.A.; Jeffrey, S. The biochemical composition of marine microalgae from the class Eustigmatophyceae 1. J. Phycol. 1993, 29, 69–78. [Google Scholar] [CrossRef]
- Marella, T.K.; Tiwari, A. Marine diatom Thalassiosira weissflogii based biorefinery for co-production of eicosapentaenoic acid and fucoxanthin. Bioresour. Technol. 2020, 307, 123245. [Google Scholar] [CrossRef]
- Vella, F.M.; Sardo, A.; Gallo, C.; Landi, S.; Fontana, A.; d’Ippolito, G. Annual outdoor cultivation of the diatom Thalassiosira weissflogii: Productivity, limits and perspectives. Algal Res. 2019, 42, 101553. [Google Scholar] [CrossRef]
- Sahin, M.S.; Khazi, M.I.; Demirel, Z.; Dalay, M.C. Variation in growth, fucoxanthin, fatty acids profile and lipid content of marine diatoms Nitzschia sp. and Nanofrustulum shiloi in response to nitrogen and iron. Biocatal. Agric. Biotechnol. 2019, 17, 390–398. [Google Scholar] [CrossRef]
- Lou, Y.; Liu, Y.; Wang, H.; Li, N.; Liu, Q.; Liu, Y.; Zhao, X. Effects of CO2 restriction on growth of Nitzschia closterium: Evidence from stable isotopes and fatty acids. Ecotoxicol. Environ. Saf. 2019, 177, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Zhuo, W.-H.; Wang, X.-W.; Chen, C.-P.; Gao, Y.-H.; Liang, J.-R. Effects of fundamental nutrient stresses on the lipid accumulation profiles in two diatom species Thalassiosira weissflogii and Chaetoceros muelleri. Bioprocess Biosyst. Eng. 2018, 41, 1213–1224. [Google Scholar] [CrossRef] [PubMed]
- Renaud, S.M.; Thinh, L.-V.; Parry, D.L. The gross chemical composition and fatty acid composition of 18 species of tropical Australian microalgae for possible use in mariculture. Aquaculture 1999, 170, 147–159. [Google Scholar] [CrossRef]
- Zhukova, N.V.; Aizdaicher, N.A. Fatty acid composition of 15 species of marine microalgae. Phytochemistry 1995, 39, 351–356. [Google Scholar] [CrossRef]
- Pratoomyot, J.; Srivilas, P.; Noiraksar, T. Fatty acids composition of 10 microalgal species. Songklanakarin J. Sci. Technol 2005, 27, 1179–1187. [Google Scholar]
- Brown, M.R.; Dunstan, G.A.; Norwood, S.J.; Miller, K.A. Effects of harvest stage and light on the biochemical composition of the diatom Thalassiosira pseudonana 1. J. Phycol. 1996, 32, 64–73. [Google Scholar] [CrossRef]
- Qiao, H.; Cong, C.; Sun, C.; Li, B.; Wang, J.; Zhang, L. Effect of culture conditions on growth, fatty acid composition and DHA/EPA ratio of Phaeodactylum tricornutum. Aquaculture 2016, 452, 311–317. [Google Scholar] [CrossRef]
- Chang, K.S.; Kim, J.; Park, H.; Hong, S.-J.; Lee, C.-G.; Jin, E. Enhanced lipid productivity in AGP knockout marine microalga Tetraselmis sp. using a DNA-free CRISPR-Cas9 RNP method. Bioresour. Technol. 2020, 303, 122932. [Google Scholar] [CrossRef] [PubMed]
- Suresh, K.S.; Suresh, P.; Kudre, T.G. Prospective ecofuel feedstocks for sustainable production. In Advances in Eco-Fuels for a Sustainable Environment; Elsevier: Amsterdam, The Netherlands, 2019; pp. 89–117. [Google Scholar]
- Gatamaneni, B.L.; Orsat, V.; Lefsrud, M. Factors affecting growth of various microalgal species. Environ. Eng. Sci. 2018, 35, 1037–1048. [Google Scholar] [CrossRef]
- Indrayani, I.; Moheimani, N.R.; de Boer, K.; Bahri, P.A.; Borowitzka, M.A. Temperature and salinity effects on growth and fatty acid composition of a halophilic diatom, Amphora sp. MUR258 (Bacillariophyceae). J. Appl. Phycol. 2020, 32, 977–987. [Google Scholar] [CrossRef]
- Chaisutyakorn, P.; Praiboon, J.; Kaewsuralikhit, C. The effect of temperature on growth and lipid and fatty acid composition on marine microalgae used for biodiesel production. J. Appl. Phycol. 2018, 30, 37–45. [Google Scholar]
- Aussant, J.; Guihéneuf, F.; Stengel, D.B. Impact of temperature on fatty acid composition and nutritional value in eight species of microalgae. Appl. Microbiol. Biotechnol. 2018, 102, 5279–5297. [Google Scholar] [CrossRef] [PubMed]
- Renaud, S.M.; Thinh, L.-V.; Lambrinidis, G.; Parry, D.L. Effect of temperature on growth, chemical composition and fatty acid composition of tropical Australian microalgae grown in batch cultures. Aquaculture 2002, 211, 195–214. [Google Scholar] [CrossRef]
- Renaud, S.M.; Zhou, H.; Parry, D.L.; Thinh, L.-V.; Woo, K. Effect of temperature on the growth, total lipid content and fatty acid composition of recently isolated tropical microalgae Isochrysis sp., Nitzschia closterium, Nitzschia paleacea, and commercial species Isochrysis sp.(clone T. ISO). J. Appl. Phycol. 1995, 7, 595–602. [Google Scholar] [CrossRef]
- Thompson, P.A.; Guo, M.x.; Harrison, P.J.; Whyte, J.N. Effects of variation in temperature. II. On the fatty acid composition of eight species of marine phytoplankton 1. J. Phycol. 1992, 28, 488–497. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus Plus Species | Class | Class Vernacular | Source Country | |
|---|---|---|---|---|
| CS-5 | Cylindrotheca closterium | Bacillariophyceae | Diatom | Australia |
| CS-29 | Phaeodactylum tricornutum | Bacillariophyceae | Diatom | United Kingdom |
| CS-176 | Chaetoceros muelleri | Coscinodiscophyceae | Diatom | USA |
| CS-178 | Chaetoceros calcitrans | Coscinodiscophyceae | Diatom | Japan |
| CS-252 | Skeletonema pseudocostatum | Coscinodiscophyceae | Diatom | Australia |
| CS-173 | Thalassiosira pseudonana | Coscinodiscophyceae | Diatom | USA |
| CS-871 | Thalassiosira weissflogii | Coscinodiscophyceae | Diatom | USA |
| CS-186 | Isochrysis galbana | Coccolothophyceae | Prymnesiophyte | USA |
| CS-177 | Tisochrysis lutea | Coccolothophyceae | Prymnesiophyte | Tahiti |
| CS-179 | Nannochloropsis oceanica | Eustigmaophyceae | Eustigmatophycea | Japan |
| Species | Tisochrysis lutea | Isochrysis galbana | Cylindrotheca closterium | Phaeodactylum tricornutum | Skeletonema pseudocostatum | Thalassiosira pseudonana | Thalassiosira weissflogii | Chaetoceros muelleri | Chaetoceros calcitrans | Nannochloropsis oceanica |
|---|---|---|---|---|---|---|---|---|---|---|
| CS-177 | CS-186 | CS-5 | CS-29 | CS-252 | CS-173 | CS-871 | CS-176 | CS-178 | CS-179 | |
| 14:0 | 16.8 ± 2.6 | 10.2 ± 5.2 | 8.1 ± 0.1 | 5.6 ± 0.9 | 14.3 ± 7.6 | 11.3 ± 4.5 | 6.9 ± 5.4 | 10.1 ± 1.2 | 13.1 ± 4.9 | 5.3 ± 0.6 |
| C16 PUFA | 0.3 ± 0.1 | 0.1 ± 0.1 | 14.2 ± 10.1 | 10.8 ± 7.7 | 11.8 ± 5.9 | 13.0 ± 9.2 | 25.8 ± 18.2 | 21.3 ± 10.6 | 13.0 ± 7.5 | 0.8 ± 0.3 |
| 16:1ω7c | 8.3 ± 2.1 | 3.5 ± 2.5 | 16.4 ± 11.6 | 19.4 ± 13.7 | 36.4 ± 18.2 | 27.8 ± 19.6 | 15.1 ± 10.7 | 24.5 ± 12.3 | 26.9 ± 15.5 | 20.2 ± 7.6 |
| 16:0 | 11.4 ± 2.2 | 17 ± 6.1 | 8.3 ± 1.5 | 11.6 ± 2.2 | 8.2 ± 8.1 | 10.0 ± 1.9 | 8.1 ± 2.4 | 15.1 ± 7.3 | 7.2 ± 3.0 | 21.8 ± 6.7 |
| 18:3ω6 + 18:5ω3 | 2.6 ± 0.7 | 6.6 ± 4.7 | 0.6 ± 0.4 | 0.5 ± 0.4 | 0.4 ± 0.2 | 0.1 ± 0.1 | 1.0 ± 0.7 | 0.5 ± 0.2 | 0.1 ± 0.0 | 1.0 ± 0.4 |
| 18:4ω3 | 21.0 ± 4.4 | 15.4 ± 10.9 | 3.2 ± 0.4 | 0.4 ± 0.5 | 0.9 ± 0.3 | 4.3 ± 1.5 | 0.6 ± 0.7 | 0.9 ± 0.6 | 0.7 ± 0.3 | 0.1 ± 0.1 |
| 18:2ω6 | 4.4 ± 3.9 | 3.1 ± 2 | 0.6 ± 0.1 | 1.3 ± 0.5 | 0.7 ± 0.5 | 0.4 ± 0.1 | 2.1 ± 2.1 | 0.6 ± 0.1 | 0.5 ± 0.3 | 1.9 ± 0.9 |
| 18:3ω3 | 5.5 ± 1.7 | 5.2 ± 3.7 | Tr | 0.2 ± 0.1 | 0.6 ± 0.4 | 0.2 ± 0.1 | 0.1 ± 0.2 | 0.1 ± 0.1 | 0.3 ± 0.3 | 0.1 ± 0.0 |
| 18:1ω9c | 7.7 ± 1.9 | 11.6 ± 8.2 | 0.6 ± 0.4 | 5.6 ± 3.9 | 3.6 ± 1.8 | 1.0 ± 0.7 | 2.2 ± 1.5 | 0.6 ± 0.3 | 0.5 ± 0.3 | 6.1 ± 2.3 |
| 18:1ω7c | 1.8 ± 0.5 | 4.1 ± 2.9 | 0.1 ± 0.0 | 0.3 ± 0.2 | 1.0 ± 0.5 | 0.2 ± 0.1 | 0.3 ± 0.2 | 0.8 ± 0.4 | 0.2 ± 0.1 | 0.3 ± 0.1 |
| 18:0 | 0.3 ± 0.2 | 4.1 ± 4.1 | 0.2 ± 0.1 | 0.4 ± 0.2 | 0.3 ± 0.3 | 0.7 ± 0.1 | 2.2 ± 3.1 | 1.9 ± 2 | 0.4 ± 0.4 | 0.4 ± 0.2 |
| 20:4ω6 ARA | 0.1 ± 0.1 | Tr | 4.8 ± 1.2 | 1.2 ± 0.5 | 0.5 ± 0.6 | 0.2 ± 0.1 | 1.3 ± 1.1 | 10.6 ± 10.7 | 2.3 ± 2.9 | 6.5 ± 1.5 |
| 20:5ω3 EPA | 0.6 ± 0.2 | 0.5 ± 0.1 | 26.9 ± 19.0 | 29.0 ± 7.8 | 18.0 ± 4.1 | 23.9 ± 6.1 | 17.2 ± 9.9 | 0.7 ± 0.3 | 21.2 ± 11.3 | 30.0 ± 7.4 |
| 22:6ω3 DHA | 11.1 ± 2.8 | 10.1 ± 0.8 | 3.4 ± 1.4 | 1.7 ± 1.2 | 3.3 ± 1.3 | 3.5 ± 0.6 | 6.2 ± 3.5 | 0.5 ± 0.2 | 0.8 ± 0.0 | Tr |
| ∑SFA | 28.5 | 31.3 | 16.5 | 17.6 | 22.9 | 22.0 | 17.2 | 27.2 | 20.7 | 27.5 |
| ∑MUFA | 17.8 | 19.2 | 17.0 | 25.3 | 41.0 | 29.0 | 17.5 | 25.9 | 27.6 | 26.6 |
| ∑PUFA | 45.4 | 41.1 | 53.8 | 45.1 | 36.1 | 45.7 | 54.5 | 35.1 | 38.9 | 40.4 |
| ∑ω-3LC-PUFA | 11.7 | 10.6 | 30.3 | 30.7 | 21.3 | 27.4 | 23.5 | 1.1 | 22.0 | 30.0 |
| Wax ester | 0.0 | 2.2 | 0.9 | 0.4 | 0.3 | 0.4 | 0.8 | 0.2 | Tr | Tr |
| Triacylglycerol | 1.8 | 2.5 | 2.2 | 1.4 | Tr | 1.8 | 1.6 | 2.0 | Tr | Tr |
| Free fatty acid | 3.0 | 0.0 | 8.3 | 0.0 | 6.3 | 5.1 | 1.8 | 5.7 | 2.5 | 2.5 |
| Sterol | 0.9 | 0.9 | 1.5 | 0.9 | 1.7 | 1.3 | 3.0 | 1.3 | 1.1 | 1.1 |
| Polar lipids | 94.3 | 94.4 | 87.2 | 97.3 | 91.6 | 91.4 | 92.7 | 90.8 | 96.3 | 96.3 |
| Total lipids mg/g | 90.3 | 61.1 | 28.6 | 45.5 | 43.5 | 16.1 | 16.1 | 43.7 | 51.9 | 55.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mai, T.D.; Lee-Chang, K.J.; Jameson, I.D.; Hoang, T.; Cai, N.B.A.; Pham, H.Q. Fatty Acid Profiles of Selected Microalgae Used as Live Feeds for Shrimp Postlarvae in Vietnam. Aquac. J. 2021, 1, 26-38. https://doi.org/10.3390/aquacj1010004
Mai TD, Lee-Chang KJ, Jameson ID, Hoang T, Cai NBA, Pham HQ. Fatty Acid Profiles of Selected Microalgae Used as Live Feeds for Shrimp Postlarvae in Vietnam. Aquaculture Journal. 2021; 1(1):26-38. https://doi.org/10.3390/aquacj1010004
Chicago/Turabian StyleMai, Thao Duc, Kim Jye Lee-Chang, Ian D. Jameson, Tung Hoang, Ngoc Bao Anh Cai, and Hung Quoc Pham. 2021. "Fatty Acid Profiles of Selected Microalgae Used as Live Feeds for Shrimp Postlarvae in Vietnam" Aquaculture Journal 1, no. 1: 26-38. https://doi.org/10.3390/aquacj1010004
APA StyleMai, T. D., Lee-Chang, K. J., Jameson, I. D., Hoang, T., Cai, N. B. A., & Pham, H. Q. (2021). Fatty Acid Profiles of Selected Microalgae Used as Live Feeds for Shrimp Postlarvae in Vietnam. Aquaculture Journal, 1(1), 26-38. https://doi.org/10.3390/aquacj1010004