
Identification of Putative Causal Relationships between Blood-Based Biomarkers and Prediabetes-Induced Senescence: A Comprehensive Review


Abstract
1. Introduction
2. Age-Related Changes in Plasma Biochemistry and Vascular Dynamics in Prediabetes
2.1. Prediabetes as a Precursor of Age-Related Vascular Changes
2.2. Altered Plasma Biochemistry and Implications for Senescence
2.3. Role of Biomarkers in Plasma Biochemistry Changes
2.3.1. Telomere Length
2.3.2. p16INK4a
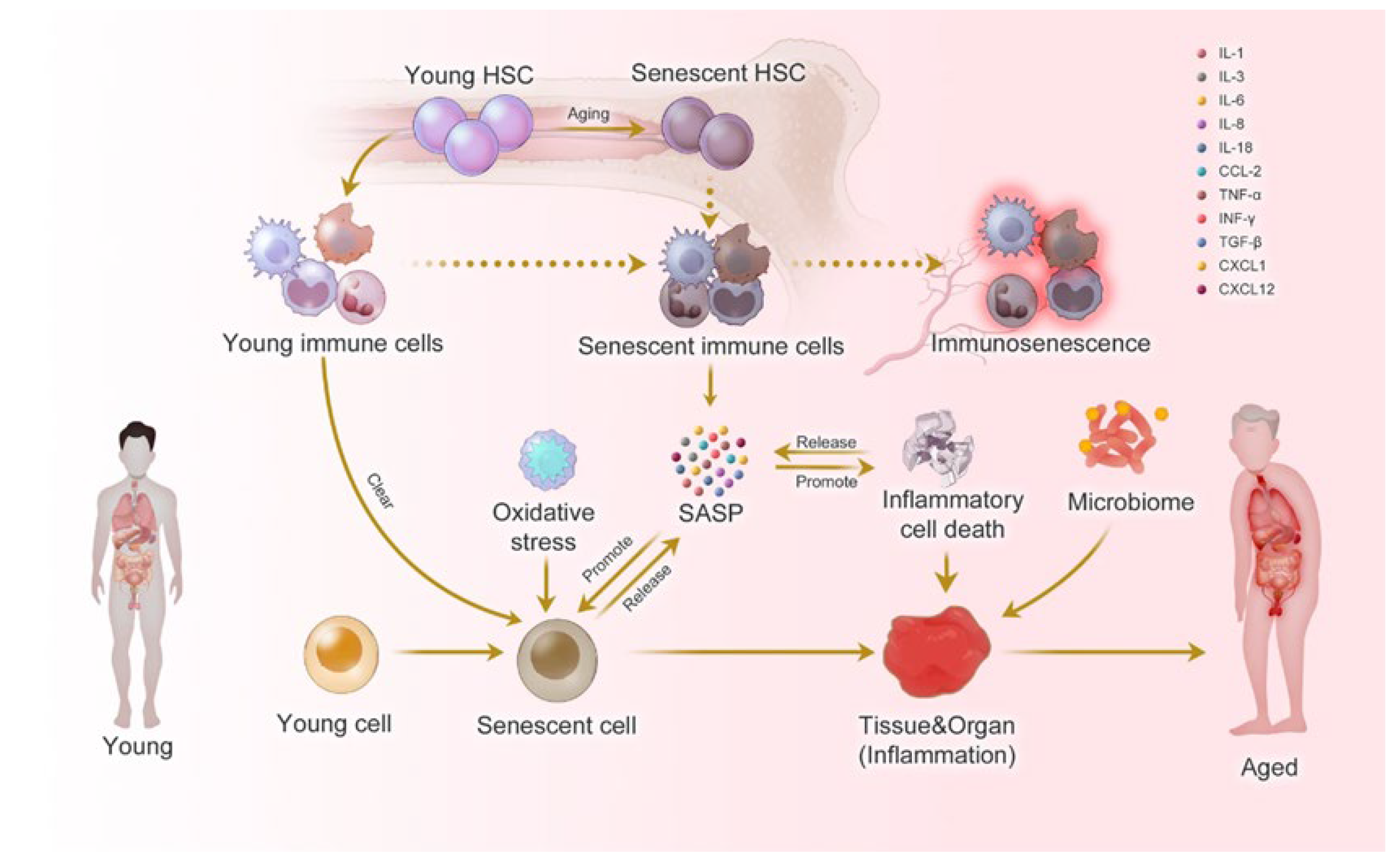
2.3.3. Senescence-Associated Secretory Phenotype (SASP) Factors
2.3.4. DNA Methylation Clocks
2.3.5. Advanced Glycation End Products (AGEs)
3. Circulating Hormones and Growth Factors Associated with Aging in Prediabetes
3.1. Hormonal Shifts in Prediabetes and Their Influence on Senescence
3.2. Growth Factors and Their Role in Age-Associated Processes
3.3. Role of Biomarkers in Hormonal Changes
3.3.1. Circulating Growth Hormone (GH) and Insulin-like Growth Factor 1 (IGF-1)
3.3.2. Dehydroepiandrosterone Sulfate (DHEA-S)
3.3.3. Testosterone (In Men)
3.3.4. Oestrogen (In Women)
4. Age-Associated Inflammatory Factors in Prediabetes
4.1. Inflammatory Mediators in Prediabetes-Induced Senescence
4.2. Chronic Inflammation and Its Implications for Senescence
4.3. Role of Biomarkers in Inflammatory Factors
4.3.1. Senescence-Associated Secretory Phenotype (SASP) Factors
4.3.2. Inflammatory Markers
5. Vascular and Neural System Aging in Patients with Prediabetes
5.1. Prediabetes-Induced Vascular Changes and Senescence
5.2. Neural System Aging and Cognitive Implications in Prediabetes
5.3. Role of Biomarkers in Vascular and Neural Aging
5.3.1. Advanced Glycation End Products (AGEs)
5.3.2. Endothelial Dysfunction
6. Systemic Inflammation in Prediabetes
6.1. Link between Prediabetes and Systemic Inflammation
6.2. Inflammatory Factors and Their Contribution to Senescence
6.3. Role of Biomarkers in Systemic Inflammation
6.3.1. Inflammatory Markers (Continued)
6.3.2. Oxidative Stress Markers
7. Regeneration and Metabolic Disorders in Prediabetes
7.1. Impaired Regeneration Mechanisms in Patients with Prediabetes
7.2. Metabolic Disorders and Their Impact on Senescence
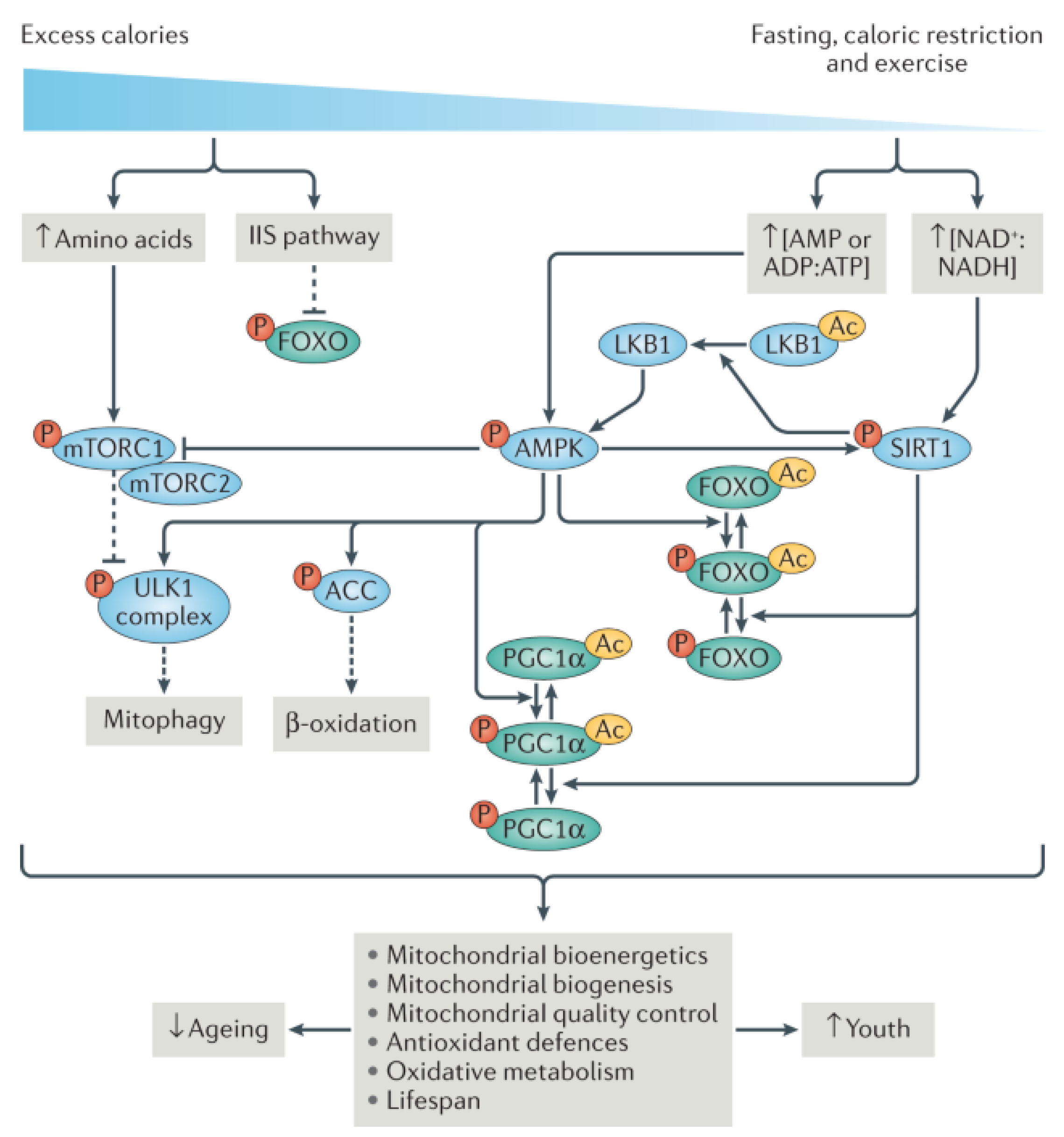
7.2.1. Mitochondrial Dysfunction
7.2.2. Red Blood Cell Distribution Width (RDW)
7.2.3. Hemoglobin A1c (HbA1c)
7.2.4. Serum Albumin
7.2.5. Folate and B12
7.2.6. Osteocalcin
7.2.7. Adiponectin
7.2.8. Leptin
7.2.9. Brain-Derived Neurotrophic Factor (BDNF)
7.2.10. IGF-Binding Proteins
7.2.11. Homocysteine
7.2.12. Insulin Resistance Markers
7.2.13. N-Terminal Pro-B-Type Natriuretic Peptide (NT-proBNP)
7.3. The Negative Effects of Senescence and Pre-Diabetes on Renal Function and Association with Early Stages of Chronic Kidney Disease
8. Perspectives for Future Research
8.1. Gaps in Current Understanding and the Need for Further Research
8.2. Potential Blood-Based Biomarkers and Intervention Strategies
9. Conclusions and Insights
9.1. Summarizing the Key Findings on Blood-Based Biomarkers in Prediabetes-Induced Senescence
9.2. Implications for Understanding Senescence and Aging-related Disorders in Prediabetes
9.3. Future Directions for Research Incorporating Diverse Biomarkers in Prediabetes-Induced Senescence
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Echouffo-Tcheugui, J.B.; Selvin, E. Prediabetes and What It Means: The Epidemiological Evidence. Annu. Rev. Public Health 2021, 42, 59–77. [Google Scholar] [CrossRef] [PubMed]
- Nolan, C.J.; Prentki, M. Insulin resistance and insulin hypersecretion in the metabolic syndrome and type 2 diabetes: Time for a conceptual framework shift. Diabetes Vasc. Dis. Res. 2019, 16, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gómez, M.E.; Zapico, S.C. Frailty, cognitive decline, neurodegenerative diseases, and nutrition interventions. Int. J. Mol. Sci. 2019, 20, 2842. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Han, X.; Zhang, X.; Wang, S. Spatiotemporal evolution of global population aging from 1960 to 2017. BMC Public Health 2019, 19, 127. [Google Scholar]
- Rizos, E.C.; Kanellopoulou, A.; Filis, P.; Markozannes, G.; Chaliasos, K.; Ntzani, E.E.; Tzamouranou, A.; Tentolouris, N. and Tsilidis, K.K. Difference on glucose profile from continuous glucose monitoring in people with prediabetes vs. normoglycemic individuals: A matched-pair analysis. J. diabetes Sci. Technol. 2024, 18, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.M.M.; Chua, Z.J.Y.; Tan, J.C.; Yang, Y.; Liao, Z.; Zhao, Y. From pre-diabetes to diabetes: Diagnosis, treatments and translational research. Medicina 2019, 55, 546. [Google Scholar] [CrossRef] [PubMed]
- Rybtsova, N.; Berezina, T.; Kagansky, A.; Rybtsov, S. Can blood-circulating factors unveil and delay your biological aging? Biomedicines 2020, 8, 615. [Google Scholar] [CrossRef] [PubMed]
- Dorcely, B.; Katz, K.; Jagannathan, R.; Chiang, S.S.; Oluwadare, B.; Goldberg, I.J.; Bergman, M. Novel biomarkers for prediabetes, diabetes, and associated complications. Diabetes Metab. Syndr. Obes. 2017, 10, 345–361. [Google Scholar] [CrossRef] [PubMed]
- Pyo, I.S.; Yun, S.; Yoon, Y.E.; Choi, J.-W.; Lee, S.-J. Mechanisms of aging and the preventive effects of resveratrol on age-related diseases. Molecules 2020, 25, 4649. [Google Scholar] [CrossRef]
- De Biasi, A.; Wolfe, M.; Carmody, J.; Fulmer, T.; Auerbach, J. Creating an age-friendly public health system. Innov. Aging 2020, 4, igz044. [Google Scholar] [CrossRef]
- Zhang, K.; Ma, Y.; Luo, Y.; Song, Y.; Xiong, G.; Ma, Y.; Sun, X.; Kan, C. Metabolic diseases and healthy aging: Identifying environmental and behavioral risk factors and promoting public health. Front. Public Health 2023, 11, 1253506. [Google Scholar] [CrossRef] [PubMed]
- Bahour, N.; Cortez, B.; Pan, H.; Shah, H.; Doria, A.; Aguayo-Mazzucato, C. Diabetes mellitus correlates with increased biological age as indicated by clinical biomarkers. Geroscience 2022, 44, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Han, J.; Jiang, C.; Zhang, Y. Biomarkers, oxidative stress and autophagy in skin aging. Aging Res. Rev. 2020, 59, 101036. [Google Scholar] [CrossRef] [PubMed]
- Chia, C.W.; Egan, J.M.; Ferrucci, L. Age-Related Changes in Glucose Metabolism, Hyperglycemia, and Cardiovascular Risk. Circ. Res. 2018, 123, 886–904. [Google Scholar] [CrossRef] [PubMed]
- Mota, R.I.; Morgan, S.E.; Bahnson, E.M. Diabetic vasculopathy: Macro and microvascular injury. Curr. Pathobiol. Rep. 2020, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Fhayli, W.; Boëté, Q.; Harki, O.; Briançon-Marjollet, A.; Jacob, M.-P.; Faury, G. Rise and fall of elastic fibers from development to aging. Consequences on arterial structure-function and therapeutical perspectives. Matrix Biol. 2019, 84, 41–56. [Google Scholar] [CrossRef]
- Wang, C.; Yuan, Y.; Zheng, M.; Pan, A.; Wang, M.; Zhao, M.; Li, Y.; Yao, S.; Chen, S.; Wu, S. Association of age of onset of hypertension with cardiovascular diseases and mortality. J. Am. Coll. Cardiol. 2020, 75, 2921–2930. [Google Scholar] [CrossRef] [PubMed]
- Brannick, B.; Dagogo-Jack, S. Prediabetes and Cardiovascular Disease: Pathophysiology and Interventions for Prevention and Risk Reduction. Endocrinol. Metab. Clin. N. Am. 2018, 47, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Luc, K.; Schramm-Luc, A.; Guzik, T.; Mikolajczyk, T. Oxidative stress and inflammatory markers in prediabetes and diabetes. J. Physiol. Pharmacol. 2019, 70, 809–824. [Google Scholar]
- Izzo, C.; Vitillo, P.; Di Pietro, P.; Visco, V.; Strianese, A.; Virtuoso, N.; Ciccarelli, M.; Galasso, G.; Carrizzo, A.; Vecchione, C. The role of oxidative stress in cardiovascular aging and cardiovascular diseases. Life 2021, 11, 60. [Google Scholar] [CrossRef]
- Dilworth, L.; Facey, A.; Omoruyi, F. Diabetes Mellitus and Its Metabolic Complications: The Role of Adipose Tissues. Int. J. Mol. Sci. 2021, 22, 7644. [Google Scholar] [CrossRef] [PubMed]
- Foretz, M.; Guigas, B.; Viollet, B. Understanding the glucoregulatory mechanisms of metformin in type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2019, 15, 569–589. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, M.R.; Chaudhary, S.; Sharma, Y.; Singh, T.A.; Mishra, A.K.; Sharma, S.; Mehdi, M.M. Aging, oxidative stress and degenerative diseases: Mechanisms, complications and emerging therapeutic strategies. Biogerontology 2023, 24, 609–662. [Google Scholar] [CrossRef] [PubMed]
- Gianazza, E.; Brioschi, M.; Baetta, R.; Mallia, A.; Banfi, C.; Tremoli, E. Platelets in healthy and disease states: From biomarkers discovery to drug targets identification by proteomics. Int. J. Mol. Sci. 2020, 21, 4541. [Google Scholar] [CrossRef]
- Aguayo-Mazzucato, C.; Andle, J.; Lee, T.B.; Midha, A.; Talemal, L.; Chipashvili, V.; Hollister-Lock, J.; Van Deursen, J.; Weir, G.; Bonner-Weir, S. Acceleration of β cell aging determines diabetes and senolysis improves disease outcomes. Cell Metab. 2019, 30, 129–142.e124. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Hossain, K.S.; Das, S.; Kundu, S.; Adegoke, E.O.; Rahman, M.A.; Hannan, M.A.; Uddin, M.J.; Pang, M.-G. Role of insulin in health and disease: An update. Int. J. Mol. Sci. 2021, 22, 6403. [Google Scholar] [CrossRef] [PubMed]
- Derrick, S.A.; Kristo, A.S.; Reaves, S.K.; Sikalidis, A.K. Effects of dietary red raspberry consumption on pre-diabetes and type 2 diabetes mellitus parameters. Int. J. Environ. Res. Public Health 2021, 18, 9364. [Google Scholar] [CrossRef] [PubMed]
- Ellulu, M.S.; Samouda, H. Clinical and biological risk factors associated with inflammation in patients with type 2 diabetes mellitus. BMC Endocr. Disord. 2022, 22, 16. [Google Scholar] [CrossRef] [PubMed]
- Barbu, E.; Popescu, M.-R.; Popescu, A.-C.; Balanescu, S.-M. Inflammation as a precursor of atherothrombosis, diabetes and early vascular aging. Int. J. Mol. Sci. 2022, 23, 963. [Google Scholar] [CrossRef]
- Li, X.; Li, C.; Zhang, W.; Wang, Y.; Qian, P.; Huang, H. Inflammation and aging: Signaling pathways and intervention therapies. Signal Transduct. Target. Ther. 2023, 8, 239. [Google Scholar] [CrossRef]
- Wan, M.; Gray-Gaillard, E.F.; Elisseeff, J.H. Cellular senescence in musculoskeletal homeostasis, diseases, and regeneration. Bone Res. 2021, 9, 41. [Google Scholar] [CrossRef] [PubMed]
- Shahwan, M.J.; Jairoun, A.A.; Farajallah, A.; Shanabli, S. Prevalence of dyslipidemia and factors affecting lipid profile in patients with type 2 diabetes. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 2387–2392. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.T.; Fernando, S.; Schwarz, N.; Tan, J.T.; Bursill, C.A.; Psaltis, P.J. Inflammation as a therapeutic target in atherosclerosis. J. Clin. Med. 2019, 8, 1109. [Google Scholar] [CrossRef]
- Honda, M. Reverse Heart Disease Naturally: Cures for High Cholesterol, Hypertension, Arteriosclerosis, Blood Clots, Aneurysms, Myocardial Infarcts and More; Hatherleigh Press: Hobart, NY, USA, 2019. [Google Scholar]
- Gilon, P. The role of α-cells in islet function and glucose homeostasis in health and type 2 diabetes. J. Mol. Biol. 2020, 432, 1367–1394. [Google Scholar] [CrossRef] [PubMed]
- Dozio, E.; Massaccesi, L.; Corsi Romanelli, M.M. Glycation and Glycosylation in Cardiovascular Remodeling: Focus on Advanced Glycation End Products and O-Linked Glycosylations as Glucose-Related Pathogenetic Factors and Disease Markers. J. Clin. Med. 2021, 10, 4792. [Google Scholar] [CrossRef]
- Kosmopoulos, M.; Drekolias, D.; Zavras, P.D.; Piperi, C.; Papavassiliou, A.G. Impact of advanced glycation end products (AGEs) signaling in coronary artery disease. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2019, 1865, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, Z.; Ren, Y.; Wang, Y.; Fang, J.; Yue, H.; Ma, S.; Guan, F. Aging and age-related diseases: From mechanisms to therapeutic strategies. Biogerontology 2021, 22, 165–187. [Google Scholar] [CrossRef] [PubMed]
- Banday, M.Z.; Sameer, A.S.; Nissar, S. Pathophysiology of diabetes: An overview. Avicenna J. Med. 2020, 10, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Biessels, G.J.; Nobili, F.; Teunissen, C.E.; Simó, R.; Scheltens, P. Understanding multifactorial brain changes in type 2 diabetes: A biomarker perspective. Lancet Neurol. 2020, 19, 699–710. [Google Scholar] [CrossRef]
- Kudryashova, K.S.; Burka, K.; Kulaga, A.Y.; Vorobyeva, N.S.; Kennedy, B.K. Aging biomarkers: From functional tests to multi-omics approaches. Proteomics 2020, 20, 1900408. [Google Scholar] [CrossRef]
- Zheng, L.; Meng, Y.; Campbell, J.L.; Shen, B. Multiple roles of DNA2 nuclease/helicase in DNA metabolism, genome stability and human diseases. Nucleic Acids Res. 2020, 48, 16–35. [Google Scholar] [CrossRef] [PubMed]
- Kamal, N.S.M.; Safuan, S.; Shamsuddin, S.; Foroozandeh, P. Aging of the cells: Insight into cellular senescence and detection Methods. Eur. J. Cell Biol. 2020, 99, 151108. [Google Scholar]
- Gavia-García, G.; Rosado-Pérez, J.; Arista-Ugalde, T.L.; Aguiñiga-Sánchez, I.; Santiago-Osorio, E.; Mendoza-Núñez, V.M. Telomere length and oxidative stress and its relation with metabolic syndrome components in the aging. Biology 2021, 10, 253. [Google Scholar] [CrossRef] [PubMed]
- Tam, B.T.; Morais, J.A.; Santosa, S. Obesity and aging: Two sides of the same coin. Obes. Rev. 2020, 21, e12991. [Google Scholar] [CrossRef] [PubMed]
- Safwan-Zaiter, H.; Wagner, N.; Wagner, K.-D. P16INK4A—More Than a Senescence Marker. Life 2022, 12, 1332. [Google Scholar] [CrossRef]
- Mbatha, N.A.; Mzimela, N.C.; Mushebenge, A.G.-A.; Khathi, A. Investigating the Impact of Prediabetes on the Aging Process through Longitudinal Analysis of Blood-Based Biomarkers: A Systematic Review and Meta-Analysis Protocol. medRxiv 2023. [Google Scholar] [CrossRef]
- Yi, H.-S.; Kim, S.Y.; Kim, J.T.; Lee, Y.-S.; Moon, J.S.; Kim, M.; Kang, Y.E.; Joung, K.H.; Lee, J.H.; Kim, H.J. T-cell senescence contributes to abnormal glucose homeostasis in humans and mice. Cell Death Dis. 2019, 10, 249. [Google Scholar] [CrossRef]
- Xiong, Y.; Zhou, L. The signaling of cellular senescence in diabetic nephropathy. Oxidative Med. Cell. Longev. 2019, 2019, 7495629. [Google Scholar] [CrossRef]
- Li, N.; Liu, F.; Yang, P.; Xiong, F.; Yu, Q.; Li, J.; Zhou, Z.; Zhang, S.; Wang, C.-Y. Aging and stress induced β cell senescence and its implication in diabetes development. Aging 2019, 11, 9947. [Google Scholar] [CrossRef]
- Cianflone, E.; Torella, M.; Biamonte, F.; De Angelis, A.; Urbanek, K.; Costanzo, F.S.; Rota, M.; Ellison-Hughes, G.M.; Torella, D. Targeting cardiac stem cell senescence to treat cardiac aging and disease. Cells 2020, 9, 1558. [Google Scholar] [CrossRef]
- Lopes-Paciencia, S.; Saint-Germain, E.; Rowell, M.-C.; Ruiz, A.F.; Kalegari, P.; Ferbeyre, G. The senescence-associated secretory phenotype and its regulation. Cytokine 2019, 117, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Lagoumtzi, S.M.; Chondrogianni, N. Senolytics and senomorphics: Natural and synthetic therapeutics in the treatment of aging and chronic diseases. Free Radic. Biol. Med. 2021, 171, 169–190. [Google Scholar] [CrossRef] [PubMed]
- Khosla, S.; Samakkarnthai, P.; Monroe, D.G.; Farr, J.N. Update on the pathogenesis and treatment of skeletal fragility in type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2021, 17, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Raucci, A.; Macrì, F.; Castiglione, S.; Badi, I.; Vinci, M.C.; Zuccolo, E. MicroRNA-34a: The bad guy in age-related vascular diseases. Cell. Mol. Life Sci. 2021, 78, 7355–7378. [Google Scholar] [CrossRef] [PubMed]
- Teissier, T.; Boulanger, E.; Cox, L.S. Interconnections between inflammaging and immunosenescence during aging. Cells 2022, 11, 359. [Google Scholar] [CrossRef]
- Khosla, S.; Farr, J.N.; Tchkonia, T.; Kirkland, J.L. The role of cellular senescence in aging and endocrine disease. Nat. Rev. Endocrinol. 2020, 16, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Tracy, E.P.; Hughes, W.; Beare, J.E.; Rowe, G.; Beyer, A.; LeBlanc, A.J. Aging-induced impairment of vascular function: Mitochondrial redox contributions and physiological/clinical implications. Antioxid. Redox Signal. 2021, 35, 974–1015. [Google Scholar] [CrossRef] [PubMed]
- Unnikrishnan, A.; Freeman, W.M.; Jackson, J.; Wren, J.D.; Porter, H.; Richardson, A. The role of DNA methylation in epigenetics of aging. Pharmacol. Ther. 2019, 195, 172–185. [Google Scholar] [CrossRef] [PubMed]
- Salameh, Y.; Bejaoui, Y.; El Hajj, N. DNA methylation biomarkers in aging and age-related diseases. Front. Genet. 2020, 11, 171. [Google Scholar] [CrossRef]
- Samblas, M.; Milagro, F.I.; Martínez, A. DNA methylation markers in obesity, metabolic syndrome, and weight loss. Epigenetics 2019, 14, 421–444. [Google Scholar] [CrossRef]
- Maldonado, E.; Morales-Pison, S.; Urbina, F.; Solari, A. Aging Hallmarks and the Role of Oxidative Stress. Antioxidants 2023, 12, 651. [Google Scholar] [CrossRef]
- Barkabi-Zanjani, S.; Ghorbanzadeh, V.; Aslani, M.; Ghalibafsabbaghi, A.; Chodari, L. Diabetes mellitus and the impairment of male reproductive function: Possible signaling pathways. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 14, 1307–1314. [Google Scholar] [CrossRef] [PubMed]
- Twarda-Clapa, A.; Olczak, A.; Białkowska, A.M.; Koziołkiewicz, M. Advanced glycation end-products (AGEs): Formation, chemistry, classification, receptors, and diseases related to AGEs. Cells 2022, 11, 1312. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef] [PubMed]
- Basisty, N.; Kale, A.; Jeon, O.H.; Kuehnemann, C.; Payne, T.; Rao, C.; Holtz, A.; Shah, S.; Sharma, V.; Ferrucci, L. A proteomic atlas of senescence-associated secretomes for aging biomarker development. PLoS Biol. 2020, 18, e3000599. [Google Scholar] [CrossRef]
- Wang, K.; Liu, H.; Hu, Q.; Wang, L.; Liu, J.; Zheng, Z.; Zhang, W.; Ren, J.; Zhu, F.; Liu, G.-H. Epigenetic regulation of aging: Implications for interventions of aging and diseases. Signal Transduct. Target. Ther. 2022, 7, 374. [Google Scholar] [CrossRef]
- Haywood, N.J.; Slater, T.A.; Matthews, C.J.; Wheatcroft, S.B. The insulin like growth factor and binding protein family: Novel therapeutic targets in obesity & diabetes. Mol. Metab. 2019, 19, 86–96. [Google Scholar]
- Barclay, R.D.; Burd, N.A.; Tyler, C.; Tillin, N.A.; Mackenzie, R.W. The role of the IGF-1 signaling cascade in muscle protein synthesis and anabolic resistance in aging skeletal muscle. Front. Nutr. 2019, 6, 146. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chi, X.; Wang, Y.; Setrerrahmane, S.; Xie, W.; Xu, H. Trends in insulin resistance: Insights into mechanisms and therapeutic strategy. Signal Transduct. Target. Ther. 2022, 7, 216. [Google Scholar] [CrossRef]
- Koffi, K.A.; Doublier, S.; Ricort, J.-M.; Babajko, S.; Nassif, A.; Isaac, J. The role of GH/IGF axis in dento-alveolar complex from development to aging and therapeutics: A narrative review. Cells 2021, 10, 1181. [Google Scholar] [CrossRef]
- Ehtewish, H.; Arredouani, A.; El-Agnaf, O. Diagnostic, prognostic, and mechanistic biomarkers of diabetes mellitus-associated cognitive decline. Int. J. Mol. Sci. 2022, 23, 6144. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Parihar, A.; Talaiya, T.; Dubey, K.; Porwal, B.; Parihar, M.S. Cognitive impairments in type 2 diabetes, risk factors and preventive strategies. J. Basic Clin. Physiol. Pharmacol. 2020, 31, 20190105. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, P.; Kiss, T.; Tarantini, S.; Nyúl-Tóth, Á.; Ahire, C.; Yabluchanskiy, A.; Csipo, T.; Lipecz, A.; Tabak, A.; Institoris, A. Obesity-induced cognitive impairment in older adults: A microvascular perspective. Am. J. Physiol.-Heart Circ. Physiol. 2021, 320, H740–H761. [Google Scholar] [CrossRef] [PubMed]
- Wondmkun, Y.T. Obesity, insulin resistance, and type 2 diabetes: Associations and therapeutic implications. Diabetes Metab. Syndr. Obes. 2020, 13, 3611–3616. [Google Scholar] [CrossRef] [PubMed]
- Kuryłowicz, A.; Koźniewski, K. Anti-inflammatory strategies targeting metaflammation in type 2 diabetes. Molecules 2020, 25, 2224. [Google Scholar] [CrossRef] [PubMed]
- Ou, M.-Y.; Zhang, H.; Tan, P.-C.; Zhou, S.-B.; Li, Q.-F. Adipose tissue aging: Mechanisms and therapeutic implications. Cell Death Dis. 2022, 13, 300. [Google Scholar] [CrossRef] [PubMed]
- Salvestrini, V.; Sell, C.; Lorenzini, A. Obesity may accelerate the aging process. Front. Endocrinol. 2019, 10, 266. [Google Scholar] [CrossRef]
- Le Couteur, D.G.; Solon-Biet, S.M.; Cogger, V.C.; Ribeiro, R.; De Cabo, R.; Raubenheimer, D.; Cooney, G.J.; Simpson, S.J. Branched chain amino acids, aging and age-related health. Aging Res. Rev. 2020, 64, 101198. [Google Scholar] [CrossRef] [PubMed]
- Slater, T.; Haywood, N.J.; Matthews, C.; Cheema, H.; Wheatcroft, S.B. Insulin-like growth factor binding proteins and angiogenesis: From cancer to cardiovascular disease. Cytokine Growth Factor Rev. 2019, 46, 28–35. [Google Scholar] [CrossRef]
- Kasprzak, A. Insulin-like growth factor 1 (IGF-1) signaling in glucose metabolism in colorectal cancer. Int. J. Mol. Sci. 2021, 22, 6434. [Google Scholar] [CrossRef]
- Barone, E.; Di Domenico, F.; Perluigi, M.; Butterfield, D.A. The interplay among oxidative stress, brain insulin resistance and AMPK dysfunction contribute to neurodegeneration in type 2 diabetes and Alzheimer disease. Free Radic. Biol. Med. 2021, 176, 16–33. [Google Scholar] [CrossRef] [PubMed]
- Forcina, L.; Miano, C.; Scicchitano, B.M.; Musarò, A. Signals from the niche: Insights into the role of IGF-1 and IL-6 in modulating skeletal muscle fibrosis. Cells 2019, 8, 232. [Google Scholar] [CrossRef] [PubMed]
- Fadini, G.P.; Albiero, M.; Bonora, B.M.; Avogaro, A. Angiogenic abnormalities in diabetes mellitus: Mechanistic and clinical aspects. J. Clin. Endocrinol. Metab. 2019, 104, 5431–5444. [Google Scholar] [CrossRef]
- Schalkwijk, C.; Stehouwer, C. Methylglyoxal, a highly reactive dicarbonyl compound, in diabetes, its vascular complications, and other age-related diseases. Physiol. Rev. 2020, 100, 407–461. [Google Scholar] [CrossRef]
- Di Micco, R.; Krizhanovsky, V.; Baker, D.; D’Adda di Fagagna, F. Cellular senescence in aging: From mechanisms to therapeutic opportunities. Nat. Rev. Mol. Cell Biol. 2021, 22, 75–95. [Google Scholar] [CrossRef] [PubMed]
- Mancinelli, R.; Checcaglini, F.; Coscia, F.; Gigliotti, P.; Fulle, S.; Fanò-Illic, G. Biological aspects of selected myokines in skeletal muscle: Focus on aging. Int. J. Mol. Sci. 2021, 22, 8520. [Google Scholar] [CrossRef] [PubMed]
- LeRoith, D.; Holly, J.M.; Forbes, B.E. Insulin-like growth factors: Ligands, binding proteins, and receptors. Mol. Metab. 2021, 52, 101245. [Google Scholar] [CrossRef] [PubMed]
- Bellary, S.; Kyrou, I.; Brown, J.E.; Bailey, C.J. Type 2 diabetes mellitus in older adults: Clinical considerations and management. Nat. Rev. Endocrinol. 2021, 17, 534–548. [Google Scholar] [CrossRef] [PubMed]
- Dichtel, L.E.; Cordoba-Chacon, J.; Kineman, R.D. Growth Hormone and Insulin-Like Growth Factor 1 Regulation of Nonalcoholic Fatty Liver Disease. J. Clin. Endocrinol. Metab. 2022, 107, 1812–1824. [Google Scholar] [CrossRef]
- Biadgo, B.; Tamir, W.; Ambachew, S. Insulin-like Growth Factor and its Therapeutic Potential for Diabetes Complications—Mechanisms and Metabolic Links: A Review. Rev. Diabet. Stud. 2020, 16, 24–34. [Google Scholar] [CrossRef]
- Elpers, A.-L.; Steptoe, A. Associations between dehydroepiandrosterone sulphate (DHEAS) and cognitive function in 5,061 older men and women in the English Longitudinal Study of Aging. Psychoneuroendocrinology 2020, 117, 104702. [Google Scholar] [CrossRef] [PubMed]
- Sinha, N.; Maiti, A.; Sinha, A.; Basu, A.; Das, T.C. Study of serum testosterone level in males with Prediabetes, before and after 3 months of metformin therapy, in patients attending a tertiary care hospital in West Bengal, Eastern India. J. Med. Sci. Clin. Res. 2019, 7, 32–43. [Google Scholar] [CrossRef]
- De Paoli, M.; Zakharia, A.; Werstuck, G.H. The Role of Estrogen in Insulin Resistance: A Review of Clinical and Preclinical Data. Am. J. Pathol. 2021, 191, 1490–1498. [Google Scholar] [CrossRef] [PubMed]
- Van den Beld, A.W.; Kaufman, J.M.; Zillikens, M.C.; Lamberts, S.W.J.; Egan, J.M.; Van der Lely, A.J. The physiology of endocrine systems with aging. Lancet Diabetes Endocrinol. 2018, 6, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Yen, H.H.; Hsieh, S.T.; Chen, C.L.; Yang, W.S.; Lee, P.C.; Lin, M.T.; Chen, C.N.; Yang, P.J. Circulating Diabetic Candidate Neurotrophic Factors, Brain-Derived Neurotrophic Factor and Fibroblast Growth Factor 21, in Sleeve Gastrectomy. Sci. Rep. 2020, 10, 5341. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.B.; Aleksic, S.; Gao, T.; Weiss, E.F.; Demetriou, E.; Verghese, J.; Holtzer, R.; Barzilai, N.; Milman, S. Insulin-like Growth Factor-1 and IGF Binding Proteins Predict All-Cause Mortality and Morbidity in Older Adults. Cells 2020, 9, 1368. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.; Betriu, À.; López-Cano, C.; Hernández, M.; Fernández, E.; Purroy, F.; Bermúdez-López, M.; Farràs-Sallés, C.; Barril, S.; Pamplona, R. Characteristics of atheromatosis in the prediabetes stage: A cross-sectional investigation of the ILERVAS project. Cardiovasc. Diabetol. 2019, 18, 154. [Google Scholar] [CrossRef]
- Mangge, H.; Herrmann, M.; Almer, G.; Zelzer, S.; Moeller, R.; Horejsi, R.; Renner, W. Telomere shortening associates with elevated insulin and nuchal fat accumulation. Sci. Rep. 2020, 10, 6863. [Google Scholar] [CrossRef] [PubMed]
- Ungvari, Z.; Tarantini, S.; Sorond, F.; Merkely, B.; Csiszar, A. Mechanisms of Vascular Aging, A Geroscience Perspective: JACC Focus Seminar. J. Am. Coll. Cardiol. 2020, 75, 931–941. [Google Scholar] [CrossRef]
- Schafer, M.J.; Zhang, X.; Kumar, A.; Atkinson, E.J.; Zhu, Y.; Jachim, S.; Mazula, D.L.; Brown, A.K.; Berning, M.; Aversa, Z.; et al. The senescence-associated secretome as an indicator of age and medical risk. JCI Insight 2020, 5, e133668. [Google Scholar] [CrossRef]
- Reale, A.; Tagliatesta, S.; Zardo, G.; Zampieri, M. Counteracting aged DNA methylation states to combat aging and age-related diseases. Mech. Aging Dev. 2022, 206, 111695. [Google Scholar] [CrossRef] [PubMed]
- Dhindsa, S.; Zhang, N.; McPhaul, M.J.; Wu, Z.; Ghoshal, A.K.; Erlich, E.C.; Mani, K.; Randolph, G.J.; Edwards, J.R.; Mudd, P.A.; et al. Association of Circulating Sex Hormones with Inflammation and Disease Severity in Patients with COVID-19. JAMA Netw. Open 2021, 4, e2111398. [Google Scholar] [CrossRef] [PubMed]
- Berezin, A.E.; Berezin, A.A. Circulating Cardiac Biomarkers in Diabetes Mellitus: A New Dawn for Risk Stratification—A Narrative Review. Diabetes Ther. 2020, 11, 1271–1291. [Google Scholar] [CrossRef] [PubMed]
- Olcha, P.; Winiarska-Mieczan, A.; Kwiecień, M.; Nowakowski, Ł.; Miturski, A.; Semczuk, A.; Kiczorowska, B.; Gałczyński, K. Antioxidative, anti-inflammatory, anti-obesogenic, and antidiabetic properties of tea polyphenols—The positive impact of regular tea consumption as an element of prophylaxis and pharmacotherapy support in endometrial cancer. Int. J. Mol. Sci. 2022, 23, 6703. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhang, S.; Wang, L.; Pan, T.; Zhong, X. The relationship between red blood cell distribution and islet β-cell function indexes in patients with type 2 diabetes. BMC Endocr. Disord. 2021, 21, 7. [Google Scholar] [CrossRef] [PubMed]
- Torres-Zegarra, C.; Sundararajan, D.; Benson, J.; Seagle, H.; Witten, M.; Walders-Abramson, N.; Simon, S.; Huguelet, P.; Nokoff, N.; Cree-Green, M. Care for adolescents with polycystic ovary syndrome: Development and prescribing patterns of a multidisciplinary clinic. J. Pediatr. Adolesc. Gynecol. 2021, 34, 617–625. [Google Scholar] [CrossRef]
- Zhang, X.; Deng, X.; Zhou, J.; Qiu, K.; Deng, M.; Lin, Z.; Mosha, S.S.; Li, W. The association of serum cortisol level with microalbuminuria in patients with type 2 diabetes and prediabetes. Int. J. Med. Sci. 2020, 17, 2998. [Google Scholar] [CrossRef] [PubMed]
- Janssen, J.A. Hyperinsulinemia and its pivotal role in aging, obesity, type 2 diabetes, cardiovascular disease and cancer. Int. J. Mol. Sci. 2021, 22, 7797. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, C.J.; Veras, K.; De Oliveira Carvalho, C.R. Dehydroepiandrosterone on metabolism and the cardiovascular system in the postmenopausal period. J. Mol. Med. 2020, 98, 39–57. [Google Scholar] [CrossRef]
- Tang, J.; Chen, L.-R.; Chen, K.-H. The Utilization of dehydroepiandrosterone as a sexual hormone precursor in premenopausal and postmenopausal women: An overview. Pharmaceuticals 2021, 15, 46. [Google Scholar] [CrossRef]
- Cappola, A.R.; Auchus, R.J.; El-Hajj Fuleihan, G.; Handelsman, D.J.; Kalyani, R.R.; McClung, M.; Stuenkel, C.A.; Thorner, M.O.; Verbalis, J.G. Hormones and Aging: An Endocrine Society Scientific Statement. J. Clin. Endocrinol. Metab. 2023, 108, 1835–1874. [Google Scholar] [CrossRef] [PubMed]
- Salonia, A.; Rastrelli, G.; Hackett, G.; Seminara, S.B.; Huhtaniemi, I.T.; Rey, R.A.; Hellstrom, W.J.; Palmert, M.R.; Corona, G.; Dohle, G.R. Paediatric and adult-onset male hypogonadism. Nat. Rev. Dis. Primers 2019, 5, 38. [Google Scholar] [CrossRef]
- Hur, M.-H.; Hong, J.H.; Yeo, S. Effects of aromatherapy on stress, fructosamine, fatigue, and sleep quality in prediabetic middle-aged women: A randomised controlled trial. Eur. J. Integr. Med. 2019, 31, 100978. [Google Scholar] [CrossRef]
- Bartz, D.; Chitnis, T.; Kaiser, U.B.; Rich-Edwards, J.W.; Rexrode, K.M.; Pennell, P.B.; Goldstein, J.M.; O’Neal, M.A.; LeBoff, M.; Behn, M. Clinical advances in sex-and gender-informed medicine to improve the health of all: A review. JAMA Intern. Med. 2020, 180, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Yun, C.; Pang, Y.; Qiao, J. The impact of the gut microbiota on the reproductive and metabolic endocrine system. Gut Microbes 2021, 13, 1894070. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, R.; Parrillo, L.; Longo, M.; Florese, P.; Desiderio, A.; Zatterale, F.; Miele, C.; Raciti, G.A.; Beguinot, F. Molecular basis of aging in chronic metabolic diseases. J. Endocrinol. Investig. 2020, 43, 1373–1389. [Google Scholar] [CrossRef] [PubMed]
- Lavillegrand, J.-R.; Garnier, M.; Spaeth, A.; Mario, N.; Hariri, G.; Pilon, A.; Berti, E.; Fieux, F.; Thietart, S.; Urbina, T. Elevated plasma IL-6 and CRP levels are associated with adverse clinical outcomes and death in critically ill SARS-CoV-2 patients: Inflammatory response of SARS-CoV-2 patients. Ann. Intensive Care 2021, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Berlanga-Acosta, J.A.; Guillén-Nieto, G.E.; Rodríguez-Rodríguez, N.; Mendoza-Mari, Y.; Bringas-Vega, M.L.; Berlanga-Saez, J.O.; García del Barco Herrera, D.; Martinez-Jimenez, I.; Hernandez-Gutierrez, S.; Valdés-Sosa, P.A. Cellular senescence as the pathogenic hub of diabetes-related wound chronicity. Front. Endocrinol. 2020, 11, 573032. [Google Scholar] [CrossRef] [PubMed]
- Oguntibeju, O.O. Type 2 diabetes mellitus, oxidative stress and inflammation: Examining the links. Int. J. Physiol. Pathophysiol. Pharmacol. 2019, 11, 45. [Google Scholar]
- Szűcs, G.; Sója, A.; Péter, M.; Sárközy, M.; Bruszel, B.; Siska, A.; Földesi, I.; Szabó, Z.; Janáky, T.; Vígh, L. Prediabetes induced by fructose-enriched diet influences cardiac lipidome and proteome and leads to deterioration of cardiac function prior to the development of excessive oxidative stress and cell damage. Oxidative Med. Cell. Longev. 2019, 2019, 3218275. [Google Scholar] [CrossRef]
- Peña, A.; Olson, M.L.; Ayers, S.L.; Sears, D.D.; Vega-López, S.; Colburn, A.T.; Shaibi, G.Q. Inflammatory Mediators and Type 2 Diabetes Risk Factors before and in Response to Lifestyle Intervention among Latino Adolescents with Obesity. Nutrients 2023, 15, 2442. [Google Scholar] [CrossRef]
- Michaud, M.; Balardy, L.; Moulis, G.; Gaudin, C.; Peyrot, C.; Vellas, B.; Cesari, M. and Nourhashemi, F. Proinflammatory cytokines, aging, and age-related diseases. JAMDA 2013, 14, 877–882. [Google Scholar]
- Pan, X.; Kaminga, A.C.; Wen, S.W.; Liu, A. Chemokines in Prediabetes and Type 2 Diabetes: A Meta-Analysis. Front. Immunol. 2021, 12, 622438. [Google Scholar] [CrossRef] [PubMed]
- Avramovic, D.; Archaimbault, S.A.; Kemble, A.M.; Gruener, S.; Lazendic, M.; Westenskow, P.D. TGFβ1 Induces Senescence and Attenuated VEGF Production in Retinal Pericytes. Biomedicines 2022, 10, 1404. [Google Scholar] [CrossRef]
- Tan, Q.; Liang, N.; Zhang, X.; Li, J. Dynamic Aging: Channeled Through Microenvironment. Front. Physiol. 2021, 12, 702276. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.; Yang, T.; Chen, H.; Fu, D.; Hu, Y.; Wang, J.; Yuan, Q.; Yu, H.; Xu, W.; Xie, X. New insights into oxidative stress and inflammation during diabetes mellitus-accelerated atherosclerosis. Redox Biol. 2019, 20, 247–260. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Xu, B.-t.; Wan, S.-r.; Ma, X.-m.; Long, Y.; Xu, Y.; Jiang, Z.-z. The role of oxidative stress in diabetes mellitus-induced vascular endothelial dysfunction. Cardiovasc. Diabetol. 2023, 22, 237. [Google Scholar] [CrossRef]
- Khalil, R.; Diab-Assaf, M.; Lemaitre, J.-M. Emerging Therapeutic Approaches to Target the Dark Side of Senescent Cells: New Hopes to Treat Aging as a Disease and to Delay Age-Related Pathologies. Cells 2023, 12, 915. [Google Scholar] [CrossRef]
- Amorim, J.A.; Coppotelli, G.; Rolo, A.P.; Palmeira, C.M.; Ross, J.M.; Sinclair, D.A. Mitochondrial and metabolic dysfunction in aging and age-related diseases. Nat. Rev. Endocrinol. 2022, 18, 243–258. [Google Scholar] [CrossRef]
- Drake, A.M.; Coughlan, M.T.; Christophersen, C.T.; Snelson, M. Resistant starch as a dietary intervention to limit the progression of diabetic kidney disease. Nutrients 2022, 14, 4547. [Google Scholar] [CrossRef]
- Liberale, L.; Montecucco, F.; Tardif, J.-C.; Libby, P.; Camici, G.G. Inflamm-aging: The role of inflammation in age-dependent cardiovascular disease. Eur. Heart J. 2020, 41, 2974–2982. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. The IL-1 family of cytokines and receptors in rheumatic diseases. Nat. Rev. Rheumatol. 2019, 15, 612–632. [Google Scholar] [CrossRef] [PubMed]
- Elimam, H.; Abdulla, A.M.; Taha, I.M. Inflammatory markers and control of type 2 diabetes mellitus. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 800–804. [Google Scholar] [CrossRef] [PubMed]
- Angulo, J.; El Assar, M.; Álvarez-Bustos, A.; Rodríguez-Mañas, L. Physical activity and exercise: Strategies to manage frailty. Redox Biol. 2020, 35, 101513. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, J.S.; Sehrawat, A.; Mishra, J.; Sidhu, I.S.; Navik, U.; Khullar, N.; Kumar, S.; Bhatti, G.K.; Reddy, P.H. Oxidative stress in the pathophysiology of type 2 diabetes and related complications: Current therapeutics strategies and future perspectives. Free Radic. Biol. Med. 2022, 184, 114–134. [Google Scholar] [CrossRef]
- Halim, M.; Halim, A. The effects of inflammation, aging and oxidative stress on the pathogenesis of diabetes mellitus (type 2 diabetes). Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Sivandzade, F.; Prasad, S.; Bhalerao, A.; Cucullo, L. NRF2 and NF-қB interplay in cerebrovascular and neurodegenerative disorders: Molecular mechanisms and possible therapeutic approaches. Redox Biol. 2019, 21, 101059. [Google Scholar] [CrossRef]
- Narasimhan, A.; Flores, R.R.; Robbins, P.D.; Niedernhofer, L.J. Role of cellular senescence in type II diabetes. Endocrinology 2021, 162, bqab136. [Google Scholar] [CrossRef] [PubMed]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W. Chronic inflammation in the etiology of disease across the life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef]
- Zabetakis, I.; Lordan, R.; Tsoupras, A. The Impact of Nutrition and Statins on Cardiovascular Diseases; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Lainampetch, J.; Panprathip, P.; Phosat, C.; Chumpathat, N.; Prangthip, P.; Soonthornworasiri, N.; Puduang, S.; Wechjakwen, N.; Kwanbunjan, K. Association of tumor necrosis factor alpha, interleukin 6, and C-reactive protein with the risk of developing type 2 diabetes: A retrospective cohort study of rural Thais. J. Diabetes Res. 2019, 2019, 9051929. [Google Scholar] [CrossRef]
- Randeria, S.N.; Thomson, G.J.; Nell, T.A.; Roberts, T.; Pretorius, E. Inflammatory cytokines in type 2 diabetes mellitus as facilitators of hypercoagulation and abnormal clot formation. Cardiovasc. Diabetol. 2019, 18, 72. [Google Scholar] [CrossRef] [PubMed]
- Moshapa, F.T.; Riches-Suman, K.; Palmer, T.M. Therapeutic targeting of the proinflammatory IL-6-JAK/STAT signalling pathways responsible for vascular restenosis in type 2 diabetes mellitus. Cardiol. Res. Pract. 2019, 2019, 9846312. [Google Scholar] [CrossRef]
- Chi, C.; Li, D.-J.; Jiang, Y.-J.; Tong, J.; Fu, H.; Wu, Y.-H.; Shen, F.-M. Vascular smooth muscle cell senescence and age-related diseases: State of the art. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2019, 1865, 1810–1821. [Google Scholar] [CrossRef] [PubMed]
- De la Cruz-Ares, S.P.; Cardelo, M.M.; Gutiérrez-Mariscal, F.D.; Torres-Peña, J.; García-Rio, A.; Katsiki, N.M.; Malagón, M.; López-Miranda, J.; Pérez-Martínez, P.M.; Yubero-Serrano, E. Endothelial dysfunction and advanced glycation end products in patients with newly diagnosed versus established diabetes: From the CORDIOPREV study. Nutrients 2020, 12, 238. [Google Scholar] [PubMed]
- Biswas, I.; Khan, G.A. Endothelial dysfunction in cardiovascular diseases. In Basic and Clinical Understanding of Microcirculation; IntechOpen: Vienna, Austria, 2020; Volume 10. [Google Scholar] [CrossRef]
- Meza, C.A.; La Favor, J.D.; Kim, D.-H.; Hickner, R.C. Endothelial dysfunction: Is there a hyperglycemia-induced imbalance of NOX and NOS? Int. J. Mol. Sci. 2019, 20, 3775. [Google Scholar] [CrossRef]
- Marseglia, A.; Fratiglioni, L.; Kalpouzos, G.; Wang, R.; Bäckman, L.; Xu, W. Prediabetes and diabetes accelerate cognitive decline and predict microvascular lesions: A population-based cohort study. Alzheimer’s Dement. 2019, 15, 25–33. [Google Scholar] [CrossRef]
- Yan, C.; Zhou, Y.; Chen, Q.; Luo, Y.; Zhang, J.H.; Huang, H.; Shao, A. Dysfunction of the neurovascular unit in diabetes-related neurodegeneration. Biomed. Pharmacother. 2020, 131, 110656. [Google Scholar] [CrossRef] [PubMed]
- Moheet, A.; Mangia, S.; Seaquist, E.R. Impact of diabetes on cognitive function and brain structure. Ann. N. Y Acad. Sci. 2015, 1353, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Vecoli, C.; Borghini, A.; Andreassi, M.G. The molecular biomarkers of vascular aging and atherosclerosis: Telomere length and mitochondrial DNA4977 common deletion. Mutat. Res./Rev. Mutat. Res. 2020, 784, 108309. [Google Scholar] [CrossRef]
- Cardoso, A.L.; Fernandes, A.; Aguilar-Pimentel, J.A.; De Angelis, M.H.; Guedes, J.R.; Brito, M.A.; Ortolano, S.; Pani, G.; Athanasopoulou, S.; Gonos, E.S.; et al. Towards frailty biomarkers: Candidates from genes and pathways regulated in aging and age-related diseases. Aging Res. Rev. 2018, 47, 214–277. [Google Scholar] [CrossRef]
- Prašnikar, E.; Borišek, J.; Perdih, A. Senescent cells as promising targets to tackle age-related diseases. Aging Res. Rev. 2021, 66, 101251. [Google Scholar] [CrossRef]
- Yeh, S.H.-H.; Shie, F.-S.; Liu, H.-K.; Yao, H.-H.; Kao, P.-C.; Lee, Y.-H.; Chen, L.-M.; Hsu, S.-M.; Chao, L.-J.; Wu, K.-W. A high-sucrose diet aggravates Alzheimer’s disease pathology, attenuates hypothalamic leptin signaling, and impairs food-anticipatory activity in APPswe/PS1dE9 mice. Neurobiol. Aging 2020, 90, 60–74. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.Y.; Song, J. The importance of BDNF and RAGE in diabetes-induced dementia. Pharmacol. Res. 2020, 160, 105083. [Google Scholar] [CrossRef] [PubMed]
- Sankar, S.B.; Infante-Garcia, C.; Weinstock, L.D.; Ramos-Rodriguez, J.J.; Hierro-Bujalance, C.; Fernandez-Ponce, C.; Wood, L.B.; Garcia-Alloza, M. Amyloid beta and diabetic pathology cooperatively stimulate cytokine expression in an Alzheimer’s mouse model. J. Neuroinflamm. 2020, 17, 38. [Google Scholar] [CrossRef] [PubMed]
- Infante-Garcia, C.; Garcia-Alloza, M. Review of the effect of natural compounds and extracts on neurodegeneration in animal models of diabetes mellitus. Int. J. Mol. Sci. 2019, 20, 2533. [Google Scholar] [CrossRef] [PubMed]
- Maneechote, C.; Kerdphoo, S.; Jaiwongkam, T.; Chattipakorn, S.C.; Chattipakorn, N. Chronic Pharmacological Modulation of Mitochondrial Dynamics Alleviates Prediabetes-Induced Myocardial Ischemia–Reperfusion Injury by Preventing Mitochondrial Dysfunction and Programmed Apoptosis. Cardiovasc. Drugs Ther. 2023, 37, 89–105. [Google Scholar] [CrossRef] [PubMed]
- Balistreri, C.R.; Pisano, C.; Bertoldo, F.; Massoud, R.; Dolci, S.; Ruvolo, G. Red Blood Cell Distribution Width, Vascular Aging Biomarkers, and Endothelial Progenitor Cells for Predicting Vascular Aging and Diagnosing/Prognosing Age-Related Degenerative Arterial Diseases. Rejuvenation Res. 2019, 22, 399–408. [Google Scholar] [CrossRef]
- Pinna, A.; Carlino, P.; Serra, R.; Boscia, F.; Dore, S.; Carru, C.; Zinellu, A. Red Cell Distribution Width (RDW) and Complete Blood Cell Count-Derived Measures in Non-Arteritic Anterior Ischemic Optic Neuropathy. Int. J. Med. Sci. 2021, 18, 2239–2244. [Google Scholar] [CrossRef] [PubMed]
- Gratuze, M.; Joly-Amado, A.; Buee, L.; Vieau, D.; Blum, D. Overview on Tau and Tauopathies. Tau Biol. 2020, 1184, 259. [Google Scholar]
- Powrie, Y.S.L.; Smith, C. Central intracrine DHEA synthesis in aging-related neuroinflammation and neurodegeneration: Therapeutic potential? J. Neuroinflamm. 2018, 15, 289. [Google Scholar] [CrossRef]
- Thonusin, C.; Pantiya, P.; Sumneang, N.; Chunchai, T.; Nawara, W.; Arunsak, B.; Siri-Angkul, N.; Sriwichaiin, S.; Chattipakorn, S.C.; Chattipakorn, N. Effectiveness of high cardiorespiratory fitness in cardiometabolic protection in prediabetic rats. Mol. Med. 2022, 28, 31. [Google Scholar] [CrossRef] [PubMed]
- Rettberg, J.R.; Yao, J.; Brinton, R.D. Estrogen: A master regulator of bioenergetic systems in the brain and body. Front. Neuroendocr. 2014, 35, 8–30. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Yan, F.; Tian, J.; Qiao, A.; Yan, D. Potential clinical biomarkers and perspectives in diabetic cardiomyopathy. Diabetol. Metab. Syndr. 2023, 15, 35. [Google Scholar] [CrossRef] [PubMed]
- Carranza-Naval, M.J.; Vargas-Soria, M.; Hierro-Bujalance, C.; Baena-Nieto, G.; Garcia-Alloza, M.; Infante-Garcia, C.; Del Marco, A. Alzheimer’s disease and diabetes: Role of diet, microbiota and inflammation in preclinical models. Biomolecules 2021, 11, 262. [Google Scholar] [CrossRef]
- Meng, J.-M.; Cao, S.-Y.; Wei, X.-L.; Gan, R.-Y.; Wang, Y.-F.; Cai, S.-X.; Xu, X.-Y.; Zhang, P.-Z.; Li, H.-B. Effects and mechanisms of tea for the prevention and management of diabetes mellitus and diabetic complications: An updated review. Antioxidants 2019, 8, 170. [Google Scholar] [CrossRef] [PubMed]
- Carrageta, D.F.; Dias, T.R.; Alves, M.G.; Oliveira, P.F.; Monteiro, M.P.; Silva, B.M. Anti-obesity potential of natural methylxanthines. J. Funct. Foods 2018, 43, 84–94. [Google Scholar] [CrossRef]
- Biessels, G.J.; Despa, F. Cognitive decline and dementia in diabetes mellitus: Mechanisms and clinical implications. Nat. Rev. Endocrinol. 2018, 14, 591–604. [Google Scholar] [CrossRef]
- Lee, J.H.; Jahrling, J.B.; Denner, L.; Dineley, K.T. Targeting insulin for Alzheimer’s disease: Mechanisms, status and potential directions. J. Alzheimer’s Dis. 2018, 64, S427–S453. [Google Scholar] [CrossRef] [PubMed]
- Van Duinkerken, E.; Ryan, C.M. Diabetes mellitus in the young and the old: Effects on cognitive functioning across the life span. Neurobiol. Dis. 2020, 134, 104608. [Google Scholar] [CrossRef]
- Lee, S.Y.; Wang, J.; Chao, C.T.; Chien, K.L.; Huang, J.W. Frailty is associated with a higher risk of developing delirium and cognitive impairment among patients with diabetic kidney disease: A longitudinal population-based cohort study. Diabet. Med. 2021, 38, e14566. [Google Scholar] [CrossRef]
- Hayden, M.R. Type 2 diabetes mellitus increases the risk of late-onset Alzheimer’s disease: Ultrastructural remodeling of the neurovascular unit and diabetic gliopathy. Brain Sci. 2019, 9, 262. [Google Scholar] [CrossRef] [PubMed]
- Horton, W.B.; Barrett, E.J. Microvascular Dysfunction in Diabetes Mellitus and Cardiometabolic Disease. Endocr. Rev. 2021, 42, 29–55. [Google Scholar] [CrossRef] [PubMed]
- Silveira, A.C.; Dias, J.P.; Santos, V.M.; Oliveira, P.F.; Alves, M.G.; Rato, L.; Silva, B.M. The action of polyphenols in diabetes mellitus and Alzheimer’s disease: A common agent for overlapping pathologies. Curr. Neuropharmacol. 2019, 17, 590–613. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.H.; Hegde, V. Obesity and diabetes mediated chronic inflammation: A potential biomarker in Alzheimer’s disease. J. Pers. Med. 2020, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Krivanek, T.J.; Gale, S.A.; McFeeley, B.M.; Nicastri, C.M.; Daffner, K.R. Promoting successful cognitive aging: A ten-year update. J. Alzheimer’s Dis. 2021, 81, 871–920. [Google Scholar] [CrossRef] [PubMed]
- Khalid, M.; Petroianu, G.; Adem, A. Advanced glycation end products and diabetes mellitus: Mechanisms and perspectives. Biomolecules 2022, 12, 542. [Google Scholar] [CrossRef]
- Medina-Leyte, D.J.; Zepeda-García, O.; Domínguez-Pérez, M.; González-Garrido, A.; Villarreal-Molina, T.; Jacobo-Albavera, L. Endothelial dysfunction, inflammation and coronary artery disease: Potential biomarkers and promising therapeutical approaches. Int. J. Mol. Sci. 2021, 22, 3850. [Google Scholar] [CrossRef] [PubMed]
- Ashby, J.W.; Mack, J.J. Endothelial control of cerebral blood flow. Am. J. Pathol. 2021, 191, 1906–1916. [Google Scholar] [CrossRef] [PubMed]
- Theofilis, P.; Sagris, M.; Oikonomou, E.; Antonopoulos, A.S.; Siasos, G.; Tsioufis, C.; Tousoulis, D. Inflammatory mechanisms contributing to endothelial dysfunction. Biomedicines 2021, 9, 781. [Google Scholar] [CrossRef]
- Coco, C.; Sgarra, L.; Potenza, M.A.; Nacci, C.; Pasculli, B.; Barbano, R.; Parrella, P.; Montagnani, M. Can epigenetics of endothelial dysfunction represent the key to precision medicine in type 2 diabetes mellitus? Int. J. Mol. Sci. 2019, 20, 2949. [Google Scholar] [CrossRef]
- Johnson, J.; Jaggers, R.M.; Gopalkrishna, S.; Dahdah, A.; Murphy, A.J.; Hanssen, N.M.; Nagareddy, P.R. Oxidative stress in neutrophils: Implications for diabetic cardiovascular complications. Antioxid. Redox Signal. 2022, 36, 652–666. [Google Scholar] [CrossRef] [PubMed]
- Stanimirovic, J.; Radovanovic, J.; Banjac, K.; Obradovic, M.; Essack, M.; Zafirovic, S.; Gluvic, Z.; Gojobori, T.; Isenovic, E.R. Role of C-reactive protein in diabetic inflammation. Mediat. Inflamm. 2022, 2022, 3706508. [Google Scholar] [CrossRef] [PubMed]
- Forrester, J.V.; Kuffova, L.; Delibegovic, M. The role of inflammation in diabetic retinopathy. Front. Immunol. 2020, 11, 583687. [Google Scholar] [CrossRef]
- Landecho, M.F.; Tuero, C.; Valentí, V.; Bilbao, I.; De la Higuera, M.; Frühbeck, G. Relevance of leptin and other adipokines in obesity-associated cardiovascular risk. Nutrients 2019, 11, 2664. [Google Scholar] [CrossRef] [PubMed]
- Placha, D.; Jampilek, J. Chronic inflammatory diseases, anti-inflammatory agents and their delivery nanosystems. Pharmaceutics 2021, 13, 64. [Google Scholar] [CrossRef]
- Zatterale, F.; Longo, M.; Naderi, J.; Raciti, G.A.; Desiderio, A.; Miele, C.; Beguinot, F. Chronic adipose tissue inflammation linking obesity to insulin resistance and type 2 diabetes. Front. Physiol. 2020, 10, 1607. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, R.; Viana, S.D.; Nunes, S.; Reis, F. Diabetic gut microbiota dysbiosis as an inflammaging and immunosenescence condition that fosters progression of retinopathy and nephropathy. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2019, 1865, 1876–1897. [Google Scholar] [CrossRef]
- Kistner, T.M.; Pedersen, B.K.; Lieberman, D.E. Interleukin 6 as an energy allocator in muscle tissue. Nat. Metab. 2022, 4, 170–179. [Google Scholar] [CrossRef]
- Battineni, G.; Sagaro, G.G.; Chintalapudi, N.; Amenta, F.; Tomassoni, D.; Tayebati, S.K. Impact of obesity-induced inflammation on cardiovascular diseases (CVD). Int. J. Mol. Sci. 2021, 22, 4798. [Google Scholar] [CrossRef]
- Bashir, H.; Bhat, S.A.; Majid, S.; Hamid, R.; Koul, R.K.; Rehman, M.U.; Din, I.; Bhat, J.A.; Qadir, J.; Masood, A. Role of inflammatory mediators (TNF-α, IL-6, CRP), biochemical and hematological parameters in type 2 diabetes mellitus patients of Kashmir, India. Med. J. Islam. Repub. Iran 2020, 34, 5. [Google Scholar] [CrossRef]
- Rasmussen, L.J.H.; Petersen, J.E.V.; Eugen-Olsen, J. Soluble urokinase plasminogen activator receptor (suPAR) as a biomarker of systemic chronic inflammation. Front. Immunol. 2021, 12, 780641. [Google Scholar] [CrossRef] [PubMed]
- Podkowińska, A.; Formanowicz, D. Chronic kidney disease as oxidative stress-and inflammatory-mediated cardiovascular disease. Antioxidants 2020, 9, 752. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Mechanistic insight into oxidative stress-triggered signaling pathways and type 2 diabetes. Molecules 2022, 27, 950. [Google Scholar] [CrossRef] [PubMed]
- Aleksandrova, K.; Koelman, L.; Rodrigues, C.E. Dietary patterns and biomarkers of oxidative stress and inflammation: A systematic review of observational and intervention studies. Redox Biol. 2021, 42, 101869. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.Y.; Kim, D.H.; Lee, E.K.; Chung, K.W.; Chung, S.; Lee, B.; Seo, A.Y.; Chung, J.H.; Jung, Y.S.; Im, E. Redefining chronic inflammation in aging and age-related diseases: Proposal of the senoinflammation concept. Aging Dis. 2019, 10, 367. [Google Scholar] [CrossRef] [PubMed]
- Dascalu, A.e.; Furman, C.; Landrieu, I.; Cantrelle, F.X.; Mortelecque, J.; Grolaux, G.; Gillery, P.; Tessier, F.; Lipka, E.; Billamboz, M. Development of Receptor for Advanced Glycation End Products (RAGE) ligands through target directed dynamic combinatorial chemistry: A novel class of possible antagonists. Chem.–A Eur. J. 2024, 20, e202303255. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Gao, Y.; Zeng, C.; Hua, R.; Guo, Y.; Wang, Y.; Wang, Z. Effects of Maillard reaction and its product AGEs on aging and age-related diseases. Food Sci. Hum. Wellness 2024, 13, 1118–1134. [Google Scholar] [CrossRef]
- Soraci, L.; Corsonello, A.; Paparazzo, E.; Montesanto, A.; Piacenza, F.; Olivieri, F.; Gambuzza, M.E.; Savedra, E.V.; Marino, S.; Lattanzio, F. Neuroinflammaging: A Tight Line between Normal Aging and Age-Related Neurodegenerative Disorders. Aging Dis. 2024, 15, 2. [Google Scholar]
- Shen, C.-Y.; Lu, C.-H.; Wu, C.-H.; Li, K.-J.; Kuo, Y.-M.; Hsieh, S.-C.; Yu, C.-L. The development of maillard reaction, and advanced glycation end product (AGE)-receptor for AGE (RAGE) signaling inhibitors as novel therapeutic strategies for patients with AGE-related diseases. Molecules 2020, 25, 5591. [Google Scholar] [CrossRef]
- Boyajian, J.L.; Ghebretatios, M.; Schaly, S.; Islam, P.; Prakash, S. Microbiome and human aging: Probiotic and prebiotic potentials in longevity, skin health and cellular senescence. Nutrients 2021, 13, 4550. [Google Scholar] [CrossRef]
- Moura, J.; Madureira, P.; Leal, E.; Fonseca, A.; Carvalho, E. Immune aging in diabetes and its implications in wound healing. Clin. Immunol. 2019, 200, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Patankar, J.V.; Becker, C. Cell death in the gut epithelium and implications for chronic inflammation. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 543–556. [Google Scholar] [CrossRef] [PubMed]
- Burgess, J.L.; Wyant, W.A.; Abdo Abujamra, B.; Kirsner, R.S.; Jozic, I. Diabetic Wound-Healing Science. Medicina 2021, 57, 1072. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Wosinski, P.; Salazar-Noratto, G.E.; Bensidhoum, M.; Bizios, R.; Marashi, S.-A.; Potier, E.; Sheng, P.; Petite, H. Glucose Metabolism: Optimizing Regenerative Functionalities of Mesenchymal Stromal Cells Postimplantation. Tissue Eng. Part B Rev. 2023, 29, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Dubey, P.; Thakur, V.; Chattopadhyay, M. Role of minerals and trace elements in diabetes and insulin resistance. Nutrients 2020, 12, 1864. [Google Scholar] [CrossRef] [PubMed]
- Prasun, P. Mitochondrial dysfunction in metabolic syndrome. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2020, 1866, 165838. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wu, K.K.; Jiang, X.; Xu, A.; Cheng, K.K. The role of adipose tissue senescence in obesity-and aging-related metabolic disorders. Clin. Sci. 2020, 134, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Madan, S.; Uttekar, B.; Chowdhary, S.; Rikhy, R. Mitochondria lead the way: Mitochondrial dynamics and function in cellular movements in development and disease. Front. Cell Dev. Biol. 2022, 9, 781933. [Google Scholar] [CrossRef]
- Teodoro, J.S.; Nunes, S.; Rolo, A.P.; Reis, F.; Palmeira, C.M. Therapeutic options targeting oxidative stress, mitochondrial dysfunction and inflammation to hinder the progression of vascular complications of diabetes. Front. Physiol. 2019, 9, 1857. [Google Scholar] [CrossRef]
- Pinti, M.V.; Fink, G.K.; Hathaway, Q.A.; Durr, A.J.; Kunovac, A.; Hollander, J.M. Mitochondrial dysfunction in type 2 diabetes mellitus: An organ-based analysis. Am. J. Physiol.-Endocrinol. Metab. 2019, 316, E268–E285. [Google Scholar] [CrossRef]
- Ferreira, J.P.; Lamiral, Z.; Bakris, G.; Mehta, C.; White, W.B.; Zannad, F. Red cell distribution width in patients with diabetes and myocardial infarction: An analysis from the EXAMINE trial. Diabetes Obes. Metab. 2021, 23, 1580–1587. [Google Scholar] [CrossRef] [PubMed]
- Arkew, M.; Gemechu, K.; Haile, K.; Asmerom, H. Red blood cell distribution width as novel biomarker in cardiovascular diseases: A literature review. J. Blood Med. 2022, 13, 413–424. [Google Scholar] [CrossRef]
- Kavvasoglu, B.; Akdemir, S.; Kurt, M. A routine but overlooked parameter for impaired glucose control: Red cell distribution width. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 2431–2436. [Google Scholar] [PubMed]
- Zhou, W.; Sailani, M.R.; Contrepois, K.; Zhou, Y.; Ahadi, S.; Leopold, S.R.; Zhang, M.J.; Rao, V.; Avina, M.; Mishra, T. Longitudinal multi-omics of host–microbe dynamics in prediabetes. Nature 2019, 569, 663–671. [Google Scholar] [CrossRef]
- Gonzalez, A.; Deng, Y.; Lane, A.; Benkeser, D.; Cui, X.; Staimez, L.; Ford, C.; Khan, F.; Markley Webster, S.; Leong, A. Impact of mismatches in HbA1c vs glucose values on the diagnostic classification of diabetes and prediabetes. Diabet. Med. 2020, 37, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Beck, R.W.; Bergenstal, R.M.; Cheng, P.; Kollman, C.; Carlson, A.L.; Johnson, M.L.; Rodbard, D. The relationships between time in range, hyperglycemia metrics, and HbA1c. J. Diabetes Sci. Technol. 2019, 13, 614–626. [Google Scholar] [CrossRef] [PubMed]
- Kesika, P.; Sivamaruthi, B.S.; Chaiyasut, C. Do probiotics improve the health status of individuals with diabetes mellitus? A review on outcomes of clinical trials. BioMed Res. Int. 2019, 2019, 1531567. [Google Scholar] [CrossRef] [PubMed]
- Yanagita, I.; Fujihara, Y.; Iwaya, C.; Kitajima, Y.; Tajima, M.; Honda, M.; Teruya, Y.; Asakawa, H.; Ito, T.; Eda, T. Low serum albumin, aspartate aminotransferase, and body mass are risk factors for frailty in elderly people with diabetes–a cross-sectional study. BMC Geriatr. 2020, 20, 200. [Google Scholar] [CrossRef] [PubMed]
- Ismail, M.T.; Elbaih, A.H. Principles of intravenous fluids therapy. EC Emerg. Med. Crit. Care 2020, 4, 24–46. [Google Scholar]
- Petroni, M.L.; Brodosi, L.; Marchignoli, F.; Sasdelli, A.S.; Caraceni, P.; Marchesini, G.; Ravaioli, F. Nutrition in patients with type 2 diabetes: Present knowledge and remaining challenges. Nutrients 2021, 13, 2748. [Google Scholar] [CrossRef]
- Barbagallo, M.; Veronese, N.; Dominguez, L.J. Magnesium in aging, health and diseases. Nutrients 2021, 13, 463. [Google Scholar] [CrossRef]
- Wang, J.; Maxwell, C.A.; Yu, F. Biological processes and biomarkers related to frailty in older adults: A state-of-the-science literature review. Biol. Res. Nurs. 2019, 21, 80–106. [Google Scholar] [CrossRef]
- Boachie, J.; Adaikalakoteswari, A.; Samavat, J.; Saravanan, P. Low vitamin B12 and lipid metabolism: Evidence from pre-clinical and clinical studies. Nutrients 2020, 12, 1925. [Google Scholar] [CrossRef]
- Tiwari, V.; Wilson, D.M. DNA damage and associated DNA repair defects in disease and premature aging. Am. J. Hum. Genet. 2019, 105, 237–257. [Google Scholar] [CrossRef]
- Andes, L.J.; Cheng, Y.J.; Rolka, D.B.; Gregg, E.W.; Imperatore, G. Prevalence of prediabetes among adolescents and young adults in the United States, 2005-2016. JAMA Pediatr. 2020, 174, e194498. [Google Scholar] [CrossRef]
- Dhar, G.A.; Saha, S.; Mitra, P.; Nag Chaudhuri, R. DNA methylation and regulation of gene expression: Guardian of our health. Nucl. 2021, 64, 259–270. [Google Scholar] [CrossRef]
- Li, X.; Wang, J.; Wang, L.; Gao, Y.; Feng, G.; Li, G.; Zou, J.; Yu, M.; Li, Y.F.; Liu, C. Lipid metabolism dysfunction induced by age-dependent DNA methylation accelerates aging. Signal Transduct. Target. Ther. 2022, 7, 162. [Google Scholar] [CrossRef]
- Zhao, H.; Qi, C.; Zheng, C.; Gan, K.; Ren, L.; Song, G. Effects of glycated hemoglobin level on bone metabolism biomarkers in patients with type 2 diabetes mellitus. Diabetes Metab. Syndr. Obes. 2020, 13, 1785–1791. [Google Scholar] [CrossRef]
- Bellido, T.; Plotkin, L.I.; Bruzzaniti, A. Bone cells. In Basic and Applied Bone Biology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 37–55. [Google Scholar]
- Costantini, S.; Conte, C. Bone health in diabetes and prediabetes. World J. Diabetes 2019, 10, 421. [Google Scholar] [CrossRef]
- Kim, J.-M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.-H. Osteoblast-osteoclast communication and bone homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef]
- Anabtawi, A.; Le, T.; Putman, M.; Tangpricha, V.; Bianchi, M.L. Cystic fibrosis bone disease: Pathophysiology, assessment and prognostic implications. J. Cyst. Fibros. 2019, 18, S48–S55. [Google Scholar] [CrossRef]
- Rodriguez, A.J.; Scott, D.; Ebeling, P.R. Exploring the links between common diseases of aging—Osteoporosis, sarcopenia and vascular calcification. Clin. Rev. Bone Miner. Metab. 2019, 17, 1–23. [Google Scholar] [CrossRef]
- Khoramipour, K.; Chamari, K.; Hekmatikar, A.A.; Ziyaiyan, A.; Taherkhani, S.; Elguindy, N.M.; Bragazzi, N.L. Adiponectin: Structure, physiological functions, role in diseases, and effects of nutrition. Nutrients 2021, 13, 1180. [Google Scholar] [CrossRef]
- Nguyen, T.M.D. Adiponectin: Role in physiology and pathophysiology. Int. J. Prev. Med. 2020, 11, 136. [Google Scholar] [CrossRef]
- Shou, J.; Chen, P.-J.; Xiao, W.-H. Mechanism of increased risk of insulin resistance in aging skeletal muscle. Diabetol. Metab. Syndr. 2020, 12, 14. [Google Scholar] [CrossRef]
- Bleve, A.; Motta, F.; Durante, B.; Pandolfo, C.; Selmi, C.; Sica, A. Immunosenescence, inflammaging, and frailty: Role of myeloid cells in age-related diseases. Clin. Rev. Allergy Immunol. 2022, 64, 1–22. [Google Scholar] [CrossRef]
- Sardu, C.; D’Onofrio, N.; Torella, M.; Portoghese, M.; Loreni, F.; Mureddu, S.; Signoriello, G.; Scisciola, L.; Barbieri, M.; Rizzo, M.R. Pericoronary fat inflammation and Major Adverse Cardiac Events (MACE) in prediabetic patients with acute myocardial infarction: Effects of metformin. Cardiovasc. Diabetol. 2019, 18, 126. [Google Scholar] [CrossRef]
- Friedman, J.M. Leptin and the endocrine control of energy balance. Nat. Metab. 2019, 1, 754–764. [Google Scholar] [CrossRef]
- Pereira, S.; Cline, D.L.; Glavas, M.M.; Covey, S.D.; Kieffer, T.J. Tissue-specific effects of leptin on glucose and lipid metabolism. Endocr. Rev. 2021, 42, 1–28. [Google Scholar] [CrossRef]
- Gruzdeva, O.; Borodkina, D.; Uchasova, E.; Dyleva, Y.; Barbarash, O. Leptin resistance: Underlying mechanisms and diagnosis. Diabetes Metab. Syndr. Obes. Targets Ther. 2019, 12, 191–198. [Google Scholar] [CrossRef]
- Rubio-Ruiz, M.E.; Guarner-Lans, V.; Pérez-Torres, I.; Soto, M.E. Mechanisms underlying metabolic syndrome-related sarcopenia and possible therapeutic measures. Int. J. Mol. Sci. 2019, 20, 647. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chen, D.; Xiu, M.; Li, J.; Zhang, X.Y. Diabetes mellitus, cognitive deficits and serum BDNF levels in chronic patients with schizophrenia: A case-control study. J. Psychiatr. Res. 2021, 134, 39–47. [Google Scholar] [CrossRef] [PubMed]
- De Vincenti, A.P.; Ríos, A.S.; Paratcha, G.; Ledda, F. Mechanisms that modulate and diversify BDNF functions: Implications for hippocampal synaptic plasticity. Front. Cell. Neurosci. 2019, 13, 135. [Google Scholar] [CrossRef] [PubMed]
- Sumbul-Sekerci, B.; Sekerci, A.; Pasin, O.; Durmus, E.; Yuksel-Salduz, Z.I. Cognition and BDNF levels in prediabetes and diabetes: A mediation analysis of a cross-sectional study. Front. Endocrinol. 2023, 14, 1120127. [Google Scholar] [CrossRef] [PubMed]
- Azman, K.F.; Zakaria, R. Recent advances on the role of brain-derived neurotrophic factor (BDNF) in neurodegenerative diseases. Int. J. Mol. Sci. 2022, 23, 6827. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-derived neurotrophic factor: A key molecule for memory in the healthy and the pathological brain. Front. Cell. Neurosci. 2019, 13, 363. [Google Scholar] [CrossRef] [PubMed]
- Sebastian, M.J.; Khan, S.K.; Pappachan, J.M.; Jeeyavudeen, M.S. Diabetes and cognitive function: An evidence-based current perspective. World J. Diabetes 2023, 14, 92–109. [Google Scholar] [CrossRef]
- Amiri, N.; Fathei, M.; Mosaferi Ziaaldini, M. Effects of resistance training on muscle strength, insulin-like growth factor-1, and insulin-like growth factor–binding protein-3 in healthy elderly subjects: A systematic review and meta-analysis of randomized controlled trials. Hormones 2021, 20, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Bailes, J.; Soloviev, M. Insulin-like growth factor-1 (IGF-1) and its monitoring in medical diagnostic and in sports. Biomolecules 2021, 11, 217. [Google Scholar] [CrossRef]
- Vitale, G.; Pellegrino, G.; Vollery, M.; Hofland, L.J. Role of IGF-1 system in the modulation of longevity: Controversies and new insights from a centenarians’ perspective. Front. Endocrinol. 2019, 10, 27. [Google Scholar] [CrossRef]
- Mutavdzin, S.S.; Djuric, D.M. Homocysteine and Related B Vitamins in Pre-diabetes and Diabetes Mellitus. In Biochemistry of Cardiovascular Dysfunction in Obesity; Springer: Berlin/Heidelberg, Germany, 2020; pp. 329–351. [Google Scholar]
- Moretti, R.; Giuffre, M.; Caruso, P.; Gazzin, S.; Tiribelli, C. Homocysteine in neurology: A possible contributing factor to small vessel disease. Int. J. Mol. Sci. 2021, 22, 2051. [Google Scholar] [CrossRef]
- Tinelli, C.; Di Pino, A.; Ficulle, E.; Marcelli, S.; Feligioni, M. Hyperhomocysteinemia as a risk factor and potential nutraceutical target for certain pathologies. Front. Nutr. 2019, 6, 49. [Google Scholar] [CrossRef]
- Sondrup, N.; Termannsen, A.-D.; Eriksen, J.N.; Hjorth, M.F.; Færch, K.; Klingenberg, L.; Quist, J.S. Effects of sleep manipulation on markers of insulin sensitivity: A systematic review and meta-analysis of randomized controlled trials. Sleep Med. Rev. 2022, 62, 101594. [Google Scholar] [CrossRef]
- Alam, S.; Sarker, M.M.R.; Sultana, T.N.; Chowdhury, M.N.R.; Rashid, M.A.; Chaity, N.I.; Zhao, C.; Xiao, J.; Hafez, E.E.; Khan, S.A. Antidiabetic phytochemicals from medicinal plants: Prospective candidates for new drug discovery and development. Front. Endocrinol. 2022, 13, 800714. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, B.; Sultana, R.; Greene, M.W. Adipose tissue and insulin resistance in obese. Biomed. Pharmacother. 2021, 137, 111315. [Google Scholar] [CrossRef]
- Kitada, M.; Ogura, Y.; Monno, I.; Koya, D. Sirtuins and type 2 diabetes: Role in inflammation, oxidative stress, and mitochondrial function. Front. Endocrinol. 2019, 10, 187. [Google Scholar] [CrossRef] [PubMed]
- Thupakula, S.; Nimmala, S.S.R.; Ravula, H.; Chekuri, S.; Padiya, R. Emerging biomarkers for the detection of cardiovascular diseases. Egypt. Heart J. 2022, 74, 77. [Google Scholar] [CrossRef]
- Mariappan, V.; Srinivasan, R.; Pratheesh, R.; Jujjuvarapu, M.R.; Pillai, A.B. Predictive biomarkers for the early detection and management of heart failure. Heart Fail. Rev. 2023, 29, 331–353. [Google Scholar] [CrossRef] [PubMed]
- Castiglione, V.; Aimo, A.; Vergaro, G.; Saccaro, L.; Passino, C.; Emdin, M. Biomarkers for the diagnosis and management of heart failure. Heart Fail. Rev. 2022, 27, 625–643. [Google Scholar] [CrossRef]
- Ceriello, A.; Catrinoiu, D.; Chandramouli, C.; Cosentino, F.; Dombrowsky, A.C.; Itzhak, B.; Lalic, N.M.; Prattichizzo, F.; Schnell, O.; Seferović, P.M. Heart failure in type 2 diabetes: Current perspectives on screening, diagnosis and management. Cardiovasc. Diabetol. 2021, 20, 218. [Google Scholar] [CrossRef]
- Saraiva, R.M.M.d.S.d.S. Aging Biomarkers-A Global View. Doctoral Thesis, PhD- University of Coimbra, Coimbra, Portugal, 2021. Available online: https://hdl.handle.net/10316/98330 (accessed on 4 April 2024).
- Cao, Z.; Jia, Y.; Zhu, B. BNP and NT-proBNP as diagnostic biomarkers for cardiac dysfunction in both clinical and forensic medicine. Int. J. Mol. Sci. 2019, 20, 1820. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, H.; Mügge, A.; Trampisch, H.J.; Scharnagl, H.; März, W.; Kara, K. NT-proBNP for risk prediction of cardiovascular events and all-cause mortality: The getABI-study. IJC Heart Vasc. 2020, 29, 100553. [Google Scholar] [CrossRef]
- Gao, P.; Zou, X.; Sun, X.; Zhang, C. Cellular senescence in metabolic-associated kidney disease: An update. Cells 2022, 11, 3443. [Google Scholar] [CrossRef] [PubMed]
- Vallon, V.; Komers, R. Pathophysiology of the diabetic kidney. Compr. Physiol. 2011, 1, 1175. [Google Scholar] [PubMed]
- Docherty, M.H.; Baird, D.P.; Hughes, J.; Ferenbach, D.A. Cellular senescence and senotherapies in the kidney: Current evidence and future directions. Front. Pharmacol. 2020, 11, 543955. [Google Scholar] [CrossRef]
- Chen, Y.; Kanwar, Y.S.; Chen, X.; Zhan, M. Aging and diabetic kidney disease: Emerging pathogenetic mechanisms and clinical implications. Curr. Med. Chem. 2024, 31, 697–725. [Google Scholar] [CrossRef]
- Baranowska-Jurkun, A.; Matuszewski, W.; Bandurska-Stankiewicz, E. Chronic microvascular complications in prediabetic states—An overview. J. Clin. Med. 2020, 9, 3289. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Ha, X.; Yang, S.; Tian, X.; Jiang, H. Advances in understanding and treating diabetic kidney disease: Focus on tubulointerstitial inflammation mechanisms. Front. Endocrinol. 2023, 14, 1232790. [Google Scholar] [CrossRef] [PubMed]
- Naidoo, K.; Ngubane, P.S.; Khathi, A. Investigating the Effects of Diet-Induced Pre-Diabetes on the Functioning of Calcium-Regulating Organs in Male Sprague Dawley Rats: Effects on Selected Markers. Front. Endocrinol 2022, 11, 13:914189. [Google Scholar] [CrossRef]
- Ma, H.; Liu, S.; Li, S.; Xia, Y. Targeting growth factor and cytokine pathways to treat idiopathic pulmonary fibrosis. Front. Pharmacol. 2022, 13, 918771. [Google Scholar] [CrossRef]
- Loomba, R.; Friedman, S.L.; Shulman, G.I. Mechanisms and disease consequences of nonalcoholic fatty liver disease. Cell 2021, 184, 2537–2564. [Google Scholar] [CrossRef] [PubMed]
- Kenner, B.; Chari, S.T.; Kelsen, D.; Klimstra, D.S.; Pandol, S.J.; Rosenthal, M.; Rustgi, A.K.; Taylor, J.A.; Yala, A.; Abul-Husn, N. Artificial intelligence and early detection of pancreatic cancer: 2020 summative review. Pancreas 2021, 50, 251. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Gan, D.; Lin, S.; Zhong, Y.; Chen, M.; Zou, X.; Shao, Z.; Xiao, G. Metformin in aging and aging-related diseases: Clinical applications and relevant mechanisms. Theranostics 2022, 12, 2722. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Li, J.T.; Sun, P.; Wang, L.L.; Sun, L.Z.; Pang, S.G. Effects of lifestyle interventions on glucose regulation and diabetes risk in adults with impaired glucose tolerance or prediabetes: A meta-analysis. Arch. Endocrinol. Metab. 2022, 66, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Jung, G.; Hernández-Illán, E.; Moreira, L.; Balaguer, F.; Goel, A. Epigenetics of colorectal cancer: Biomarker and therapeutic potential. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 111–130. [Google Scholar] [CrossRef]
- Han, H. Clinical-community linkage strategies. In Applied Population Health Approaches for Asian American Communities, 1st ed.; John Wiley & Sons: Hoboken, NJ, Canada, 2022; p. 151. [Google Scholar]
- Justice, J.N.; Leng, I.; LeBrasseur, N.K.; Tchkonia, T.; Kirkland, J.L.; Mitin, N.; Liu, Y.; Kritchevsky, S.B.; Nicklas, B.J.; Ding, J. Caloric restriction intervention alters specific circulating biomarkers of the senescence-associated secretome in middle-aged and older adults with obesity and prediabetes in an 18-week randomized controlled trial. J. Gerontol. Ser. A 2024, 79, glad214. [Google Scholar] [CrossRef]
- Bönhof, G.J.; Herder, C.; Strom, A.; Papanas, N.; Roden, M.; Ziegler, D. Emerging biomarkers, tools, and treatments for diabetic polyneuropathy. Endocr. Rev. 2019, 40, 153–192. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Martínez, M.; González-González, M.; Martagón, A.J.; Hlavinka, V.; Willson, R.C.; Rito-Palomares, M. Recent developments in biomarkers for diagnosis and screening of type 2 diabetes mellitus. Curr. Diabetes Rep. 2022, 22, 95–115. [Google Scholar] [CrossRef] [PubMed]
- Green, S.; Hillersdal, L. Aging biomarkers and the measurement of health and risk. Hist. Philos. Life Sci. 2021, 43, 28. [Google Scholar] [CrossRef]
- Akbulut, S. An assessment of serum vitamin B12 and folate in patients with Crohn’s disease. Medicine 2022, 101, e31892. [Google Scholar] [CrossRef]
- Wu, Q.; Gao, Z.-J.; Yu, X.; Wang, P. Dietary regulation in health and disease. Signal Transduct. Target. Ther. 2022, 7, 252. [Google Scholar] [CrossRef] [PubMed]
- Bottani, M.; Banfi, G.; Lombardi, G. The clinical potential of circulating miRNAs as biomarkers: Present and future applications for diagnosis and prognosis of age-associated bone diseases. Biomolecules 2020, 10, 589. [Google Scholar] [CrossRef] [PubMed]
- Paton, B.; Suarez, M.; Herrero, P.; Canela, N. Glycosylation biomarkers associated with age-related diseases and current methods for glycan analysis. Int. J. Mol. Sci. 2021, 22, 5788. [Google Scholar] [CrossRef] [PubMed]
- Sławińska, N.; Krupa, R. Molecular Aspects of Senescence and Organismal Aging-DNA Damage Response, Telomeres, Inflammation and Chromatin. Int. J. Mol. Sci. 2021, 22, 590. [Google Scholar] [CrossRef]
- Jänig, W. The Integrative Action of the Autonomic Nervous System: Neurobiology of Homeostasis; Cambridge University Press: Cambridge, UK, 2022. [Google Scholar]
- Strasser, B.; Wolters, M.; Weyh, C.; Krüger, K.; Ticinesi, A. The effects of lifestyle and diet on gut microbiota composition, inflammation and muscle performance in our aging society. Nutrients 2021, 13, 2045. [Google Scholar] [CrossRef]
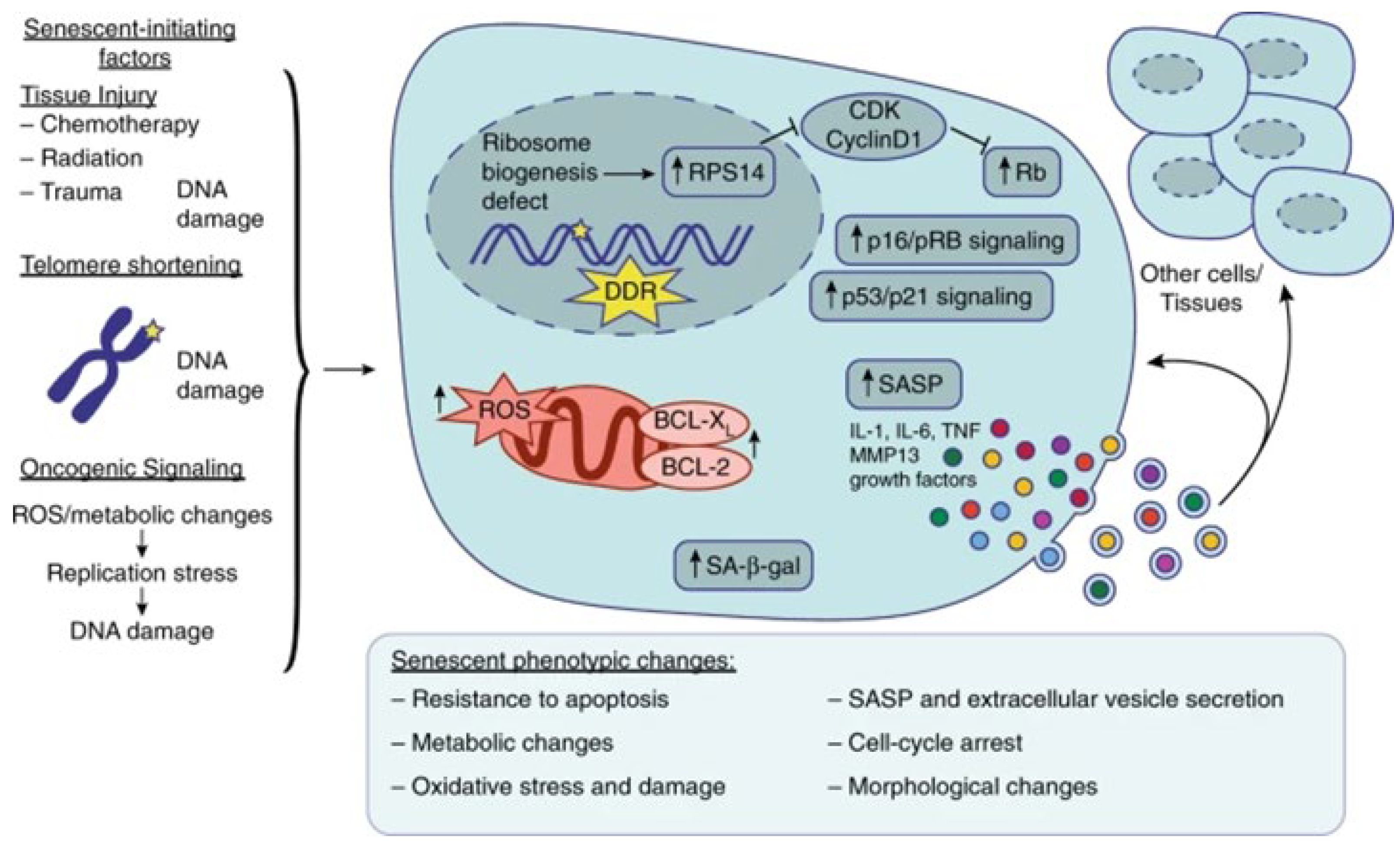
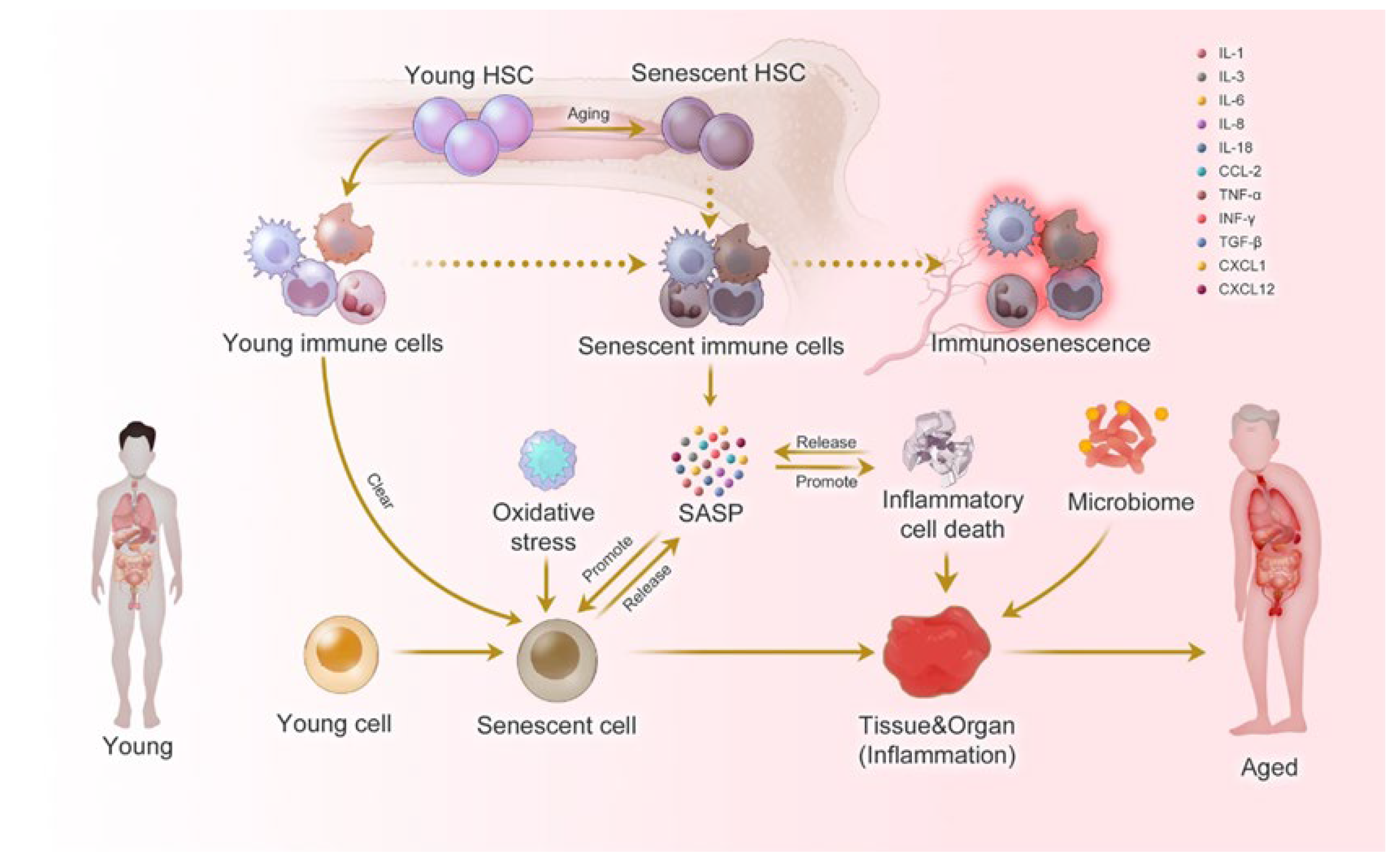
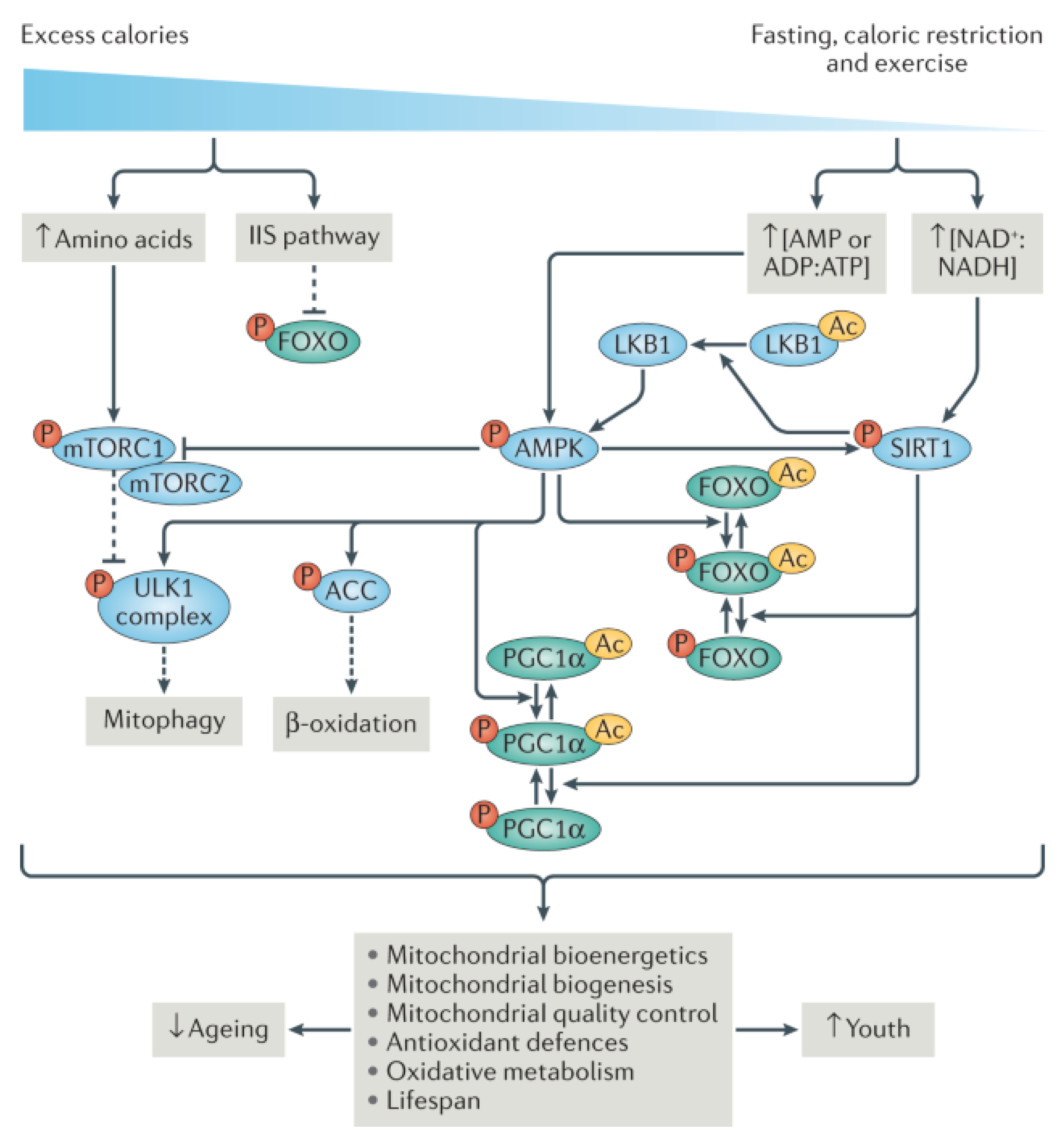

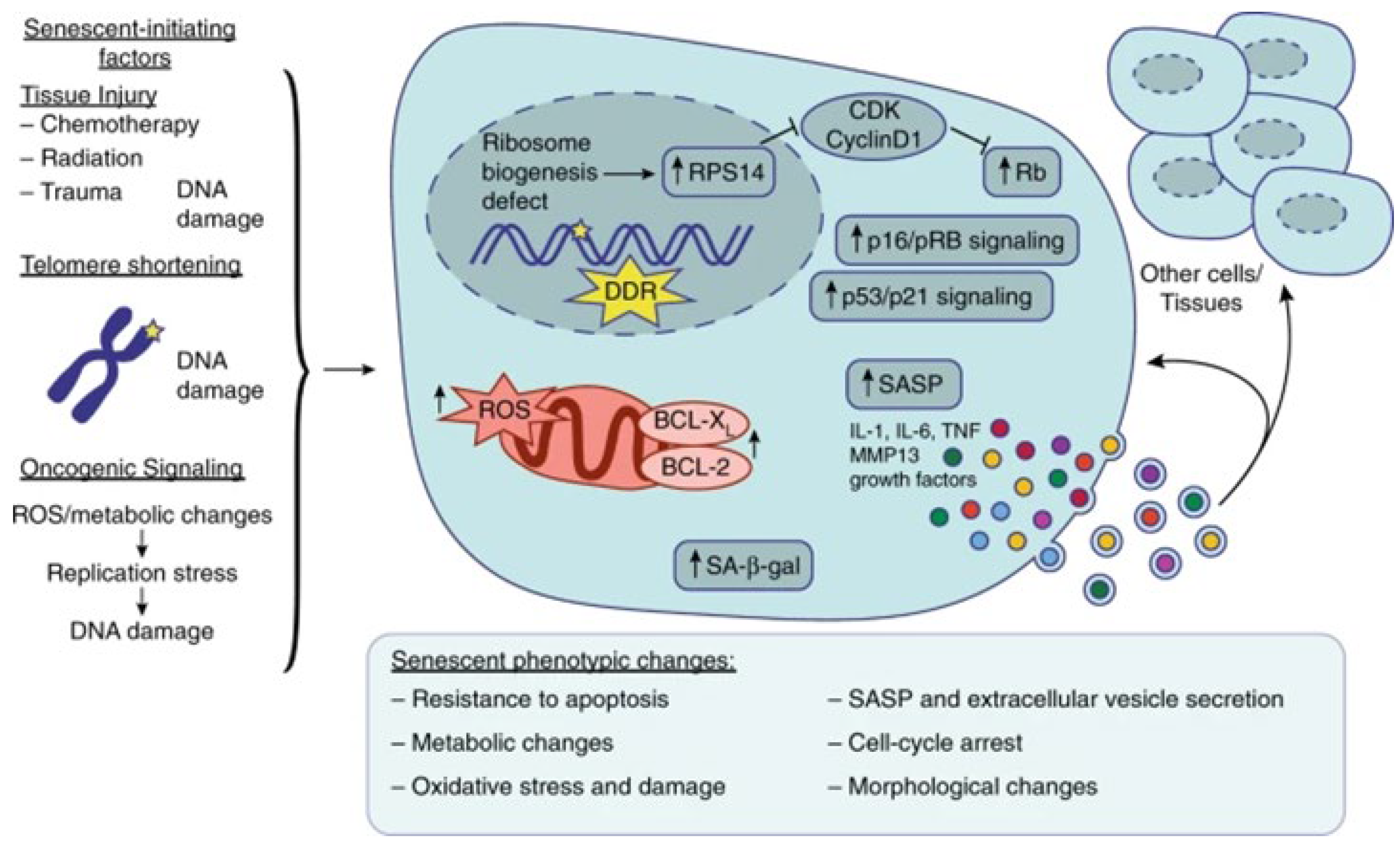{kind=link}
{kind=link}
{kind=link}
| Circulating Indicators of Aging | Dynamics during Aging | Function/Risk Factor | Reasons for the Condition | Lifespan Influence | Ref. |
|---|---|---|---|---|---|
| Growth hormone (GH) | Altered levels | Impact on muscle mass and bone density | Insulin resistance | Influence on aging | [90] |
| Insulin-like growth factor 1 | Variations during aging | Regulation of cell growth and repair | Metabolic changes | Potential lifespan influence | [91] |
| Dehydroepiandrosterone sulphate (DHEA-S) | Decreased levels | Hormonal changes | Prediabetes | Aging effect | [92] |
| Testosterone (in men) | Changes in aging | Impact on muscle and bone health | Hormonal alterations | Potential influence on the lifespan | [93] |
| Estrogen (in women) | Hormonal shifts during aging | Effects on bone density and cardiovascular health | Menopause and prediabetes | Aging impact | [94] |
| Circulating growth factors | Alterations with age | Role in cell growth, repair, and regeneration | Aging process | Lifespan variations | [95] |
| Brain-derived neurotrophic factor (BDNF) | Age-related changes | Cognitive health in aging | Prediabetes and aging | Potential influence on the lifespan | [96] |
| Insulin-like growth factor-binding proteins | Age-related alterations | Modulation of IGF-1 effects | Metabolic changes | Aging and lifespan | [97] |
| Additional factors in aging | Dynamics during aging | Various influences on aging | Prediabetes and aging | Lifespan variations | [98] |
| Telomere Length | Shortening with age | Cellular aging indicator | Oxidative stress and inflammation | Influence on aging | [99] |
| p16INK4a | Increased levels with age | Cellular senescence regulator | Prediabetes and aging | Accelerated aging | [100] |
| Senescence-Associated Secretory Phenotype (SASP) Factors | Elevated levels with age | Impact on inflammation and biochemistry | Chronic inflammation | Aging implications | [101] |
| DNA methylation clocks | Accelerated aging with age | Epigenetic changes indicator | Metabolic and Oxidative Stress | Influence on aging | [102] |
| Advanced Glycation End Products (AGEs) | Increased levels with age | Age-related complications indicator | Glycation and oxidative stress | Accelerated aging | [64] |
| Inflammatory Markers | Elevated with age | Indicators of chronic inflammation | Prediabetes and aging | Aging and inflammation | [103,104] |
| Oxidative stress markers | Increased with age | Oxidative damage indicators | Prediabetes and aging | Influence on aging | [105] |
| Red blood cell distribution width (RDW) | Increased with age | Inflammation and metabolic changes | Prediabetes and aging | Influence on aging | [106] |
| Hemoglobin A1c (HbA1c) | Elevated with age | Impact of hyperglycaemia on tissues and systems | Chronic hyperglycaemia | Aging and diabetes | [107] |
| Serum Albumin | Decreased with age | Nutritional status indicator | Prediabetes and aging | Influence on aging | [108] |
| Circulating Biomarker | Dynamics during Aging | Function and Risk Factors in Aging | Molecule Longevity Influence | Ref. |
|---|---|---|---|---|
| Inflammatory Mediators | Changes during aging | Role in chronic inflammation and aging | May influence lifespan | [122] |
| Pro-inflammatory Cytokines (e.g., IL-6) | Increased levels | Chronic inflammation and aging | May shorten the lifespan | [30,123] |
| Chemokines (e.g., MCP-1) | Altered dynamics | Recruitment of immune cells and aging | May impact lifespan | [124] |
| Growth factors (e.g., TGF-β1) | Variation with age | Modulation of cell growth and aging | Influence on the lifespan | [125] |
| Senescence-Associated Secretory Phenotype (SASP) Factors | Increased with age | Promotion of inflammation and aging | May influence lifespan | [126] |
| Inflammatory markers (e.g., CRP) | Elevated with age | Indicators of chronic inflammation | May impact lifespan | [56] |
| Oxidative stress markers (e.g., ROS) | Increased with age | Indicators of oxidative damage | May influence lifespan | [127] |
| Endothelial markers (e.g., vWF) | Altered dynamics | Indicators of endothelial dysfunction | May impact lifespan | [128] |
| DNA damage markers (e.g., 8-OHdG) | Increased levels | Indicators of DNA damage and aging | May influence lifespan | [129] |
| Mitochondrial dysfunction markers (e.g., mtDNA) | Changes during aging | Indicators of impaired mitochondrial function | May impact lifespan | [130] |
| Immune system biomarkers (e.g., CD4+ T cells) | Altered dynamics | Immune system indicators of aging | May influence lifespan | [56,123] |
| Biomarker | Role in Aging | Implications for Senescence | Ref. |
|---|---|---|---|
| Telomere length | Reflects cellular aging and senescence | Accelerated aging and cellular senescence | [152] |
| p16INK4a | Regulates cellular senescence | Increased cellular senescence | [153] |
| Senescence-associated secretory phenotype (SASP) Factors | Reflect senescent cell secretions | Promote inflammation and senescence | [154] |
| DNA methylation clocks | Epigenetic aging indicators | Accelerated epigenetic aging | [155] |
| Advanced glycation end products (AGEs) | Reflect glycation and oxidative stress | Contributions to accelerated aging and age-related complications | [156] |
| Inflammatory markers | Indicators of inflammation | Contributions to inflammation associated with aging | [157] |
| Oxidative stress markers | Indicators of oxidative damage and stress | Exacerbation of age-related oxidative damage | [156] |
| Endothelial dysfunction | Indicators of vascular dysfunction | Exacerbation of endothelial dysfunction and impact on vascular health | [158] |
| Mitochondrial dysfunction | Reflect mitochondrial function and health | Impairment of mitochondrial function associated with aging | [159] |
| Red blood cell distribution width (RDW) | Reflect changes in erythrocytes | Indicate inflammation and metabolic changes affecting aging | [160] |
| Haemoglobin A1c (HbA1c) | Reflects long-term blood glucose levels | Accelerating aging due to chronic hyperglycaemia | [161] |
| Serum albumin | Reflects nutritional status and frailty | Affecting nutritional status and frailty | [159] |
| Circulating Growth Hormone (GH) and Insulin-like Growth Factor 1 (IGF-1) | Reflect hormonal changes | Influence of insulin resistance and metabolic changes | [162] |
| DHEA-S | Reflects hormonal changes | Contributions to hormonal changes associated with aging | [163] |
| Testosterone (in men) | Reflects hormonal changes in men | Impact of muscle mass, bone density, and aging | [63,164] |
| Estrogen (in women) | Reflects hormonal changes in women | Affects bone density, cardiovascular health, and aging | [165] |
| Brain-derived neurotrophic factor (BDNF) | Reflects changes in neurotrophic factors | Influence on cognitive health, particularly in aging | [156] |
| IGF-binding proteins | Reflect changes in IGF-1 bioavailability | Contributions to metabolic and aging-related effects | [166] |
| Folate and B12 levels | Reflect nutritional deficiencies | Impact of DNA methylation and repair essential for aging | [167] |
| Osteocalcin | Reflect changes in bone health | Affecting bone health, a key consideration in aging | [168] |
| Adiponectin | Reflects changes in metabolic health | Impact of insulin resistance and metabolic changes | [167] |
| Leptin | Reflects changes in adipose tissue | Impact of metabolism and aging | [155] |
| Homocysteine | Reflects cardiovascular risk | Maybe more prevalent in prediabetic individuals | [156,169] |
| Insulin resistance markers | Reflect insulin resistance | May worsen with age in prediabetic individuals | [170,171] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mbatha, N.A.; Mushebenge, A.G.-A.; Khathi, A. Identification of Putative Causal Relationships between Blood-Based Biomarkers and Prediabetes-Induced Senescence: A Comprehensive Review. Physiologia 2024, 4, 149-181. https://doi.org/10.3390/physiologia4020009
Mbatha NA, Mushebenge AG-A, Khathi A. Identification of Putative Causal Relationships between Blood-Based Biomarkers and Prediabetes-Induced Senescence: A Comprehensive Review. Physiologia. 2024; 4(2):149-181. https://doi.org/10.3390/physiologia4020009
Chicago/Turabian StyleMbatha, Nonkululeko Avril, Aganze Gloire-Aimé Mushebenge, and Andile Khathi. 2024. "Identification of Putative Causal Relationships between Blood-Based Biomarkers and Prediabetes-Induced Senescence: A Comprehensive Review" Physiologia 4, no. 2: 149-181. https://doi.org/10.3390/physiologia4020009
APA StyleMbatha, N. A., Mushebenge, A. G.-A., & Khathi, A. (2024). Identification of Putative Causal Relationships between Blood-Based Biomarkers and Prediabetes-Induced Senescence: A Comprehensive Review. Physiologia, 4(2), 149-181. https://doi.org/10.3390/physiologia4020009