

Gene Expression of Monoterpene Synthases Is Affected Rhythmically during the Day in Lavandula angustifolia Flowers

 , ,
, , 
Abstract

1. Introduction
2. Results and Discussion
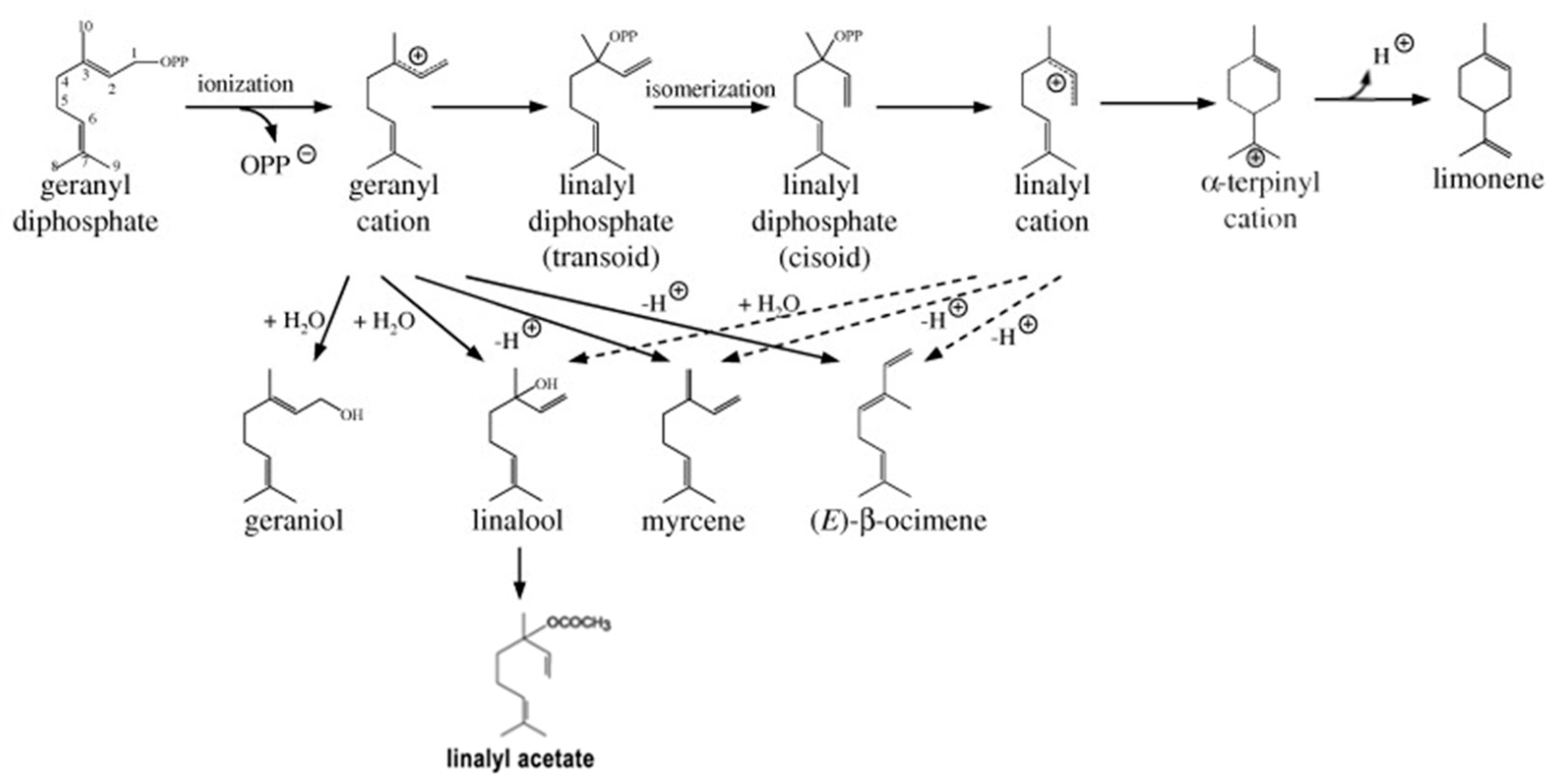
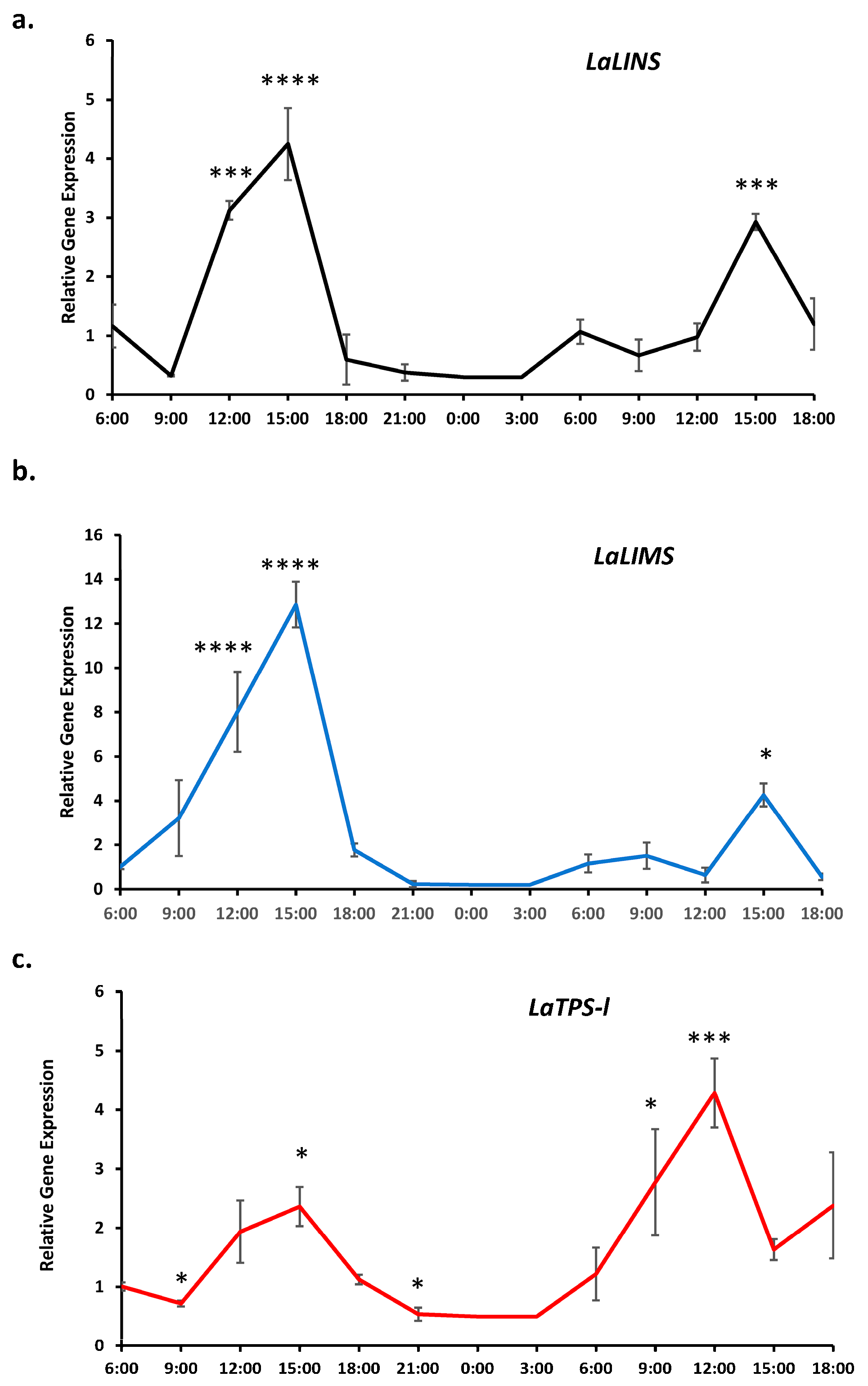
2.1. Gene Expression of Monoterpenes Synthases Displays a Circadian Rhythm Pattern
2.2. The Lavender EO Composition Is Affected by Harvest Time
3. Materials and Methods
3.1. Plant Material and EO Extraction
3.2. EO Analysis
3.3. RNA Isolation and Gene Expression Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jullien, F.; Fontez, M.; Bony, A.; Nicole, F.; Moja, S. Lavandula angustifolia Mill. a model of aromatic and medicinal plant to study volatile organic compounds synthesis, evolution and ecological functions. Bot. Let. 2023, 170, 65–76. [Google Scholar] [CrossRef]
- Paul, J.P.; Brophy, J.J.; Goldsack, R.J.; Fontaniella, B. Analysis of volatile components of Lavandula canariensis (L.) Mill., a Canary Islands endemic species, growing in Australia. Biochem. Syst. Ecol. 2004, 32, 55–62. [Google Scholar] [CrossRef]
- Lis-Balchin, M.; Hart, S. Studies on the mode of action of the essential oil of lavender (Lavandula angustifolia P. Miller). Phytother. Res. 1999, 13, 540–542. [Google Scholar] [CrossRef]
- Soltani, R.; Soheilipour, S.; Hajhashemi, V.; Asghari, G.; Bagheri, M.; Molavi, M. Evaluation of the effect of aromatherapy with lavender essential oil on post-tonsillectomy pain in pediatric patients: A randomized controlled trial. Inter. J. Pediatr. Otorhinolaryngol. 2013, 77, 1579–1581. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.S.; Lee, D.S. Comparison of different extraction methods for the analysis of fragrances from Lavandula species by gas chromatography–mass spectrometry. J. Chromatogr. A 2002, 982, 31–47. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, H.M.; Wilkinson, J.M. Biological activities of lavender essential oil. Phytother. Res. 2002, 16, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Kashani, M.S.; Tavirani, M.R.; Talaei, S.A.; Salami, M. Aqueous extract of lavender (Lavandula angustifolia) improves the spatial performance of a rat model of Alzheimer’s disease. Neurosci. Bull. 2011, 27, 99–106. [Google Scholar] [CrossRef]
- Sienkiewicz, M.; Lysakowska, M.; Ciecwierz, J.; Denys, P.; Kowalczyk, E. Antibacterial activity of thyme and lavender essential oils. Med. Chem. 2011, 7, 674–689. [Google Scholar] [CrossRef] [PubMed]
- Wells, R.; Truong, F.; Adal, A.M.; Sarker, L.S.; Mahmoud, S.S. Lavandula essential oils: A current review of applications in medicinal, food, and cosmetic industries of lavender. Nat. Prod. Commun. 2018, 13, 1934578x1801301. [Google Scholar] [CrossRef]
- Jullien, F.; Moja, S.; Bony, A.; Legrand, S.; Petit, C.; Benabdelkader, T.; Poirot, K.; Fiorucci, S.; Guitton, Y.; Nicolè, F.; et al. Isolation and functional characterization of a-cadinol synthase, a new sesquiterpene synthase from Lavandula angustifolia. Plant Mol. Biol. 2014, 84, 227–241. [Google Scholar] [CrossRef]
- Franks, S.J.; Wheeler, G.S.; Goodnight, C. Genetic variation and evolution of secondary compounds in native and introduced populations of the invasive plant Melaleuca qunquenervia. Evolution 2012, 66, 1398–1412. [Google Scholar] [CrossRef] [PubMed]
- Gershenzone, J.; Croteau, R. Herbivores: Their interactions with secondary plant metabolites. In Terpenoids: The Chemical Participants; Rosenthal, G.A., Berenbaum, M., Eds.; Academic Press: San Diego, CA, USA, 1991; pp. 165–219. [Google Scholar]
- Southwell, I.A.; Russell, M.F.; Maddox, C.D.A.; Wheeler, G.S. Differential metabolism of 1,8-cineole in insects. J. Chem. Ecol. 2003, 29, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Degenhardt, J.; Köllner, T.G.; Gershenzon, J. Monoterpene and sesquiterpene synthases and the origin of terpene skeletal diversity in plants. Phytochemistry 2009, 70, 1621–1637. [Google Scholar] [CrossRef] [PubMed]
- Zaks, A.; Davidovich-Rikanati, R.; Bar, E.; Inbar, M.; Lewinsohn, E. Biosynthesis of linalyl acetate and other terpenes in lemon mint (Mentha aquatica var. citrata, Lamiaceae) glandular trichomes. Israel J. Plant Sci. 2008, 56, 233–244. [Google Scholar] [CrossRef]
- Fopa Fomeju, B.; Brunel, D.; Bérard, A.; Rivoal, J.-B.; Gallois, P.; Le Paslier, M.-C.; Bouverat-Bernier, J.-P. Quick and effi-cient approach to develop genomic resources in orphan species: Application in Lavandula angustifolia. PLoS ONE 2020, 15, e0243853. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.; Kang, K.; Wang, P.; Li, M.; Huang, X. Transcriptome profiling of spike provides expression features of genes related to terpene biosynthesis in lavender. Sci. Rep. 2020, 10, 6933. [Google Scholar] [CrossRef]
- Lane, A.; Boecklemann, A.; Woronuk, G.N.; Sarker, L.; Mahmoud, S.S. A genomics resource for investigating regulation of essential oil production in Lavandula angustifolia. Planta 2010, 231, 835–845. [Google Scholar] [CrossRef]
- Li, H.; Li, J.; Dong, Y.; Hao, H.; Ling, Z.; Bai, H.; Wang, H.; Cui, H.; Shi, L. Time-series transcriptome provides insights into the gene regulation network involved in the volatile terpenoid metabolism during the flower development of lavender. BMC Plant Biol. 2019, 19, 313. [Google Scholar] [CrossRef]
- Malli, R.P.N.; Adal, A.M.; Sarker, L.S.; Liang, P.; Mahmoud, S.S. De novo sequencing of the Lavandula angustifolia genome reveals highly duplicated and optimized features for essential oil production. Planta 2019, 249, 251–256. [Google Scholar] [CrossRef]
- Li, J.; Wang, Y.; Dong, Y.; Zhang, W.; Wang, D.; Bai, H.; Li, K.; Li, H.; Shi, L. The chromosome-based lavender genome provides new insights into Lamiaceae evolution and terpenoid biosynthesis. Hortic. Res. 2021, 8, 53. [Google Scholar] [CrossRef]
- Hassiotis, C.N.; Ntana, F.; Lazari, D.M.; Poulios, S.; Vlachonasios, K.E. Environmental and developmental factors affect essential oil production and quality of Lavandula angustifolia during flowering period. Ind. Crops Prod. 2014, 62, 359–366. [Google Scholar] [CrossRef]
- Hassiotis, C.N.; Lazari, D.M.; Vlachonasios, K.E. The effects of habitat type and diurnal harvest on essential oil yield and composition of Lavandula angustifolia mill. Fresen. Environ. Bull. 2010, 19, 1491–1498. [Google Scholar]
- Picazo-Aragonés, J.; Terrab, A.; Balao, F. Plant volatile organic compounds evolution: Transcriptional regulation, epigenetics and polyploidy. Inter. J. Mol. Sci. 2020, 21, 8956. [Google Scholar] [CrossRef] [PubMed]
- Colquhoun, T.A.; Verdonk, J.C.; Schimmel, B.C.; Tieman, D.M.; Underwood, B.A.; Clark, D.G. Petunia floral volatile benzenoid/phenylpropanoid genes are regulated in a similar manner. Phytochemistry 2010, 71, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Guitton, Y.; Nicolè, F.; Moja, S.; Valot, N.; Legrand, S.; Jullien, F.; Legendre, L. Differential accumulation of volatile terpene and terpene synthase mRNAs during lavender (Lavandula angustifolia and L. × intermedia) inflorescence development. Physiol. Plant. 2010, 138, 150–163. [Google Scholar] [CrossRef] [PubMed]
- Demissie, Z.A.; Sarker, L.S.; Mahmoud, S.S. Cloning and functional characterization of β-phellandrene synthase from Lavandula angustifolia. Planta 2011, 233, 685–696. [Google Scholar] [CrossRef]
- Mostafa, S.; Wang, Y.; Zeng, W.; Jin, B. Floral Scents and Fruit Aromas: Functions, Compositions, Biosynthesis, and Regulation. Front. Plant Sci. 2022, 13, 860157. [Google Scholar] [CrossRef]
- Fenske, M.P.; Imaizumi, T. Circadian rhythms in floral scent emission. Front. Plant Sci. 2016, 7, 462. [Google Scholar] [CrossRef]
- Zeng, L.; Wang, X.; Kang, M.; Dong, F.; Yang, Z. Regulation of the rhythmic emission of plant volatiles by the circadian clock. Inter. J. Mol. Sci. 2017, 18, 2408. [Google Scholar] [CrossRef]
- Chuang, Y.C.; Lee, M.C.; Chang, Y.L.; Chen, W.H.; Chen, H.H. Diurnal regulation of the floral scent emission by light and circadian rhythm in the Phalaenopsis orchids. Bot. Stud. 2017, 58, 50. [Google Scholar] [CrossRef]
- Abbas, F.; Ke, Y.; Yu, R.; Fan, Y. Functional characterization and expression analysis of two terpene synthases involved in foral scent formation in Lilium ‘Siberia’. Planta 2019, 249, 71–93. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef]
- Yeh, C.-W.; Zhong, H.-Q.; Ho, Y.-F.; Tian, Z.-H.; Yeh, K.-W. The diurnal emission of foral scent in Oncidium hybrid orchid is controlled by CIRCADIAN CLOCK ASSOCIATED 1 (CCA1) through the direct regulation on terpene synthase. BMC Plant Biol. 2022, 22, 472. [Google Scholar] [CrossRef] [PubMed]
- Fenske, M.P.; Hazelton, K.D.H.; Hempton, A.K.; Shim, J.S.; Yamamoto, B.M.; Riffell, J.A.; Imaizumi, T. Circadian clock gene LATE ELONGATED HYPOCOTYL directly regulates the timing of floral scent emission in Petunia. Proc. Natl. Acad. Sci. USA 2015, 112, 9775–9780. [Google Scholar] [CrossRef] [PubMed]
- Sarker, L.S.; Adal, A.M.; Mahmoud, S.S. Diverse transcription factors control monoterpene synthase expression in lavender (Lavandula). Planta 2020, 251, 5. [Google Scholar] [CrossRef] [PubMed]
- Hassiotis, C.N.; Tarantilis, P.A.; Daferera, D.; Polissiou, M.G. Etherio, a new variety of Lavandula angustifolia with improved essential oil production and composition from natural selected genotypes growing in Greece. Ind. Crops Prod. 2010, 32, 77–82. [Google Scholar] [CrossRef]
- Van den Dool, H.; Kratz, P.D. A generalization of the retention index system including linear temperature programmed gas-liquid partition chromatography. J. Chromatogr. 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Massada, Y. Analysis of Essential Oil by Gas Chromatography and Spectrometry; John Wiley & Sons: New York, NY, USA, 1976. [Google Scholar]
- Adams, R.P. Identification of Essential Oils Components by Gas Chromatography/Quadrupole Mass Spectroscopy; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Bisio, A.; Ciarallo, G.; Romussi, G.; Fontana, N.; Mascolo, N.; Capasso, R.; Biscardi, D. Chemical composition of essential oils from some Salvia species. Phytother. Res. 1998, 12, 117–120. [Google Scholar] [CrossRef]
- Davies, N.N. Gas chromatographic retention indices of monoterpenes and sesquiterpenes on methyl silicone and Carbowax 20 M phases. J. Chromatogr. 1990, 503, 1–24. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds 1 | RI. | Compound Concentration (Percentage in Total EO.) | Identification 2 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 9:00 | 12:00 | 15:00 | 18:00 | |||||||
| 3-Octanone | 988 | 0.48 | ±0.01 | 0.47 | ±0.04 | 0.41 | ±0.04 | 0.10 | ±0.00 | I, MS |
| β-Myrcene | 991 | 0.94 | ±0.00 | 1.13 | ±0.12 | 1.21 | ±0.02 | 1.13 | ±0.05 | I, MS |
| Hexyl acetate | 1016 | 0.79 | ±0.08 | 0.64 | ±0.07 | 0.58 | ±0.01 | 0.25 | ±0.01 | I, MS |
| Limonene | 1028 | 0.53 | ±0.01 | 0.74 | ±0.01 | 0.53 | ±0.02 | 0.30 | ±0.01 | I, MS |
| 1,8-Cineol | 1029 | 1.94 | ±0.03 | 1.75 | ±0.00 | 1.18 | ±0.13 | 0.46 | ±0.02 | I, MS |
| cis-Ocimene | 1039 | 1.18 | ±0.04 | 0.99 | ±0.10 | 0.96 | ±0.02 | 0.44 | ±0.04 | I, MS |
| trans-Ocimene | 1049 | 1.84 | ±0.16 | 1.50 | ±0.03 | 1.35 | ±0.05 | 0.73 | ±0.08 | I, MS |
| Terpinolene | 1087 | 0.15 | ±0.00 | 0.15 | ±0.00 | 0.13 | ±0.00 | 0.07 | ±0.01 | I, MS |
| Linalool | 1100 | 33.02 | ±1.38 | 32.48 | ±1.36 | 32.51 | ±0.67 | 29.05 | ±0.58 | I, MS |
| 1-Octen-3-ol acetate | 1114 | 0.30 | ±0.03 | 0.23 | ±0.02 | 0.22 | ±0.01 | 0.13 | ±0.01 | I, MS |
| 3-Octanol acetate | 1126 | 0.17 | ±0.01 | 0.11 | ±0.01 | 0.12 | ±0.00 | 0.07 | ±0.01 | I, MS |
| Camphor | 1143 | 3.79 | ±0.35 | 3.30 | ±0.07 | 3.31 | ±0.06 | 2.65 | ±0.05 | I, MS |
| Hexyl isobutanoate | 1151 | 0.22 | ±0.02 | 0.16 | ±0.00 | 0.16 | ±0.02 | 0.14 | ±0.00 | I, MS |
| Borneol | 1165 | 2.75 | ±0.30 | 2.80 | ±0.31 | 2.87 | ±0.35 | 2.73 | ±0.28 | I, MS |
| Menthol | 1172 | 0.19 | ±0.01 | 0.25 | ±0.01 | 0.32 | ±0.03 | 0.28 | ±0.01 | I, MS |
| Terpinen-4-ol | 1176 | tr | tr | tr | tr | I, MS | ||||
| Cryptone | 1187 | 0.14 | ±0.01 | 0.13 | ±0.01 | 0.08 | ±0.00 | 0.07 | ±0.00 | I, MS |
| α-Terpineol | 1190 | 0.77 | ±0.08 | 1.02 | ±0.04 | 0.64 | ±0.01 | 0.70 | ±0.03 | I, MS |
| Hexyl butanoate | 1193 | 0.92 | ±0.04 | 0.94 | ±0.04 | 1.04 | ±0.04 | 1.05 | ±0.04 | I, MS |
| Dodecane | 1200 | 0.09 | ±0.01 | 0.12 | ±0.01 | 0.14 | ±0.01 | 0.11 | ±0.00 | I, MS |
| Octyl acetate | 1213 | tr | tr | tr | tr | I, MS | ||||
| Hexyl 2-methyl butanoate | 1238 | 0.15 | ±0.01 | 0.12 | ±0.01 | 0.15 | ±0.01 | 0.14 | ±0.01 | I, MS |
| Hexyl isovalerate | 1243 | 0.11 | ±0.01 | 0.14 | ±0.00 | 0.14 | ±0.01 | 0.15 | ±0.02 | I, MS |
| Linalyl acetate | 1257 | 40.92 | ±3.93 | 41.50 | ±0.00 | 43.00 | ±0.83 | 46.80 | ±0.95 | I, MS |
| Bornyl acetate | 1288 | 0.08 | ±0.01 | 0.09 | ±0.01 | 0.10 | ±0.01 | 0.10 | ±0.00 | I, MS |
| Lavandulyl acetate | 1292 | 1.59 | ±0.07 | 1.80 | ±0.19 | 2.04 | ±0.19 | 2.37 | ±0.00 | I, MS |
| Tridecane | 1300 | - | tr | 0.11 | ±0.01 | tr | I, MS | |||
| Hexyl tiglate | 1332 | 0.33 | ±0.03 | 0.32 | ±0.01 | 0.34 | ±0.03 | 0.39 | ±0.01 | I, MS |
| Neryl acetate | 1366 | 0.43 | ±0.08 | 0.38 | ±0.03 | 0.39 | ±0.02 | 0.48 | ±0.01 | I, MS |
| Geranyl acetate | 1385 | 0.95 | ±0.10 | 0.96 | ±0.10 | 0.91 | ±0.09 | 1.13 | ±0.05 | I, MS |
| 7-epi-Sesquithujene | 1391 | 0.06 | ±0.00 | 0.09 | ±0.01 | 0.08 | ±0.01 | 0.11 | ±0.01 | I, MS |
| β-Caryophyllene | 1421 | 1.12 | ±0.12 | 1.32 | ±0.03 | 1.48 | ±0.14 | 1.95 | ±0.04 | I, MS, Co-GC |
| trans-β-Farnesene | 1458 | 0.64 | ±0.05 | 0.66 | ±0.00 | 0.68 | ±0.07 | 1.14 | ±0.11 | I, MS |
| Germacrene D | 1483 | 0.30 | ±0.03 | 0.34 | ±0.04 | 0.37 | ±0.03 | 0.56 | ±0.01 | I, MS |
| Lavandulyl isovalerate | 1511 | 0.21 | ±0.01 | 0.24 | ±0.00 | 0.29 | ±0.03 | 0.42 | ±0.01 | I, MS |
| α-Bisabolol | 1687 | 0.52 | ±0.05 | 0.74 | ±0.01 | 0.74 | ±0.02 | 0.92 | ±0.03 | I, MS |
| Total | 97.62 | 97.72 | 98.57 | 97.20 | ||||||
| Alcohols | 37.30 | 37.30 | 37.08 | 33.67 | ||||||
| Esters | 47.20 | 47.63 | 49.48 | 53.62 | ||||||
| Ethers | 1.94 | 1.75 | 1.18 | 0.46 | ||||||
| Hydrocarbons | 6.85 | 7.14 | 7.03 | 6.63 | ||||||
| Ketones | 4.42 | 3.90 | 3.80 | 2.83 | ||||||
| KB494 | F: CCCTTCTTGAGGCTCTTGAC | EF1-a [26] |
| KB495 | R: GCACAGTTCCAATACCACC | |
| KB479 | F: ACACGCACGACAATTTGCCA | LINS [27] |
| KB480 | R: AGCCCTCCAATGAAGTGGGAT | |
| KB481 | F: GCGCCACACAACTAGAAATTAAGT | LIMS [26] |
| KB482 | R: TTGCACAGTCAGCTCAGCG | |
| KB477 | F: ACTACACTGGAGGGTGCAAAGA | TPS-1 [27] |
| KB478 | R: AATCTGGAACTCGCATTTGGCG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seira, E.; Poulaki, S.; Hassiotis, C.; Poulios, S.; Vlachonasios, K.E. Gene Expression of Monoterpene Synthases Is Affected Rhythmically during the Day in Lavandula angustifolia Flowers. Physiologia 2023, 3, 433-441. https://doi.org/10.3390/physiologia3030030
Seira E, Poulaki S, Hassiotis C, Poulios S, Vlachonasios KE. Gene Expression of Monoterpene Synthases Is Affected Rhythmically during the Day in Lavandula angustifolia Flowers. Physiologia. 2023; 3(3):433-441. https://doi.org/10.3390/physiologia3030030
Chicago/Turabian StyleSeira, Eleftheria, Stefania Poulaki, Christos Hassiotis, Stylianos Poulios, and Konstantinos E. Vlachonasios. 2023. "Gene Expression of Monoterpene Synthases Is Affected Rhythmically during the Day in Lavandula angustifolia Flowers" Physiologia 3, no. 3: 433-441. https://doi.org/10.3390/physiologia3030030
APA StyleSeira, E., Poulaki, S., Hassiotis, C., Poulios, S., & Vlachonasios, K. E. (2023). Gene Expression of Monoterpene Synthases Is Affected Rhythmically during the Day in Lavandula angustifolia Flowers. Physiologia, 3(3), 433-441. https://doi.org/10.3390/physiologia3030030