Abstract
Drought is an important abiotic stress factor limiting crop productivity worldwide and its impact is increasing with climate change. Regardless of the plant growth period, drought has a deadly and yield-reducing effect on the plant at every stage of development. As with many environmental stressors, drought-exposed plants trigger a series of molecular, biochemical, and physiological responses to overcome the effect of drought stress. Currently, researchers are trying to determine the complex functioning of drought stress response in plants with different approaches. Plants are more sensitive to drought stress during certain critical stages like germination, seedling formation, flowering, fertilization, and grain formation periods. Plants have high success in reducing the effects of drought stress in vegetative development periods with the activity of tolerance mechanisms. On the other hand, drought stress during the generative period can cause irreversible losses in yield. This review focuses on the progression of molecular, biochemical, and physiological mechanisms involved in the drought stress tolerance in plants and the responses of field crops to drought stress at different development stages.
1. Introduction
Agricultural production is directly affected by climatic conditions. The direct or indirect effects of climatic change (e.g., temperature differences to seasonal norms and irregularities in the precipitation regime) limit plant development and yield [1,2,3]. Water is one of the essential factors for the sustainability of life of all living organisms, including plants. Plants need water for photosynthesis and metabolic activities [4]. Besides, the plants should use the maximum level of water from the environment to continue their growth performance [5]. Drought is a physiological form of water deficiency in which the soil water available to the plant is insufficient and adversely affects its metabolism [6]. Against the negative effects caused by drought, plants manage this process with a complex set of related mechanisms [7]. Physiological and metabolic changes that occur as a result of the interaction of these mechanisms help tolerate the negative effects of stress [8].
Plant stress response mechanisms are controlled by complex networks determined by environmental and genetic factors. Traditional methods are insufficient to control and explain the complex tolerance mechanism [9]. In this respect, omic technologies are promising for improving drought stress tolerance with many biotechnological approaches [8]. The focus of these studies is genome-wide research to discover stress-related candidate regions and genes for stress resilience [2]. Many studies have been carried out on functional genes involved in the stress response, with methods such as QTL (quantitative trait loci) analysis, transcriptomic analysis, and GWAS (genome-wide association study) in important crop species [10,11,12,13]. The identified target genes contributed to the improvement of tolerance to stress through gene silencing techniques, transgenic approaches, and genome engineering (CRISPR/Cas9) methods [2,8].
Different defense mechanisms help plants deal with the stress of drought. The plant responds to drought stress with biochemical (antioxidant content, chlorophyll content, proline accumulation, hormonal content, secondary metabolite, etc.), physiological (activity of stomata, photosynthesis, osmotic balance, transpiration, leaf water content, water transmission), and morphological changes (decreased leaf area, number of leaves, increase in root length, leaf aging, early maturation, change in growth stages, etc.). This is due to several molecular mechanisms that are put into action (increased expression of transcription factor genes) [14,15,16,17]. Additionally, the plant’s stress response and coping mechanisms depend on its growth stage when it experiences drought stress [18]. Depending on the stage of their growth, plants may be more or less sensitive to drought stress. Abnormalities occur in the turgor pressure, leaf water content, stomatal movement, leaf coloration, photosynthesis and respiration, leaf vitality, and ultimately growth activities when drought stress is experienced during the vegetative development cycle. These responses might encourage the plant to keep its vegetative period brief and move quickly through the generative stage [7]. Drought stress exposure during the generative development period causes reductions in flowering rate, fertilization, seed setting and product quality [19,20]. Many researchers have investigated the effects of drought stress in sorghum [21,22,23], maize [24,25], wheat [19,26,27], rice [20,28], mung bean [29,30,31], soybean [32,33] and lentil [3]. Nevertheless, depending on the severity and duration of drought stress, the growth period of the plant is an important factor in managing its response to stress [34].
This review focuses on explaining the responses of field crops to drought stress, especially during important developmental stages, by providing information about the interaction between physiological, biochemical and molecular mechanisms underlying drought stress tolerance in plants.
2. Drought Stress Signaling and Molecular Regulation
Drought stress negatively affects plant growth with various physiological and biochemical processes such as respiration, translocation, ion uptake, water potential, stomatal closure, photosynthesis, sugar and nutrient metabolism, antioxidant system, and phytohormones [35]. The activation of numerous genes with different functions causes the plant to undergo physiological and biochemical reactions in response to drought stress [36,37]. Molecular mechanisms related to drought tolerance are studied under two main categories. The first is signal transduction factors, including protein kinases, transcription factors, and ABA receptors. The other is functional factors, including proteins involved in metabolism, osmotic regulation, protein conversion, protein modification, and ROS transport [38].
Stress signal activation occurs through protein molecules that are activated by disruption of the cell wall [39]. The diverse signaling pathways of water deficiency stress in plants consist of several proteins, including transcription factor (TFs), enzymes, molecular chaperones, and several metabolites [40]. Many genes have been identified that are differentially expressed in plants in response to drought [41,42]. These genes function in different cellular signaling pathways and cellular responses such as transcriptional regulation [43,44]. Transcriptional factors consist of the DREB, WRKY, bZIP, bHLH, NAC, MYC, MYB gene families and protein kinases (mitogen-activated protein kinases (MAPK), calcium-dependent protein kinases (CDPK), are composed of receptor protein [45,46]. Numerous TF genes associated with stress have been discovered in various plant species [6]. Plants use both pathways categorized as abscisic acid (ABA)-dependent and ABA-independent signaling pathways to sense and respond to drought stress [47]. During signal transduction, ABA-independent TFs serve as molecular switches, directly regulating the expression of associated genes by interacting with cis-elements in the promoter region of genes [48,49]. This is based on the specific nature of the DNA binding sites themselves [41,50]. In this way, TF genes are involved in the expression of a specific gene in the event of drought stress [49,51,52].
Another mechanism implicated in stress signaling is an increase in the generation of reactive oxygen species (ROS). ROS signaling is associated with abscisic acid (ABA) and Ca2+ increase under drought stress in plants [53]. Overproduction and accumulation of ROS in different tissues and cells of the plant are considered stress signals [54]. In addition, protective molecules such as low molecular weight osmolytes (sugars, polyols, amino acids such as proline), heat shock proteins, aquaporins, and LEA proteins are involved in the responses to stress in plants [55]. Proteins are synthesized in the plant cell by gene expression as a result of stress signaling. Synthesized proteins are responsible for biochemical, physiological, and morphological activities such as transcriptional regulation, cell membrane protection, antioxidant biosynthesis, initiating or stopping physiological activities, and uptake of water and ions [56].
3. Biochemical Reactions of Plants in Drought Stress
Drought resistance is a complex set of events involving the interaction of different stress-sensitive mechanisms [57]. Arid and semi-arid environmental conditions induce the formation of ROS in plants and cause oxidative damage in plant cells. ROS signaling is involved in the initiation of stress-induced molecular, biochemical, physiological, and morphological responses [58,59,60]. ABA, a crucial component of stress signaling, is produced when ROS production increases in response to stress [61]. In this way, it can regulate gene expression for biochemical responses by producing superoxide dismutase (SOD) and catalase (CAT) [62].
High levels of ROS production can damage various physiological and metabolic processes such as photosynthesis and the antioxidant defense system in plants [58]. The antioxidant system and osmotic regulation are the main defense systems that provide the tolerance of plants against water deficiency stress conditions. CAT, peroxidase (POD), SOD, glutathione reductase (GR), ascorbate peroxidase (APX), glutathione peroxidase (GPX) are enzymatic antioxidants, and phenolic compounds are non-enzymatic antioxidants (e.g., ascorbic acid, vitamins, carotenoids, phenolic compounds) [63].
SOD is the first line of defense in the presence of ROS. CAT and APX detoxify ROS and prevent its accumulation in cells and tissues [64]. However, non-enzymatic antioxidants such as flavonoids and tannins, which are phenolic compounds, play a significant role in ROS detoxification and mitigate the effects of oxidative stress [65]. Natural antioxidants maintain cellular redox balance by binding and neutralizing free radicals for plant survival under stress conditions [66]. The antioxidant defense system, consisting of the coordinated action of enzymatic and non-enzymatic antioxidants, provides an effective mechanism to control the toxicity induced by ROS. Plants respond to drought stress by accumulating soluble chemicals in the cytoplasm, such as proline, glycine-betaine, glucose, fructose, mannitol, inositol, valine, isoleucine, etc. These metabolites don’t interfere with the functioning of metabolic reactions under normal conditions. However, in case of stress conditions, they act as an osmoprotectant to regulate the osmotic balance of the plant, maintain water flow and molecular stability, and prevent the accumulation of stress-related free radicals [51,67]. Among the osmoprotectants, proline is one of the important amino acid that have high antioxidant properties and plays a role in the prevention of cell death [68]. Many researchers consider stress-related proline accumulation as a biochemical marker for tolerant cultivar selection [69]. Glycine-betaine acts as a protective in protein unfolding and denaturation through direct and indirect interaction with macromolecules [70]. Mannitol increases enzymatic antioxidant activities in plants. Exogenous application of mannitol increased catalase and ascorbate peroxidase activities in wheat shoots and roots [71]. Sucrose is another important osmolyte found in abundance in the plant. It supports anthocyanin accumulation to scavenge reactive oxygen species and plays an important role in reducing the effect of ROS [72,73].
4. Physiological Responses and Mechanisms of Plants against Drought Stress
The effects of drought stress include decreased plant cell growth, stomatal closure, irregular turgor pressure, decreased leaf water content, accumulation of biochemical substances, poor root-absorption function, reduced photosynthetic activity, impaired metabolism, and plant mortality [74]. Plant response to drought stress is managed by molecular, biochemical and physiological mechanisms (Figure 1).
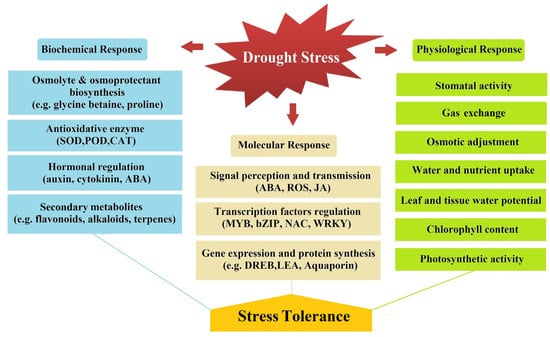
Figure 1.
Relation of molecular, biochemical, and physiological responses to drought stress.
The physiological response of plants to drought stress consists of long-term and short-term responses [75]. The long-term negative impact of drought stress on the plant includes processes disruption of leaf/root physiological cycles, changes in maturity times (early productive maturity), and yield losses [76]. Short-term reactions to drought in plants include changes in stomatal conductivity, water potential across tissues, water and nutrient uptake movements of roots, turgor pressure, and biochemical composition [77]. Plants can transmit positive and negative signals between roots and shoots for adaptation to environmental conditions [78]. The stress factor in the environment can cause a reaction in the shoots with the signals transmitted from the roots. As a result, the vital functions of the plant may decrease with some active physiological processes [43,79]. Many factors including abscisic acid (ABA), auxin, cytokinins, ethylene, gibberellins, strigolactone (SL), jasmonic acid (JA), and proline act as signal molecules under variable environmental stresses and play a role in the regulation of physiological processes [39,80,81,82]. Strigolactone (SL) is a plant hormone that affects physiological processes such as shoot branching, root elongation, and leaf senescence [83]. Besides, SL acts as a signal molecule for drought stress tolerance [84]. The increased level of SL biosynthesis gene expression under drought stress is one of the important regulators in plant response to stress tolerance [85,86].
Alterations in the cellular ROS due to biochemical response affect various metabolic and physiological reactions in the plant. Certain ROS also acts as a signaling molecule in stress adaptation [87,88] addition, the roots create stress-related hormones and osmoprotectants when they detect a scarcity of water in the soil, and they then direct these substances to the shoot via transpiration current [80]. These substances accumulate in leaf tissues and cause the initiation of molecular, biochemical, and physiological processes. Oğuz et al. [89] stated that under the influence of drought stress, leaf tissues were physiologically more affected than root tissues and also displayed relatively higher TF gene expression.
The first physiological response of plants under the influence of drought stress is to reduce transpiration by stomata [90]. The closure of the stomata and the reduction of water loss by the plant is a physiological response to avoid drought [91,92]. On the other hand, the stomata’s closure influences physiological and biochemical processes, such as a reduction in leaf water content, chlorophyll quantity, chloroplast fragmentation, gas interaction, ion exchange between root and shoot, and photosynthesis, while suppressing leaf expansion morphologically [93,94,95,96,97]. As a result, all these processes and physiological events affected photosynthetic activity directly or indirectly [98,99,100]. Plants control gas and water flow through the stomata in leaf tissue. The closure of stomata due to drought prevents the use of CO2, which is of great importance for photosynthesis [101]. The reduction of CO2 uptake by the plant directly causes low photosynthetic activity [102]. Decreased transpiration due to the closure of stomata under water-deficient conditions also limits the absorption of nutrients from the soil through the roots and their translocation to the upper parts of the plant [24,103] (Figure 2). This situation causes a dramatic decrease in the nutrient concentration of plant tissues and ion balance [79,103,104]. Many processes are adversely affected due to the disruption of nutrient, mineral, water, and gas flow in plant tissues [105,106,107]. Relative water content (RWC) is another important physiological feature that affects leaf water potential, stomatal resistance, transpiration rate, and plant water relations [90]. Relative water content is considered a marker of plant water status, which regulates metabolic activity in tissues. RWC is formed as a result of water loss by transpiration and uptake by roots [18,108,109] (Figure 2). Leaf water potential, which is important for plant survival and photosynthetic processes; turgor pressure is closely related to stomatal closure and cell growth [110,111]. Maintaining the leaf water potential allows for the tolerance of low to moderate water stress. However, the reduced efficiency of photosynthesis is brought on by the rise in leaf water potential loss brought on by increased water stress [112].
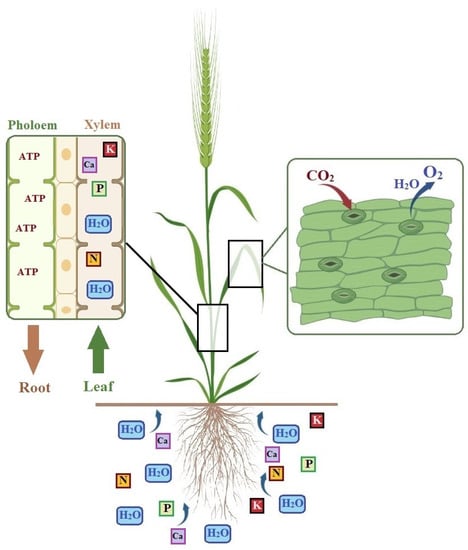
Figure 2.
Transpiration through stomata and the movement of water and nutrients from the soil. Leaf transpiration and gas exchange are controlled by stomata. The water movement in the plant is controlled by the loss of water by transpiration from the leaves and the movement of water uptake from the root. Water and dissolved compounds are absorbed from the soil by the roots and transported to the upper parts of the plant through the xylem. The energy (ATP) produced in the upper parts of the plant is moved to the other parts by the phloem. Illustrations such as Ca (calcium), K (potassium), and N (nitrogen) are representative.
Photosynthesis is the most important physiological process directly related to growth, development, and yield in all green plants [113]. Chloroplasts are cellular organelles and are important for photosynthesis. With the help of metabolites synthesized during photosynthesis and key proteins involved in the metabolic process, chloroplasts provide resistance against various abiotic stresses such as drought [114]. Deterioration in the chloroplast structure due to drought adversely affects the synthesis of chlorophyll [113]. Chlorophyll is one of the main chloroplast components for photosynthesis, and chlorophyll content has a positive relationship with the rate of photosynthesis. The decrease in chlorophyll content under drought stress has been considered a typical manifestation of oxidative stress [115]. The reduction in chlorophyll content due to drought stress is the result of pigment photo-oxidation and chlorophyll degradation [18]. The reduction in photosynthetic pigment concentrations such as chlorophyll due to environmental stressors could directly limit the production of photosynthetic activities [79].
5. Management of Drought Stress in Plants
From past to present, a number of important agronomic strategies have been developed to increase plant adaptation to abiotic stress factors caused by climate change [116]. Fertilization and irrigation treatment according to the development periods of the plants and the selection of the appropriate tillage system are of great importance in preventing yield losses in plants under the drought stress [117]. In addition, some strategies such as sowing time, sowing frequency, sowing to stubble, crop rotation, selection of plant varieties with short life cycles, optimum irrigation practices, and use of bio-fertilizers are the key management techniques to obtain higher productivity of crops [118,119,120,121]. Except that the methods and practices developed by farmers and researchers for stress management, there are a number of mechanisms that plants have developed to manage drought stress. The effect of drought on the plant depends on the severity of the stress and the developmental stage of the plant [122]. The effects of continuous and intense drought stress and the effects of short-term and low-level drought stress effects different on the plant. The severity and timing of drought stress change the plant’s response to drought stress. Stress responses of the plant can be grouped under three different headings as escape, avoidance, and tolerance (Figure 3).
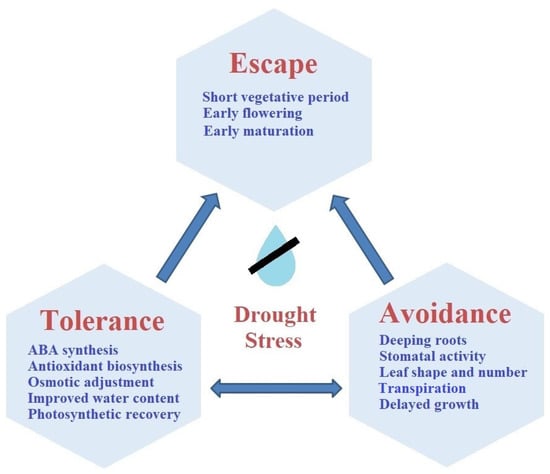
Figure 3.
Drought stress management of the plant.
Tolerance includes molecular, biochemical, and physiological responses mediated by osmotic regulation, accumulation of osmoprotectant (e.g., proline), ABA biosynthesis, and stomatal activities [7]. Stress avoidance by plants is the ability of the root to move deeper to reach water, the closing of stomata, leaf rolling, and efficient use of available water by plants [18]. To escape drought the vegetative cycle is completed in a short time. Plants pass the generative stage quickly. This situation leads to early flowering and seed formation [123]. These reactions occur as a result of the effect of multiple stress tolerance mechanisms operating within the plant.
6. Drought Stress Effect on Different Development Stages
Many processes that affect growth and development in plants are impacted by water deficiency stress. Yield is the final stage of these processes. The effect of drought on the plant varies according to the genotype, the intensity and duration of the drought stress, and the growth stage of the plant [14,124].
Growth is established through cell division, cell growth, and differentiation. Low turgor pressure greatly limits cell growth [125]. The mitosis process disrupted by drought causes decreased cell growth and development [126]. Consequently, cell growth is one of the most drought-sensitive physiological processes. It has been stated that cell growth responds to even mild drought stress and can be adversely affected [127]. With the continuation of drought stress and the increase in stress severity, cell death can occur. This situation causes the metabolism to be disrupted and thus the physiological functioning to be damaged [128].
Plants can be exposed to drought stress for many periods from germination to harvest. Drought stress is an important factor that directs germination, seedling formation, root and shoot growth, tillering, flowering initiation, pollination, fertilization, seed yield, and quality [129]. Plants are susceptible to drought stress during all growth periods [130]. The growth and development phases of the plant, such as germination, seedling formation, and tillering constitute the vegetative development stages. Generative growth consists of flowering, fertilization, seed formation, and grain-filling periods. Drought can affect the vegetative and generative stages of the plant differently [14,131]. Researchers focused on examining the effects of drought on yield and quality in crop production during these critical developmental periods [18].
6.1. Drought Stress in the Vegetative Stage
The germination of plant seeds marks the start of vegetative growth. A complicated series of biochemical and physiological processes occurs inside the seed to start plant development and embryo growth. Seeds undergo biochemical changes quickly after absorbing water. The amount of water in the environment affects how much water is taken in and absorbed [132]. For successful germination, seeds must reach a sufficient level of hydration during the absorption phase to reactivate the metabolic processes and stimulate the growth of the embryonic axis. Under drought stress, more time is needed to adjust the osmotic potential of the seed [133]. Therefore, there is a delay in the absorption process [21]. As the germination rate of seeds that do not reach the required hydration level decreases and the germination delay increases, poor seedling formation occurs [134]. Decreased germination rate and poor seedling growth are early signs of drought stress [132]. The negativities encountered in the early stage of plant growth due to drought stress reduce plant establishment per unit area [135,136]. Many researchers have reported that drought has negative effects on physiological processes in the early development stage such as germination, coleoptile length, shoot, and root length [21,137,138,139,140,141]. Coleoptile length is an important parameter that affects the emergence of the germinated seed in the soil and the success of seedling formation. Under the condition of water deficiency, suppression of coleoptile and root formation may occur due to turgor pressure-induced negativities in early cell division and growth. This has important consequences that affect shoot elongation and root growth in the afterward growth stages of the plant [21,133,142].
Varieties that show long and widespread root development in tolerance to drought stress can have successful seedling formation with advanced root systems that can reach deep for water uptake from the soil [143]. This feature is one of the mechanisms developed by the plant to avoid drought stress. Besides, root characteristics such as the number of roots, diameter, angle, depth, total length, distribution, and biomass of the plant in the later stages of the vegetative growth period are closely related to drought tolerance [144,145].
Physiologically drought-resistant varieties have high water use efficiency. They can produce better photosynthetic activity and energy with low water consumption and low transpiration rate mechanism [146,147]. Mega et al. [148] reported that the phytohormone abscisic acid (ABA) is involved in regulating water use, directly regulating stomatal opening and perspiration. Plant growth is positively affected as a result of decreased transpiration and the accompanying increase in photosynthetic activity and increased water use efficiency.
To maintain a balance between the water received by their roots and the hydration status of plant tissues, plants can restrict leaf elongation when they are under water stress [149]. A reduction in the number of leaves per plant, a reduction in leaf size, and an increase in leaf senescence are only a few of the detrimental effects of drought stress during the vegetative period [150,151,152,153]. Another important physiological response that occurs in response to water stress is leaf rolling. It is thought that leaf rolling aims to reduce the transpiration rate of the plant [38,154]. The purpose of the plant with these physiological responses is to keep water loss to a minimum [18]. However, photosynthetic activities may be damaged due to decreased gas assimilation, decreased amount of chlorophyll, and impaired physiological and biochemical balance such as RWC [14,79,155,156,157].
Drought has a great effect on the intake of minerals and nutrients needed by the plant from the soil. The decrease in soil water content due to drought adversely affects the water content in the cells and tissues of the plant [158,159]. In addition, water is an important factor in dissolving the supplements necessary for plant growth and development in the soil. Due to the decrease in the assimilation of the roots, there are problems in the uptake of these nutrients [160,161,162]. The effects of drought in the vegetative period are decreased fresh and dry biomass production, delayed tillering, shorter first internodes, early maturity and unexpected plant losses [163]. According to Semerci et al. [164] pointed to a significant reduction in growth, including shoot length, biomass, and leaf number, due to low turgor pressure that drives the plant to stunted growth during drought stress. Panda et al. [165]; reported that drought stress during the vegetative growth period was effective on RWC, the number of siblings, plant biomass, and grain yield. In addition, significant and positive correlations were observed between yield and physiological and biochemical properties such as proline content, relative water content, catalase activity, peroxidase activity, total chlorophyll content, and plant biomass under drought stress conditions [165]. Hossain et al. [166] reported a significant decrease in the number of days to flowering, plant height, seeds, and yield per plant under drought stress. According to Bangar et al. [30] stated that significant decreases in RWC, membrane stability index (MSI), proline content of leaves, leaf area, plant height, and yield occur under drought stress in the vegetative and reproductive stages.
Most of the molecular, physiological, and morphological studies carried out in the explanation of drought stress are generally carried out at the seedling stage (Vegetative period). However, it has been suggested that the most important period for sensitivity to drought is the vegetative and generative phases merge. The physiological merge stage represents vegetative growth ends and flower formation begins, and the transition to the generative stage.
6.2. Drought Stress in the Generative Stage
Drought stress on the vegetative growth and development stages of the plant can affect the yield. However, the plant’s exposure to stress in the generative stage has a more severe effect on fertilization and grain yield. The plant vegetative stage takes longer than the generative period. Therefore the plant has more time to respond to stress and improve physiological processes. Conversely, the stresses encountered in the generative stage can cause irreversible results. The period of plants such as pollination, fertilization, grain formation, and grain filling in the generative stage are critical periods when yield losses can be high [22,157,167].
Plants undergo substantial changes during their flowering phase after the vegetative phase has ended. Both the beginning and the duration of these crucial developmental stages are impacted by drought. Plants typically reduce the amount of time between the start of flowering and blooming when there is a moderate drought, in an effort to avoid it. However, under the effect of a severe drought, this time frame could be extended [168].
Arid conditions decrease the development progress as a result of the decrease in the amount of photosynthesis resulting in a loss in flowering formation, grain filling, and yield [102]. Drought during flowering often results in sterility. One of the most important causes of sterility is the insufficient flow of nutrients and minerals to the developing generative area [79,169,170]. On the other hand, drought stress causes anthesis (blooming and dying of the flower bud). The reproductive phase shortened by anthesis occurs as a result of the tendency of plants to escape from stress [171]. Ranawake et al. [172] stated that water stress significantly affects the flowering and pod-filling period.
Vadez et al. [173] stated that the plant’s tolerance to drought stress increases as a result of the improvement of physiological activities such as transpiration rate and water use efficiency in the pre-blooming period. Physiological adaptations such as efficient use of water during the vegetative process, low stomatal movement, and maintaining the balance in turgor pressure ensure the preservation of the water needed by the plant during the grain filling phase [174]. Besides, high chlorophyll content positively influences flowering and reproduction periods with increased photosynthetic activities [175]. On the other hand, plants are more sensitive to drought stress during the flowering period [176]. Drought stress delays the flower formation stage and negatively affects fertilization, cluster development, and seed formation processes [177]. Therefore, drought stress at the flowering stage has a strong irreversible effect on yield [28].
During the generative period, drought stress significantly affects grain yield and quality by reducing seed size, number, and seed weight per plant [22,157]. The decrease in grain filling rate occurs due to inactivation in energy synthesis, carbohydrate metabolism, sucrose and starch production as a result of disruptions in the photosynthesis process [19,161]. Drought stress at the stage of fertilization and cob formation in maize led to a significant decrease in components such as grain order, grain number, 1000 grain weight, grain yield per plant, biological yield, and harvest index [14]. Cakir [34] reported that water stress during the cob formation period causes yield loss of up to 40% in maize. Rizza et al. [152] reported that drought in the reproductive growth stage of wheat reduced the yield in the number of grains by up to 72% due to anthesis. One of the adaptation mechanisms that plants have developed to escape from drought stress is the short seed-filling period. However, losses in seed yield occur as a result of the shortening of seed filling time and reduced seed size due to drought stress [178]. Felisberto et al. [32] stated that the lack of water encountered in the grain-filling stage of soybean is of critical importance for yield.
7. Future Scope of Research for Agricultural Sustainability of Crops
In order to meet the increasing food demand of the world population, preventing yield losses due to abiotic stresses in agricultural production is crucial. A prerequisite for increasing plant stress tolerance is an understanding of plant response mechanisms to stress. Innovative and biotechnological methods are of great importance in increasing tolerance to abiotic stress [8]. In this respect, researchers use different omic approaches to develop plant stress tolerance [2].
Local populations are important resources that should be used in plant breeding and selection studies that are resistant to drought stress thanks to their wide genetic variety [117,179]. Especially, agronomic plant traits of local plant populations used in drought-resistant cultivar breeding and their performance under stress conditions should be considered. Because the yield in field crops depends on some agronomic plant traits such as plant height, number of spikes, grain weight, harvest index, thousand seed weight and grain yield [180].
In recent years, researchers have focused to prevent yield losses in plants with environmentally friendly innovative approaches. In changing climatic conditions and under abiotic stress factors, the use of bio-fertilizers, bio-stimulants and agro-industrial wastes as a compost is important for sustainable agriculture [181,182,183].
Most of the studies to increase stress tolerance target a specific developmental period of the plant. However, the reactions of plants vary according to the developmental stages. For this reason, the responses of plants to stress in different growth periods should be targeted. These approaches need to be integrated into agronomy and supported by field trials. Furthermore, conducting field trials in different locations and climatic conditions will make a significant contribution to supporting the arguments.
8. Conclusions
It is obvious that, as it is today and will continue to be in the future owing to global climate change, dryness will be the primary factor restricting crop production. Drought stress affects plant growth and yield. The timing, duration, severity, and speed of stress undoubtedly play an important role in determining a plant’s response to a lack of water. However, drought is a difficult situation to control under natural conditions. The response of plants to stress at different growth stages is an important criterion for the development of varieties with high-stress tolerance. The response of plants to stress occurs as a result of the cooperation of molecular, biochemical, physiological, and morphological mechanisms. Each of these mechanisms is very complex to be considered separately. Focusing on the differences in the activation and regulation of these mechanisms during important development stages of the plant may lead to new approaches. In this review, we tried to explain the response to drought stress in the critical vegetative and generative periods. Consequently, determining the effect of drought on the critical growth stages will guide the studies to be carried out to prevent yield losses.
Author Contributions
M.C.O. design and writing; M.A., E.O., I.P. and M.Y. review and editing; M.C.O. and E.O. editing figures; M.A. and I.P. editing references; M.A. submission; M.Y. supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the FOSC project (Sus-Agri-CC) from the European Union’s Horizon 2020 research and innovation program under grant agreement 220N247 to M.Y.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yohannes, G.; Kidane, L.; Abraha, B.; Beyene, T. Effect of Salt Stresses on Seed Germination and Early Seedling Growth of Camelina sativa L. Momona Ethiop. J. Sci. 2020, 12, 1–19. [Google Scholar] [CrossRef]
- Billah, M.; Aktar, S.; Brestic, M.; Zivcak, M.; Khaldun, A.B.M.; Uddin, M.S.; Bagum, S.A.; Yang, X.; Skalicky, M.; Mehari, T.G.; et al. Progressive Genomic Approaches to Explore Drought- and Salt-Induced Oxidative Stress Responses in Plants under Changing Climate. Plants 2021, 10, 1910. [Google Scholar] [CrossRef]
- el Haddad, N.; Choukri, H.; Ghanem, M.E.; Smouni, A.; Mentag, R.; Rajendran, K.; Hejjaoui, K.; Maalouf, F.; Kumar, S. High-Temperature and Drought Stress Effects on Growth, Yield and Nutritional Quality with Transpiration Response to Vapor Pressure Deficit in Lentil. Plants 2022, 11, 95. [Google Scholar] [CrossRef]
- Oo, A.T.; van Huylenbroeck, G.; Speelman, S. Measuring the Economic Impact of Climate Change on Crop Production in the Dry Zone of Myanmar: A Ricardian Approach. Climate 2020, 8, 9. [Google Scholar] [CrossRef]
- Tátrai, Z.A.; Sanoubar, R.; Pluhár, Z.; Mancarella, S.; Orsini, F.; Gianquinto, G. Morphological and Physiological Plant Responses to Drought Stress in Thymus citriodorus. Int. J. Agron. 2016, 2016, 4165750. [Google Scholar] [CrossRef]
- Kumar, S.; Sachdeva, S.; Bhat, K.V.; Vats, S. Plant Responses to Drought Stress: Physiological, Biochemical and Molecular Basis. In Biotic and Abiotic Stress Tolerance in Plants; Springer: Berlin/Heidelberg, Germany, 2018; pp. 1–25. ISBN 978-981-10-9029-5. [Google Scholar]
- Pamungkas, S.S.T.; Suwarto; Suprayogi; Farid, N. Drought Stress: Responses and Mechanism in Plants. Rev. Agric. Sci. 2022, 10, 168–185. [Google Scholar] [CrossRef]
- Hossain, A.; Farooq, M.; el Sabagh, A.; Hasanuzzaman, M.; Erman, M.; Islam, T. Morphological, Physiobiochemical and Molecular Adaptability of Legumes of Fabaceae to Drought Stress, with Special Reference to Medicago sativa L. In The Plant Family Fabaceae; Springer: Berlin/Heidelberg, Germany, 2021; pp. 289–317. [Google Scholar] [CrossRef]
- Sinclair, T.R. Challenges in Breeding for Yield Increase for Drought. Trends Plant Sci. 2011, 16, 289–293. [Google Scholar] [CrossRef]
- Gahlaut, V.; Jaiswal, V.; Singh, S.; Balyan, H.S.; Gupta, P.K. Multi-Locus Genome Wide Association Mapping for Yield and Its Contributing Traits in Hexaploid Wheat under Different Water Regimes. Sci. Rep. 2019, 9, 19486. [Google Scholar] [CrossRef]
- Ballesta, P.; Mora, F.; del Pozo, A. Association Mapping of Drought Tolerance Indices in Wheat: QTL-Rich Regions on Chromosome 4A. Sci. Agric. 2020, 77, e20180153. [Google Scholar] [CrossRef]
- Swamy, B.P.M.; Kaladhar, K.; Anuradha, K.; Batchu, A.K.; Longvah, T.; Sarla, N. QTL Analysis for Grain Iron and Zinc Concentrations in Two O. Nivara Derived Backcross Populations. Rice Sci. 2018, 25, 197–207. [Google Scholar] [CrossRef]
- Cormier, F.; le Gouis, J.; Dubreuil, P.; Lafarge, S.; Praud, S. A Genome-Wide Identification of Chromosomal Regions Determining Nitrogen Use Efficiency Components in Wheat (Triticum aestivum L.). Theor. Appl. Genet. 2014, 127, 2679–2693. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.A.; Wang, L.C.; Farooq, M.; Hussain, M.; Xue, L.L.; Zou, C.M. Brassinolide Application Improves the Drought Tolerance in Maize Through Modulation of Enzymatic Antioxidants and Leaf Gas Exchange. J. Agron. Crop Sci. 2011, 197, 177–185. [Google Scholar] [CrossRef]
- Haworth, M.; Elliott-Kingston, C.; McElwain, J.C. Co-Ordination of Physiological and Morphological Responses of Stomata to Elevated [CO2] in Vascular Plants. Oecologia 2013, 171, 71–82. [Google Scholar] [CrossRef]
- Conesa, M.R.; de la Rosa, J.M.; Domingo, R.; Bañon, S.; Pérez-Pastor, A. Changes Induced by Water Stress on Water Relations, Stomatal Behaviour and Morphology of Table Grapes (Cv. Crimson Seedless) Grown in Pots. Sci. Hortic. 2016, 202, 9–16. [Google Scholar] [CrossRef]
- Ammar, M.H.; Anwar, F.; El-Harty, E.H.; Migdadi, H.M.; Abdel-Khalik, S.M.; Al-Faifi, S.A.; Farooq, M.; Alghamdi, S.S. Physiological and Yield Responses of Faba Bean (Vicia faba L.) to Drought Stress in Managed and Open Field Environments. J. Agron. Crop Sci. 2015, 201, 280–287. [Google Scholar] [CrossRef]
- Nezhadahmadi, A.; Prodhan, Z.H.; Faruq, G. Drought Tolerance in Wheat. Sci. World J. 2013, 2013, 610721. [Google Scholar] [CrossRef]
- Mahla, R.; Madan, S.; Kaur, V.; Munjal, R.; Behl, R.K.; Midathala, R. Activities of Sucrose to Starch Metabolizing Enzymes during Grain Filling in Late Sown Wheat under Water Stress. J. Appl. Nat. Sci. 2017, 9, 338–343. [Google Scholar] [CrossRef]
- Akram, R.; Fahad, S.; Masood, N.; Rasool, A.; Ijaz, M.; Ihsan, M.Z.; Maqbool, M.M.; Ahmad, S.; Hussain, S.; Ahmed, M.; et al. Plant Growth and Morphological Changes in Rice under Abiotic Stress. In Advances in Rice Research for Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Nahar, K., Biwas, J.K., Eds.; Woodhead Publishing: Shaxton, UK, 2018; pp. 69–85. [Google Scholar]
- Queiroz, M.S.; Oliveira, C.E.S.; Steiner, F.; Zuffo, A.M.; Zoz, T.; Vendruscolo, E.P.; Silva, M.V.; Mello, B.F.F.R.; Cabral, R.C.; Menis, F.T. Drought Stresses on Seed Germination and Early Growth of Maize and Sorghum. J. Agri. Sci. 2019, 11, 310–318. [Google Scholar] [CrossRef]
- Sarshad, A.; Talei, D.; Torabi, M.; Rafiei, F.; Nejatkhah, P. Morphological and Biochemical Responses of Sorghum bicolor (L.) Moench under Drought Stress. SN Appl. Sci. 2021, 3, 81. [Google Scholar] [CrossRef]
- Sanjari, S.; Shobbar, Z.S.; Ghanati, F.; Afshari-Behbahanizadeh, S.; Farajpour, M.; Jokar, M.; Khazaei, A.; Shahbazi, M. Molecular, Chemical, and Physiological Analyses of Sorghum Leaf Wax under Post-Flowering Drought Stress. Plant Physiol. Biochem. 2021, 159, 383–391. [Google Scholar] [CrossRef]
- Hammad, H.M.; Farhad, W.; Abbas, F.; Fahad, S.; Saeed, S.; Nasim, W.; Bakhat, H.F. Maize Plant Nitrogen Uptake Dynamics at Limited Irrigation Water and Nitrogen. Environ. Scie Pollut. Res. 2017, 24, 2549–2557. [Google Scholar] [CrossRef] [PubMed]
- Goodarzian Ghahfarokhi, M.; Mansurifar, S.; Taghizadeh-Mehrjardi, R.; Saeidi, M.; Jamshidi, A.M.; Ghasemi, E. Effects of Drought Stress and Rewatering on Antioxidant Systems and Relative Water Content in Different Growth Stages of Maize (Zea mays L.) Hybrids. Arch. Agron. Soil Sci. 2015, 61, 493–506. [Google Scholar] [CrossRef]
- Kulczycki, G.; Sacała, E.; Chohura, P.; Załuska, J. Maize and Wheat Response to Drought Stress under Varied Sulphur Fertilisation. Agronomy 2022, 12, 1076. [Google Scholar] [CrossRef]
- Jin, N.; Ren, W.; Tao, B.; He, L.; Ren, Q.; Li, S.; Yu, Q. Effects of Water Stress on Water Use Efficiency of Irrigated and Rainfed Wheat in the Loess Plateau, China. Sci. Total Environ. 2018, 642, 1–11. [Google Scholar] [CrossRef]
- Yang, X.; Wang, B.; Chen, L.; Li, P.; Cao, C. The Different Influences of Drought Stress at the Flowering Stage on Rice Physiological Traits, Grain Yield, and Quality. Sci. Rep. 2019, 9, 3742. [Google Scholar] [CrossRef]
- Prakash, M.; Sunilkumar, B.; Sathiyanarayanan, G.; Gokulakrishnan, J. Screening for Drought Tolerance in Mungbean. Legume Rese 2017, 40, 423–428. [Google Scholar] [CrossRef]
- Bangar, P.; Chaudhury, A.; Tiwari, B.; Kumar, S.; Kumari, R.; Bhat, K.V. Morphophysiological and Biochemical Response of Mungbean [Vigna radiata (L.) Wilczek] Varieties at Different Developmental Stages under Drought Stress. Turk. J. Biol. 2019, 43, 58–69. [Google Scholar] [CrossRef]
- Jincy, M.; Prasad, V.B.R.; Jeyakumar, P.; Senthil, A.; Manivannan, N. Evaluation of Green Gram Genotypes for Drought Tolerance by PEG (Polyethylene Glycol) Induced Drought Stress at Seedling Stage. Legume Res. 2021, 44, 684–691. [Google Scholar] [CrossRef]
- Felisberto, G.; Schwerz, F.; Umburanas, R.C.; Dourado-Neto, D.; Reichardt, K. Physiological and Yield Responses of Soybean under Water Deficit. J. Crop Sci. Biotechnol. 2022. [Google Scholar] [CrossRef]
- Dong, S.; Jiang, Y.; Dong, Y.; Wang, L.; Wang, W.; Ma, Z.; Yan, C.; Ma, C.; Liu, L. A Study on Soybean Responses to Drought Stress and Rehydration. Saudi J. Biol. Sci. 2019, 26, 2006–2017. [Google Scholar] [CrossRef]
- Çakir, R. Effect of Water Stress at Different Development Stages on Vegetative and Reproductive Growth of Corn. Field Crop. Res. 2004, 89, 1–16. [Google Scholar] [CrossRef]
- Singh, A. Soil Salinization Management for Sustainable Development: A Review. J. Environ. Manag. 2021, 277, 111383. [Google Scholar] [CrossRef]
- Tovignan, T.K.; Adoukonou-Sagbadja, H.; Diatta, C.; Clément-Vidal, A.; Soutiras, A.; Cisse, N.; Luquet, D. Terminal Drought Effect on Sugar Partitioning and Metabolism Is Modulated by Leaf Stay-Green and Panicle Size in the Stem of Sweet Sorghum (Sorghum bicolor L. Moench). CABI Agric. Biosci. 2020, 1, 4. [Google Scholar] [CrossRef]
- Impa, S.M.; Sunoj, V.S.J.; Krassovskaya, I.; Bheemanahalli, R.; Obata, T.; Jagadish, S.V.K. Carbon Balance and Source-Sink Metabolic Changes in Winter Wheat Exposed to High Night-Time Temperature. Plant Cell Environ. 2019, 42, 1233–1246. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, C.; Li, L.; Reynolds, M.; Mao, X.; Jing, R. Exploitation of Drought Tolerance-Related Genes for Crop Improvement. Int. J. Mol. Sci. 2021, 22, 10265. [Google Scholar] [CrossRef] [PubMed]
- Razi, K.; Muneer, S. Drought Stress-Induced Physiological Mechanisms, Signaling Pathways and Molecular Response of Chloroplasts in Common Vegetable Crops. Crit. Rev. Biotechnol. 2021, 41, 669–691. [Google Scholar] [CrossRef]
- Song, X.; Li, Y.; Hou, X. Genome-Wide Analysis of the AP2/ERF Transcription Factor Superfamily in Chinese Cabbage (Brassica rapa ssp. Pekinensis). BMC Genom. 2013, 14, 573. [Google Scholar] [CrossRef]
- Joshi, R.; Wani, S.H.; Singh, B.; Bohra, A.; Dar, Z.A.; Lone, A.A.; Pareek, A.; Singla-Pareek, S.L. Transcription Factors and Plants Response to Drought Stress: Current Understanding and Future Directions. Front. Plant Sci. 2016, 7, 1029. [Google Scholar] [CrossRef]
- Rasheed, S.; Bashir, K.; Matsui, A.; Tanaka, M.; Seki, M. Transcriptomic Analysis of Soil-Grown Arabidopsis thaliana Roots and Shoots in Response to a Drought Stress. Front. Plant Sci. 2016, 7, 180. [Google Scholar] [CrossRef]
- Bashir, S.S.; Hussain, A.; Hussain, S.J.; Wani, O.A.; Zahid Nabi, S.; Dar, N.A.; Baloch, F.S.; Mansoor, S. Plant Drought Stress Tolerance: Understanding Its Physiological, Biochemical and Molecular Mechanisms. Biotechnol. Biotechnol. Equip. 2021, 35, 1912–1925. [Google Scholar] [CrossRef]
- Wei, W.; Huang, J.; Hao, Y.J.; Zou, H.F.; Wang, H.W.; Zhao, J.Y.; Liu, X.Y.; Zhang, W.K.; Ma, B.; Zhang, J.S.; et al. Soybean GmPHD-Type Transcription Regulators Improve Stress Tolerance in Transgenic Arabidopsis Plants. PLoS ONE 2009, 4, e7209. [Google Scholar] [CrossRef] [PubMed]
- Fàbregas, N.; Yoshida, T.; Fernie, A.R. Role of Raf-like Kinases in SnRK2 Activation and Osmotic Stress Response in Plants. Nat. Commun. 2020, 11, 6184. [Google Scholar] [CrossRef] [PubMed]
- Lata, C.; Muthamilarasan, M.; Prasad, M. Drought Stress Responses and Signal Transduction in Plants. In Elucidation of Abiotic Stress Signaling in Plants: Functional Genomics Perspectives; Pandey, G., Ed.; Springer: New York, NY, USA, 2015; pp. 195–225. ISBN 978-1-4939-2540-7. [Google Scholar]
- Kim, T.H. Mechanism of ABA Signal Transduction: Agricultural Highlights for Improving Drought Tolerance. J. Plant Biol. 2014, 57, 1–8. [Google Scholar] [CrossRef]
- Villano, C.; Esposito, S.; D’Amelia, V.; Garramone, R.; Alioto, D.; Zoina, A.; Aversano, R.; Carputo, D. WRKY Genes Family Study Reveals Tissue-Specific and Stress-Responsive TFs in Wild Potato Species. Sci. Rep. 2020, 10, 7196. [Google Scholar] [CrossRef]
- Franco-Zorrilla, J.M.; López-Vidriero, I.; Carrasco, J.L.; Godoy, M.; Vera, P.; Solano, R. DNA-Binding Specificities of Plant Transcription Factors and Their Potential to Define Target Genes. Proc. Natl. Acad. Sci. USA 2014, 111, 2367–2372. [Google Scholar] [CrossRef]
- Jin, J.; Zhang, H.; Kong, L.; Gao, G.; Luo, J. PlantTFDB 3.0: A Portal for the Functional and Evolutionary Study of Plant Transcription Factors. Nucleic Acids Res. 2014, 42, D1182–D1187. [Google Scholar] [CrossRef]
- Rao, D.E.; Chaitanya, K.V. Photosynthesis and Antioxidative Defense Mechanisms in Deciphering Drought Stress Tolerance of Crop Plants. Biol. Plant 2016, 60, 201–218. [Google Scholar] [CrossRef]
- Anbazhagan, K.; Bhatnagar-Mathur, P.; Vadez, V.; Dumbala, S.R.; Kishor, P.B.K.; Sharma, K.K. DREB1A Overexpression in Transgenic Chickpea Alters Key Traits Influencing Plant Water Budget across Water Regimes. Plant Cell Rep. 2015, 34, 199–210. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Molecular Responses to Drought Stress in Plants. Biol. Plant 2017, 61, 201–209. [Google Scholar] [CrossRef]
- Hasanuzzaman, M. Plant Ecophysiology and Adaptation under Climate Change: Mechanisms and Perspectives II: Mechanisms of Adaptation and Stress Amelioration; Springer: Singapore, 2020; ISBN 978-981-15-2155-3. [Google Scholar] [CrossRef]
- Kumari, V.V.; Roy, A.; Vijayan, R.; Banerjee, P.; Verma, V.C.; Nalia, A.; Pramanik, M.; Mukherjee, B.; Ghosh, A.; Reja, M.H.; et al. Drought and Heat Stress in Cool-Season Food Legumes in Sub-Tropical Regions: Consequences, Adaptation, and Mitigation Strategies. Plants 2021, 10, 1038. [Google Scholar] [CrossRef]
- Nakashima, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. The Transcriptional Regulatory Network in the Drought Response and Its Crosstalk in Abiotic Stress Responses Including Drought, Cold, and Heat. Front. Plant Sci. 2014, 5, 170. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Selim, M.M.; Alderfasi, A.A.; Afzal, M. Effect of Drought Stress on Mungbean ( Vigna radiata L.) under Arid Climatic Conditions of Saudi Arabia. In Ecosystems and Sustainable Development X; WIT Press: Southampton, UK, 2015; pp. 185–193. ISBN 978-1-84564-900-5. [Google Scholar]
- Zou, Y.N.; Wu, Q.S.; Kuča, K. Unravelling the Role of Arbuscular Mycorrhizal Fungi in Mitigating the Oxidative Burst of Plants under Drought Stress. Plant Biol. 2021, 23 (Suppl. S1), 50–57. [Google Scholar] [CrossRef] [PubMed]
- Saeidnejad, A.H.; Rajaei, P. Antioxidative Responses to Drought and Salinity Stress in Plants, a Comprehensive Review. Int. J. Life Sci. 2015, 9, 1–8. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species Generation, Hazards, and Defense Mechanisms in Plants under Environmental (Abiotic and Biotic) Stress Conditions. In Handbook of Plant and Crop Physiology; CRC Press: Boca Raton, FL, USA, 2021; pp. 509–548. ISBN 978-100-309-364-0. [Google Scholar]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.M.; Zhao, J.; Scandalios, J.G. Cis-Elements and Trans-Factors That Regulate Expression of the Maize Cat1 Antioxidant Gene in Response to ABA and Osmotic Stress: H2O2 Is the Likely Intermediary Signaling Molecule for the Response. Plant J. 2000, 22, 87–95. [Google Scholar] [CrossRef]
- Hossain, M.A.; Mostofa, M.G.; Fujita, M. Cross Protection by Cold-Shock to Salinity and Drought Stress-Induced Oxidative Stress in Mustard (Brassica campestris L.) Seedlings. Mol. Plant Breed. 2013, 4, 50–70. [Google Scholar] [CrossRef]
- Cruz De Carvalho, M.H. Drought Stress and Reactive Oxygen Species: Production, Scavenging and Signaling. Plant Signal Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, S.; Chunduri, V.; Kaur, A.; Kaur, S.; Malhotra, N.; Kumar, A.; Kapoor, P.; Kumari, A.; Kaur, J.; et al. Genome-Wide Identification and Characterization of Heat Shock Protein Family Reveals Role in Development and Stress Conditions in Triticum aestivum L. Sci. Rep. 2020, 10, 7858. [Google Scholar] [CrossRef]
- Most, P.; Papenbrock, J. Possible Roles of Plant Sulfurtransferases in Detoxification of Cyanide, Reactive Oxygen Species, Selected Heavy Metals and Arsenate. Molecules 2015, 20, 1410–1423. [Google Scholar] [CrossRef]
- Padmavathi, T.A.V.; Rao, D.M. Differential Accumulation of Osmolytes in 4 Cultivars of Peanut (Arachis hypogaea L.) under Drought Stress. J. Crop Sci. Biotechnol. 2013, 16, 151–159. [Google Scholar] [CrossRef]
- Bhardwaj, J.; Yadav, S.K. Comparative Study on Biochemical Parameters and Antioxidant Enzymes in a Drought Tolerant and a Sensitive Variety of Horsegram (Macrotyloma uniflorum) under Drought Stress. Am. J. Plant Physiol. 2012, 7, 17–29. [Google Scholar] [CrossRef]
- Mwadzingeni, L.; Shimelis, H.; Tesfay, S.; Tsilo, T.J. Screening of Bread Wheat Genotypes for Drought Tolerance Using Phenotypic and Proline Analyses. Front. Plant Sci. 2016, 7, 1276. [Google Scholar] [CrossRef] [PubMed]
- Giri, J. Glycinebetaine and Abiotic Stress Tolerance in Plants. Plant Signal Behav. 2011, 6, 1746–1751. [Google Scholar] [CrossRef] [PubMed]
- Adrees, M.; Ali, S.; Iqbal, M.; Aslam Bharwana, S.; Siddiqi, Z.; Farid, M.; Ali, Q.; Saeed, R.; Rizwan, M. Mannitol Alleviates Chromium Toxicity in Wheat Plants in Relation to Growth, Yield, Stimulation of Anti-Oxidative Enzymes, Oxidative Stress and Cr Uptake in Sand and Soil Media. Ecotoxicol. Environ. Saf. 2015, 122, 1–8. [Google Scholar] [CrossRef]
- He, Y.F.; Xie, Y.F.; Li, X.; Yang, J. Drought Tolerance of Transgenic Rice Overexpressing Maize C4-Pepc Gene Related to Increased Anthocyanin Synthesis Regulated by Sucrose and Calcium. Biol. Plant 2020, 64, 136–149. [Google Scholar] [CrossRef]
- Zhang, J.; Lei, Y.; Wang, B.; Li, S.; Yu, S.; Wang, Y.; Li, H.; Liu, Y.; Ma, Y.; Dai, H.; et al. The High-Quality Genome of Diploid Strawberry (Fragaria nilgerrensis) Provides New Insights into Anthocyanin Accumulation. Plant Biotechnol. J. 2020, 18, 1908–1924. [Google Scholar] [CrossRef]
- Tarafdar, M.; Bahadur, V.; Rana, S.; Singh, R.K. A Review: Abiotic Stress on Transpiration, Stomatal Diffusive Resistance and Photosynthetic Rate. Pharma Innov. J. 2022, 11, 1632–1635, ISSN (E) 2277-7695. [Google Scholar]
- Tardieu, F.; Simonneau, T.; Muller, B. The Physiological Basis of Drought Tolerance in Crop Plants: A Scenario-Dependent Probabilistic Approach. Annu. Rev. Plant Biol. 2018, 69, 733–759. [Google Scholar] [CrossRef]
- Demidchik, V. ROS-Activated Ion Channels in Plants: Biophysical Characteristics, Physiological Functions and Molecular Nature. Int. J. Mol. Sci. 2018, 19, 1263. [Google Scholar] [CrossRef]
- Batool, S.; Uslu, V.V.; Rajab, H.; Ahmad, N.; Waadt, R.; Geiger, D.; Malagoli, M.; bin Xiang, C.; Hedrich, R.; Rennenberg, H.; et al. Sulfate Is Incorporated into Cysteine to Trigger ABA Production and Stomatal Closure. Plant Cell 2018, 30, 2973–2987. [Google Scholar] [CrossRef]
- Roblero, M.; Pineda, J.; León, C.; Castellanos, J.S. Oxygen in the Root Zone and Its Effect on Plants. Rev. Mex. Cienc. Agric. 2020, 11, 931–943. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. In Sustainable Agriculture; Springer: Dordrech, The Netherlands, 2009; pp. 153–188. [Google Scholar] [CrossRef]
- Schachtman, D.P.; Goodger, J.Q.D. Chemical Root to Shoot Signaling under Drought. Trends Plant Sci. 2008, 13, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Blumwald, E. The Roles of ROS and ABA in Systemic Acquired Acclimation. Plant Cell 2015, 27, 64–70. [Google Scholar] [CrossRef]
- Rameau, C.; Goormachtig, S.; Cardinale, F.; Bennett, T.; Cubas, P. Strigolactones as Plant Hormones. In Strigolactones—Biology and Applications; Springer: Cham, Switzerland, 2019; pp. 47–87. [Google Scholar] [CrossRef]
- Yamada, Y.; Umehara, M. Possible Roles of Strigolactones during Leaf Senescence. Plants 2015, 4, 664–677. [Google Scholar] [CrossRef]
- Visentin, I.; Vitali, M.; Ferrero, M.; Zhang, Y.; Ruyter-Spira, C.; Novák, O.; Strnad, M.; Lovisolo, C.; Schubert, A.; Cardinale, F. Low Levels of Strigolactones in Roots as a Component of the Systemic Signal of Drought Stress in Tomato. New Phytol. 2016, 212, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Min, Z.; Li, R.; Chen, L.; Zhang, Y.; Li, Z.; Liu, M.; Ju, Y.; Fang, Y. Alleviation of Drought Stress in Grapevine by Foliar-Applied Strigolactones. Plant Physiol. Biochem. 2019, 135, 99–110. [Google Scholar] [CrossRef]
- Wang, L.; Wang, B.; Yu, H.; Guo, H.; Lin, T.; Kou, L.; Wang, A.; Shao, N.; Ma, H.; Xiong, G.; et al. Transcriptional Regulation of Strigolactone Signalling in Arabidopsis. Nature 2020, 583, 277–281. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Oxidant and Antioxidant Signalling in Plants: A Re-Evaluation of the Concept of Oxidative Stress in a Physiological Context. Plant Cell Environ. 2005, 28, 1056–1071. [Google Scholar] [CrossRef]
- Jaspers, P.; Kangasjärvi, J. Reactive Oxygen Species in Abiotic Stress Signaling. Physiol. Plant 2010, 138, 405–413. [Google Scholar] [CrossRef]
- Oğuz, M.Ç.; Mujtaba, M.; Yüksel Özmen, C.; Kibar, U.; Kumlay, A.M.; Ergül, A. Expression Analysis of Transcription-Factor Genes Related to Endoplasmic Reticulum Stress Signaling Pathway in Alfalfa (Medicago sativa L.). Acta Physiol. Plant 2022, 44, 37. [Google Scholar] [CrossRef]
- Hartmann, H.; Ziegler, W.; Kolle, O.; Trumbore, S. Thirst Beats Hunger—Declining Hydration during Drought Prevents Carbon Starvation in Norway Spruce Saplings. New Phytol. 2013, 200, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under Drought and Salt Stress: Regulation Mechanisms from Whole Plant to Cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Murata, Y.; Mori, I.C. Stomatal Regulation of Plant Water Status. In Plant Abiotic Stress, 2nd ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; pp. 47–67. [Google Scholar] [CrossRef]
- Rollins, J.A.; Habte, E.; Templer, S.E.; Colby, T.; Schmidt, J.; von Korff, M. Leaf Proteome Alterations in the Context of Physiological and Morphological Responses to Drought and Heat Stress in Barley (Hordeum vulgare L.). J. Exp. Bot. 2013, 64, 3201–3212. [Google Scholar] [CrossRef] [PubMed]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef]
- Bray, E.A. Classification of Genes Differentially Expressed during Water-Deficit Stress in Arabidopsis thaliana: An Analysis Using Microarray and Differential Expression Data. Ann. Bot. 2002, 89, 803–811. [Google Scholar] [CrossRef]
- Mumm, P.; Wolf, T.; Fromm, J.; Roelfsema, M.R.G.; Marten, I. Cell Type-Specific Regulation of Ion Channels within the Maize Stomatal Complex. Plant Cell Physiol. 2011, 52, 1365–1375. [Google Scholar] [CrossRef]
- Potopová, V.; Boroneanţ, C.; Boincean, B.; Soukup, J. Impact of Agricultural Drought on Main Crop Yields in the Republic of Moldova. Int. J. Climatol. 2016, 36, 2063–2082. [Google Scholar] [CrossRef]
- Zhang, Q. Strategies for Developing Green Super Rice. Proc. Natl. Acad. Sci. USA 2007, 104, 16402–16409. [Google Scholar] [CrossRef]
- Muhammad, I.; Shalmani, A.; Ali, M.; Yang, Q.H.; Ahmad, H.; Li, F.B. Mechanisms Regulating the Dynamics of Photosynthesis Under Abiotic Stresses. Front. Plant Sci. 2021, 11, 615942. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Kumar, V.; Kohli, S.K.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Bhardwaj, R.; Zheng, B. Phytohormones Regulate Accumulation of Osmolytes under Abiotic Stress. Biomolecules 2019, 9, 285. [Google Scholar] [CrossRef]
- Sevanto, S. Phloem Transport and Drought. J. Exp. Bot. 2014, 65, 1751–1759. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T.D. Diffusive and Metabolic Limitations to Photosynthesis under Drought and Salinity in C3 Plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Amin Kheradmand, M.; Shahmoradzadeh Fahraji, S.; Fatahi, E.; Mahdi Raoofi, M. Effect of Water Stress on Oil Yield and Some Characteristics of Brassica napus. Int. Res. J. Appl. Basic Sci. 2014, 8, 1447–1453. [Google Scholar]
- Ahanger, M.A.; Morad-Talab, N.; Abd-Allah, E.F.; Ahmad, P.; Hajiboland, R. Plant Growth under Drought Stress: Significance of Mineral Nutrients. In Water Stress and Crop Plants: A Sustainable Approach; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; pp. 649–668. ISBN 978-1-1190-5436-8. [Google Scholar]
- Bhargava, S.; Sawant, K. Drought Stress Adaptation: Metabolic Adjustment and Regulation of Gene Expression. Plant Breed. 2013, 132, 21–32. [Google Scholar] [CrossRef]
- Ying, Y.; Yue, Y.; Huang, X.; Wang, H.; Mei, L.; Yu, W.; Zheng, B.; Wu, J. Salicylic Acid Induces Physiological and Biochemical Changes in Three Red Bayberry (Myric rubra) Genotypes under Water Stress. Plant Growth Regul. 2013, 71, 181–189. [Google Scholar] [CrossRef]
- Rivas, R.; Falcão, H.M.; Ribeiro, R.V.; Machado, E.C.; Pimentel, C.; Santos, M.G. Drought Tolerance in Cowpea Species Is Driven by Less Sensitivity of Leaf Gas Exchange to Water Deficit and Rapid Recovery of Photosynthesis after Rehydration. S. Afr J. Bot. 2016, 103, 101–107. [Google Scholar] [CrossRef]
- Nayyar, H.; Gupta, D. Differential Sensitivity of C3 and C4 Plants to Water Deficit Stress: Association with Oxidative Stress and Antioxidants. Environ. Exp. Bot. 2006, 58, 106–113. [Google Scholar] [CrossRef]
- Georgii, E.; Jin, M.; Zhao, J.; Kanawati, B.; Schmitt-Kopplin, P.; Albert, A.; Winkler, J.B.; Schäffner, A.R. Relationships between Drought, Heat and Air Humidity Responses Revealed by Transcriptome-Metabolome Co-Analysis. BMC Plant Biol. 2017, 17, 120. [Google Scholar] [CrossRef]
- Alghabari, F.; Ihsan, M.Z.; Hussain, S.; Aishia, G.; Daur, I. Effect of Rht Alleles on Wheat Grain Yield and Quality under High Temperature and Drought Stress during Booting and Anthesis. Environ. Sci. Pollut. Res. 2015, 22, 15506–15515. [Google Scholar] [CrossRef]
- Sun, X.L.; Sun, M.; Luo, X.; Ding, X.D.; Ji, W.; Cai, H.; Bai, X.; Liu, X.F.; Zhu, Y.M. A Glycine Soja ABA-Responsive Receptor-like Cytoplasmic Kinase, GsRLCK, Positively Controls Plant Tolerance to Salt and Drought Stresses. Planta 2013, 237, 1527–1545. [Google Scholar] [CrossRef]
- Nikinmaa, E.; Hölttä, T.; Hari, P.; Kolari, P.; Mäkelä, A.; Sevanto, S.; Vesala, T. Assimilate Transport in Phloem Sets Conditions for Leaf Gas Exchange. Plant Cell Environ. 2013, 36, 655–669. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under Stressful Environments: An Overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Sun, Q.; Zybailov, B.; Majeran, W.; Friso, G.; Olinares, P.D.B.; van Wijk, K.J. PPDB, the Plant Proteomics Database at Cornell. Nucleic Acids Res. 2009, 37, D969–D974. [Google Scholar] [CrossRef] [PubMed]
- Faisal, S.; Mujtaba, S.M.; Asma; Mahboob, W. Polyethylene Glycol Mediated Osmotic Stress Impacts on Growth and Biochemical Aspects of Wheat (Triticum aestivum L.). J. Crop Sci. Biotechnol. 2019, 22, 213–223. [Google Scholar] [CrossRef]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of Climate Change on Crops Adaptation and Strategies to Tackle Its Outcome: A Review. Plants 2019, 8, 34. [Google Scholar] [CrossRef]
- Karavidas, I.; Ntatsi, G.; Vougeleka, V.; Karkanis, A.; Ntanasi, T.; Saitanis, C.; Agathokleous, E.; Ropokis, A.; Sabatino, L.; Tran, F.; et al. Agronomic Practices to Increase the Yield and Quality of Common Bean (Phaseolus vulgaris L.): A Systematic Review. Agronomy 2022, 12, 271. [Google Scholar] [CrossRef]
- Abebe, A.; Tsige, A.; Work, M.; Enyew, A. Optimizing Irrigation Frequency and Amount on Yield and Water Productivity of Snap Bean (Phaseolus vulgaris L.) in NW Amhara, Ethiopia: A Case Study in Koga and Ribb Irrigation Scheme. Cogent Food Agric. 2020, 6, 1773690. [Google Scholar] [CrossRef]
- Chojnacka, K.; Moustakas, K.; Witek-Krowiak, A. Bio-Based Fertilizers: A Practical Approach towards Circular Economy. Bioresour. Technol. 2020, 295, 122223. [Google Scholar] [CrossRef]
- Deligios, P.A.; Chergia, A.P.; Sanna, G.; Solinas, S.; Todde, G.; Narvarte, L.; Ledda, L. Climate Change Adaptation and Water Saving by Innovative Irrigation Management Applied on Open Field Globe Artichoke. Sci. Total Environ. 2019, 649, 461–472. [Google Scholar] [CrossRef]
- Marcinkowski, P.; Piniewski, M. Effect of Climate Change on Sowing and Harvest Dates of Spring Barley and Maize in Poland. Int. Agrophys. 2018, 32, 265–271. [Google Scholar] [CrossRef]
- Cooper, M.; van Eeuwijk, F.; Chapman, S.C.; Podlich, D.W.; Löffler, C. Genotype-by environment interactions under water-limited conditions. In Drought Adaptaton in Cereals; CRC Press: New York, NY, USA, 2006; pp. 51–96. ISBN 978-1-5602-2278-1. [Google Scholar]
- Turyagyenda, L.F.; Kizito, E.B.; Ferguson, M.; Baguma, Y.; Agaba, M.; Harvey, J.J.W.; Osiru, D.S.O. Physiological and Molecular Characterization of Drought Responses and Identification of Candidate Tolerance Genes in Cassava. AoB Plants 2013, 5, plt007. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.B.; Chu, L.Y.; Jaleel, C.A.; Manivannan, P.; Panneerselvam, R.; Shao, M.A. Understanding Water Deficit Stress-Induced Changes in the Basic Metabolism of Higher Plants-Biotechnologically and Sustainably Improving Agriculture and the Ecoenvironment in Arid Regions of the Globe. Crit. Rev. Biotechnol. 2009, 29, 131–151. [Google Scholar] [CrossRef] [PubMed]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Al-Juburi, H.J.; Somasundaram, R.; Panneerselvam, R. Drought Stress in Plants: A Review on Morphological Characteristics and Pigments Composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Hussain, M.; Malik, M.A.; Farooq, M.; Ashraf, M.Y.; Cheema, M.A. Improving Drought Tolerance by Exogenous Application of Glycinebetaine and Salicylic Acid in Sunflower. J. Agron. Crop Sci. 2008, 194, 193–199. [Google Scholar] [CrossRef]
- Alves, A.A.C.; Setter, T.L. Response of Cassava Leaf Area Expansion to Water Deficit: Cell Proliferation, Cell Expansion and Delayed Development. Ann. Bot. 2004, 94, 605–613. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive Oxygen Species: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Sankar, B.; Kishorekumar, A.; Gopi, R.; Somasundaram, R.; Panneerselvam, R. Water Deficit Stress Mitigation by Calcium Chloride in Catharanthus Roseus: Effects on Oxidative Stress, Proline Metabolism and Indole Alkaloid Accumulation. Colloids Surf. B Biointerfaces 2007, 60, 110–116. [Google Scholar] [CrossRef]
- Pratap, A.; Douglas, C.; Prajapati, U.; Kumari, G.; War, A.R.; Tomar, R.; Pandey, A.K.; Dubey, S. Breeding Progress and Future Challenges: Biotic Stresses. In The Mungbean Genome; Springer: Cham, Switzerland, 2020; pp. 55–80. [Google Scholar] [CrossRef]
- Shi, J.F.; Mao, X.G.; Jing, R.L.; Pang, X.B.; Wang, Y.G.; Chang, X.P. Gene Expression Profiles of Response to Water Stress at the Jointing Stage in Wheat. Agric. Sci. China 2010, 9, 325–330. [Google Scholar] [CrossRef]
- Harris, D.; Pathan, A.K.; Gothkar, P.; Joshi, A.; Chivasa, W.; Nyamudeza, P. On-Farm Seed Priming: Using Participatory Methods to Revive and Refine a Key Technology. Agric. Syst. 2001, 69, 151–164. [Google Scholar] [CrossRef]
- Abreha, K.B.; Enyew, M.; Carlsson, A.S.; Vetukuri, R.R.; Feyissa, T.; Motlhaodi, T.; Ng’uni, D.; Geleta, M. Sorghum in Dryland: Morphological, Physiological, and Molecular Responses of Sorghum under Drought Stress. Planta 2022, 255, 1–23. [Google Scholar] [CrossRef]
- Liu, M.; Li, M.; Liu, K.; Sui, N. Effects of Drought Stress on Seed Germination and Seedling Growth of Different Maize Varieties. J. Agric. Sci. 2015, 7, 231. [Google Scholar] [CrossRef]
- Kaya, M.D.; Okçu, G.; Atak, M.; Çikili, Y.; Kolsarici, Ö. Seed Treatments to Overcome Salt and Drought Stress during Germination in Sunflower (Helianthus annuus L.). Eur. J. Agron. 2006, 24, 291–295. [Google Scholar] [CrossRef]
- Okçu, G.; Kaya, M.D.; Atak, M. Effects of Salt and Drought Stresses on Germination and Seedling Growth of Pea (Pisum sativum L.). Turk. J. Agric. For. 2005, 29, 237–242. [Google Scholar] [CrossRef]
- Tawfik, K.M. Effect of Water Stress in Addition to Potassiomag Application on Mungbean. Aust. J. Basic Appl. Sci. 2008, 2, 42–52. [Google Scholar]
- Chaniago, I.; Syarif, A.; Riviona, P. Sorghum Seedling Drought Response: In Search of Tolerant Genotypes. Int. J. Adv Sci. Eng. Inf. Technol. 2017, 7, 892–897. [Google Scholar] [CrossRef]
- Ali, Q.; Javed, M.T.; Noman, A.; Haider, M.Z.; Waseem, M.; Iqbal, N.; Waseem, M.; Shah, M.S.; Shahzad, F.; Perveen, R. Assessment of Drought Tolerance in Mung Bean Cultivars/Lines as Depicted by the Activities of Germination Enzymes, Seedling’s Antioxidative Potential and Nutrient Acquisition. Arch. Agron. Soil Sci. 2018, 64, 84–102. [Google Scholar] [CrossRef]
- Li, L.; Mao, X.; Wang, J.; Chang, X.; Reynolds, M.; Jing, R. Genetic Dissection of Drought and Heat-Responsive Agronomic Traits in Wheat. Plant Cell Environ. 2019, 42, 2540–2553. [Google Scholar] [CrossRef]
- Gano, B.; Dembele, J.S.B.; Tovignan, T.K.; Sine, B.; Vadez, V.; Diouf, D.; Audebert, A. Article Adaptation Responses to Early Drought Stress of West Africa Sorghum Varieties. Agronomy 2021, 11, 443. [Google Scholar] [CrossRef]
- Rana, M.S.; Hasan, M.A.; Bahadur, M.M.; Islam, M.R. Effect of polyethylene glycol induced water stress on germination and seedling growth of wheat (Triticum aestivum). Agriculture 2017, 15, 81–91. [Google Scholar] [CrossRef][Green Version]
- Fadoul, H.E.; el Siddig, M.A.; Abdalla, A.W.H.; el Hussein, A.A. Physiological and Proteomic Analysis of Two Contrasting Sorghum bicolor Genotypes in Response to Drought Stress. Aust. J. Crop Sci. 2018, 12, 1543–1551. [Google Scholar] [CrossRef]
- Wang, C.; Han, B. Twenty Years of Rice Genomics Research: From Sequencing and Functional Genomics to Quantitative Genomics. Mol. Plant 2022, 15, 593–619. [Google Scholar] [CrossRef]
- Lopes, M.S.; Reynolds, M.P. Partitioning of Assimilates to Deeper Roots Is Associated with Cooler Canopies and Increased Yield under Drought in Wheat. Funct. Plant Biol. 2010, 37, 147–156. [Google Scholar] [CrossRef]
- Abbate, P.E.; Dardanelli, J.L.; Cantarero, M.G.; Maturano, M.; Melchiori, R.J.M.; Suero, E.E. Climatic and Water Availability Effects on Water-Use Efficiency in Wheat. Crop Sci. 2004, 44, 474–483. [Google Scholar] [CrossRef]
- Monclus, R.; Dreyer, E.; Villar, M.; Delmotte, F.M.; Delay, D.; Petit, J.M.; Barbaroux, C.; le Thiec, D.; Bréchet, C.; Brignolas, F. Impact of Drought on Productivity and Water Use Efficiency in 29 Genotypes of Populus Deltoides x Populus Nigra. New Phytol. 2006, 169, 765–777. [Google Scholar] [CrossRef]
- Mega, R.; Abe, F.; Kim, J.S.; Tsuboi, Y.; Tanaka, K.; Kobayashi, H.; Sakata, Y.; Hanada, K.; Tsujimoto, H.; Kikuchi, J.; et al. Tuning Water-Use Efficiency and Drought Tolerance in Wheat Using Abscisic Acid Receptors. Nat. Plants 2019, 5, 153–159. [Google Scholar] [CrossRef]
- Rucker, K.S.; Kvien, C.K.; Holbrook, C.C.; Hook, J.E. Identification of Peanut Genotypes with Improved Drought Avoidance Traits1. Peanut Sci. 1995, 22, 14–18. [Google Scholar] [CrossRef]
- Shao, H.B.; Chu, L.Y.; Jaleel, C.A.; Zhao, C.X. Water-Deficit Stress-Induced Anatomical Changes in Higher Plants. Comptes Rendus Biol. 2008, 331, 215–225. [Google Scholar] [CrossRef]
- Zhang, J.; Jia, W.; Yang, J.; Ismail, A.M. Role of ABA in Integrating Plant Responses to Drought and Salt Stresses. Field Crop. Res. 2006, 97, 111–119. [Google Scholar] [CrossRef]
- Rizza, F.; Badeck, F.W.; Cattivelli, L.; Lidestri, O.; di Fonzo, N.; Stanca, A.M. Use of a Water Stress Index to Identify Barley Genotypes Adapted to Rainfed and Irrigated Conditions. Crop Sci. 2004, 44, 2127–2137. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Alegre, L. Die and Let Live: Leaf Senescence Contributes to Plant Survival under Drought Stress. Funct. Plant Biol. 2004, 31, 203–216. [Google Scholar] [CrossRef]
- Zhang, G.; Hou, X.; Wang, L.; Xu, J.; Chen, J.; Fu, X.; Shen, N.; Nian, J.; Jiang, Z.; Hu, J.; et al. PHOTO-SENSITIVE LEAF ROLLING 1 Encodes a Polygalacturonase That Modifies Cell Wall Structure and Drought Tolerance in Rice. New Phytol. 2021, 229, 890–901. [Google Scholar] [CrossRef]
- Fu, J.; Huang, B. Involvement of Antioxidants and Lipid Peroxidation in the Adaptation of Two Cool-Season Grasses to Localized Drought Stress. Environ. Exp. Bot. 2001, 45, 105–114. [Google Scholar] [CrossRef]
- Anjum, A.S.; Zada, R.; Tareen, W.H. Organic Farming: Hope for the Sustainable Livelihoods of Future Generations in Pakistan. J. Rural Dev. Agric. 2016, 1, 20–29. [Google Scholar]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and Heat-Stress Effects on Seed Filling in Food Crops: Impacts on Functional Biochemistry, Seed Yields, and Nutritional Quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef]
- Jabran, K.; Ullah, E.; Akbar, N.; Yasin, M.; Zaman, U.; Nasim, W.; Riaz, M.; Arjumend, T.; Azhar, M.F.; Hussain, M. Growth and Physiology of Basmati Rice under Conventional and Water-Saving Production Systems. Arch. Agron. Soil Sci. 2017, 63, 1465–1476. [Google Scholar] [CrossRef]
- Manickavelu, A.; Nadarajan, N.; Ganesh, S.K.; Gnanamalar, R.P.; Chandra Babu, R. Drought Tolerance in Rice: Morphological and Molecular Genetic Consideration. Plant Growth Regul. 2006, 50, 121–138. [Google Scholar] [CrossRef]
- Awais, M.; Wajid, A.; Bashir, M.U.; Habib-ur-Rahman, M.; Raza, M.A.S.; Ahmad, A.; Saleem, M.F.; Hammad, H.M.; Mubeen, M.; Saeed, U.; et al. Nitrogen and Plant Population Change Radiation Capture and Utilization Capacity of Sunflower in Semi-Arid Environment. Environ. Sci. Pollut. Res. 2017, 24, 17511–17525. [Google Scholar] [CrossRef]
- Nasim, W.; Ahmad, A.; Belhouchette, H.; Fahad, S.; Hoogenboom, G. Evaluation of the OILCROP-SUN Model for Sunflower Hybrids under Different Agro-Meteorological Conditions of Punjab-Pakistan. Field Crop. Res. 2016, 188, 17–30. [Google Scholar] [CrossRef]
- Selvakumar, K.; Bavithra, S.; Suganthi, M.; Benson, C.S.; Elumalai, P.; Arunkumar, R.; Krishnamoorthy, G.; Venkataraman, P.; Arunakaran, J. Protective Role of Quercetin on PCBs-Induced Oxidative Stress and Apoptosis in Hippocampus of Adult Rats. Neurochem. Res. 2012, 37, 708–721. [Google Scholar] [CrossRef]
- Zlatev, Z.; Lidon, F.C. An Overview on Drought Induced Changes in Plant Growth, Water Relations and Photosynthesis. Emir J. Food Agric. 2012, 24, 57–72. [Google Scholar] [CrossRef]
- Semercı, A.; Çıçek, N.; Karahan, F.A.; Özyürek, E.; Kaşko Arici, Y.; Ekmekçı, Y. Some Growth and Chlorophyll Fluorescence Parameters of Black and Hybrid Poplar Clones under Water Stress. Turk. J. Agric. For. 2017, 41, 348–356. [Google Scholar] [CrossRef]
- Panda, R.K.; Pandit, E.; Swain, A.; Mohanty, D.; Baig, M.J.; Kar, M.; Pradhan, S.K. Response of Physiological and Biochemical Parameters in Deeper Rooting Rice Genotypes under Irrigated and Water Stress Conditions. ORYZA—Int. J. Rice 2016, 53, 422–427. [Google Scholar]
- Hossain, M.A.; Fujita, M. Evidence for a Role of Exogenous Glycinebetaine and Proline in Antioxidant Defense and Methylglyoxal Detoxification Systems in Mung Bean Seedlings under Salt Stress. Physiol. Mol. Biol. Plants 2010, 16, 19–29. [Google Scholar] [CrossRef]
- de Souza, A.P.; Cocuron, J.C.; Garcia, A.C.; Alonso, A.P.; Buckeridge, M.S. Changes in Whole-Plant Metabolism during the Grain-Filling Stage in Sorghum Grown under Elevated CO2 and Drought. Plant Physiol. 2015, 169, 1755–1765. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Staggenborg, S.A.; Ristic, Z. Impacts of Drought and/or Heat Stress on Physiological, Developmental, Growth, and Yield Processes of Crop Plants. In Response of Crops to Limited Water: Understanding and Modeling Water Stress Effects on Plant Growth Processes; American Society of Agronomy: Madison, WI, USA, 2008; pp. 301–355. [Google Scholar] [CrossRef]
- Murtaza, G.; Rasool, F.; Habib, R.; Javed, T.; Sardar, K.; Mohsin, M.; Ayub, M.A.; Rasool, A. A Review of Morphological, Physiological and Biochemical Responses of Plants under Drought Stress Conditions. Imp. J. Interdiscip. Res. 2016, 2, 1600–1606. [Google Scholar]
- Yadav, R.S.; Hash, C.T.; Bidinger, F.R.; Devos, K.M.; Howarth, C.J. Genomic Regions Associated with Grain Yield and Aspects of Post-Flowering Drought Tolerance in Pearl Millet across Stress Environments and Tester Background. Euphytica 2004, 136, 265–277. [Google Scholar] [CrossRef]
- Basu, P.S.; Pratap, A.; Gupta, S.; Sharma, K.; Tomar, R.; Singh, N.P. Physiological Traits for Shortening Crop Duration and Improving Productivity of Greengram (Vigna radiata L. Wilczek) Under High Temperature. Front. Plant Sci. 2019, 10, 1508. [Google Scholar] [CrossRef]
- Ranawake, A.; Dahanayaka, N.; Amarasingha, U.; Rodrigo, W.; Rodrigo, U. Effect of Water Stress on Growth and Yield of Mung Bean (Vigna radiata L.). Trop. Agric. Res. Ext. 2012, 14, 76–79. [Google Scholar] [CrossRef]
- Vadez, V.; Krishnamurthy, L.; Hash, C.T.; Upadhyaya, H.D.; Borrell, A.K. Yield, Transpiration Efficiency, and Water-Use Variations and Their Interrelationships in the Sorghum Reference Collection. Crop Pasture Sci. 2011, 62, 645–655. [Google Scholar] [CrossRef]
- Lopez, J.R.; Erickson, J.E.; Munoz, P.; Saballos, A.; Felderhoff, T.J.; Vermerris, W. QTLs Associated with Crown Root Angle, Stomatal Conductance, and Maturity in Sorghum. Plant Genome 2017, 10, plantgenome2016.04.0038. [Google Scholar] [CrossRef]
- Rama Reddy, N.R.; Ragimasalawada, M.; Sabbavarapu, M.M.; Nadoor, S.; Patil, J.V. Detection and Validation of Stay-Green QTL in Post-Rainy Sorghum Involving Widely Adapted Cultivar, M35-1 and a Popular Stay-Green Genotype B35. BMC Genom. 2014, 15, 909. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, H.; Rad, A.H.S.; Moghadam, H.R.T. Effects of Zeolite and Selenium Applications on Some Agronomic Traits of Three Canola Cultivars under Drought Stress. Pesqui Agropecu Trop 2011, 41, 179–185. [Google Scholar] [CrossRef][Green Version]
- Ndlovu, E.; van Staden, J.; Maphosa, M. Morpho-Physiological Effects of Moisture, Heat and Combined Stresses on Sorghum bicolor [Moench (L.)] and Its Acclimation Mechanisms. Plant Stress 2021, 2, 100018. [Google Scholar] [CrossRef]
- Pervez, M.A.; Ayub, C.M.; Khan, H.A.; Shahid, M.A.; Ashraf, I. Effect of Drought Stress on Growth, Yield and Seed Quality of Tomato ( Lycopersicon esculentum L.). Pak. J. Agri. Sci. 2009, 46, 174–178. [Google Scholar]
- Lopes, M.S.; El-Basyoni, I.; Baenziger, P.S.; Singh, S.; Royo, C.; Ozbek, K.; Aktas, H.; Ozer, E.; Ozdemir, F.; Manickavelu, A.; et al. Exploiting Genetic Diversity from Landraces in Wheat Breeding for Adaptation to Climate Change. J. Exp. Bot. 2015, 66, 3477–3486. [Google Scholar] [CrossRef]
- Sabella, E.; Aprile, A.; Negro, C.; Nicolì, F.; Nutricati, E.; Vergine, M.; Luvisi, A.; de Bellis, L. Impact of Climate Change on Durum Wheat Yield. Agronomy 2020, 10, 793. [Google Scholar] [CrossRef]
- Ait-El-Mokhtar, M.; ben Laouane, R.; Anli, M.; Boutasknit, A.; Wahbi, S.; Meddich, A. Use of Mycorrhizal Fungi in Improving Tolerance of the Date Palm (Phoenix dactylifera L.) Seedlings to Salt Stress. Sci. Hortic. 2019, 253, 429–438. [Google Scholar] [CrossRef]
- Meddich, A.; Oufdou, K.; Boutasknit, A.; Raklami, A.; Tahiri, A.; Ben-Laouane, R.; Ait-El-Mokhtar, M.; Anli, M.; Mitsui, T.; Wahbi, S.; et al. Use of Organic and Biological Fertilizers as Strategies to Improve Crop Biomass, Yields and Physicochemical Parameters of Soil. In Nutrient Dynamics for Sustainable Crop Production; Springer: Singapore, 2019; pp. 247–288. [Google Scholar] [CrossRef]
- Boutasknit, A.; Baslam, M.; Ait-El-mokhtar, M.; Anli, M.; Ben-Laouane, R.; Douira, A.; el Modafar, C.; Mitsui, T.; Wahbi, S.; Meddich, A. Arbuscular Mycorrhizal Fungi Mediate Drought Tolerance and Recovery in Two Contrasting Carob (Ceratonia siliqua L.) Ecotypes by Regulating Stomatal, Water Relations, and (in)Organic Adjustments. Plants 2020, 9, 80. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).