
Generalization Increases the Adaptive Value of Mate Choice Copying When Immediate Copying Is Costly


Abstract
1. Introduction
2. Models
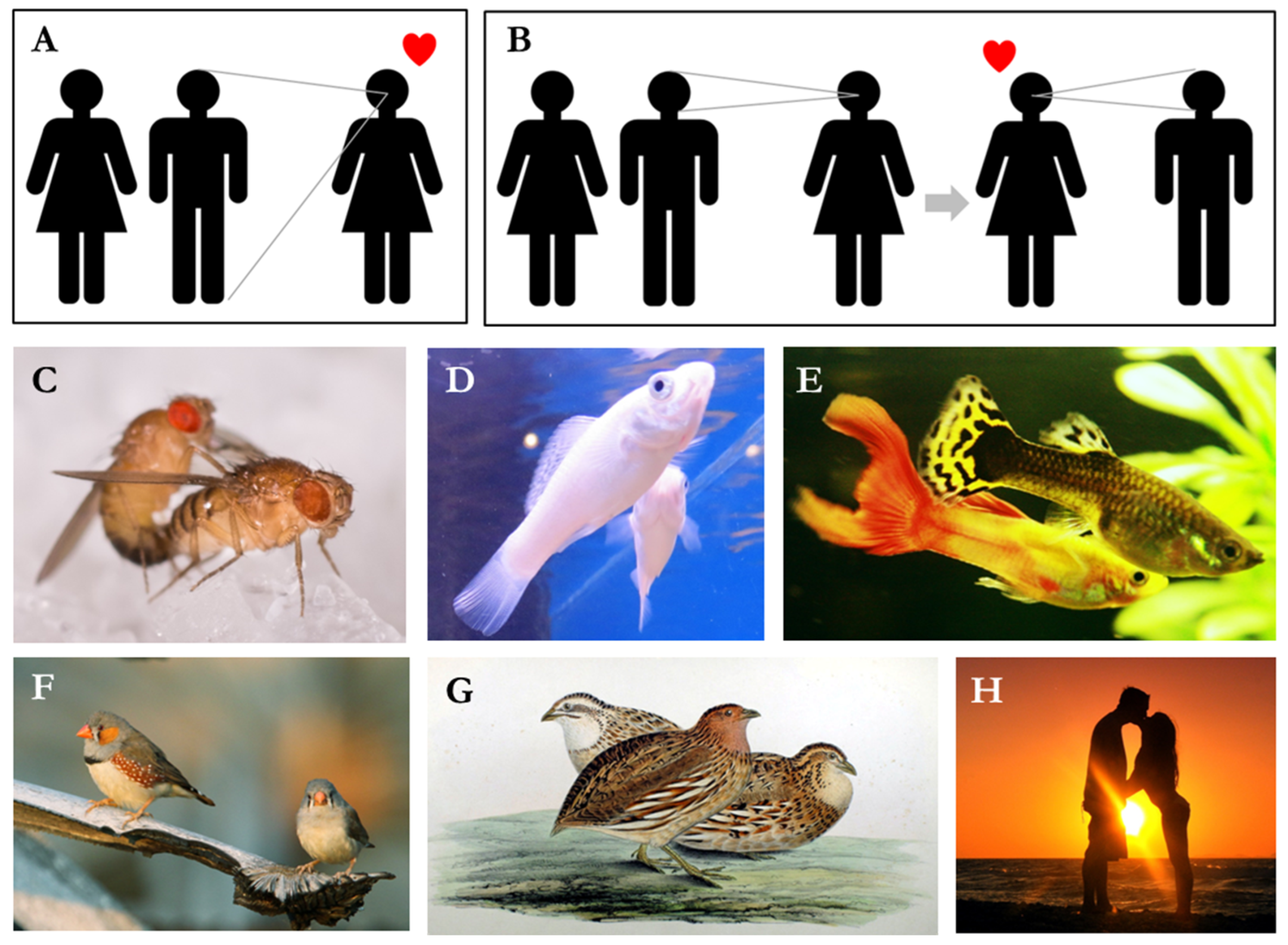
2.1. Original Model
2.2. Extension 1: Generalization
2.3. Extension 2: Costs to Immediate Copying
2.4. Extension 3: Incorporating Both Extensions
3. Discussion
Funding
Data Availability Statement
Conflicts of Interest
References
- Westneat, D.F.; Walters, A.; McCarthy, T.M.; Hatch, M.I.; Hein, W.K. Alternative mechanisms of nonindependent mate choice. Anim. Behav. 2000, 59, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Hoglund, J.; Alatalo, R.V.; Lundberg, A. Copying the mate choice of others? observations on the female black grouse. Behaviour 1990, 114, 221–231. [Google Scholar] [CrossRef]
- Dugatkin, L.A. Sexual selection and imitation: Females copy the mate choice of others. Am. Nat. 1992, 139, 1384–1389. [Google Scholar] [CrossRef]
- Vakirtzis, A. Mate choice copying and nonindependent mate choice: A critical review. Ann. Zool. Fenn. 2011, 48, 91–107. [Google Scholar] [CrossRef]
- Gouda-Vossos, A.; Nakagawa, S.; Dixson, B.J.W.; Brooks, R.C. Mate Choice Copying in Humans: A Systematic Review and Meta-Analysis. Adapt. Hum. Behav. Physiol. 2018, 4, 364–386. [Google Scholar] [CrossRef]
- Wade, M.J.; Pruett-Jones, S.G. Female copying increases the variance in male mating success. Proc. Nat. Acad. Sci. USA 1990, 87, 5749–5753. [Google Scholar] [CrossRef]
- Brooks, R. The importance of mate copying and cultural inheritance of mating preferences. Trends Ecol. Evol. 1998, 13, 45–46. [Google Scholar] [CrossRef]
- Little, A.C.; Jones, B.C.; DeBruine, L.M.; Caldwell, C.A. Social learning and human mate preferences: A potential mechanism for generating and maintaining between-population diversity in attraction. Phil Trans. R. Soc. B 2011, 366, 366–375. [Google Scholar] [CrossRef]
- Nöbel, S.; Jacquet, A.; Isabel, G.; Pocheville, A.; Seabright, P.; Danchin, E. Conformity in mate choice, the overlooked social component of animal and human culture. Biol. Rev. 2023, 98, 132–149. [Google Scholar] [CrossRef]
- Bowers, R.I.; Place, S.S.; Todd, P.M.; Penke, L.; Asendorpf, J.B. Generalization in mate-choice copying in humans. Behav. Ecol. 2012, 23, 112–124. [Google Scholar] [CrossRef]
- Danchin, E.; Giraldeau, L.-A.; Valone, T.J.; Wagner, R.H. Public information: From nosy neighbors to cultural evolution. Science 2004, 305, 487–491. [Google Scholar] [CrossRef] [PubMed]
- White, D.J.; Galef, B.G., Jr. ‘Culture’ in quail: Social influences on mate choices of female Coturnix japonica. Anim. Behav. 2000, 59, 975–979. [Google Scholar] [CrossRef] [PubMed]
- Godin, J.-G.J.; Herdman, E.J.E.; Dugatkin, L.A. Social influences on female mate choice in the guppy, Poecilia reticulata: Generalized and repeatable trait-copying behaviour. Anim. Behav. 2005, 69, 999–1005. [Google Scholar] [CrossRef]
- Brown, G.R.; Fawcett, T.W. Sexual selection: Copycat mating in birds. Curr. Biol. 2005, 15, R626–R628. [Google Scholar] [CrossRef] [PubMed]
- Witte, K.; Noltemeier, B. The role of information in mate-choice copying in female sailfin mollies (Poecilia latipinna). Behav. Ecol. Sociobiol. 2002, 52, 194–202. [Google Scholar] [CrossRef]
- Swaddle, J.P.; Cathey, M.G.; Correll, M.; Hodkinson, B.P. Socially transmitted mate preferences in a monogamous bird: A non-genetic mechanism of sexual selection. Proc. R. Soc. B Biol. Sci. 2005, 272, 1053–1058. [Google Scholar] [CrossRef]
- Mery, F.; Varela, S.A.M.; Danchin, É.; Blanchet, S.; Parejo, D.; Coolen, I.; Wagner, R.H. Public versus personal information for mate copying in an invertebrate. Curr. Biol. 2009, 19, 730–734. [Google Scholar] [CrossRef]
- Danchin, E.; Nöbel, S.; Pocheville, A.; Dagaeff, A.-C.; Demay, L.; Alphand, M.; Ranty-Roby, S.; van Renssen, L.; Monier, M.; Gazagne, E.; et al. Cultural flies: Conformist social learning in fruitflies predicts long-lasting mate-choice traditions. Science 2018, 362, 1025–1030. [Google Scholar] [CrossRef]
- Pruett-Jones, S. Independent versus nonindependent mate choice: Do females copy each other? Am. Nat. 1992, 140, 1000–1009. [Google Scholar] [CrossRef]
- Maynard Smith, J. Evolution and the Theory of Games; Princeton University Press: Princeton, NJ, USA, 1982. [Google Scholar]
- Maynard Smith, J.; Price, G.R. The logic of animal conflict. Nature 1973, 246, 15–18. [Google Scholar] [CrossRef]
- Barrett, H.C. The Shape of Thought: How Mental Adaptations Evolve; Oxford University Press: New York, NY, USA, 2015. [Google Scholar]
- Galef, B.G.; White, D.J. Evidence of social effects on mate choice in vertebrates. Behav. Process. 2000, 51, 167–175. [Google Scholar] [CrossRef] [PubMed]
- White, D.J. Influences of social learning on mate-choice decisions. Learn. Behav. 2004, 32, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Vakirtzis, A.; Roberts, S.C. Nonindependent mate choice in monogamy. Behav. Ecol. 2010, 21, 898–901. [Google Scholar] [CrossRef]
- Dubois, F. Mate choice copying in a monogamous species: Should females use public information to choose extrapair males? Anim. Behav. 2007, 74, 1785–1793. [Google Scholar] [CrossRef]
- Agrawal, A.F. The evolutionary consequences of mate choice copying on male traits. Behav. Ecol. Sociobiol. 2001, 51, 33–40. [Google Scholar] [CrossRef]
- Verzijden, M.N.; ten Cate, C.; Servedio, M.R.; Kozak, G.M.; Boughman, J.W.; Svensson, E.I. The impact of learning on sexual selection and speciation. Trends Ecol. Evol. 2012, 27, 511–519. [Google Scholar] [CrossRef]
- Gibson, R.M.; Höglund, J. Copying and sexual selection. Trends Ecol. Evol. 1992, 7, 229–232. [Google Scholar] [CrossRef]
- Kirkpatrick, M.; Dugatkin, L.A. Sexual selection and the evolutionary effects of copying mate choice. Behav. Ecol. Sociobiol. 1994, 34, 443–449. [Google Scholar] [CrossRef]
- Bailey, N.W.; Moore, A.J. Runaway sexual selection without genetic correlations: Social environments and flexible mate choice initiate and enhance the Fisher process. Evolution 2012, 66, 2674–2684. [Google Scholar] [CrossRef]
- Richerson, P.J.; Boyd, R. The role of evolved predispositions in cultural evolution: Or, Human Sociobiology meets Pascal’s Wager. Ethol. Sociobiol. 1989, 10, 195–219. [Google Scholar] [CrossRef]
- Laland, K.N. Sexual selection with a culturally transmitted mating preference. Theor. Popul. Biol. 1994, 45, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M. An evolutionarily stable strategy may be inaccessible. J. Theor. Biol. 1990, 142, 237–241. [Google Scholar] [CrossRef] [PubMed]
- McNamara, J.M.; Weissing, F.J. Evolutionary game theory. In Social Behaviour: Genes, Ecology and Evolution; Székely, T., Moore, A.J., Komdeur, J., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 88–106. [Google Scholar]
- Huttegger, S.M.; Zollman, K.J.S. Methodology in biological game theory. Br. J. Philos. Sci. 2013, 64, 637–658. [Google Scholar] [CrossRef]
- Taylor, P.D.; Jonker, L.B. Evolutionary stable strategies and game dynamics. Math. Biosci. 1978, 40, 145–156. [Google Scholar] [CrossRef]
- Kokko, H. Modeling for Field Biologists and Other Interesting People; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- DuVal, E.H.; Fitzpatrick, C.L.; Hobson, E.A.; Servedio, M.R. Inferred attractiveness: A generalized mechanism for sexual selection that can maintain variation in traits and preferences over time. PLOS Biol. 2023, 21, e3002269. [Google Scholar] [CrossRef]
- Levins, R. The strategy of model building in population biology. Am. Sci. 1966, 54, 421–431. [Google Scholar]
- Darwin, C. The Descent of Man and Sex in Relation to Sex; John Murray: London, UK, 1871. [Google Scholar]
- Ford, C.S.; Beach, F.A. Patterns of Sexual Behavior; Harper: New York, NY, USA, 1951. [Google Scholar]
- Dugatkin, L.A. The interface between culturally based preferences and genetic mate preferences: Female mate choice in Pocilia reticulata. Proc. Nat. Acad. Sci. USA 1996, 93, 2770–2773. [Google Scholar] [CrossRef]
- Santos, M.; Varela, S.A.M. Genetic and Cultural Evolution of Mate Choice. In Illuminating Human Evolution: 150 Years After Darwin; Bertranpetit, J., Peretó, J., Eds.; Springer Nature: Singapore, 2022; pp. 187–199. [Google Scholar]
- Street, S.E.; Morgan, T.J.H.; Thornton, A.; Brown, G.R.; Laland, K.N.; Cross, C.P. Human mate-choice copying is domain-general social learning. Sci. Rep. 2018, 8, 1715. [Google Scholar] [CrossRef]
- Dougherty, L.R. Designing mate choice experiments. Biol. Rev. 2020, 95, 759–781. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| When Paired with: | |||
|---|---|---|---|
| Chooser | Copier | ||
| Payoff to: | Chooser | W + f − k | W + f − k |
| Copier | W + f | W | |
| When Paired with: | |||||
|---|---|---|---|---|---|
| Chooser | Copier | ||||
| Immediate | Future | Immediate | Future | ||
| Payoff to: | Chooser | W + f − k | W + f − k | W + f − k | W + f − k |
| Copier | W + f | m (W + f) + (1 − m) W | W | W | |
| When Paired with: | |||
|---|---|---|---|
| Chooser | Copier | ||
| Payoff to: | Chooser | W + f − k | W + f − k |
| Copier | W + f − c | W − c | |
| When Paired with: | |||||
|---|---|---|---|---|---|
| Chooser | Copier | ||||
| Immediate | Future | Immediate | Future | ||
| Payoff to: | Chooser | W + f − k | W + f − k | W + f − k | W + f − k |
| Copier | W + f − c | m (W + f) + (1 − m) W | W − c | W | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kushnick, G. Generalization Increases the Adaptive Value of Mate Choice Copying When Immediate Copying Is Costly. Humans 2024, 4, 340-350. https://doi.org/10.3390/humans4040022
Kushnick G. Generalization Increases the Adaptive Value of Mate Choice Copying When Immediate Copying Is Costly. Humans. 2024; 4(4):340-350. https://doi.org/10.3390/humans4040022
Chicago/Turabian StyleKushnick, Geoff. 2024. "Generalization Increases the Adaptive Value of Mate Choice Copying When Immediate Copying Is Costly" Humans 4, no. 4: 340-350. https://doi.org/10.3390/humans4040022
APA StyleKushnick, G. (2024). Generalization Increases the Adaptive Value of Mate Choice Copying When Immediate Copying Is Costly. Humans, 4(4), 340-350. https://doi.org/10.3390/humans4040022