
Optimizing Early Growth of Laminaria hyperborea in Controlled Settings: A Pathway to Improved Restoration Efforts

 , , , , , and
, , , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Fieldwork and Laboratory Procedure
2.2. Data Analysis
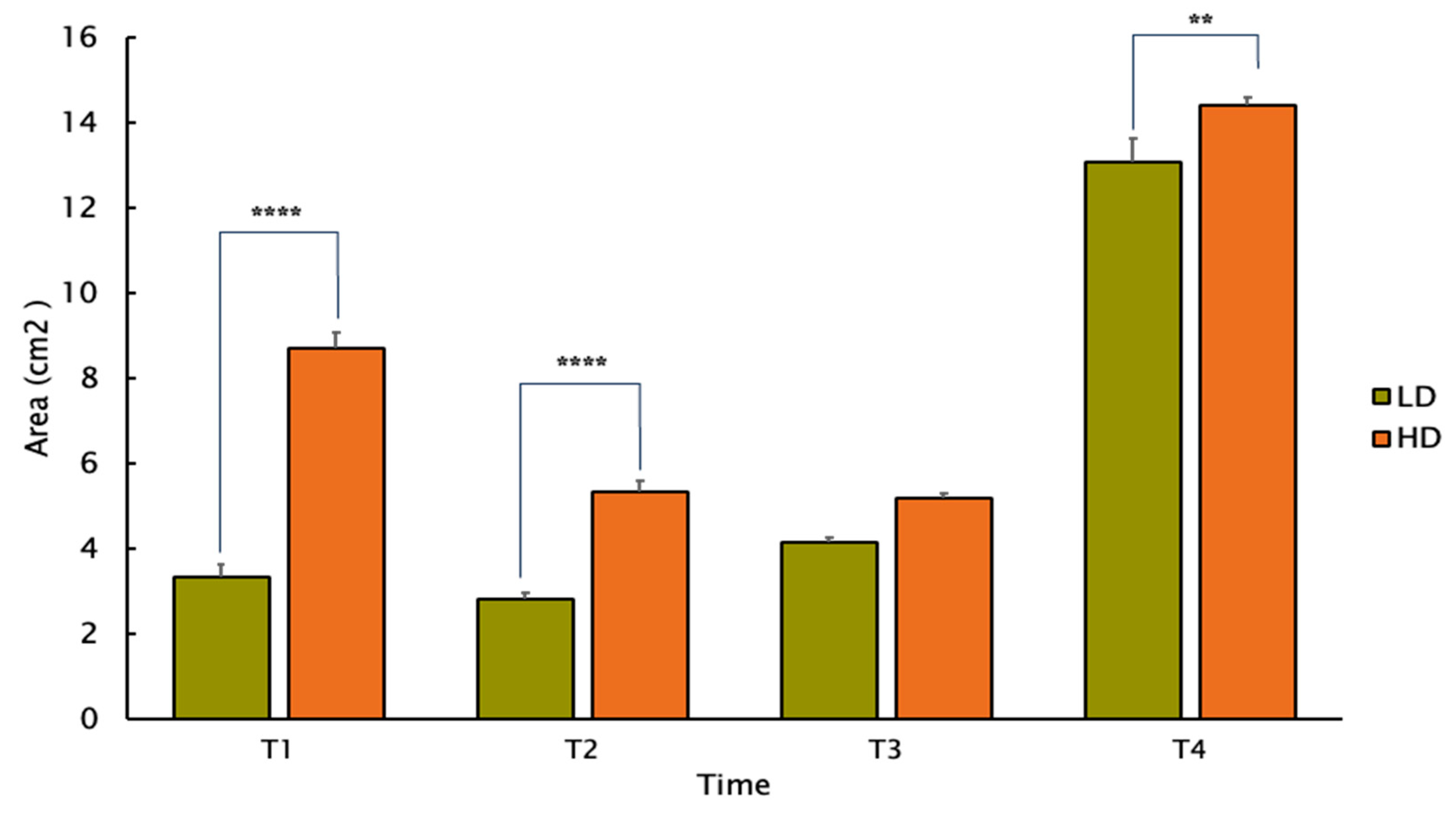
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bourque, B.J.; Corbett, D.; Erlandson, J.M.; Estes, J.A.; Graham, M.H.; Steneck, R.S.; Tegner, M.J. Kelp forest ecosystems: Biodiversity, stability, resilience and future. Environ. Conserv. 2002, 29, 436–459. [Google Scholar] [CrossRef]
- Wernberg, T.; Thomsen, M.S.; Tuya, F.; Kendrick, G.A.; Staehr, P.A.; Toohey, B.D. Decreasing resilience of kelp beds along a latitudinal temperature gradient: Potential implications for a warmer future. Ecol. Lett. 2010, 13, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Araújo, R.M.; Assis, J.; Aguillar, R.; Airoldi, L.; Bárbara, I.; Bartsch, I.; Bekkby, T.; Christie, H.; Davoult, D.; Derrien-Courtel, S.; et al. Status, trends and drivers of kelp forests in Europe: An expert assessment. Biodivers. Conserv. 2016, 25, 1319–1348. [Google Scholar] [CrossRef]
- Verdura, J.; Sales, M.; Ballesteros, E.; Cefalì, M.E.; Cebrian, E. Restoration of a Canopy-Forming Alga Based on Recruitment Enhancement: Methods and Long-Term Success Assessment. Front. Plant Sci. 2018, 9, 1832. [Google Scholar] [CrossRef]
- Araujo, R.M.; Bartsch, I.; Bekkby, T.; Erzini, K.; Sousa-Pinto, I. What is the impact of kelp forest density and/or area on fisheries? Environ. Evid. 2013, 2, 15. [Google Scholar] [CrossRef]
- Turner, R.K.; Burgess, D.; Hadley, D.; Coombes, E.; Jackson, N. A cost–benefit appraisal of coastal managed realignment policy. Glob. Environ. Chang. 2007, 17, 397–407. [Google Scholar] [CrossRef]
- Temmerman, S.; Meire, P.; Bouma, T.J.; Herman, P.M.J.; Ysebaert, T.; De Vriend, H.J. Ecosystem-based coastal defence in the face of global change. Nature 2013, 504, 79–83. [Google Scholar] [CrossRef]
- Belkin, I.M. Rapid warming of large marine ecosystems. Prog. Oceanogr. 2009, 81, 207–213. [Google Scholar] [CrossRef]
- Casado-Amezúa, P.; Araújo, R.; Bárbara, I.; Bermejo, R.; Borja, Á.; Díez, I.; Fernández, C.; Gorostiaga, J.M.; Guinda, X.; Hernández, I.; et al. Distributional shifts of canopy-forming seaweeds from the Atlantic coast of Southern Europe. Biodivers. Conserv. 2019, 28, 1151–1172. [Google Scholar] [CrossRef]
- Pinho, D.; Bertocci, I.; Arenas, F.; Franco, J.N.; Jacinto, D.; Castro, J.J.; Vieira, R.; Sousa-Pinto, I.; Wernberg, T.; Tuya, F. Spatial and temporal variation of kelp forests and associated macroalgal assemblages along the Portuguese coast. Mar. Freshw. Res. 2015, 67, 113–122. [Google Scholar] [CrossRef]
- Fernández, C. The retreat of large brown seaweeds on the north coast of Spain: The case of Saccorhiza polyschides. Eur. J. Phycol. 2011, 46, 352–360. [Google Scholar] [CrossRef]
- Tanaka, K.; Taino, S.; Haraguchi, H.; Prendergast, G.; Hiraoka, M. Warming off southwestern Japan linked to distributional shifts of subtidal canopy-forming seaweeds. Ecol. Evol. 2012, 2, 2854–2865. [Google Scholar] [CrossRef] [PubMed]
- Wernberg, T.; de Bettignies, T.; Joy, B.A.; Finnegan, P.M. Physiological responses of habitat-forming seaweeds to increasing temperatures. Limnol. Oceanogr. 2016, 61, 2180–2190. [Google Scholar] [CrossRef]
- Smale, D.A.; Burrows, M.T.; Moore, P.; O’Connor, N.; Hawkins, S.J. Threats and knowledge gaps for ecosystem services provided by kelp forests: A northeast Atlantic perspective. Ecol. Evol. 2013, 3, 4016–4038. [Google Scholar] [CrossRef]
- Assis, J.; Lucas, A.V.; Bárbara, I.; Serrão, E.Á. Future climate change is predicted to shift long-term persistence zones in the cold-temperate kelp Laminaria hyperborea. Mar. Environ. Res. 2016, 113, 174–182. [Google Scholar] [CrossRef]
- García, L.M.; Rancel-Rodríguez, N.M.; Sangil, C.; Reyes, J.; Benito, B.; Orellana, S.; Sansón, M. Environmental and human factors drive the subtropical marine forests of Gongolaria abies-marina to extinction. Mar. Environ. Res. 2022, 181, 105759. [Google Scholar] [CrossRef]
- Vea, J.; Ask, E. Creating a sustainable commercial harvest of Laminaria hyperborea in Norway. J. Appl. Phycol. 2011, 23, 489–494. [Google Scholar] [CrossRef]
- Voerman, S.E.; Llera, E.; Rico, J.M. Climate driven changes in subtidal kelp forest communities in NW Spain. Mar. Environ. Res. 2013, 90, 119–127. [Google Scholar] [CrossRef]
- Eger, A.; Aguirre, J.D.; Altamirano, M.; Arafeh-Dalmau, N.; Arroyo, N.L.; Bauer-Civiello, A.M.; Beas-Luna, R.; Bekkby, T.; Bellgrove, A.; Bennett, S.; et al. The Kelp Forest Challenge: A collaborative global movement to protect and restore 4 million hectares of kelp forests. J. Appl. Phycol. 2024, 36, 951–964. [Google Scholar] [CrossRef]
- Harden, M.; Kovalev, M.; Molano, G.; Yorke, C.; Miller, R.; Reed, D.; Alberto, F.; Koos, D.S.; Lansford, R.; Nuzhdin, S. Heat stress analysis suggests a genetic basis for tolerance in Macrocystis pyrifera across developmental stages. Commun. Biol. 2024, 7, 1147. [Google Scholar] [CrossRef]
- Perrow, M.R.; Davy, A.J. Handbook of Ecological Restoration; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Bayraktarov, E.; Saunders, M.I.; Abdullah, S.; Mills, M.; Beher, J.; Possingham, H.P.; Mumby, P.J.; Lovelock, C.E. The cost and feasibility of marine coastal restoration. Ecol. Appl. 2016, 26, 1055–1074. [Google Scholar] [CrossRef]
- Ebbing, A.; Pierik, R.; Bouma, T.; Kromkamp, J.C.; Timmermans, K. How light and biomass density influence the reproduction of delayed Saccharina latissima gametophytes (Phaeophyceae). J. Phycol. 2020, 56, 709–718. [Google Scholar] [CrossRef]
- Peteiro, C.; Freire, Ó. Biomass yield and morphological features ofthe seaweed Saccharina latissima cultivated at two different sites in acoastal bay in the Atlantic coast of Spain. J. Appl. Phycol. 2013, 25, 205–213. [Google Scholar] [CrossRef]
- Forbord, S.; Steinhovden, K.B.; Solvang, T.; Handå, A.; Skjermo, J. Effect of Seeding Methods and Hatchery Periods on Sea Cultivation of Saccharina latissima (Phaeophyceae): A Norwegian Case Study. J. Appl. Phycol. 2020, 32, 2201–2212. [Google Scholar] [CrossRef]
- Martins, N.; Tanttu, H.; Pearson, G.A.; Bartsch, I. Interactions of Daylength, Temperature and Nutrients Affect Thresholds for Life Stage Transitions in the Kelp Laminaria digitata (Phaeophyceae). Bot. Mar. 2017, 60, 109–121. [Google Scholar] [CrossRef]
- Kerrison, P.D.; Stanley, M.S.; Kelly, M.; MacLeod, A.; Black, K.D.; Hughes, A.D. Optimising the Settlement and Hatchery Culture of Saccharina latissima (Phaeophyta) by Manipulation of Growth Medium and Substrate Surface Condition. J. Appl. Phycol. 2016, 28, 1181–1191. [Google Scholar] [CrossRef]
- Nardelli, A.E.; Visch, W.; Farrington, G.; Sanderson, J.C.; Bellgrove, A.; Wright, J.T.; Macleod, C.; Hurd, C.L. A New Nursery Approach Enhances at—Sea Performance in the Kelp Lessonia corrugata. J. Appl. Phycol. 2023, 36, 591–603. [Google Scholar] [CrossRef]
- Dieck, T.I. North Pacific and North Atlantic digitate Laminaria species (Phaeophyta): Hybridization experiments and temperature responses. Phycologia 1992, 31, 147–163. [Google Scholar]
- Harrison, P.J.; Berges, J.A. Marine Culture Media. In Algal Culturing Techniques; Andersen, R., Ed.; Academic Press: New York, NY, USA, 2005; pp. 21–33. [Google Scholar]
- Su, L.; Pang, S.J.; Shan, T.F.; Li, X. Large-scale hatchery of the kelp Saccharina japonica: A case study experience at Lvshun in northern China. J. Appl. Phycol. 2017, 29, 3003–3013. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Brawley, S.H.; Johnson, L.E. Survival Of Fucoid Embryos In The Intertidal Zone Depends Upon Developmental Stage And Microhabitat1. J. Phycol. 1991, 27, 179–186. [Google Scholar] [CrossRef]
- Steen, H.; Scrosati, R. Intraspecific competition in Fucus serratus and F. evanescens (Phaeophyceae: Fucales) germlings: Effects of settlement density, nutrient concentration, and temperature. Mar. Biol. 2004, 144, 61–70. [Google Scholar] [CrossRef]
- Arenas, F.; Viejo, R.M.; Fernández, C. Density-dependent regulation in an invasive seaweed: Responses at plant and modular levels. J. Ecol. 2002, 90, 820–829. [Google Scholar] [CrossRef]
- Alsuwaiyan, N.A.; Vranken, S.; Burkholz, C.; Cambridge, M.; Coleman, M.A.; Wernberg, T. Green gravel as a vector of dispersal for kelp restoration. Front. Mar. Sci. 2022, 9, 910417. [Google Scholar] [CrossRef]
- Lawton, R.J.; Magnusson, M. Effects of seeding twine type and seeding density on hatchery performance and initial at-sea cultivation performance of the kelp Ecklonia radiata. Alg. Res. 2024, 84, 103777. [Google Scholar] [CrossRef]
- Kerrison, P.D.; Twigg, G.; Stanley, M.; De Smet, D.; Buyle, G.; Martínez Pina, A.; Hughes, A.D. Twine selection is essential for successful hatchery cultivation of Saccharina latissima, seeded with either meiospores or juvenile sporophytes. J. Appl. Phycol. 2019, 31, 3051–3060. [Google Scholar] [CrossRef]
- Arantzamendi, L.; Andrés, M.; Basurko, O.C.; Suárez, M.J. Circular and lower impact mussel and seaweed aquaculture by a shift towards bio-based ropes. Rev. Aquac. 2023, 15, 1010–1019. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chemello, S.; Coutinho, A.; Sá, M.F.; Borges, D.; Arenas, F.; Sousa-Pinto, I.; Costa, I. Optimizing Early Growth of Laminaria hyperborea in Controlled Settings: A Pathway to Improved Restoration Efforts. Phycology 2025, 5, 5. https://doi.org/10.3390/phycology5010005
Chemello S, Coutinho A, Sá MF, Borges D, Arenas F, Sousa-Pinto I, Costa I. Optimizing Early Growth of Laminaria hyperborea in Controlled Settings: A Pathway to Improved Restoration Efforts. Phycology. 2025; 5(1):5. https://doi.org/10.3390/phycology5010005
Chicago/Turabian StyleChemello, Sílvia, Ana Coutinho, M. Francisca Sá, Débora Borges, Francisco Arenas, Isabel Sousa-Pinto, and Isabel Costa. 2025. "Optimizing Early Growth of Laminaria hyperborea in Controlled Settings: A Pathway to Improved Restoration Efforts" Phycology 5, no. 1: 5. https://doi.org/10.3390/phycology5010005
APA StyleChemello, S., Coutinho, A., Sá, M. F., Borges, D., Arenas, F., Sousa-Pinto, I., & Costa, I. (2025). Optimizing Early Growth of Laminaria hyperborea in Controlled Settings: A Pathway to Improved Restoration Efforts. Phycology, 5(1), 5. https://doi.org/10.3390/phycology5010005