
Cyanobacteria-Based Bioprocess for Cosmetic Products—Cyanobium sp. as a Novel Source of Bioactive Pigments




Abstract
1. Introduction
2. Cyanobacteria in Biotechnological Processes
3. Cyanobium sp. a Novel Biotechnological Cyanobacteria
4. Production Strategies
5. Downstream Strategies
6. Applicability in Cosmetic Industry
7. Cosmetic Ingredients Regulation
8. Life Cycle Assessment and Scale-up Simulation
9. The Remaining Challenges on Cyanobium Bioprocess
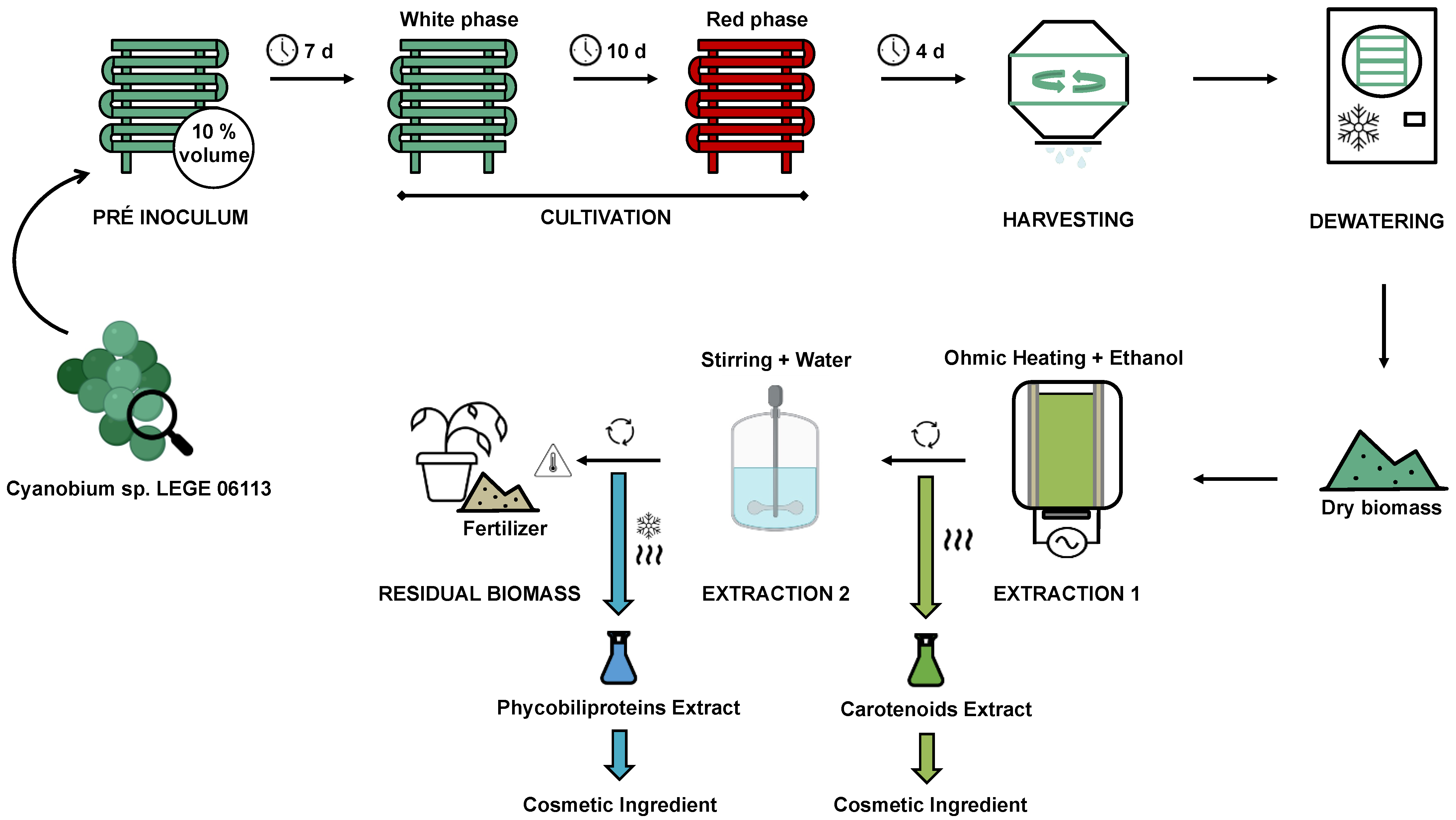
10. Process Overview
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghosh, S.; Sarkar, T.; Das, A.; Chakraborty, R. Natural Colorants from Plant Pigments and Their Encapsulation: An Emerging Window for the Food Industry. LWT 2022, 153, 112527. [Google Scholar] [CrossRef]
- Pagels, F.; Salvaterra, D.; Amaro, H.M.; Guedes, A.C. Pigments from Microalgae. In Handbook of Microalgae-Based Processes and Products; Academc Press: Cambridge, MA, USA, 2020; pp. 465–492. [Google Scholar]
- Masojdek, J.; Koblek, M.; Torzillo, G. Photosynthesis in Microalgae. In Handbook of Microalgal Culture; John Wiley & Sons: Hoboken, NJ, USA, 2007; pp. 20–39. ISBN 9780470673898. [Google Scholar]
- Stewart, S. Painted Faces: A Colourful History of Cosmetics; Amberley Publishing Limited: Stroud, UK, 2017; ISBN 9781445654003. [Google Scholar]
- Pagels, F.; Lopes, G.; Vasconcelos, V.; Guedes, A.C. Cosmetic Applications of Microalgal and Cyanobacterial Pigments. In Algal Genetic Resources Cosmeceuticals, Nutraceuticals, and Pharmaceuticals from Algae; Sangeetha, J., Thangadurai, D., Eds.; Apple Academic Press: Palm Bay, FL, USA, 2022; Volume 1. [Google Scholar]
- Scott, D.A. A Review of Ancient Egyptian Pigments and Cosmetics. Stud. Conserv. 2016, 61, 185–202. [Google Scholar] [CrossRef]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as Sources of High Added-Value Compounds—A Brief Review of Recent Work. Biotechnol. Prog. 2011, 27, 597–613. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.; Bernal-Castillo, J.; Rozo, C.; Rodríguez, I. Sp/Rulina (Arthrospira) an Edible Microorganism a Review. Univ. Sci. 2003, 8, 7–24. [Google Scholar]
- Joshi, S.; Kumari, R.; Upasani, V.N. Applications of Algae in Cosmetics: An Overview. Int. J. Innov. Res. Sci. Eng. Technol. 2018, 7, 1269–1278. [Google Scholar]
- Morone, J.; Alfeus, A.; Vasconcelos, V.; Martins, R. Revealing the Potential of Cyanobacteria in Cosmetics and Cosmeceuticals —A New Bioactive Approach. Algal Res. 2019, 41, 101541. [Google Scholar] [CrossRef]
- Anunciato, T.P.; Rocha-Filho, P.A. Carotenoids and Polyphenols in Nutricosmetics, Nutraceuticals, and Cosmeceuticals. J. Cosmet. Derm. 2012, 11, 51–54. [Google Scholar] [CrossRef]
- Pagels, F.; Vasconcelos, V.; Guedes, A.C. Carotenoids from Cyanobacteria: Biotechnological Potential and Optimization Strategies. Biomolecules 2021, 11, 735. [Google Scholar] [CrossRef]
- Shegokar, R.; Mitri, K. Carotenoid Lutein: A Promising Candidate for Pharmaceutical and Nutraceutical Applications. J. Diet. Suppl. 2012, 9, 183–210. [Google Scholar] [CrossRef]
- Pagels, F.; Guedes, A.C.; Amaro, H.M.; Kijjoa, A.; Vasconcelos, V. Phycobiliproteins from Cyanobacteria: Chemistry and Biotechnological Applications. Biotechnol. Adv. 2019, 37, 422–443. [Google Scholar] [CrossRef]
- Balboa, E.M.; Conde, E.; Soto, M.L.; Pérez-Armada, L.; Domínguez, H. Cosmetics from Marine Sources. In Springer Handbook of Marine Biotechnology; Kim, S.-K., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 1015–1042. ISBN 9783642539718. [Google Scholar]
- Acién, F.G.; Fernández, J.M.; Magán, J.J.; Molina, E. Production Cost of a Real Microalgae Production Plant and Strategies to Reduce It. Biotechnol. Adv. 2012, 30, 1344–1353. [Google Scholar] [CrossRef] [PubMed]
- Richmond, A. Microalgal Biotechnology at the Turn of the Millennium: A Personal View. J. Appl. Phycol. 2000, 12, 441–451. [Google Scholar] [CrossRef]
- Rizwan, M.; Mujtaba, G.; Memon, S.A.; Lee, K.; Rashid, N. Exploring the Potential of Microalgae for New Biotechnology Applications and beyond: A Review. Renew. Sustain. Energy Rev. 2018, 92, 394–404. [Google Scholar] [CrossRef]
- Chen, H.; Li, T.; Wang, Q. Ten Years of Algal Biofuel and Bioproducts: Gains and Pains. Planta 2019, 249, 195–219. [Google Scholar] [CrossRef] [PubMed]
- Beck, C.; Knoop, H.; Axmann, I.M.; Steuer, R. The Diversity of Cyanobacterial Metabolism: Genome Analysis of Multiple Phototrophic Microorganisms. BMC Genom. 2012, 13, 56. [Google Scholar] [CrossRef]
- Sánchez-Baracaldo, P.; Hayes, P.K.; Blank, C.E. Morphological and Habitat Evolution in the Cyanobacteria Using a Compartmentalization Approach. Geobiology 2005, 3, 145–165. [Google Scholar] [CrossRef]
- Uzair, B.; Tabassum, S.; Rasheed, M.; Rehman, S.F. Exploring Marine Cyanobacteria for Lead Compounds of Pharmaceutical Importance. Sci. World J. 2012, 2012, 179782. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Shedid, E.S.; Saied, E.M.; Jassbi, A.R.; Jamebozorgi, F.H.; Rateb, M.E.; Du, M.; Abdel-Daim, M.M.; Kai, G.Y.; Al-Hammady, M.A.M.; et al. Cyanobacteria—From the Oceans to the Potential Biotechnological and Biomedical Applications. Mar. Drugs 2021, 19, 241. [Google Scholar] [CrossRef]
- Grewe, C.B.; Pulz, O. The Biotechnology of Cyanobacteria. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; Volume 9789400738553. [Google Scholar]
- Soni, R.A.; Sudhakar, K.; Rana, R.S. Spirulina—From Growth to Nutritional Product: A Review. Trends Food Sci. Technol. 2017, 69, 157–171. [Google Scholar] [CrossRef]
- Kamravamanesh, D.; Lackner, M.; Herwig, C. Bioprocess Engineering Aspects of Sustainable Polyhydroxyalkanoate Production in Cyanobacteria. Bioengineering 2018, 5, 111. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for Biodiesel Production and Other Applications: A Review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef]
- Anderson, L.K.; Rayner, M.C.; Sweet, R.M.; Eiserling, F.A. Regulation of Nostoc Sp. Phycobilisome Structure by Light and Temperature. J. Bacteriol. 1983, 155, 1407–1416. [Google Scholar] [CrossRef] [PubMed]
- Graham, L.E.; Graham, J.M.; Wilcox, L.W.; Cook, M.E. Algae, 3rd ed.; LJLM Press: Madison, WI, USA, 2016; ISBN 978-0321559654. [Google Scholar]
- Schulze, P.S.C.; Barreira, L.A.; Pereira, H.G.C.; Perales, J.A.; Varela, J.C.S. Light Emitting Diodes (LEDs) Applied to Microalgal Production. Trends Biotechnol. 2014, 32, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Assunção, J.; Malcata, F.X. Enclosed “Non-Conventional” Photobioreactors for Microalga Production: A Review. Algal Res. 2020, 52, 102107. [Google Scholar] [CrossRef]
- Pagels, F.; Pereira, R.N.; Vicente, A.A.; Guedes, A.C. Extraction of Pigments from Microalgae and Cyanobacteria—A Review on Current Methodologies. Appl. Sci. 2021, 11, 5187. [Google Scholar] [CrossRef]
- Molina-Grima, E.; Belarbi, E.H.; Acién Fernández, F.G.; Robles Medina, A.; Chisti, Y. Recovery of Microalgal Biomass and Metabolites: Process Options and Economics. Biotechnol. Adv. 2003, 20, 491–515. [Google Scholar] [CrossRef]
- de Morais, M.G.; da Fontoura Prates, D.; Moreira, J.B.; Duarte, J.H.; Costa, J.A.V. Phycocyanin from Microalgae: Properties, Extraction and Purification, with Some Recent Applications. Ind. Biotechnol. 2018, 14, 30–37. [Google Scholar] [CrossRef]
- Bastiaens, L.; van Roy, S.; Thomassen, G.; Elst, K. Biorefinery of Algae: Technical and Economic Considerations. In Microalgae-Based Biofuels and Bioproducts: From Feedstock Cultivation to End-Products; Gonzalez-Fernandez, C., Muñoz, R., Eds.; Woodhead Publishing: Cambridge, UK, 2017; pp. 327–345. ISBN 9780081010273. [Google Scholar]
- Ramos, V.; Morais, J.; Castelo-Branco, R.; Pinheiro, Â.; Martins, J.; Regueiras, A.; Pereira, A.L.; Lopes, V.R.; Frazão, B.; Gomes, D.; et al. Cyanobacterial Diversity Held in Microbial Biological Resource Centers as a Biotechnological Asset: The Case Study of the Newly Established LEGE Culture Collection. J. Appl. Phycol. 2018, 30, 1437–1451. [Google Scholar] [CrossRef]
- Costa, M.; Sampaio-Dias, I.E.; Castelo-Branco, R.; Scharfenstein, H.; Rezende De Castro, R.; Silva, A.; Schneider, M.P.C.; Araújo, M.J.; Martins, R.; Domingues, V.F.; et al. Structure of Hierridin c, Synthesis of Hierridins b and c, and Evidence for Prevalent Alkylresorcinol Biosynthesis in Picocyanobacteria. J. Nat. Prod. 2019, 82, 393–402. [Google Scholar] [CrossRef]
- Freitas, S.; Martins, R.; Costa, M.; Leão, P.N.; Vitorino, R.; Vasconcelos, V.; Urbatzka, R. Hierridin B Isolated from a Marine Cyanobacterium Alters VDAC1, Mitochondrial Activity, and Cell Cycle Genes on HT-29 Colon Adenocarcinoma Cells. Mar. Drugs 2016, 14, 158. [Google Scholar] [CrossRef]
- Leão, P.N.; Costa, M.; Ramos, V.; Pereira, A.R.; Fernandes, V.C.; Domingues, V.F.; Gerwick, W.H.; Vasconcelos, V.M.; Martins, R. Antitumor Activity of Hierridin B, a Cyanobacterial Secondary Metabolite Found in Both Filamentous and Unicellular Marine Strains. PLoS ONE 2013, 8, e69562. [Google Scholar] [CrossRef] [PubMed]
- Leão, P.N.; Martins, M.D.R.; Costa, M.; Vasconcelos, V.; Domingues, V.; Nogueira, F. Antimalarial Agent, Methods and Uses Thereof. Patent No. WO2016207869A1, 29 December 2016. [Google Scholar]
- Brandão, P.; Moreira, J.; Almeida, J.; Nazareth, N.; Sampaio-Dias, I.E.; Vasconcelos, V.; Martins, R.; Leão, P.; Pinto, M.; Saraíva, L.; et al. Norhierridin B, a New Hierridin B-Based Hydroquinone with Improved Antiproliferative Activity. Molecules 2020, 25, 1578. [Google Scholar] [CrossRef] [PubMed]
- Pagels, F.; Barufi, J.B.; Vega, J.; Abdala-Díaz, R.; Vasconcelos, V.; Guedes, A.C.; Figueroa, F.L. Light Regulating Metabolic Responses of Cyanobium Sp. (Cyanobacteria). Fundam. Appl. Limnol. 2020, 193, 285–297. [Google Scholar] [CrossRef]
- Pagels, F.; Bonomi-Barufi, J.; Vega, J.; Abdala-Díaz, R.; Vasconcelos, V.; Guedes, A.C.; Figueroa, F.L. Light Quality Triggers Biochemical Modulation of Cyanobium Sp.—Photobiology as Tool for Biotechnological Optimization. J. Appl. Phycol. 2020, 32, 2851–2861. [Google Scholar] [CrossRef]
- Pagels, F.; Salvaterra, D.; Amaro, H.M.; Lopes, G.; Sousa-Pinto, I.; Vasconcelos, V.; Guedes, A.C. Factorial Optimization of Upstream Process for Cyanobium Sp. Pigments Production. J. Appl. Phycol. 2020, 32, 3861–3872. [Google Scholar] [CrossRef]
- Pagels, F.; Lopes, G.; Vasconcelos, V.; Guedes, A.C. White and Red LEDs as Two-Phase Batch for Cyanobacterial Pigments Production. Bioresour. Technol. 2020, 307, 123105. [Google Scholar] [CrossRef]
- Pagels, F.; Salvaterra, D.; Amaro, H.M.; Lopes, G.; Sousa-Pinto, I.; Vasconcelos, V.; Guedes, A.C. Bioactive Potential of Cyanobium Sp. Pigment-Rich Extracts. J. Appl. Phycol. 2020, 32, 3031–3040. [Google Scholar] [CrossRef]
- Pagels, F.; Pereira, R.N.; Amaro, H.M.; Vasconcelos, V.; Guedes, A.C.; Vicente, A.A. Continuous Pressurized Extraction versus Electric Fields-Assisted Extraction of Cyanobacterial Pigments. J. Biotechnol. 2021, 334, 35–42. [Google Scholar] [CrossRef]
- Pagels, F.; Almeida, C.; Vasconcelos, V.; Guedes, A.C. Cosmetic Potential of Pigments Extracts from the Marine Cyanobacterium Cyanobium Sp. Mar. Drugs 2022, 20, 481. [Google Scholar] [CrossRef]
- Pagels, F.; Arias, A.; Guedes, A.C.; Vicente, A.A.; Vasconcelos, V.; Moreira, M.T. Identifying Key Environmental Indicators in the Assessment of the Proof-of-Concept in Pigment Production from the Marine Cyanobacterium Cyanobium Sp. Appl. Sci. 2022, 12, 12999. [Google Scholar] [CrossRef]
- Rippka, R.; Cohen-Bazire, G. The Cyanobacteriales: A Legitimate Order Based on the Type Strain Cyanobacterium Stanieri? Ann. Inst. Pasteur Microbiol. 1983, 134, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Guiry, M.D. How Many Species of Algae Are There? J. Phycol. 2012, 48, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.R.J.; Siqueira, A.S.; dos Santos, B.G.S.; da Silva, F.D.F.; Lima, C.P.; Cardoso, J.F.; Vianez, J.L.D.S.G.; Dall’Agnol, L.T.; McCulloch, J.A.; Nunes, M.R.T.; et al. Draft Genome Sequence of the Brazilian Cyanobium sp. Strain CACIAM 14. Genome Announc. 2014, 2, 669–714. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Shimura, Y.; Suzuki, S.; Yamagishi, T.; Tatarazako, N.; Kawachi, M. Complete Genome Sequence of Cyanobium Sp. NIES-981, a Marine Strain Potentially Useful for Ecotoxicological Bioassays. Genome Announc. 2016, 4, e00736-16. [Google Scholar] [CrossRef] [PubMed]
- Komárek, J.; Kopecký, J.; Cepák, V. Generic Characters of the Simplest Cyanoprokaryotes Cyanobium, Cyanobacterium and Synechococcus. Cryptogam. Algol. 1999, 20, 209–222. [Google Scholar] [CrossRef]
- Pérez-Portilla, P.; Araya, J.; Gallardo, K.; Aránguiz-Acuña, A. Potential of Arsenic Bioremediation by a Cyanobacterium Isolated from the Salado River in the Atacama Desert. J. Plankton Res. 2021, 43, 156–160. [Google Scholar] [CrossRef]
- Armstrong, L.; Vaz, M.G.M.V.; Genuário, D.B.; Fiore, M.F.; Debonsi, H.M. Volatile Compounds Produced by Cyanobacteria Isolated from Mangrove Environment. Curr. Microbiol. 2019, 76, 575–582. [Google Scholar] [CrossRef]
- Lopes, G.; Clarinha, D.; Vasconcelos, V. Carotenoids from Cyanobacteria: A Biotechnological Approach for the Topical Treatment of Psoriasis. Microorganisms 2020, 8, 302. [Google Scholar] [CrossRef]
- Morone, J.; Lopes, G.; Preto, M.; Vasconcelos, V.; Martins, R. Exploitation of Filamentous and Picoplanktonic Cyanobacteria for Cosmetic Applications: Potential to Improve Skin Structure and Preserve Dermal Matrix Components. Mar. Drugs 2020, 18, 486. [Google Scholar] [CrossRef]
- Adriano, A.H.; Gabriel, M.D.R.; Luiza, M.; Michele, G.D.M.; Jorge, A.V.C. The Cultivation of Microalgae Cyanobium Sp. and Chlorella Sp. in Different Culture Media and Stirring Setting. Afr. J. Microbiol. Res. 2015, 9, 1431–1439. [Google Scholar] [CrossRef]
- Henrard, A.A.; de Morais, M.G.; Costa, J.A.V. Vertical Tubular Photobioreactor for Semicontinuous Culture of Cyanobium sp. Bioresour. Technol. 2011, 102, 4897–4900. [Google Scholar] [CrossRef] [PubMed]
- Gutu, A.; Kehoe, D.M. Emerging Perspectives on the Mechanisms, Regulation, and Distribution of Light Color Acclimation in Cyanobacteria. Mol. Plant. 2012, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, B.L. Seeing New Light: Recent Insights into the Occurrence and Regulation of Chromatic Acclimation in Cyanobacteria. Curr. Opin. Plant Biol. 2017, 37, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Wiltbank, L.B.; Kehoe, D.M. Diverse Light Responses of Cyanobacteria Mediated by Phytochrome Superfamily Photoreceptors. Nat. Rev. Microbiol. 2019, 17, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Hemlata; Fatma, T. Screening of Cyanobacteria for Phycobiliproteins and Effect of Different Environmental Stress on Its Yield. Bull. Env. Contam. Toxicol. 2009, 83, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Luna, L.D.; Bezerra, R.P.; Matsudo, M.C.; Sato, S.; Converti, A.; de Carvalho, J.C.M. Influence of PH, Temperature, and Urea Molar Flowrate on Arthrospira Platensis Fed-Batch Cultivation: A Kinetic and Thermodynamic Approach. Biotechnol. Bioeng. 2007, 96, 702–711. [Google Scholar] [CrossRef]
- Keithellakpam, O.S.; Nath, T.O.; Oinam, A.S.; Thingujam, I.; Oinam, G.; Dutt, S.G. Effect of External PH on Cyanobacterial Phycobiliproteins Production and Ammonium Excretion. J. Appl. Biol. Biotechnol. 2015, 3, 38–42. [Google Scholar] [CrossRef]
- Mishra, S.K.; Shrivastav, A.; Maurya, R.R.; Patidar, S.K.; Haldar, S.; Mishra, S. Effect of Light Quality on the C-Phycoerythrin Production in Marine Cyanobacteria Pseudanabaena sp. Isolated from Gujarat Coast, India. Protein Expr. Purif. 2012, 81, 5–10. [Google Scholar] [CrossRef]
- Khattar, J.I.S.; Kaur, S.; Kaushal, S.; Singh, Y.; Singh, D.P.; Rana, S.; Gulati, A. Hyperproduction of Phycobiliproteins by the Cyanobacterium Anabaena Fertilissima PUPCCC 410.5 under Optimized Culture Conditions. Algal Res. 2015, 12, 463–469. [Google Scholar] [CrossRef]
- Vijaya, V.; Anand, N. Blue light enhance the pigment synthesis in cyanobacterium Anabaena Ambigua Rao (NOSTACALES). ARPN J. Agric. Biol. Sci. 2009, 4, 36–43. [Google Scholar]
- Ma, R.; Lu, F.; Bi, Y.; Hu, Z. Effects of Light Intensity and Quality on Phycobiliprotein Accumulation in the Cyanobacterium Nostoc Sphaeroides Kützing. Biotechnol. Lett. 2015, 37, 1663–1669. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.N.; Shin, H.S.; Park, H.G.; Lee, J.; Kil, G.S.; Choi, C.Y. Profiles of Photosynthetic Pigment Accumulation and Expression of Photosynthesis-Related Genes in the Marine Cyanobacteria Synechococcus Sp.: Effects of LED Wavelengths. Biotechnol. Bioprocess Eng. 2014, 19, 250–256. [Google Scholar] [CrossRef]
- Kumar, M.; Kulshreshtha, J.; Singh, G.P. Growth and Biopigment Accumulation of Cyanobacterium Spirulina Platensis at Different Light Intensities and Temperature. Braz. J. Microbiol. 2011, 42, 1128–1135. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Zhao, F.; Wei, W.; Wen, Z.; Qin, S. Carotenoid Biosynthesis in Cyanobacteria: Structural and Evolutionary Scenarios Based on Comparative Genomics. Int. J. Biol. Sci. 2006, 2, 197–207. [Google Scholar] [CrossRef]
- Lima, G.M.; Teixeira, P.C.N.; Teixeira, C.M.L.L.; Filócomo, D.; Lage, C.L.S. Influence of Spectral Light Quality on the Pigment Concentrations and Biomass Productivity of Arthrospira Platensis. Algal Res. 2018, 31, 157–166. [Google Scholar] [CrossRef]
- Olaizola, M.; Duerr, E.O. Effects of Light Intensity and Quality on the Growth Rate and Photosynthetic Pigment Content of Spirulina Platensis. J. Appl. Phycol. 1990, 2, 97–104. [Google Scholar] [CrossRef]
- Stirk, W.A.; Bálint, P.; Vambe, M.; Lovász, C.; Molnár, Z.; van Staden, J.; Ördög, V. Effect of Cell Disruption Methods on the Extraction of Bioactive Metabolites from Microalgal Biomass. J. Biotechnol. 2020, 307, 35–43. [Google Scholar] [CrossRef]
- Salinas-Salazar, C.; Garcia-Perez, J.S.; Chandra, R.; Castillo-Zacarias, C.; Iqbal, H.M.N.; Parra-Saldívar, R. Methods for Extraction of Valuable Products from Microalgae Biomass. In Microalgae Biotechnology for Development of Biofuel and Wastewater Treatment; Alam, M.A., Wang, Z., Eds.; Springer: Singapore, 2019; pp. 245–263. ISBN 9789811322648. [Google Scholar]
- Sarada, R.; Pillai, M.G.; Ravishankar, G.A. Phycocyanin from Spirulina Sp: Influence of Processing of Biomass on Phycocyanin Yield, Analysis of Efficacy of Extraction Methods and Stability Studies on Phycocyanin. Process Biochem. 1999, 34, 795–801. [Google Scholar] [CrossRef]
- Rao, A.R.; Sindhuja, H.N.; Dharmesh, S.M.; Sankar, K.U.; Sarada, R.; Ravishankar, G.A. Effective Inhibition of Skin Cancer, Tyrosinase, and Antioxidative Properties by Astaxanthin and Astaxanthin Esters from the Green Alga Haematococcus Pluvialis. J. Agric. Food Chem. 2013, 61, 3842–3851. [Google Scholar] [CrossRef]
- Rodríguez-Meizoso, I.; Jaime, L.; Santoyo, S.; Cifuentes, A.; García-Blairsy Reina, G.; Señoráns, F.J.; Ibáñez, E. Pressurized Fluid Extraction of Bioactive Compounds from Phormidium Species. J. Agric. Food Chem. 2008, 56, 3517–3523. [Google Scholar] [CrossRef]
- Martínez, J.M.; Luengo, E.; Saldaña, G.; Álvarez, I.; Raso, J. C-Phycocyanin Extraction Assisted by Pulsed Electric Field from Artrosphira Platensis. Food Res. Int. 2017, 99, 1042–1047. [Google Scholar] [CrossRef]
- Favas, R.; Morone, J.; Martins, R.; Vasconcelos, V.; Lopes, G. Cyanobacteria and Microalgae Bioactive Compounds in Skin-Ageing: Potential to Restore Extracellular Matrix Filling and Overcome Hyperpigmentation. J. Enzym. Inhib. Med. Chem. 2021, 36, 1829–1838. [Google Scholar] [CrossRef] [PubMed]
- Montalvo, G.E.B.; Thomaz-Soccol, V.; Vandenberghe, L.P.S.; Carvalho, J.C.; Faulds, C.B.; Bertrand, E.; Prado, M.R.M.; Bonatto, S.J.R.; Soccol, C.R. Arthrospira Maxima OF15 Biomass Cultivation at Laboratory and Pilot Scale from Sugarcane Vinasse for Potential Biological New Peptides Production. Bioresour. Technol. 2019, 273, 103–113. [Google Scholar] [CrossRef]
- Boon, C.S.; McClements, D.J.; Weiss, J.; Decker, E.A. Factors Influencing the Chemical Stability of Carotenoids in Foods. Crit. Rev. Food Sci. Nutr. 2010, 50, 515–532. [Google Scholar] [CrossRef] [PubMed]
- Galetović, A.; Dufossé, L. Phycobiliproteins as Food Additives. In Pigments from Microalgae Handbook; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Szterk, A.; Roszko, M.; Górnicka, E. Chemical Stability of the Lipid Phase in Concentrated Beverage Emulsions Colored with Natural β-Carotene. JAOCS J. Am. Oil Chem. Soc. 2013, 90, 483–491. [Google Scholar] [CrossRef]
- Couteau, C.; Coiffard, L. Microalgal Application in Cosmetics. In Microalgae in Health and Disease Prevention; Academic Press: Cambridge, MA, USA, 2018; pp. 317–323. ISBN 9780128114056. [Google Scholar]
- Sahin, S.C. The Potential of Arthrospira Platensis Extract as a Tyrosinase Inhibitor for Pharmaceutical or Cosmetic Applications. S. Afr. J. Bot. 2018, 119, 236–243. [Google Scholar] [CrossRef]
- Singh, D.P.; Prabha, R.; Meena, K.K.; Sharma, L.; Sharma, A.K. Induced Accumulation of Polyphenolics and Flavonoids in Cyanobacteria under Salt Stress Protects Organisms through Enhanced Antioxidant Activity. Am. J. Plant. Sci. 2014, 5, 726–735. [Google Scholar] [CrossRef]
- Castangia, I.; Manca, M.L.; Catalán-Latorre, A.; Maccioni, A.M.; Fadda, A.M.; Manconi, M. Phycocyanin-Encapsulating Hyalurosomes as Carrier for Skin Delivery and Protection from Oxidative Stress Damage. J. Mater. Sci. Mater. Med. 2016, 27, 75. [Google Scholar] [CrossRef]
- Scoglio, S.; Canestrari, F.; Benedetti, S.; Zolla, L. Extracts of Aphanizomenon Flos Aquae and Nutritional Cosmetic and Pharmaceutical Compositons Containing the Same. U.S. Patent US 2010/0021493 A1, 28 January 2010. [Google Scholar]
- Ruiz Canovas, E.; López Cerro, M.T.; Latil de Ros, D.G.; Durany Turk, O.; Segura De Yebra, J.; Mercadé Roca, J. Compositions for Protecting Skin Comprising DNA Repair Enzymes and Phycobiliprotein. U.S. Patent US 11071707, 20 July 2016. [Google Scholar]
- Dorato, S. General Concepts: Current Legislation on Cosmetics in Various Countries. In Analysis of Cosmetic Products, 2nd ed.; Salvador, A., Chsvert, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar] [CrossRef]
- Novoveská, L.; Ross, M.E.; Stanley, M.S.; Pradelles, R.; Wasiolek, V.; Sassi, J.F. Microalgal Carotenoids: A Review of Production, Current Markets, Regulations, and Future Direction. Mar. Drugs 2019, 17, 640. [Google Scholar] [CrossRef]
- Nikitakis, J.; Lange, B. International Cosmetic Ingredient Dictionary and Handbook; Personal Care Products Council: Washington, DC, USA, 2016. [Google Scholar]
- Pistollato, F.; Madia, F.; Corvi, R.; Munn, S.; Grignard, E.; Paini, A.; Worth, A.; Bal-Price, A.; Prieto, P.; Casati, S.; et al. Current EU Regulatory Requirements for the Assessment of Chemicals and Cosmetic Products: Challenges and Opportunities for Introducing New Approach Methodologies. Arch. Toxicol. 2021, 95, 1867–1897. [Google Scholar] [CrossRef]
- Lopes, T.F.; Cabanas, C.; Silva, A.; Fonseca, D.; Santos, E.; Guerra, L.T.; Sheahan, C.; Reis, A.; Gírio, F. Process Simulation and Techno-Economic Assessment for Direct Production of Advanced Bioethanol Using a Genetically Modified Synechocystis sp. Bioresour. Technol. Rep. 2019, 6, 113–122. [Google Scholar] [CrossRef]
- Rodríguez, R.; Espada, J.J.; Moreno, J.; Vicente, G.; Bautista, L.F.; Morales, V.; Sánchez-Bayo, A.; Dufour, J. Environmental Analysis of Spirulina Cultivation and Biogas Production Using Experimental and Simulation Approach. Renew. Energy 2018, 129, 724–732. [Google Scholar] [CrossRef]
- Zhang, Y.I.; Singh, S.; Bakshi, B.R. Accounting for Ecosystem Sewices in Life Cycle Assessment Part I: A Critical Review. Env. Sci. Technol. 2010, 44, 2232–2242. [Google Scholar] [CrossRef] [PubMed]
- Acién, F.G.; Molina, E.; Fernández-Sevilla, J.M.; Barbosa, M.; Gouveia, L.; Sepúlveda, C.; Bazaes, J.; Arbib, Z. Economics of Microalgae Production. In Microalgae-Based Biofuels and Bioproducts: From Feedstock Cultivation to End-Products; Gonzalez-Fernandez, C., Muñoz, R., Eds.; Woodhead Publishing: Cambridge, UK, 2017; pp. 485–503. ISBN 9780081010273. [Google Scholar]
- Trabelsi, L.; ben Ouada, H.; Zili, F.; Mazhoud, N.; Ammar, J. Evaluation of Arthrospira Platensis Extracellular Polymeric Substances Production in Photoautotrophic, Heterotrophic and Mixotrophic Conditions. Folia Microbiol. 2013, 58, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Pereira, H.; Páramo, J.; Silva, J.; Marques, A.; Barros, A.; Maurício, D.; Santos, T.; Schulze, P.; Barros, R.; Gouveia, L.; et al. Scale-up and Large-Scale Production of Tetraselmis Sp. CTP4 (Chlorophyta) for CO2 Mitigation: From an Agar Plate to 100-M3 Industrial Photobioreactors. Sci. Rep. 2018, 8, 5112. [Google Scholar] [CrossRef]
- Heasman, M.; Diemar, J.; O’Connor, W.; Sushames, T.; Foulkes, L. Development of Extended Shelf-Life Microalgae Concentrate Diets Harvested by Centrifugation for Bivalve Molluscs—A Summary. Aquac. Res. 2000, 31, 637–659. [Google Scholar] [CrossRef]
- Guedes, A.C.; Katkam, N.G.; Varela, J.; Malcata, F.X. Photobioreactors for Cyanobacterial Culturing. In Cyanobacteria: An Economic Perspective; John Wiley and Sons: Hoboken, NJ, USA, 2014; pp. 270–292. [Google Scholar] [CrossRef]
- Jeevanandam, J.; Danquah, M.K. Chapter 9—Dewatering and Drying of Algal Cultures. In Handbook of Microalgae-Based Processes and Products; Jacob-Lopes, E., Maroneze, M.M., Queiroz, M.I., Zepka, L.Q., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 207–224. ISBN 978-0-12-818536-0. [Google Scholar]
- Seghiri, R.; Legrand, J.; Hsissou, R.; Essamri, A. Comparative Study of the Impact of Conventional and Unconventional Drying Processes on Phycobiliproteins from Arthrospira Platensis. Algal Res. 2021, 53, 102165. [Google Scholar] [CrossRef]
- Pérez-López, P.; González-García, S.; Jeffryes, C.; Agathos, S.N.; McHugh, E.; Walsh, D.; Murray, P.; Moane, S.; Feijoo, G.; Moreira, M.T. Life Cycle Assessment of the Production of the Red Antioxidant Carotenoid Astaxanthin by Microalgae: From Lab to Pilot Scale. J. Clean Prod. 2014, 64, 332–344. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameter | Optimized Condition | Comments |
|---|---|---|
| Growth time | 14 days | The culture requires a pre-inoculum of 7 days, and has no lag phase |
| Light source | White and Red LEDs | Ten days of white and four days of red increase the pigment content by up to 1.5-fold |
| Light intensity | 200 μmol photons m−2 s−1 | Also found to be the saturation intensity for the photosynthetic metabolism |
| Temperature | 20 °C | The versatility of a strain to adapt to a wider range of temperatures allows its cultivation in different areas or climatic conditions |
| pH | 9.0 | In general, cyanobacteria tend to grow better in neutral to alkaline environments, although the range also varies between species |
| Salinity | 10 g L−1 of NaCl (10 ‰) | Salinity alone had low impact, and only showed expressive results when evaluated with temperature and pH |
| Culture medium | BG 11 with doubled nitrate and phosphate | Increase of about 10 % of biomass and pigments productivity |
| Other parameters | Photoperiod: 16:8 h light:dark Aeration: 0.7 vvm | These parameters were not optimized in experimental trials, although considered in preliminary studies |
| Outputs | Value | Comments |
| Biomass productivity | 137.4 g L−1 d−1 | Final biomass concentration can get up to 2 g L−1 |
| Carotenoids content | 3.3 %DW | Mainly composed of β-carotene, zeaxanthin, lutein, and echinenone |
| Phycobiliproteins content | 12.4 %DW | Ratio 1:2 phycocyanin: allophycocyanin, with no presence of phycoerythrin |
| Parameter | Optimized Condition | Comments |
|---|---|---|
| Solvent route | Ethanol -> Water | No cytotoxic effect and recovery of about 50 %DW |
| Extraction method | Ohmic heating | Scalable technology with current use in food industry, allows an efficient use of thermal and electrical effects |
| Time | 5 min | 4-times lower when compared to CPSE |
| Temperature | 70 °C | Temperature was the most influent factor both in CPSE and ohmic heating optimizations |
| Frequency | 20 kHz | The advantage of a frequency higher than 1 kHz is the absence of corrosion on the electrodes |
| Outputs | Value | Comments |
| Carotenoids extract | 0.27 g gDW−1 | 13 % of carotenoids |
| Phycobiliproteins extract | 0.25 g gDW−1 | 57 % of phycobiliproteins; purity ca. 2.5, in agreement to food grade |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pagels, F.; Guedes, A.C.; Vicente, A.A.; Vasconcelos, V. Cyanobacteria-Based Bioprocess for Cosmetic Products—Cyanobium sp. as a Novel Source of Bioactive Pigments. Phycology 2023, 3, 47-64. https://doi.org/10.3390/phycology3010004
Pagels F, Guedes AC, Vicente AA, Vasconcelos V. Cyanobacteria-Based Bioprocess for Cosmetic Products—Cyanobium sp. as a Novel Source of Bioactive Pigments. Phycology. 2023; 3(1):47-64. https://doi.org/10.3390/phycology3010004
Chicago/Turabian StylePagels, Fernando, A. Catarina Guedes, António A. Vicente, and Vitor Vasconcelos. 2023. "Cyanobacteria-Based Bioprocess for Cosmetic Products—Cyanobium sp. as a Novel Source of Bioactive Pigments" Phycology 3, no. 1: 47-64. https://doi.org/10.3390/phycology3010004
APA StylePagels, F., Guedes, A. C., Vicente, A. A., & Vasconcelos, V. (2023). Cyanobacteria-Based Bioprocess for Cosmetic Products—Cyanobium sp. as a Novel Source of Bioactive Pigments. Phycology, 3(1), 47-64. https://doi.org/10.3390/phycology3010004