
SARS-CoV-2 Replication Revisited: Molecular Insights and Current and Emerging Antiviral Strategies


Abstract

1. Introduction
2. Molecular Mechanisms of SARS-CoV-2 Replication
2.1. Replication Complex and Enzymatic Machinery
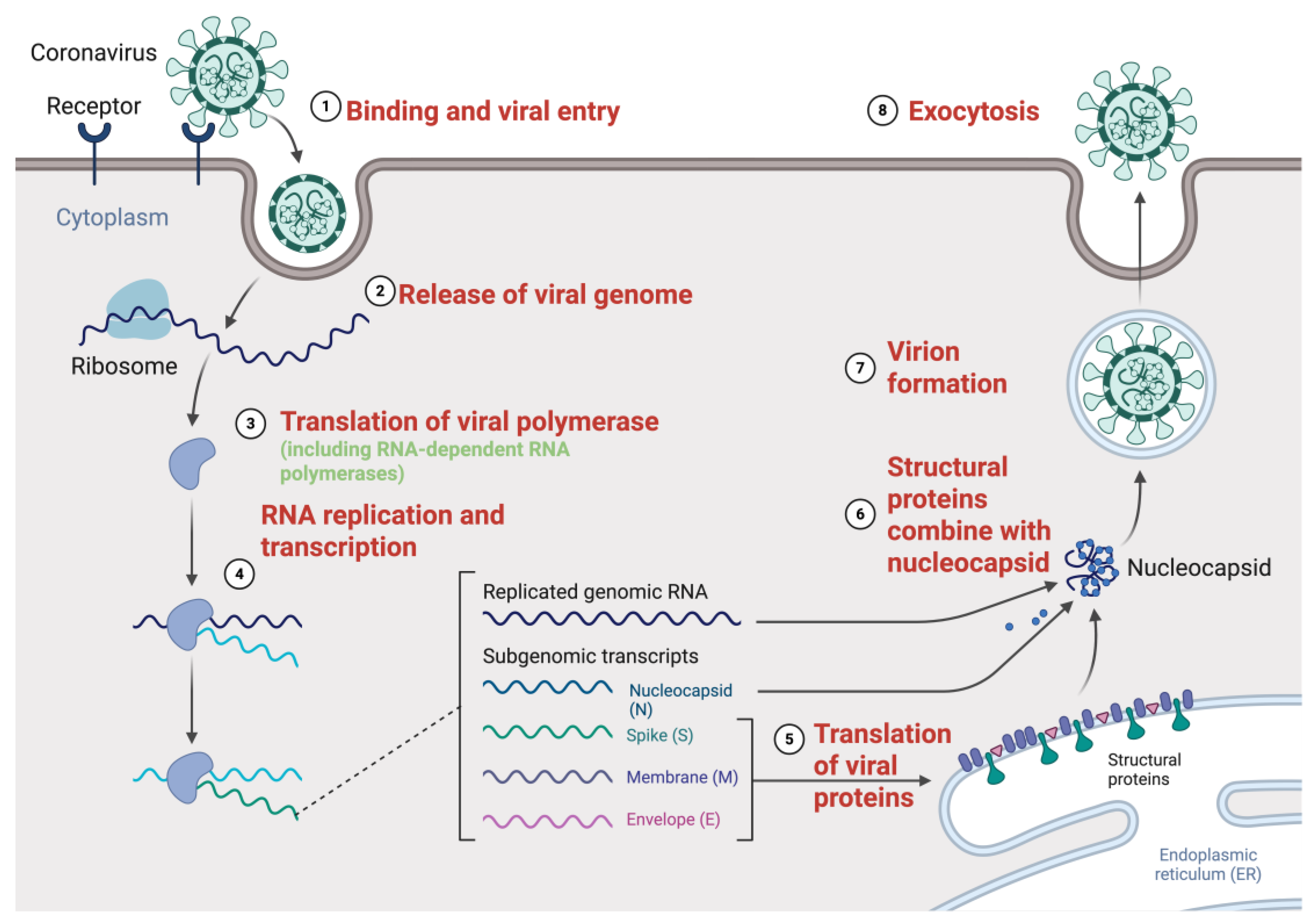
2.2. Genome Replication Cycle
2.3. Host Factors in Viral Replication
3. Drugs Targeting SARS-CoV-2 Replication
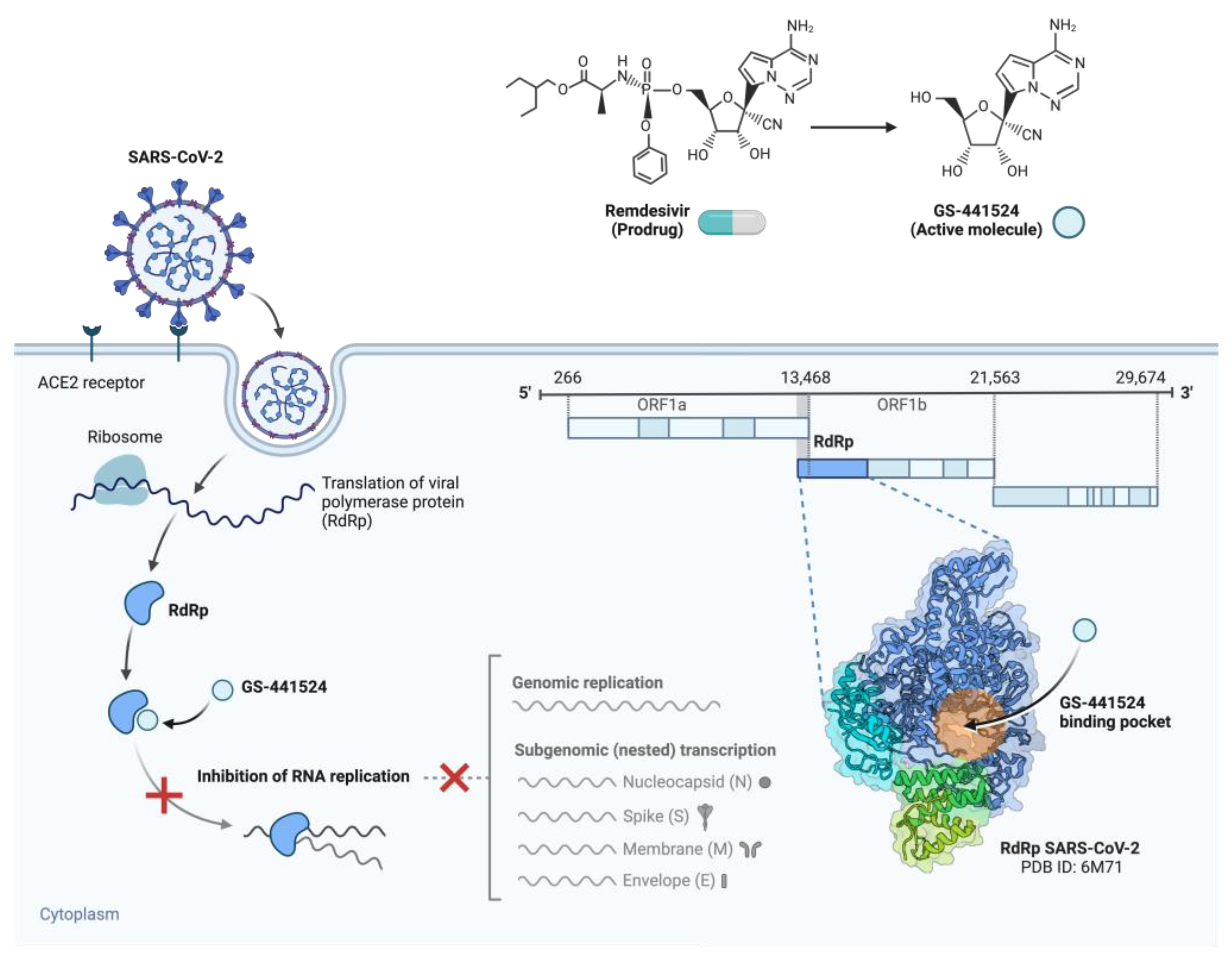
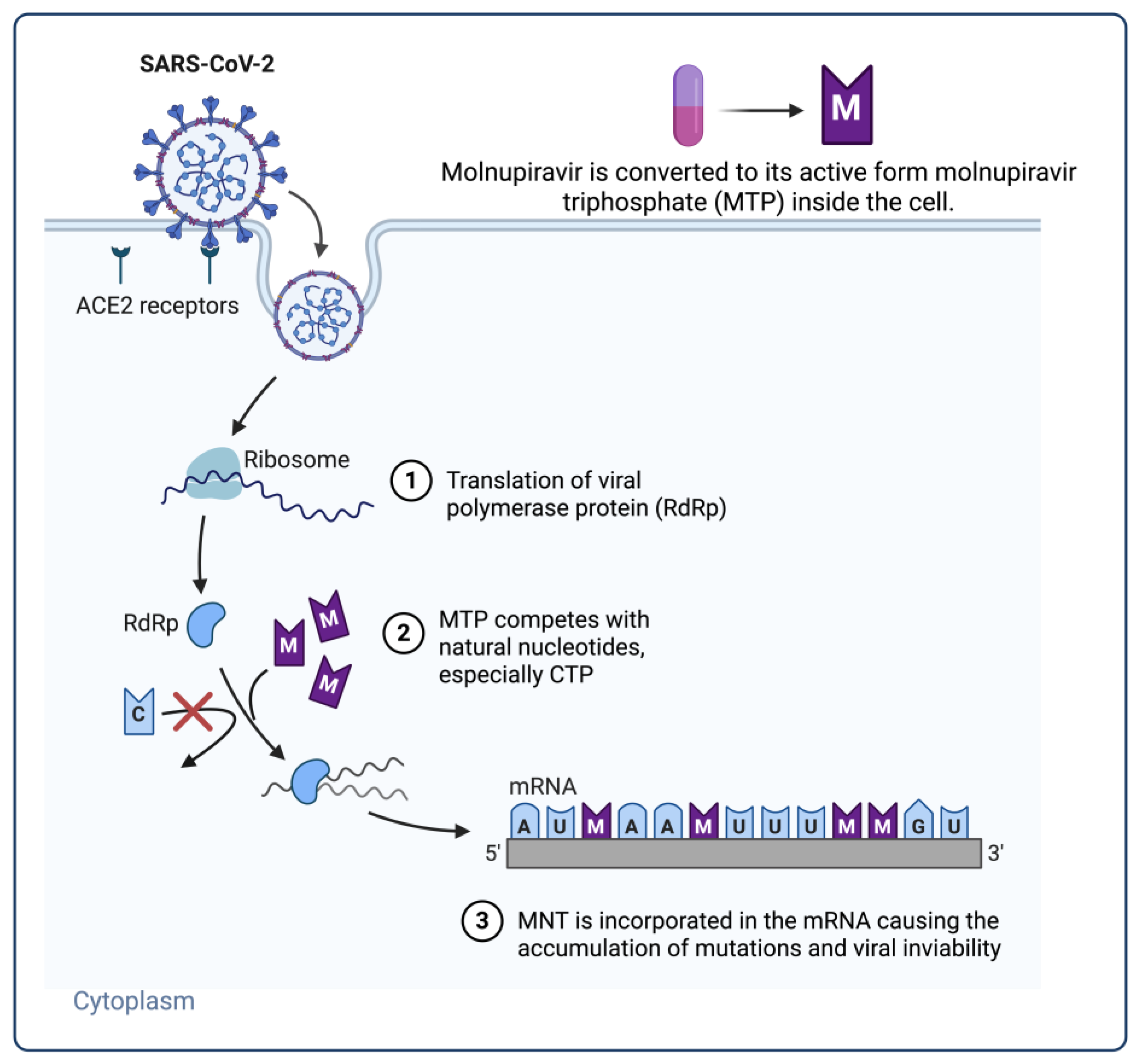
3.1. Nucleoside Analogue Polymerase Inhibitors
3.2. Protease Inhibitors
3.3. Host-Directed Antivirals
3.4. Combination Therapies and Drug Synergy
3.5. Antiviral Resistance Considerations
4. Future Trends and Emerging Antiviral Strategies
4.1. Novel Targets in the Viral Replication Cycle
4.2. Emerging and Novel Host-Directed Antivirals
4.3. AI and Computational Drug Discovery
4.4. Gene Silencing, RNA-Targeting Therapies, Molecular Glues and Targeted Protein Degraders
4.5. Pan-Coronavirus and Broad-Spectrum Antivirals
5. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.-L. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2021, 19, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, A.; Preiss, A.J.; Xiao, X.; Brannock, M.D.; Alexander, G.C.; Chew, R.F.; Davis, H.; Fitzgerald, M.; Hill, E.; Kelly, E.P.; et al. Effect of nirmatrelvir/ritonavir (Paxlovid) on hospitalization among adults with COVID-19: An electronic health record-based target trial emulation from N3C. PLoS Med. 2025, 22, e1004493. [Google Scholar] [CrossRef]
- Faisal, H.M.N.; Katti, K.S.; Katti, D.R. Differences in Interactions Within Viral Replication Complexes of SARS-CoV-2 (COVID-19) and SARS-CoV Coronaviruses Control RNA Replication Ability. JOM (1989) 2021, 73, 1684–1695. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Mao, C.; Luan, X.; Shen, D.-D.; Shen, Q.; Su, H.; Wang, X.; Zhou, F.; Zhao, W.; Gao, M.; et al. Structural Basis for Inhibition of the RNA-Dependent RNA Polymerase from SARS-CoV-2 by Remdesivir. Science 2020, 368, 1499–1504. [Google Scholar] [CrossRef]
- Saberi, A.; Gulyaeva, A.A.; Brubacher, J.L.; Newmark, P.A.; Gorbalenya, A.E. A Planarian Nidovirus Expands the Limits of RNA Genome Size. PLoS Pathog. 2018, 14, e1007314. [Google Scholar] [CrossRef] [PubMed]
- Krell, P.J. An Introduction to Viruses of Invertebrates. In Encyclopedia of Virology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 699–723. ISBN 978-0-12-814516-6. [Google Scholar]
- Bettini, A.; Lapa, D.; Garbuglia, A.R. Diagnostics of Ebola Virus. Front. Public. Health 2023, 11, 1123024. [Google Scholar] [CrossRef]
- Hillen, H.S. Structure and Function of SARS-CoV-2 Polymerase. Curr. Opin. Virol. 2021, 48, 82–90. [Google Scholar] [CrossRef]
- Terrier, O.; Si-Tahar, M.; Ducatez, M.; Chevalier, C.; Pizzorno, A.; Le Goffic, R.; Crépin, T.; Simon, G.; Naffakh, N. Influenza Viruses and Coronaviruses: Knowns, Unknowns, and Common Research Challenges. PLoS Pathog. 2021, 17, e1010106. [Google Scholar] [CrossRef]
- Singh, A.K.; Singh, A.; Singh, R.; Misra, A. Molnupiravir in COVID-19: A Systematic Review of Literature. Diabetes Metab. Syndr. Clin. Res. Rev. 2021, 15, 102329. [Google Scholar] [CrossRef]
- Qiu, Y.; Wen, H.; Wang, H.; Sun, W.; Li, G.; Li, S.; Wang, Y.; Zhai, J.; Zhan, Y.; Su, Y.; et al. Real-World Effectiveness and Safety of Nirmatrelvir-Ritonavir (Paxlovid)-Treated for COVID-19 Patients with Onset of More than 5 Days: A Retrospective Cohort Study. Front. Pharmacol. 2024, 15, 1401658. [Google Scholar] [CrossRef]
- Naqvi, A.A.T.; Fatima, K.; Mohammad, T.; Fatima, U.; Singh, I.K.; Singh, A.; Atif, S.M.; Hariprasad, G.; Hasan, G.M.; Hassan, M.I. Insights into SARS-CoV-2 Genome, Structure, Evolution, Pathogenesis and Therapies: Structural Genomics Approach. Biochim. Et. Biophys. Acta (BBA)—Mol. Basis Disease 2020, 1866, 165878. [Google Scholar] [CrossRef] [PubMed]
- Brant, A.C.; Tian, W.; Majerciak, V.; Yang, W.; Zheng, Z.-M. SARS-CoV-2: From Its Discovery to Genome Structure, Transcription, and Replication. Cell Biosci. 2021, 11, 136. [Google Scholar] [CrossRef] [PubMed]
- Polatoğlu, I.; Oncu-Oner, T.; Dalman, I.; Ozdogan, S. COVID-19 in Early 2023: Structure, Replication Mechanism, Variants of SARS-CoV-2, Diagnostic Tests, and Vaccine & Drug Development Studies. MedComm 2023, 4, e228. [Google Scholar] [CrossRef] [PubMed]
- Baggen, J.; Vanstreels, E.; Jansen, S.; Daelemans, D. Cellular Host Factors for SARS-CoV-2 Infection. Nat. Microbiol. 2021, 6, 1219–1232. [Google Scholar] [CrossRef]
- Yan, W.; Zheng, Y.; Zeng, X.; He, B.; Cheng, W. Structural Biology of SARS-CoV-2: Open the Door for Novel Therapies. Sig. Transduct. Target. Ther. 2022, 7, 26. [Google Scholar] [CrossRef]
- Guo, Y.-R.; Cao, Q.-D.; Hong, Z.-S.; Tan, Y.-Y.; Chen, S.-D.; Jin, H.-J.; Tan, K.-S.; Wang, D.-Y.; Yan, Y. The Origin, Transmission and Clinical Therapies on Coronavirus Disease 2019 (COVID-19) Outbreak—An Update on the Status. Mil. Med. Res. 2020, 7, 11. [Google Scholar] [CrossRef]
- Fehr, A.R.; Perlman, S. Coronaviruses: An Overview of Their Replication and Pathogenesis. Methods Mol. Biol. 2015, 1282, 1–23. [Google Scholar] [CrossRef]
- Subong, B.J.J.; Ozawa, T. Bio-Chemoinformatics-Driven Analysis of Nsp7 and Nsp8 Mutations and Their Effects on Viral Replication Protein Complex Stability. CIMB 2024, 46, 2598–2619. [Google Scholar] [CrossRef]
- Reshamwala, S.M.S.; Likhite, V.; Degani, M.S.; Deb, S.S.; Noronha, S.B. Mutations in SARS-CoV-2 Nsp7 and Nsp8 Proteins and Their Predicted Impact on Replication/Transcription Complex Structure. J. Med. Virol. 2021, 93, 4616–4619. [Google Scholar] [CrossRef]
- Gao, Y.; Yan, L.; Huang, Y.; Liu, F.; Zhao, Y.; Cao, L.; Wang, T.; Sun, Q.; Ming, Z.; Zhang, L.; et al. Structure of the RNA-Dependent RNA Polymerase from COVID-19 Virus. Science 2020, 368, 779–782. [Google Scholar] [CrossRef]
- Slanina, H.; Madhugiri, R.; Bylapudi, G.; Schultheiß, K.; Karl, N.; Gulyaeva, A.; Gorbalenya, A.E.; Linne, U.; Ziebuhr, J. Coronavirus Replication–Transcription Complex: Vital and Selective NMPylation of a Conserved Site in Nsp9 by the NiRAN-RdRp Subunit. Proc. Natl. Acad. Sci. USA 2021, 118, e2022310118. [Google Scholar] [CrossRef] [PubMed]
- Mickolajczyk, K.J.; Shelton, P.M.M.; Grasso, M.; Cao, X.; Warrington, S.E.; Aher, A.; Liu, S.; Kapoor, T.M. Force-Dependent Stimulation of RNA Unwinding by SARS-CoV-2 Nsp13 Helicase. Biophys. J. 2021, 120, 1020–1030. [Google Scholar] [CrossRef]
- Chen, J.; Wang, Q.; Malone, B.; Llewellyn, E.; Pechersky, Y.; Maruthi, K.; Eng, E.T.; Perry, J.K.; Campbell, E.A.; Shaw, D.E.; et al. Ensemble Cryo-EM Reveals Conformational States of the Nsp13 Helicase in the SARS-CoV-2 Helicase Replication–Transcription Complex. Nat. Struct. Mol. Biol. 2022, 29, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Chinthapatla, R.; Sotoudegan, M.; Srivastava, P.; Anderson, T.K.; Moustafa, I.M.; Passow, K.T.; Kennelly, S.A.; Moorthy, R.; Dulin, D.; Feng, J.Y.; et al. Interfering with Nucleotide Excision by the Coronavirus 3′-to-5′ Exoribonuclease. Nucleic Acids Res. 2023, 51, 315–336. [Google Scholar] [CrossRef]
- Niu, X.; Kong, F.; Hou, Y.J.; Wang, Q. Crucial Mutation in the Exoribonuclease Domain of Nsp14 of PEDV Leads to High Genetic Instability during Viral Replication. Cell Biosci. 2021, 11, 106. [Google Scholar] [CrossRef]
- Bouvet, M.; Lugari, A.; Posthuma, C.C.; Zevenhoven, J.C.; Bernard, S.; Betzi, S.; Imbert, I.; Canard, B.; Guillemot, J.-C.; Lécine, P.; et al. Coronavirus Nsp10, a Critical Co-Factor for Activation of Multiple Replicative Enzymes. J. Biol. Chem. 2014, 289, 25783–25796. [Google Scholar] [CrossRef]
- Yang, H.; Rao, Z. Structural Biology of SARS-CoV-2 and Implications for Therapeutic Development. Nat. Rev. Microbiol. 2021, 19, 685–700. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.; Cortese, M.; Winter, S.L.; Wachsmuth-Melm, M.; Neufeldt, C.J.; Cerikan, B.; Stanifer, M.L.; Boulant, S.; Bartenschlager, R.; Chlanda, P. SARS-CoV-2 Structure and Replication Characterized by in Situ Cryo-Electron Tomography. Nat. Commun. 2020, 11, 5885. [Google Scholar] [CrossRef]
- Malone, B.; Urakova, N.; Snijder, E.J.; Campbell, E.A. Structures and Functions of Coronavirus Replication–Transcription Complexes and Their Relevance for SARS-CoV-2 Drug Design. Nat. Rev. Mol. Cell Biol. 2022, 23, 21–39. [Google Scholar] [CrossRef]
- Wong, L.H.; Edgar, J.R.; Martello, A.; Ferguson, B.J.; Eden, E.R. Exploiting Connections for Viral Replication. Front. Cell Dev. Biol. 2021, 9, 640456. [Google Scholar] [CrossRef]
- Roingeard, P.; Eymieux, S.; Burlaud-Gaillard, J.; Hourioux, C.; Patient, R.; Blanchard, E. The Double-Membrane Vesicle (DMV): A Virus-Induced Organelle Dedicated to the Replication of SARS-CoV-2 and Other Positive-Sense Single-Stranded RNA Viruses. Cell. Mol. Life Sci. 2022, 79, 425. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, T.; Zhong, L.; Zhang, W.; Zhang, Y.; Yu, X.; Yuan, S.; Ni, T. Molecular Architecture of Coronavirus Double-Membrane Vesicle Pore Complex. Nature 2024, 633, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.M.; Chen, Y.-J.; Cho, W.J.; Tai, A.W.; Tsai, B. Reticulons Promote Formation of ER-Derived Double-Membrane Vesicles That Facilitate SARS-CoV-2 Replication. J. Cell Biol. 2023, 222, e202203060. [Google Scholar] [CrossRef] [PubMed]
- Hagemeijer, M.C.; Monastyrska, I.; Griffith, J.; Van Der Sluijs, P.; Voortman, J.; Van Bergen En Henegouwen, P.M.; Vonk, A.M.; Rottier, P.J.M.; Reggiori, F.; De Haan, C.A.M. Membrane Rearrangements Mediated by Coronavirus Nonstructural Proteins 3 and 4. Virology 2014, 458–459, 125–135. [Google Scholar] [CrossRef]
- Alexandersen, S.; Chamings, A.; Bhatta, T.R. SARS-CoV-2 Genomic and Subgenomic RNAs in Diagnostic Samples Are Not an Indicator of Active Replication. Nat. Commun. 2020, 11, 6059. [Google Scholar] [CrossRef]
- Telwatte, S.; Martin, H.A.; Marczak, R.; Fozouni, P.; Vallejo-Gracia, A.; Kumar, G.R.; Murray, V.; Lee, S.; Ott, M.; Wong, J.K.; et al. Novel RT-ddPCR Assays for Measuring the Levels of Subgenomic and Genomic SARS-CoV-2 Transcripts. Methods 2022, 201, 15–25. [Google Scholar] [CrossRef]
- Markov, P.V.; Ghafari, M.; Beer, M.; Lythgoe, K.; Simmonds, P.; Stilianakis, N.I.; Katzourakis, A. The Evolution of SARS-CoV-2. Nat. Rev. Microbiol. 2023, 21, 361–379. [Google Scholar] [CrossRef]
- Mack, A.H.; Menzies, G.; Southgate, A.; Jones, D.D.; Connor, T.R. A Proofreading Mutation with an Allosteric Effect Allows a Cluster of SARS-CoV-2 Viruses to Rapidly Evolve. Mol. Biol. Evol. 2023, 40, msad209. [Google Scholar] [CrossRef]
- Prydz, K.; Saraste, J. The Life Cycle and Enigmatic Egress of Coronaviruses. Mol. Microbiol. 2022, 117, 1308–1316. [Google Scholar] [CrossRef]
- Perdikari, T.M.; Murthy, A.C.; Ryan, V.H.; Watters, S.; Naik, M.T.; Fawzi, N.L. SARS-CoV-2 Nucleocapsid Protein Phase-separates with RNA and with Human hnRNPs. EMBO J. 2020, 39, e106478. [Google Scholar] [CrossRef]
- Pearson, G.J.; Mears, H.V.; Broncel, M.; Snijders, A.P.; Bauer, D.L.V.; Carlton, J.G. ER-Export and ARFRP1/AP-1–Dependent Delivery of SARS-CoV-2 Envelope to Lysosomes Controls Late Stages of Viral Replication. Sci. Adv. 2024, 10, eadl5012. [Google Scholar] [CrossRef]
- Gordon, D.E.; Hiatt, J.; Bouhaddou, M.; Rezelj, V.V.; Ulferts, S.; Braberg, H.; Jureka, A.S.; Obernier, K.; Guo, J.Z.; Batra, J.; et al. Comparative Host-Coronavirus Protein Interaction Networks Reveal Pan-Viral Disease Mechanisms. Science 2020, 370, eabe9403. [Google Scholar] [CrossRef]
- Smith, T.; Wang, S.; Kwon, Y.; Reid, A.A.; Robison, R.; Shen, P.; Willardson, B. Folding of the SARS-CoV-2 RNA Polymerase by the Cytosolic Chaperonin CCT. FASEB J. 2022, 36. [Google Scholar] [CrossRef]
- Alruwaili, M.; Armstrong, S.; Prince, T.; Erdmann, M.; Matthews, D.A.; Davidson, A.; Aljabr, W.; Hiscox, J.A. SARS-CoV-2 NSP12 Associates with the TRiC Complex and the P323L Substitution Is a Host Adaption. bioRxiv 2023. [Google Scholar]
- Liu, Q.; Zhao, S.; Hou, Y.; Ye, S.; Sha, T.; Su, Y.; Zhao, W.; Bao, Y.; Xue, Y.; Chen, H. Ongoing Natural Selection Drives the Evolution of SARS-CoV-2 Genomes. MedRxiv 2020. [Google Scholar]
- Shi, J.; Du, T.; Wang, J.; Tang, C.; Lei, M.; Yu, W.; Yang, Y.; Ma, Y.; Huang, P.; Chen, H.; et al. Aryl Hydrocarbon Receptor Is a Proviral Host Factor and a Candidate Pan-SARS-CoV-2 Therapeutic Target. Sci. Adv. 2023, 9, eadf0211. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Pu, Y.; Yuan, S.; Pache, L.; Churas, C.; Weston, S.; Riva, L.; Simons, L.M.; Cisneros, W.; Clausen, T.; et al. Global siRNA Screen Reveals Critical Human Host Factors of SARS-CoV-2 Multicycle Replication. bioRxiv 2024. [Google Scholar]
- Jockusch, S.; Tao, C.; Li, X.; Anderson, T.K.; Chien, M.; Kumar, S.; Russo, J.J.; Kirchdoerfer, R.N.; Ju, J. A Library of Nucleotide Analogues Terminate RNA Synthesis Catalyzed by Polymerases of Coronaviruses That Cause SARS and COVID-19. Antivir. Res. 2020, 180, 104857. [Google Scholar] [CrossRef]
- Li, G.; De Clercq, E. Therapeutic Options for the 2019 Novel Coronavirus (2019-nCoV). Nat. Rev. Drug Discov. 2020, 19, 149–150. [Google Scholar] [CrossRef]
- Beigel, J.H.; Tomashek, K.M.; Dodd, L.E.; Mehta, A.K.; Zingman, B.S.; Kalil, A.C.; Hohmann, E.; Chu, H.Y.; Luetkemeyer, A.; Kline, S.; et al. Remdesivir for the Treatment of Covid-19—Final Report. N. Engl. J. Med. 2020, 383, 1813–1826. [Google Scholar] [CrossRef]
- Olender, S.A.; Perez, K.K.; Go, A.S.; Balani, B.; Price-Haywood, E.G.; Shah, N.S.; Wang, S.; Walunas, T.L.; Swaminathan, S.; Slim, J.; et al. Remdesivir for Severe Coronavirus Disease 2019 (COVID-19) Versus a Cohort Receiving Standard of Care. Clin. Infect. Dis. 2021, 73, e4166–e4174. [Google Scholar] [CrossRef] [PubMed]
- Ogando, N.S.; Zevenhoven-Dobbe, J.C.; Van Der Meer, Y.; Bredenbeek, P.J.; Posthuma, C.C.; Snijder, E.J. The Enzymatic Activity of the Nsp14 Exoribonuclease Is Critical for Replication of MERS-CoV and SARS-CoV-2. J. Virol. 2020, 94, e01246-20. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.-Y.; Lahser, F.; Warren, C.; He, X.; Murray, E.; Wang, D. The Impact of SARS-CoV-2 Nsp14 Proofreading on Nucleoside Antiviral Activity: Insights from Genetic and Pharmacological Investigations. bioRxiv 2024. [Google Scholar]
- Lee, C.-C.; Hsieh, C.-C.; Ko, W.-C. Molnupiravir—A Novel Oral Anti-SARS-CoV-2 Agent. Antibiotics 2021, 10, 1294. [Google Scholar] [CrossRef]
- Kabinger, F.; Stiller, C.; Schmitzová, J.; Dienemann, C.; Kokic, G.; Hillen, H.S.; Höbartner, C.; Cramer, P. Mechanism of Molnupiravir-Induced SARS-CoV-2 Mutagenesis. Nat. Struct. Mol. Biol. 2021, 28, 740–746. [Google Scholar] [CrossRef]
- Tian, L.; Pang, Z.; Li, M.; Lou, F.; An, X.; Zhu, S.; Song, L.; Tong, Y.; Fan, H.; Fan, J. Molnupiravir and Its Antiviral Activity Against COVID-19. Front. Immunol. 2022, 13, 855496. [Google Scholar] [CrossRef]
- Malone, B.; Campbell, E.A. Molnupiravir: Coding for Catastrophe. Nat. Struct. Mol. Biol. 2021, 28, 706–708. [Google Scholar] [CrossRef]
- Masyeni, S.; Iqhrammullah, M.; Frediansyah, A.; Nainu, F.; Tallei, T.; Emran, T.B.; Ophinni, Y.; Dhama, K.; Harapan, H. Molnupiravir: A Lethal Mutagenic Drug against Rapidly Mutating Severe Acute Respiratory Syndrome Coronavirus 2—A Narrative Review. J. Med. Virol. 2022, 94, 3006–3016. [Google Scholar] [CrossRef]
- Caraco, Y.; Crofoot, G.E.; Moncada, P.A.; Galustyan, A.N.; Musungaie, D.B.; Payne, B.; Kovalchuk, E.; Gonzalez, A.; Brown, M.L.; Williams-Diaz, A.; et al. Phase 2/3 Trial of Molnupiravir for Treatment of Covid-19 in Nonhospitalized Adults. NEJM Evid. 2022, 1. [Google Scholar] [CrossRef]
- Strizki, J.M.; Gaspar, J.M.; Howe, J.A.; Hutchins, B.; Mohri, H.; Nair, M.S.; Kinek, K.C.; McKenna, P.; Goh, S.L.; Murgolo, N. Molnupiravir Maintains Antiviral Activity against SARS-CoV-2 Variants and Exhibits a High Barrier to the Development of Resistance. Antimicrob. Agents Chemother. 2024, 68, e00953-23. [Google Scholar] [CrossRef]
- Erdem, H.A.; Korkmaz Ekren, P.; Çağlayan, D.; Işikgöz Taşbakan, M.; Yamazhan, T.; Taşbakan, M.S.; Sayiner, A.; Gökengi̇n, D. Treatment of SARS-CoV-2 Pneumonia with Favipiravir: Early Results from the Ege University Cohort, Turkey. Turk. J. Med. Sci. 2021, 51, 912–920. [Google Scholar] [CrossRef] [PubMed]
- Ghasemnejad-Berenji, M.; Pashapour, S. Favipiravir and COVID-19: A Simplified Summary. Drug Res. 2021, 71, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Manabe, T.; Kambayashi, D.; Akatsu, H.; Kudo, K. Favipiravir for the Treatment of Patients with COVID-19: A Systematic Review and Meta-Analysis. BMC Infect. Dis. 2021, 21, 489. [Google Scholar] [CrossRef]
- Juul, S.; Nielsen, E.E.; Feinberg, J.; Siddiqui, F.; Jørgensen, C.K.; Barot, E.; Holgersson, J.; Nielsen, N.; Bentzer, P.; Veroniki, A.A.; et al. Interventions for Treatment of COVID-19: Second Edition of a Living Systematic Review with Meta-Analyses and Trial Sequential Analyses (The LIVING Project). PLoS ONE 2021, 16, e0248132. [Google Scholar] [CrossRef]
- Shrestha, D.B.; Budhathoki, P.; Khadka, S.; Shah, P.B.; Pokharel, N.; Rashmi, P. Favipiravir versus Other Antiviral or Standard of Care for COVID-19 Treatment: A Rapid Systematic Review and Meta-Analysis. Virol. J. 2020, 17, 141. [Google Scholar] [CrossRef]
- Tong, S.; Su, Y.; Yu, Y.; Wu, C.; Chen, J.; Wang, S.; Jiang, J. Ribavirin Therapy for Severe COVID-19: A Retrospective Cohort Study. Int. J. Antimicrob. Agents 2020, 56, 106114. [Google Scholar] [CrossRef] [PubMed]
- Messina, E.; Danise, A.; Ferrari, G.; Andolina, A.; Chiurlo, M.; Razanakolona, M.; Barakat, M.; Israel, R.J.; Castagna, A. Ribavirin Aerosol in the Treatment of SARS-CoV-2: A Case Series. Infect. Dis. Ther. 2021, 10, 2791–2804. [Google Scholar] [CrossRef]
- Qian, H.; Wang, Y.; Zhang, M.; Xie, Y.; Wu, Q.; Liang, L.; Cao, Y.; Duan, H.; Tian, G.; Ma, J.; et al. Safety, Tolerability, and Pharmacokinetics of VV116, an Oral Nucleoside Analog against SARS-CoV-2, in Chinese Healthy Subjects. Acta Pharmacol. Sin. 2022, 43, 3130–3138. [Google Scholar] [CrossRef]
- Xiao, N.; Huang, X.; Kang, X.; Zang, W.; Li, B.; Kiselev, S. The Safety and Efficacy of Oral Antiviral Drug VV116 for Treatment of COVID-19: A Systematic Review. Medicine 2023, 102, e34105. [Google Scholar] [CrossRef]
- McCarthy, M.W. VV116 as a Potential Treatment for COVID-19. Expert Opin. Pharmacother. 2023, 24, 675–678. [Google Scholar] [CrossRef]
- Hammond, J.; Leister-Tebbe, H.; Gardner, A.; Abreu, P.; Bao, W.; Wisemandle, W.; Baniecki, M.; Hendrick, V.M.; Damle, B.; Simón-Campos, A.; et al. Oral Nirmatrelvir for High-Risk, Nonhospitalized Adults with Covid-19. N. Engl. J. Med. 2022, 386, 1397–1408. [Google Scholar] [CrossRef]
- Marzi, M.; Vakil, M.K.; Bahmanyar, M.; Zarenezhad, E. Paxlovid: Mechanism of Action, Synthesis, and In Silico Study. BioMed Res. Int. 2022, 2022, 7341493. [Google Scholar] [CrossRef] [PubMed]
- Marzolini, C.; Kuritzkes, D.R.; Marra, F.; Boyle, A.; Gibbons, S.; Flexner, C.; Pozniak, A.; Boffito, M.; Waters, L.; Burger, D.; et al. Recommendations for the Management of Drug–Drug Interactions Between the COVID-19 Antiviral Nirmatrelvir/Ritonavir (Paxlovid) and Comedications. Clin. Pharma Ther. 2022, 112, 1191–1200. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Gammeltoft, K.A.; Ryberg, L.A.; Pham, L.V.; Fahnøe, U.; Binderup, A.; Hernandez, C.R.D.; Offersgaard, A.; Fernandez-Antunez, C.; Peters, G.H.J.; et al. Nirmatrelvir Resistant SARS-CoV-2 Variants with High Fitness in Vitro. Sci. Adv. 2022, 51, eadd7197. [Google Scholar] [CrossRef] [PubMed]
- Iketani, S.; Mohri, H.; Culbertson, B.; Hong, S.J.; Duan, Y.; Luck, M.I.; Annavajhala, M.K.; Guo, Y.; Sheng, Z.; Uhlemann, A.-C.; et al. Multiple Pathways for SARS-CoV-2 Resistance to Nirmatrelvir. Nature 2022, 613, 558–564. [Google Scholar] [CrossRef]
- Costacurta, F.; Dodaro, A.; Bante, D.; Schöppe, H.; Peng, J.-Y.; Sprenger, B.; He, X.; Moghadasi, S.A.; Egger, L.M.; Fleischmann, J.; et al. A Comprehensive Study of SARS-CoV-2 Main Protease (Mpro) Inhibitor-Resistant Mutants Selected in a VSV-Based System. PLoS Pathog. 2024, 20, e1012522. [Google Scholar] [CrossRef]
- Cao, B.; Wang, Y.; Wen, D.; Liu, W.; Wang, J.; Fan, G.; Ruan, L.; Song, B.; Cai, Y.; Wei, M.; et al. A Trial of Lopinavir–Ritonavir in Adults Hospitalized with Severe Covid-19. N. Engl. J. Med. 2020, 382, 1787–1799. [Google Scholar] [CrossRef]
- Kuroda, T.; Nobori, H.; Fukao, K.; Baba, K.; Matsumoto, K.; Yoshida, S.; Tanaka, Y.; Watari, R.; Oka, R.; Kasai, Y.; et al. Efficacy Comparison of 3CL Protease Inhibitors Ensitrelvir and Nirmatrelvir against SARS-CoV-2 in Vitro and in Vivo. J. Antimicrob. Chemother. 2023, 78, 946–952. [Google Scholar] [CrossRef]
- Garnsey, M.R.; Robinson, M.C.; Nguyen, L.T.; Cardin, R.; Tillotson, J.; Mashalidis, E.; Yu, A.; Aschenbrenner, L.; Balesano, A.; Behzadi, A.; et al. Discovery of SARS-CoV-2 Papain-like Protease (PLpro ) Inhibitors with Efficacy in a Murine Infection Model. Sci. Adv. 2024, 10, eado4288. [Google Scholar] [CrossRef]
- Brunotte, L.; Zheng, S.; Mecate-Zambrano, A.; Tang, J.; Ludwig, S.; Rescher, U.; Schloer, S. Combination Therapy with Fluoxetine and the Nucleoside Analog GS-441524 Exerts Synergistic Antiviral Effects against Different SARS-CoV-2 Variants In Vitro. Pharmaceutics 2021, 13, 1400. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Liu, M.; Wang, C.; Xu, W.; Lan, Q.; Feng, S.; Qi, F.; Bao, L.; Du, L.; Liu, S.; et al. Inhibition of SARS-CoV-2 (Previously 2019-nCoV) Infection by a Highly Potent Pan-Coronavirus Fusion Inhibitor Targeting Its Spike Protein That Harbors a High Capacity to Mediate Membrane Fusion. Cell Res. 2020, 30, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.; Chen, X.; Wu, J.; Duan, X.; Men, K. Small Molecules in the Treatment of COVID-19. Signal Transduct. Target. Ther. 2022, 7, 387. [Google Scholar] [CrossRef]
- Batool, S.; Chokkakula, S.; Jeong, J.H.; Baek, Y.H.; Song, M.-S. SARS-CoV-2 Drug Resistance and Therapeutic Approaches. Heliyon 2025, 11, e41980. [Google Scholar] [CrossRef]
- Rahmah, L.; Abarikwu, S.O.; Arero, A.G.; Essouma, M.; Jibril, A.T.; Fal, A.; Flisiak, R.; Makuku, R.; Marquez, L.; Mohamed, K.; et al. Oral Antiviral Treatments for COVID-19: Opportunities and Challenges. Pharmacol. Rep. 2022, 74, 1255–1278. [Google Scholar] [CrossRef]
- Gidari, A.; Sabbatini, S.; Schiaroli, E.; Bastianelli, S.; Pierucci, S.; Busti, C.; Saraca, L.M.; Capogrossi, L.; Pasticci, M.B.; Francisci, D. Synergistic Activity of Remdesivir–Nirmatrelvir Combination on a SARS-CoV-2 In Vitro Model and a Case Report. Viruses 2023, 15, 1577. [Google Scholar] [CrossRef]
- Gidari, A.; Sabbatini, S.; Schiaroli, E.; Bastianelli, S.; Pierucci, S.; Busti, C.; Comez, L.; Libera, V.; Macchiarulo, A.; Paciaroni, A.; et al. The Combination of Molnupiravir with Nirmatrelvir or GC376 Has a Synergic Role in the Inhibition of SARS-CoV-2 Replication In Vitro. Microorganisms 2022, 10, 1475. [Google Scholar] [CrossRef] [PubMed]
- Woodall, M.; Ellis, S.; Zhang, S.; Kembou-Ringert, J.; Kite, K.-A.; Buggiotti, L.; Jacobs, A.I.; Agyeman, A.A.; Masonou, T.; Palor, M.; et al. Efficient in Vitro Assay for Evaluating Drug Efficacy and Synergy against Emerging SARS-CoV-2 Strains. Antimicrob. Agents Chemother. 2025, 69, e01233-24. [Google Scholar] [CrossRef]
- García-Crespo, C.; De Ávila, A.I.; Gallego, I.; Soria, M.E.; Durán-Pastor, A.; Somovilla, P.; Martínez-González, B.; Muñoz-Flores, J.; Mínguez, P.; Salar-Vidal, L.; et al. Synergism between Remdesivir and Ribavirin Leads to SARS-CoV-2 Extinction in Cell Culture. Br. J. Pharmacol. 2024, 181, 2636–2654. [Google Scholar] [CrossRef]
- Robson, F.; Khan, K.S.; Le, T.K.; Paris, C.; Demirbag, S.; Barfuss, P.; Rocchi, P.; Ng, W.-L. Coronavirus RNA Proofreading: Molecular Basis and Therapeutic Targeting. Mol. Cell 2020, 79, 710–727. [Google Scholar] [CrossRef]
- Do, T.N.D.; Abdelnabi, R.; Boda, B.; Constant, S.; Neyts, J.; Jochmans, D. The Triple Combination of Remdesivir (GS-441524), Molnupiravir and Ribavirin Is Highly Efficient in Inhibiting Coronavirus Replication in Human Nasal Airway Epithelial Cell Cultures and in a Hamster Infection Model. Antivir. Res. 2024, 231, 105994. [Google Scholar] [CrossRef] [PubMed]
- Mayor, J.; Engler, O.; Rothenberger, S. Antiviral Efficacy of Ribavirin and Favipiravir against Hantaan Virus. Microorganisms 2021, 9, 1306. [Google Scholar] [CrossRef]
- Wagoner, J.; Herring, S.; Hsiang, T.-Y.; Ianevski, A.; Biering, S.B.; Xu, S.; Hoffmann, M.; Pöhlmann, S.; Gale, M.; Aittokallio, T.; et al. Combinations of Host- and Virus-Targeting Antiviral Drugs Confer Synergistic Suppression of SARS-CoV-2. Microbiol. Spectr. 2022, 10, e03331-22. [Google Scholar] [CrossRef]
- White, J.M.; Schiffer, J.T.; Bender Ignacio, R.A.; Xu, S.; Kainov, D.; Ianevski, A.; Aittokallio, T.; Frieman, M.; Olinger, G.G.; Polyak, S.J. Drug Combinations as a First Line of Defense against Coronaviruses and Other Emerging Viruses. mBio 2021, 12, e03347-21. [Google Scholar] [CrossRef]
- Jeong, J.H.; Chokkakula, S.; Min, S.C.; Kim, B.K.; Choi, W.-S.; Oh, S.; Yun, Y.S.; Kang, D.H.; Lee, O.-J.; Kim, E.-G.; et al. Combination Therapy with Nirmatrelvir and Molnupiravir Improves the Survival of SARS-CoV-2 Infected Mice. Antivir. Res. 2022, 208, 105430. [Google Scholar] [CrossRef]
- Mahendran, T.R.; Cynthia, B.; Thevendran, R.; Maheswaran, S. Prospects of Innovative Therapeutics in Combating the COVID-19 Pandemic. Mol. Biotechnol. 2024, 67, 2598–2606. [Google Scholar] [CrossRef]
- Abdulaziz, L.; Elhadi, E.; Abdallah, E.A.; Alnoor, F.A.; Yousef, B.A. Antiviral Activity of Approved Antibacterial, Antifungal, Antiprotozoal and Anthelmintic Drugs: Chances for Drug Repurposing for Antiviral Drug Discovery. JEP 2022, 14, 97–115. [Google Scholar] [CrossRef] [PubMed]
- Hamid, A.; Mäser, P.; Mahmoud, A.B. Drug Repurposing in the Chemotherapy of Infectious Diseases. Molecules 2024, 29, 635. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, B.; Thakur, S.S. Remdesivir and Its Combination With Repurposed Drugs as COVID-19 Therapeutics. Front. Immunol. 2022, 13, 830990. [Google Scholar] [CrossRef]
- Mahdi, M.; Hermán, L.; Réthelyi, J.M.; Bálint, B.L. Potential Role of the Antidepressants Fluoxetine and Fluvoxamine in the Treatment of COVID-19. IJMS 2022, 23, 3812. [Google Scholar] [CrossRef]
- Al-Hajeri, H.; Baroun, F.; Abutiban, F.; Al-Mutairi, M.; Ali, Y.; Alawadhi, A.; Albasri, A.; Aldei, A.; AlEnizi, A.; Alhadhood, N.; et al. Therapeutic Role of Immunomodulators during the COVID-19 Pandemic– a Narrative Review. Postgrad. Med. 2022, 134, 160–179. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, S.C.J.; Tse, C.L.Y.; Burry, L.; Dresser, L.D. Baricitinib: A Review of Pharmacology, Safety, and Emerging Clinical Experience in COVID-19. Pharmacotherapy 2020, 40, 843–856. [Google Scholar] [CrossRef]
- Frediansyah, A.; Tiwari, R.; Sharun, K.; Dhama, K.; Harapan, H. Antivirals for COVID-19: A Critical Review. Clin. Epidemiol. Glob. Health 2021, 9, 90–98. [Google Scholar] [CrossRef]
- Villa, T.G.; Abril, A.G.; Sánchez, S.; De Miguel, T.; Sánchez-Pérez, A. Animal and Human RNA Viruses: Genetic Variability and Ability to Overcome Vaccines. Arch. Microbiol. 2021, 203, 443–464. [Google Scholar] [CrossRef]
- Checkmahomed, L.; Carbonneau, J.; Du Pont, V.; Riola, N.C.; Perry, J.K.; Li, J.; Paré, B.; Simpson, S.M.; Smith, M.A.; Porter, D.P.; et al. In Vitro Selection of Remdesivir-Resistant SARS-CoV-2 Demonstrates High Barrier to Resistance. Antimicrob. Agents Chemother. 2022, 66, e00198-22. [Google Scholar] [CrossRef] [PubMed]
- Szemiel, A.M.; Merits, A.; Orton, R.J.; MacLean, O.A.; Pinto, R.M.; Wickenhagen, A.; Lieber, G.; Turnbull, M.L.; Wang, S.; Furnon, W.; et al. In Vitro Selection of Remdesivir Resistance Suggests Evolutionary Predictability of SARS-CoV-2. PLoS Pathog. 2021, 17, e1009929. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, T.; Hisner, R.; Donovan-Banfield, I.; Hartman, H.; Løchen, A.; Peacock, T.P.; Ruis, C. A Molnupiravir-Associated Mutational Signature in Global SARS-CoV-2 Genomes. Nature 2023, 623, 594–600. [Google Scholar] [CrossRef]
- Zhao, Y.; He, G.; Huang, W. A Novel Model of Molnupiravir against SARS-CoV-2 Replication: Accumulated RNA Mutations to Induce Error Catastrophe. Sig Transduct. Target. Ther. 2021, 6, 410. [Google Scholar] [CrossRef]
- Lee, J.T.; Yang, Q.; Gribenko, A.; Perrin, B.S.; Zhu, Y.; Cardin, R.; Liberator, P.A.; Anderson, A.S.; Hao, L. Genetic Surveillance of SARS-CoV-2 Mpro Reveals High Sequence and Structural Conservation Prior to the Introduction of Protease Inhibitor Paxlovid. mBio 2022, 13, e00869-22. [Google Scholar] [CrossRef]
- Peluso, M.J.; Deeks, S.G. Mechanisms of Long COVID and the Path toward Therapeutics. Cell 2024, 187, 5500–5529. [Google Scholar] [CrossRef]
- Lupașcu (Moisi), R.E.; Ilie, M.I.; Velescu, B.Ș.; Udeanu, D.I.; Sultana, C.; Ruță, S.; Arsene, A.L. COVID-19-Current Therapeutical Approaches and Future Perspectives. Processes 2022, 10, 1053. [Google Scholar] [CrossRef]
- Chala, B.; Tilaye, T.; Waktole, G. Re-Emerging COVID-19: Controversy of Its Zoonotic Origin, Risks of Severity of Reinfection and Management. IJGM 2023, 16, 4307–4319. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Zheng, M.; Gao, Y.; Lin, S.; Zhang, X.; Chen, C.; Zhu, H.; Sun, W.; Zhang, Y. Overview of Host-Directed Antiviral Targets for Future Research and Drug Development. Acta Pharm. Sin. B 2025, 15, 1723–1751. [Google Scholar] [CrossRef]
- Schuller, M.; Zarganes-Tzitzikas, T.; Bennett, J.; De Cesco, S.; Fearon, D.; Von Delft, F.; Fedorov, O.; Brennan, P.E.; Ahel, I. Discovery and Development Strategies for SARS-CoV-2 NSP3 Macrodomain Inhibitors. Pathogens 2023, 12, 324. [Google Scholar] [CrossRef]
- Petushkova, A.I.; Zamyatnin, A.A. Papain-Like Proteases as Coronaviral Drug Targets: Current Inhibitors, Opportunities, and Limitations. Pharmaceuticals 2020, 13, 277. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, M.; Damalanka, V.C.; Tartell, M.A.; Chung, D.H.; Lourenço, A.L.; Pwee, D.; Mayer Bridwell, A.E.; Hoffmann, M.; Voss, J.; Karmakar, P.; et al. A Novel Class of TMPRSS2 Inhibitors Potently Block SARS-CoV-2 and MERS-CoV Viral Entry and Protect Human Epithelial Lung Cells. Proc. Natl. Acad. Sci. USA 2021, 118, e2108728118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yu, W. Heat Shock Proteins and Viral Infection. Front. Immunol. 2022, 13, 947789. [Google Scholar] [CrossRef]
- Wickramaratne, A.C.; Wickner, S.; Kravats, A.N. Hsp90, a Team Player in Protein Quality Control and the Stress Response in Bacteria. Microbiol. Mol. Biol. Rev. 2024, 88, e00176-22. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, T.; Song, M.; Jin, M.; Liu, S.; Guo, K.; Zhang, Y. Rab1b-GBF1-ARFs Mediated Intracellular Trafficking Is Required for Classical Swine Fever Virus Replication in Swine Umbilical Vein Endothelial Cells. Vet. Microbiol. 2020, 246, 108743. [Google Scholar] [CrossRef]
- Chikhoune, L.; Poggi, C.; Moreau, J.; Dubucquoi, S.; Hachulla, E.; Collet, A.; Launay, D. JAK Inhibitors (JAKi): Mechanisms of Action and Perspectives in Systemic and Autoimmune Diseases. La. Rev. De Médecine Interne 2025, 46, 89–106. [Google Scholar] [CrossRef]
- Sodeifian, F.; Nikfarjam, M.; Kian, N.; Mohamed, K.; Rezaei, N. The Role of Type I Interferon in the Treatment of COVID-19. J. Med. Virol. 2022, 94, 63–81. [Google Scholar] [CrossRef] [PubMed]
- Savan, R.; Gale, M. Innate Immunity and Interferon in SARS-CoV-2 Infection Outcome. Immunity 2023, 56, 1443–1450. [Google Scholar] [CrossRef] [PubMed]
- Floresta, G.; Zagni, C.; Gentile, D.; Patamia, V.; Rescifina, A. Artificial Intelligence Technologies for COVID-19 De Novo Drug Design. IJMS 2022, 23, 3261. [Google Scholar] [CrossRef]
- Chodera, J.; Lee, A.A.; London, N.; Von Delft, F. Crowdsourcing Drug Discovery for Pandemics. Nat. Chem. 2020, 12, 581. [Google Scholar] [CrossRef]
- Faisal, S.; Badshah, S.L.; Kubra, B.; Sharaf, M.; Emwas, A.-H.; Jaremko, M.; Abdalla, M. Computational Study of SARS-CoV-2 RNA Dependent RNA Polymerase Allosteric Site Inhibition. Molecules 2021, 27, 223. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Yang, S.; He, S.; Li, F. AI Drug Discovery Tools and Analysis Technology: New Methods Aid in Studying the Compatibility of Traditional Chinese Medicine. Pharmacol. Res. Mod. Chin. Med. 2025, 14, 100566. [Google Scholar] [CrossRef]
- Maghsoudi, S.; Taghavi Shahraki, B.; Rameh, F.; Nazarabi, M.; Fatahi, Y.; Akhavan, O.; Rabiee, M.; Mostafavi, E.; Lima, E.C.; Saeb, M.R.; et al. A Review on Computer-aided Chemogenomics and Drug Repositioning for Rational COVID-19 Drug Discovery. Chem. Biol. Drug Des. 2022, 100, 699–721. [Google Scholar] [CrossRef]
- Mottaqi, M.S.; Mohammadipanah, F.; Sajedi, H. Contribution of Machine Learning Approaches in Response to SARS-CoV-2 Infection. Inform. Med. Unlocked 2021, 23, 100526. [Google Scholar] [CrossRef]
- Rajput, A.; Thakur, A.; Mukhopadhyay, A.; Kamboj, S.; Rastogi, A.; Gautam, S.; Jassal, H.; Kumar, M. Prediction of Repurposed Drugs for Coronaviruses Using Artificial Intelligence and Machine Learning. Comput. Struct. Biotechnol. J. 2021, 19, 3133–3148. [Google Scholar] [CrossRef]
- Huchting, J. Targeting Viral Genome Synthesis as Broad-Spectrum Approach against RNA Virus Infections. Antivir. Chem. Chemother. 2020, 28, 204020662097678. [Google Scholar] [CrossRef]
- Tolksdorf, B.; Heinze, J.; Niemeyer, D.; Röhrs, V.; Berg, J.; Drosten, C.; Kurreck, J. Development of a Highly Stable, Active Small Interfering RNA with Broad Activity against SARS-CoV Viruses. Antivir. Res. 2024, 226, 105879. [Google Scholar] [CrossRef] [PubMed]
- Abbott, T.R.; Dhamdhere, G.; Liu, Y.; Lin, X.; Goudy, L.; Zeng, L.; Chemparathy, A.; Chmura, S.; Heaton, N.S.; Debs, R.; et al. Development of CRISPR as an Antiviral Strategy to Combat SARS-CoV-2 and Influenza. Cell 2020, 181, 865–876.e12. [Google Scholar] [CrossRef]
- Li, T.; Yang, Y.; Qi, H.; Cui, W.; Zhang, L.; Fu, X.; He, X.; Liu, M.; Li, P.; Yu, T. CRISPR/Cas9 Therapeutics: Progress and Prospects. Signal Transduct. Target. Ther. 2023, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Najafi, S.; Tan, S.C.; Aghamiri, S.; Raee, P.; Ebrahimi, Z.; Jahromi, Z.K.; Rahmati, Y.; Sadri Nahand, J.; Piroozmand, A.; Jajarmi, V.; et al. Therapeutic Potentials of CRISPR-Cas Genome Editing Technology in Human Viral Infections. Biomed. Pharmacother. 2022, 148, 112743. [Google Scholar] [CrossRef] [PubMed]
- Zhong, G.; Chang, X.; Xie, W.; Zhou, X. Targeted Protein Degradation: Advances in Drug Discovery and Clinical Practice. Signal Transduct. Target. Ther. 2024, 9, 308. [Google Scholar] [CrossRef]
- Kubryń, N.; Fijałkowski, Ł.; Nowaczyk, J.; Jamil, A.; Nowaczyk, A. PROTAC Technology as a New Tool for Modern Pharmacotherapy. Molecules 2025, 30, 2123. [Google Scholar] [CrossRef]
- Liu, Z.; Hu, M.; Yang, Y.; Du, C.; Zhou, H.; Liu, C.; Chen, Y.; Fan, L.; Ma, H.; Gong, Y.; et al. An Overview of PROTACs: A Promising Drug Discovery Paradigm. Mol. Biomed. 2022, 3, 46. [Google Scholar] [CrossRef]
- Reboud-Ravaux, M.; El Amri, C. COVID-19 Therapies: Protease Inhibitions and Novel Degrader Strategies. Front. Drug Discov. 2022, 2, 892057. [Google Scholar] [CrossRef]
- Low, Z.Y.; Zabidi, N.Z.; Yip, A.J.W.; Puniyamurti, A.; Chow, V.T.K.; Lal, S.K. SARS-CoV-2 Non-Structural Proteins and Their Roles in Host Immune Evasion. Viruses 2022, 14, 1991. [Google Scholar] [CrossRef]
- Alugubelli, Y.R.; Xiao, J.; Khatua, K.; Kumar, S.; Sun, L.; Ma, Y.; Ma, X.R.; Vulupala, V.R.; Atla, S.; Blankenship, L.R.; et al. Discovery of First-in-Class PROTAC Degraders of SARS-CoV-2 Main Protease. J. Med. Chem. 2024, 67, 6495–6507. [Google Scholar] [CrossRef]
- Pan, B.; Mountford, S.J.; Kiso, M.; Anderson, D.E.; Papadakis, G.; Jarman, K.E.; Tilmanis, D.R.; Maher, B.; Tran, T.; Shortt, J.; et al. Targeted Protein Degraders of SARS-CoV-2 Mpro Are More Active than Enzymatic Inhibition Alone with Activity against Nirmatrelvir Resistant Virus. Commun. Med. 2025, 5, 140. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhao, J.; Zhong, K.; Tong, A.; Jia, D. Targeted Protein Degradation: Mechanisms, Strategies and Application. Signal Transduct. Target. Ther. 2022, 7, 113. [Google Scholar] [CrossRef] [PubMed]
- Von Delft, A.; Hall, M.D.; Kwong, A.D.; Purcell, L.A.; Saikatendu, K.S.; Schmitz, U.; Tallarico, J.A.; Lee, A.A. Accelerating Antiviral Drug Discovery: Lessons from COVID-19. Nat. Rev. Drug Discov. 2023, 22, 585–603. [Google Scholar] [CrossRef]
- Schreiber, A.; Ludwig, S. Host-Targeted Antivirals against SARS-CoV-2 in Clinical Development—Prospect or Disappointment? Antivir. Res. 2025, 235, 106101. [Google Scholar] [CrossRef]
- Seley-Radtke, K.L.; Thames, J.E.; Waters, C.D. Broad Spectrum Antiviral Nucleosides—Our Best Hope for the Future. In Annual Reports in Medicinal Chemistry; Elsevier: Amsterdam, The Netherlands, 2021; Volume 57, pp. 109–132. ISBN 978-0-323-91511-3. [Google Scholar]
- Kirchdoerfer, R.N.; Ward, A.B. Structure of the SARS-CoV Nsp12 Polymerase Bound to Nsp7 and Nsp8 Co-Factors. Nat. Commun. 2019, 10, 2342. [Google Scholar] [CrossRef]
- Xiong, R.; Zhang, L.; Li, S.; Sun, Y.; Ding, M.; Wang, Y.; Zhao, Y.; Wu, Y.; Shang, W.; Jiang, X.; et al. Novel and Potent Inhibitors Targeting DHODH Are Broad-Spectrum Antivirals against RNA Viruses Including Newly-Emerged Coronavirus SARS-CoV-2. Protein Cell 2020, 11, 723–739. [Google Scholar] [CrossRef]
- Sievers, B.L.; Cheng, M.T.K.; Csiba, K.; Meng, B.; Gupta, R.K. SARS-CoV-2 and Innate Immunity: The Good, the Bad, and the “Goldilocks”. Cell Mol. Immunol. 2023, 21, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Italiya, J.; Bhavsar, T.; Černý, J. Assessment and Strategy Development for SARS-CoV-2 Screening in Wildlife: A Review. Vet. World 2023, 16, 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- Karim, M.; Lo, C.-W.; Einav, S. Preparing for the next Viral Threat with Broad-Spectrum Antivirals. J. Clin. Investig. 2023, 133, e170236. [Google Scholar] [CrossRef]
- Barghash, R.F.; Gemmati, D.; Awad, A.M.; Elbakry, M.M.M.; Tisato, V.; Awad, K.; Singh, A.V. Navigating the COVID-19 Therapeutic Landscape: Unveiling Novel Perspectives on FDA-Approved Medications, Vaccination Targets, and Emerging Novel Strategies. Molecules 2024, 29, 5564. [Google Scholar] [CrossRef]
- Aligolighasemabadi, F.; Bakinowska, E.; Kiełbowski, K.; Sadeghdoust, M.; Coombs, K.M.; Mehrbod, P.; Ghavami, S. Autophagy and Respiratory Viruses: Mechanisms, Viral Exploitation, and Therapeutic Insights. Cells 2025, 14, 418. [Google Scholar] [CrossRef] [PubMed]
- Farkaš, B.; Minneci, M.; Misevicius, M.; Rozas, I. A Tale of Two Proteases: MPro and TMPRSS2 as Targets for COVID-19 Therapies. Pharmaceuticals 2023, 16, 834. [Google Scholar] [CrossRef] [PubMed]
- Tani, H. Recent Advances and Prospects in RNA Drug Development. IJMS 2024, 25, 12284. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Hilles, A.R.; Almurisi, S.H.; Bhatia, A.; Mahmood, S. Micro and Nano-Carriers-Based Pulmonary Drug Delivery System: Their Current Updates, Challenges, and Limitations—A Review. JCIS Open 2023, 12, 100095. [Google Scholar] [CrossRef]
- Rehman, A.U.; Li, M.; Wu, B.; Ali, Y.; Rasheed, S.; Shaheen, S.; Liu, X.; Luo, R.; Zhang, J. Role of Artificial Intelligence in Revolutionizing Drug Discovery. Fundam. Res. 2024, 5, 1273–1287. [Google Scholar] [CrossRef]
- Li, G.; Hilgenfeld, R.; Whitley, R.; De Clercq, E. Therapeutic Strategies for COVID-19: Progress and Lessons Learned. Nat. Rev. Drug Discov. 2023, 22, 449–475. [Google Scholar] [CrossRef]
- Paunovska, K.; Loughrey, D.; Dahlman, J.E. Drug Delivery Systems for RNA Therapeutics. Nat. Rev. Genet. 2022, 23, 265–280. [Google Scholar] [CrossRef]
- De Jesús-González, L.A.; León-Juárez, M.; Lira-Hernández, F.I.; Rivas-Santiago, B.; Velázquez-Cervantes, M.A.; Méndez-Delgado, I.M.; Macías-Guerrero, D.I.; Hernández-Castillo, J.; Hernández-Rodríguez, X.; Calderón-Sandate, D.N.; et al. Advances and Challenges in Antiviral Development for Respiratory Viruses. Pathogens 2024, 14, 20. [Google Scholar] [CrossRef]
- Huang, Y.; Guo, X.; Wu, Y.; Chen, X.; Feng, L.; Xie, N.; Shen, G. Nanotechnology’s Frontier in Combatting Infectious and Inflammatory Diseases: Prevention and Treatment. Signal Transduct. Target. Ther. 2024, 9, 34. [Google Scholar] [CrossRef]
- Fu, C.; Chen, Q. The Future of Pharmaceuticals: Artificial Intelligence in Drug Discovery and Development. J. Pharm. Anal. 2025, 101248. [Google Scholar] [CrossRef]
- Chakravarty, A.; Yang, P.L. Targeted Protein Degradation as an Antiviral Approach. Antivir. Res. 2023, 210, 105480. [Google Scholar] [CrossRef] [PubMed]
- Chitalia, V.C.; Munawar, A.H. A Painful Lesson from the COVID-19 Pandemic: The Need for Broad-Spectrum, Host-Directed Antivirals. J. Transl. Med. 2020, 18, 390. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug/Compound | Target/Mechanism | Drug Class | Mode of Action | Clinical Use/Efficacy | Resistance/Limitations |
|---|---|---|---|---|---|
| Remdesivir | RdRp (nsp12) | Nucleoside analogue | Adenosine analogue; chain termination via delayed RNA translocation | IV use; moderately accelerates recovery in hospitalized patients | Partial resistance via nsp12 mutations (e.g., V166, E802); fitness cost limits spread |
| Molnupiravir | RdRp | Nucleoside analogue | Cytidine/uridine analogue; induces lethal mutagenesis | Oral use; reduces hospitalization in mild-to-moderate cases | Theoretical mutagenic risks; not fully neutralized by proofreading |
| Favipiravir | RdRp | Nucleoside analogue | Purine analogue; weak RNA synthesis inhibition | Limited effect; not a frontline drug | Requires high concentrations; inconsistent trial results |
| Ribavirin | RdRp | Nucleoside analogue | Guanosine analogue; promotes mutagenesis | Alone: limited effect; synergistic in combinations | High toxicity; synergizes with remdesivir |
| VV116 (Deuviridine) | RdRp | Nucleoside analogue | Oral prodrug of GS-441524 | Comparable to Paxlovid in trials; high bioavailability | Not widely approved |
| Nirmatrelvir + Ritonavir (Paxlovid) | 3CLpro | Protease inhibitor | Blocks polyprotein cleavage; halts replication complex maturation | Oral use; ~89% reduction in hospitalization if early | Resistance via 3CLpro mutations (in vitro); clinical relevance not yet clear |
| Ensitrelvir | 3CLpro | Protease inhibitor | Similar to nirmatrelvir | Effective in Japan trials | Investigational |
| Lopinavir + Ritonavir | 3CLpro | Protease inhibitor | HIV protease inhibitor; weak SARS-CoV-2 activity | No benefit in early COVID-19 trials | Largely abandoned |
| GRL-0617 | PLpro | Protease inhibitor | Preclinical inhibitor of papain-like protease | Preclinical stage | Not clinically available |
| Fluoxetine | Host (cell pathways) | SSRI/host-directed | Synergistic with GS-441524; may block viral egress | In vitro synergy with remdesivir | Lysosomotropic mechanism; repurposed drug |
| Itraconazole | Host (cholesterol trafficking) | Antifungal/host-directed | Disrupts sterol trafficking needed for replication | Enhances remdesivir efficacy in vitro | Repurposed, not virus-specific |
| Baricitinib | Host (JAK/STAT + endocytosis) | JAK inhibitor/immunomodulator | Blocks cytokine storm; may block viral entry | EUA with remdesivir; reduces inflammation | Minor antiviral activity; adjunctive |
| Camostat | TMPRSS2 | Host entry inhibitor | Blocks spike protein priming | In vitro efficacy; used in combinations | Investigational |
| Brequinar | DHODH (pyrimidine synthesis) | Host-targeting | Reduces the nucleotide pools needed for RNA synthesis | Strong synergy with molnupiravir | Host toxicity concerns |
| Remdesivir + Nirmatrelvir | RdRp + 3CLpro | Antiviral synergy | Blocks RNA synthesis and protein processing | Superior in vitro and in case reports | Dual targeting reduces the resistance potential |
| Remdesivir + Ribavirin | RdRp | Chain terminator + mutagen | Complete viral extinction in vitro | Enhances the mutational burden and replication block | Not in clinical use |
| Molnupiravir + Nirmatrelvir | RdRp + 3CLpro | Dual-action antiviral | Polymerase + protease inhibition | Enhanced synergy; potential for resistant strains | Preclinical and early trial stages |
| Molnupiravir + Camostat + Brequinar | Multi-target | Polymerase + entry + nucleotide synthesis | Maximal suppression in vitro | For persistent/resistant infections | Not tested clinically |
| Strategy | Targets/Approaches | Key Compounds/Tools | Potential Impact |
|---|---|---|---|
| Novel Viral Targets | nsp14 exonuclease (ExoN), the NiRAN domain of nsp12, PLpro, and macrodomains | PLpro inhibitors (e.g., GRL-0617), and experimental NiRAN inhibitors | Enhances polymerase inhibitor efficacy; reduces immune suppression; introduces new viral drug targets |
| Host-Directed Antivirals | TMPRSS2, pyrimidine biosynthesis (e.g., DHODH), TRiC, Hsp90, JAK-STAT pathways, IFN-λ system | Camostat, nafamostat, brequinar, baricitinib, teriflunomide, interferon lambda (IFN-λ) | Reduced viral resistance potential; broad-spectrum activity; may pose host toxicity risks |
| AI and Computational Drug Discovery | Structure-based virtual screening, machine learning prediction, synergy modeling, allosteric pockets on RdRp (nsp12) | COVID Moonshot, flavonoid derivatives (e.g., myricetin), ML-based compound ranking | Accelerates drug discovery; identifies novel scaffolds; enables variant-specific antiviral tailoring |
| Gene silencing and RNA-Targeting Therapies | siRNA targeting conserved genome regions (e.g., RdRp); CRISPR-Cas13-mediated cleavage of viral RNA | siRNA-nanoparticle delivery systems and Cas13-based antiviral platforms | Programable and rapidly deployable antivirals; adaptable to emerging viruses |
| Pan-Coronavirus and Broad-Spectrum Antivirals | Conserved viral domains (nsp12-nsp7/nsp8 interface, nsp13 helicase); shared host dependency factors (e.g., DHODH, IFN) | Ribavirin, NHC, DHODH inhibitors, innate immune modulators | Enables pandemic preparedness; broad viral coverage; strategic stockpiling for future zoonotic outbreaks |
| Strategy | Type | Evidence of Activity | Preclinical/Clinical Status |
|---|---|---|---|
| GRL-0617 | PLpro (nsp3) inhibitor | Inhibits viral polyprotein cleavage in vitro | Preclinical |
| VV116 (Deuviridine) | Nucleoside analogue | Comparable to Paxlovid in early clinical trials | Clinical (Phase 3, China) |
| Camostat | Host-directed antiviral | Blocks viral entry; enhances effects in combinations | Clinical (investigational) |
| Brequinar | Host-directed antiviral | Synergistic with molnupiravir in lung cell models | Preclinical/Early trials |
| Baricitinib | Host-directed antiviral/immunomodulator | EUA with remdesivir; reduces inflammation & viral entry | Approved (with remdesivir) |
| Molnupiravir | Nucleoside analogue | Reduces hospitalization in mild/moderate cases | Approved (oral) |
| Remdesivir | Nucleoside analogue | Accelerates recovery in hospitalized patients | Approved (intravenous) |
| Fluoxetine | Host-directed/repurposed | Enhances remdesivir efficacy in vitro | Preclinical |
| Cas13-based platforms | Gene-targeting modality | Inhibits viral RNA replication in vitro | Preclinical (proof-of-concept) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subong, B.J.J.; Forteza, I.L. SARS-CoV-2 Replication Revisited: Molecular Insights and Current and Emerging Antiviral Strategies. COVID 2025, 5, 85. https://doi.org/10.3390/covid5060085
Subong BJJ, Forteza IL. SARS-CoV-2 Replication Revisited: Molecular Insights and Current and Emerging Antiviral Strategies. COVID. 2025; 5(6):85. https://doi.org/10.3390/covid5060085
Chicago/Turabian StyleSubong, Bryan John J., and Imelda L. Forteza. 2025. "SARS-CoV-2 Replication Revisited: Molecular Insights and Current and Emerging Antiviral Strategies" COVID 5, no. 6: 85. https://doi.org/10.3390/covid5060085
APA StyleSubong, B. J. J., & Forteza, I. L. (2025). SARS-CoV-2 Replication Revisited: Molecular Insights and Current and Emerging Antiviral Strategies. COVID, 5(6), 85. https://doi.org/10.3390/covid5060085